
Abstract
Background
The aim of this study was to analyze the response of selected components of the immune system in rowers to maximal physical exercise, and to verify if this response can be modulated by supplementation with spirulina (cyanobacterium Spirulina platensis).
Method
The double-blind study included 19 members of the Polish Rowing Team. The subjects were randomly assigned to the supplemented group (n = 10), receiving 1500 mg of spirulina extract for 6 weeks, or to the placebo group (n = 9). The participants performed a 2000-m test on a rowing ergometer at the beginning (1st examination) and at the end of the supplementation period (2nd examination). Blood samples were obtained from the antecubital vein prior to each exercise test, 1 min after completing the test, and after a 24-h recovery period. Subpopulations of T regulatory lymphocytes (Tregs) [CD4+/CD25+/CD127-], cytotoxic lymphocytes (CTLs) [CD8+/TCRαβ+], natural killer (NK) cells [CD3-/CD16+/CD56+] and TCRδγ-positive (Tδγ) cells were determined by means of flow cytometry.
Results
On the 2nd examination, athletes from the supplemented group showed neither a post-exercise increase in Treg count nor a post-recovery decrease in Tδγ cell count (both observed in the placebo group), and presented with significantly lower values of Treg/CTL prior to and after the exercise. During the same examination, rowers from the placebo group showed a significant post-recovery increase in Treg/(NK + Tδγ + CTL) ratio, which was absent in the supplemented group.
Conclusion
The results of this study imply that supplementation with spirulina extract may protect athletes against a deficit in immune function (especially, anti-infectious function) associated with strenuous exercise, and may cause a beneficial shift in “overtraining threshold” preventing a radical deterioration of immunity.
Background
Previous studies demonstrated that influence of physical exercise on the immune system may vary considerably depending on the type of training load. Regular, moderate physical exercise improves immunity, exerting a beneficial effect on the immune system [Citation1]. However, an opposite effect may be exerted by strenuous exercise which was shown to activate immunological and endocrine mechanisms similar to those prevailing in chronic stress, trauma and sepsis [Citation2, Citation3]. This may lead, in particular in high-performance athletes, to impairment of immune response and resultant greater susceptibility to infections, e.g. respiratory tract infections [Citation4, Citation5]. Previous studies demonstrated that a decrease in salivary IgA concentrations and enhanced synthesis of antigen-stimulated IL-10 correlate strongly with an increase in the incidence of upper respiratory tract infections in athletes. However, the exact etiological mechanism of the immunity impairment is still not fully understood [Citation6, Citation7]. Moreover, no other markers of the exercise-induced immunity impairment predisposing to upper respiratory tract infections have been established thus far [Citation7].
Krüger et al. [Citation8] and Adams et al. [Citation9] showed that strenuous exercise leads to a transient increase in circulating lymphocyte count, followed by a persistent decrease in this parameter, even to the point of lymphopenia. According to these authors, after strenuous exercise, lymphocytes may migrate to some critical regions, including muscles, particularly resulting in a shift in lymphocyte subpopulations [Citation10]. Effector lymphocytes, such as natural killer (NK) cells, cytotoxic T CD8+ cells (CTLs), TCRδγ-positive (Tδγ) cells and regulatory T cells (Tregs), are a key component of human immune system and play a crucial role in cell-mediated immune response. Interactions between cytotoxic lymphocytes and Tregs are of particular interest to researchers dealing with various pathological conditions (e.g. autoimmune diseases and neoplasms) [Citation11–Citation19]. In contrast, only few previous studies have addressed the problem associated with a shift in lymphocyte subpopulations and interactions between various types of immune cells in athletes. Furthermore, the results of these sparse studies are generally inconclusive.
Many recent studies dealt with Tregs, a lymphocyte subpopulation that can be identified on flow cytometry based on the expression of CD4, CD25 and FOXP3 antigens (or lack of CD 127) [Citation20]. This growing research interest is associated with the leading role of Tregs in immune tolerance, as well as with their involvement in the control of an overactivated immune system. Under physiological conditions, Tregs represent no more than 5–10% of CD4+ lymphocytes present in peripheral blood [Citation20, Citation21] . Tregs prevent autoimmune reactions, allograft rejection, food intolerance and rejection of the fetus by the maternal immune system [Citation20]. However, aside from these beneficial effects, they may also interfere with effector function and proliferation of cells involved in the specific and non-specific immune response, such as CTLs, NK cells, monocytes, macrophages and dendritic cells [Citation22, Citation23]. Available evidence suggests that overactivity of Tregs may result in immune impairment, potentially predisposing individuals to chronic infections or facilitate formation of metastases in cancer patients [Citation13, Citation15, Citation18, Citation19, Citation24, Citation25]. Kunzmann et al. [Citation26] showed that Tregs can inhibit phosphoantigen-induced proliferation of non-conventional highly-cytotoxic Tδγ cells. Furthermore, few previous studies demonstrated that repeated strenuous exercise may persistently alter absolute and relative size of lymphocyte subpopulations, causing a shift toward Tregs. This phenomenon may be inter alia reflected by an increase in the level of IL-10 synthesized by these cells, and is at least in part responsible for the resulting immunosuppression [Citation27]. Furthermore, strenuous physical exercise may cause a decrease in the number of NK cells and CTLs [Citation10, Citation28, Citation29].
However, the results of previous studies analyzing immunomodulatory effects of physical exercise, either in humans or in animal models, are inconclusive. Wang et al. [Citation30] demonstrated that 6 weeks of exhaustive high-intensity exercise resulted in an increase in the number of Tregs, and altered proportions of CD4+ and CD25+ cells in murine spleen. Similar effects, however, were not observed in mice subjected to moderate-intensity exercise. These findings were partially confirmed by Wilson et al. [Citation31] who showed that acute, intense swimming exercise contributed to an increase in the number of circulating Tregs in adolescent athletes. However, quite contradictive results were obtained by Perry et al. [Citation32] when analyzing the effects of strenuous endurance exercise in marathon runners and half-ironman triathletes, reporting a substantial post-exercise decrease in the number of circulating Tregs.
Addition of supplements with established immunomodulatory properties to athletes’ diet may constitute a form of mild and safe intervention to restore immune balance. A cyanobacterium, Spirulina platensis (SPR) is a supplement with a good safety profile and established immunomodulatory potential [Citation33, Citation34]. Many previous studies document beneficial effects of SPR on the immune system in both animals and humans [Citation34, Citation35]. SPR is rich in protein, vitamins, minerals and unsaturated fatty acids, especially gamma-linolenic acid. Moreover, it contains phycocyanins and lipopolysaccharides, key components of cell walls in Gram-negative bacteria [Citation36]. Many authors showed that SPR may stimulate macrophages to synthesize IL-12, a cytokine that promotes production of INT-γ by NK cells [Citation37]. Published evidence suggests that supplementation with SPR may also prevent replication of viruses, and thus may be helpful in control of many viral infections, such as chickenpox, cytomegaly, measles, mumps, influenza and HIV [Citation38, Citation39]. Løbner et al. [Citation40] showed that supplementation with a polysaccharide extract of SPR stimulated production of IL-2, INT-γ and TNF-alpha, while down-regulating the synthesis of IL-4 in antigen-activated lymphocytes from healthy volunteers. A decrease in IL-4 production was also observed in SPR-supplemented (2 g daily for 12 weeks) patients with allergic rhinitis. Since IL-4 was previously shown to inhibit spontaneous apoptosis of Tregs, its down-regulation likely enhances programmed death of these cells. Theoretically, the resultant decrease in Treg count should promote a cell-mediated immune response [Citation41, Citation42]. Indeed, a 7-day supplementation with SPR extract increased NK cell activity in healthy persons [Citation43]. Moreover, administration of SPR enhanced antitumor activity in syngeneic tumor-implanted mice [Citation35].
Many previous studies demonstrated that SPR may cause a shift in Th1/Th2 balance toward Th1-dominant immunity. This justifies the analysis of SPR’s effects in athletes, in whom strenuous physical exercise may impair immunity and disrupt balance between cytotoxic lymphocytes and Tregs. However, to the best of our knowledge, none of the previous studies analyzed the effects of SPR on the number of Tregs, NK cells, CTLs and Tδγ cells in this group. Nevertheless, the studies analyzing suppressive effects of Tregs and cytotoxicity of NK cells, Tδγ cells and CTLs may produce flawed results unless other components of the immune system are considered. Exposure to an extreme stressor, such as maximal exercise, results in an increase/decrease in the number of cytotoxic lymphocytes, counterbalanced with a decrease/increase in Treg count. It is the outcome of these processes, which eventually reflects the influence of exercise on immune function. Since previous studies documented a plethora of interactions (either direct or cytokine-mediated) between various lymphocyte subpopulations, we searched for possibly most accurate marker of these complex relationships. Other authors [Citation12, Citation17, Citation26, Citation44] demonstrated that Treg to effector T cell ratio reflects suppressive effects of the former on various effector cells more accurately than a simple determination of their counts. To the best of our knowledge, none of the previous studies examined exercise- induced changes in these parameters in sportspersons. Therefore, the aim of this study was to analyze changes in Treg/Tδγ, Treg/NK, Treg/CTL and Treg/(NK + Tδγ + CTL) ratios in SPR-supplemented athletes subjected to strenuous exercise; to the best of our knowledge, none of the previous studies examined exercise-induced changes in these parameters in sportspersons.
Methods
Study population
The study included 19 men, all members of the Polish Rowing Team (15 heavy-weight and 4 light-weight rowers). Basic characteristics and sport classes of the athletes are presented in Table . The study was conducted between March and May, during a 6-week training camp, scheduled between the preparatory and competitive phase of the yearly training cycle. The characteristics of the training profile, such as its intensity, volume (in minutes) and type (specific, i.e. rowing: endurance, technical, speed, etc., and nonspecific: jogging, strength) were recorded on a daily basis. The intensity of the training was classified based on the lactate acid (LA) threshold (4 mmol/L), as extensive (below the LA threshold), highly intensive (above the LA threshold), and extremely intensive (control tests) (Table ).
Table 1 Basic characteristics of the study groups
Table 2 Training schedule for the week preceding blood sampling during the 1st and the 2nd examination
Food intake
Throughout the study period, the athletes accommodated at one of the Olympic Training Centers, whereby they had all their meals. Their regular menu consisted of a mixed diet, providing the recommended dietary allowance (RDA) of carbohydrates, proteins, fats and micronutrients (vitamins and minerals), in line with the Polish Nutrition Society guidelines [Citation45]. Daily intakes of food, calories, fruits and vegetables were the same throughout the study period. All athletes were provided with a bottled mineral water and instructed about the necessity of immediate fluid repletion, in particular prior to and after the exercise.
The study subjects declared that they had ceased all drugs, medications and dietary supplements at least 2 weeks prior to the study, and did not use them throughout the entire study period.
Experimental procedure
Athletes who were randomized to the supplemented group (n = 10) received capsules with Spirulina platensis extract, manufactured by GAL (Poznan, Poland). A single 596-mg capsule was made of 500 mg Spirulina platensis containing 5.9 mg chlorophyll, 0.093 mg vitamin B6, 6.5 μg vitamin K, 0.8 μg vitamin B12, 3.5 μg selenium and 9.0 μg iodine, coated with 96 mg gelatin. The subjects were asked to take one capsule before each of their three main meals during the day, for a period of 6 weeks, which corresponded to 1500 mg of spirulina extract per day. Athletes randomized to the placebo group (n = 9) received visually identical capsules with calcium gluconate (500 mg per capsule).
Training program
Training volumes (expressed in minutes per day) during a week preceding the 1st and the 2nd examination, particularly for extensive rowing, intensive rowing, kilometers, and extensive non-specific training, are shown in Table . During the load training phase (before the 1st examination), the training volume amounted to 1130 min· wk.− 1, including approximately 49.6% of extensive rowing, 26.5% of non-specific training (e.g. power training) and 23.9% of intensive rowing. Total training volume before the 2nd examination was 940 min·wk.− 1, and included approximately 66.9% of extensive rowing, 10.7% of intensive rowing (with 5.2% of maximum-intensity control tests) and 11.7% of land training.
Rowing performance test
The athletes performed a controlled 2000-m time trial on the first day (prior to the supplementation) and at the end of training camp (after the supplementation). Each subject had to cover the 2000-m distance on a rowing ergometer (Concept II, Morrisville, VT USA) in as short time as possible. Because the results of both tests were taken into consideration during selection to the championship team, the athletes were well motivated to perform both tests at maximal effort. Prior to each test, the subjects performed a 5-min individual warm-up on the rowing ergometer Concept II, according to their own freely selected program.
Sample treatment
Blood samples from the antecubital vein were collected to tubes with dipotassium ethylene diamine tetra-acetic acid (K2EDTA) as an anticoagulant. Blood was collected before each 2000-m test (after 7–8 h of overnight fasting), 1 min after completing the test, and after a 24-h recovery period. The samples were centrifuged at 2200 rpm for 10 min. After removing the plasma and adding 1× Lysing Buffer (BD Biosciences), the samples were incubated in darkness for 15 min. Then, a PBS buffer was added, and the cells were washed twice to remove all erythrocytes.
Moreover, capillary blood samples from an ear lobe were collected prior to and after each exercise test, to assess lactic acid (LA) level.
Measurements
Cytometric analysis of lymphocyte subpopulations: Tregs [CD4+/CD25+/CD127-], CTLs [CD8+/TCRαβ+], NK cells [CD3-/CD16+/CD56+] Tδγ cells was conducted after their labeling with fluorochrome-conjugated antibodies from BD Biosciences. Cells obtained after hypotonic lysis of peripheral blood were incubated in darkness at room temperature for 20 min with the respective antibody at a concentration specified by the manufacturer to identify each lymphocyte subpopulation (Table ). Then, the cells were washed twice with PBS buffer and left in darkness in 3.7% formaldehyde solution for 10 min. Afterwards, the cells were again washed with PBP buffer and 100 μL of DAPI solution (1 mg/mL, Thermo Fisher Scientific) were added to stain cell nuclei. The cells were incubated in darkness at room temperature for 5 min, washed twice, and suspended in 250 μL of PBS buffer. After labeling, the cells were analyzed with a LSRII flow cytometer from BD Biosciences, coupled with BD FACSDiva software.
Table 3 Antibodies used for identification of lymphocyte subpopulations
Statistical analysis
Statistical analyses were performed with STATISTICA v. 10.0 software package (StatSoft, Cracow, Poland). All parameters were compared using 2 (supplementation: supplemented vs. placebo) × 3 (exercise: pre-exercise vs. post-exercise vs. post-recovery) repeated measures analysis of variance (ANOVA). Normal distribution of the study variables was verified with Shapiro-Wilk test. Whenever the result of ANOVA was statistically significant, Fisher’s post-hoc test was conducted to identify the source of significant differences. Anthropometric characteristics of the study groups were compared with Student’s unpaired t-test. Except from the rowing time, the results of the 2000-m tests performed prior to and after the supplementation were compared with Student’s paired t-test, and intergroup comparisons were conducted with Student’s unpaired t-test. The results of the 2000-m simulated rowing test were subjected to one-way ANOVA. Statistical characteristics of the study variables are presented as means ± SD, and the threshold of statistical significance for all tests was set at P < 0.05.
Results
Athletes from the supplemented group and the placebo group did not differ significantly in terms of their mean age, height, body weight, and years of training (Table ).
No significant intergroup differences were found in mean power output and total run time during the 2000-m test performed at the beginning of the training camp. Furthermore, no significant differences in the pre- and post-test blood LA levels were documented when the results of the 1st examination were compared to those of the 2nd examination (Table ).
Table 4 Changes in 2,000 m rowing ergometer performance before and after supplementation
Treg counts determined during the 1st and the 2nd examination are shown on Fig. . ANOVA revealed a significant main effect of exercise on Treg count (p < 0.001). During the 2nd examination, athletes from the placebo group showed a significant post-exercise increase in Treg count, with subsequent normalization of this parameter post-recovery (p < 0.05). In contrast, no significant exercise-induced changes in Treg count were observed in the supplemented group during the 2nd examination, as well as in both study groups during the 1st examination. Based on the results of ANOVA, spirulina supplementation did not exert a statistically significant effect on Treg count (main effect, p = 0.069).
Fig. 1 Changes in Treg (a), NK cell (b), Tδγ cell (c) and CTL (d) counts during exercise tests performed prior to and after the supplementation (mean ± SD). Note. Tregs = regulatory T cells; NK = natural killer cells; Tδγ = gamma delta T cells; CLT = cytotoxic T lymphocytes; gray box - SUPPL = supplemented group; □ - PLA = placebo group; B = baseline; Ex = post-exercise; R = after a 1-day recovery; # - significantly different compared to the baseline level; * - significantly different compared to the post-exercise level
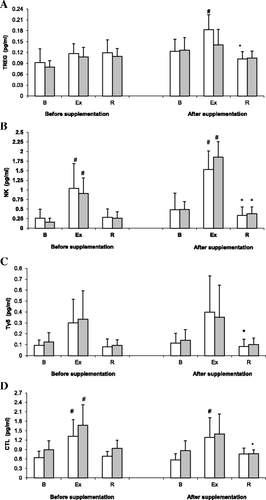
NK cell counts determined prior to and after supplementation with SPR extract are shown in Fig. . ANOVA demonstrated a significant main effect of exercise on NK cell count (p < 0.001). During both the 1st and the 2nd examination, a significant post-exercise increase in this parameter was observed in both the supplemented group and the placebo group, followed by a post-recovery normalization (solely during the 2nd examination).
Tδγ cell counts in the study subjects are presented on Fig. . Like for other previously mentioned lymphocyte subpopulations, ANOVA demonstrated a significant main effect of exercise on Tδγ cell count (p < 0.001). During the 1st examination, pre-exercise, post-exercise and post-recovery Tδγ cell counts did not differ significantly, in the supplemented group or placebo group. However, during the 2nd examination, a significant post-exercise decrease in Tδγ cell count was observed in the placebo group. Nevertheless, the main effect of supplementation on Tδγ cell count was not statistically significant (p = 0.691).
ANOVA demonstrated that exercise exerted a significant main effect on CTL count (p < 0.001). Irrespective of the group, a significant post-exercise increase in CTL count was observed during the 1st examination (Fig. ). However, during the 2nd examination, the significant post-exercise increase in this parameter was observed in the placebo group, but not in the supplemented group. Furthermore, athletes from the supplemented group showed a post-recovery decrease in CTL count during the 2nd examination. However, the main effect of supplementation on CTL count did not turn out to be statistically significant on ANOVA (p = 0.333).
Values of the Treg/(NK + Tδγ + CTL) ratio are presented on Fig. . During the 1st examination, a significant post-recovery increase in Treg/(NK+ Tδγ + CTL) ratio was observed in both study groups. In turn, during the 2nd examination, post-exercise values of Treg/(NK+ Tδγ + CTL) ratio in the supplemented group and the placebo group turned out to be significantly lower than pre-exercise values of this parameter. Nevertheless, ANOVA showed that the main effect of supplementation on Treg/(NK+ Tδγ + CTL) ratio was not statistically significant (p = 0.563).
Fig. 2 Treg/(NK + Tδγ + CTL) ratio (a), Treg/NK ratio (b), Treg/Tδγ ratio (c) and Treg/CTL (d) ratio during exercise tests performed prior to and after the supplementation (mean ± SD). Note. Tregs = regulatory T cells; NK = natural killer cells; Tδγ = gamma delta T cells; CTL = cytotoxic T lymphocytes; gray box - SUPPL = supplemented group; □ − PLA = placebo group; B = baseline; Ex = post-exercise; R = after a 1-day recovery; # - significantly different compared to baseline level; * - significantly different compared to post-exercise level; † − significantly different compared to PLA group

Values of Treg/NK ratio were modulated solely by exercise. Specifically, the post-exercise values of this parameter in both study groups were significantly lower than pre-exercise levels (Fig. ). However, this relationship was observed solely during the 1st examination, as no significant exercise-induced changes in Treg/NK ratio were found during the 2nd examination. Supplementation exerted no significant effect on the values of Treg/NK ratio.
Exercise-induced changes in Treg/Tδγ ratio are presented in Fig. . Irrespective of the study group, strenuous physical exercise contributed to statistically significant changes in this parameter during both the 1st and the 2nd examination (main effect, p < 0.001). The post-recovery increase in Treg/Tδγ ratio in the supplemented and control group corresponded to 154% and 274%, respectively, prior to the supplementation, and to 133% and 101%, respectively, after the supplementation. Based on the results of ANOVA, spirulina supplementation did not exert a statistically significant effect on Treg/Tδγ ratio (main effect, p = 0.129).
Statistical analysis showed that both exercise and supplementation exerted a significant main effect on the values of Treg/CTL ratio (p = 0.01 and p < 0.001, respectively) (Fig. ). Irrespective of the group, no statistically significant changes in this parameter were documented during the 1st examination. However, during the 2nd examination, a significant post-recovery decrease in this parameter was observed in the supplemented group, but not in the placebo group. Furthermore, post-exercise and post-recovery values of Treg/CTL ratio in supplemented athletes turned out to be significantly lower than in subjects from the placebo group.
Discussion
During the 2nd examination, a significant post-exercise increase in Treg count was observed solely in the placebo group, with subsequent normalization of this parameter after a 24-h recovery (Fig. ). Since lymphocytes are known to constantly migrate between the blood and other tissues, the post-exercise increase in circulating Tregs observed in the placebo group likely reflected a concomitant increase in the tissue count of these cells (Table ). Previous research showed that Tregs may leave the circulation and migrate to lymph nodes and inflamed tissues [Citation46, Citation47], whereby they mitigate the activity of cytotoxic cells and antigen presentation by dendritic cells (DCs) [Citation48]. This local suppressive effect of Tregs may exert an unfavorable effect on systemic immunity. Morgado et al. [Citation49] demonstrated that an increase in the training loads of swimmers contributed to a significant reduction of cytokine synthesis by monocytes and dendritic cells. Immune impairment associated with heavy training loads may occur primarily in immunologically active tissues, at least partially explaining the difficulties in identification of accurate markers of immunosuppression that could be determined in peripheral blood. Published evidence suggests that the immune impairment caused by escalation of training loads has no specific laboratory profile [Citation7, Citation50, Citation51]. It should be emphasized that circulating lymphocytes represent only 2% of lymphocyte population; the vast majority of lymphocytes can be found in immunologically active tissues, such as lymph nodes, spleen, intestines, blood marrow, thymus and skin [Citation52]. During heavy training, some tissue lymphocytes may migrate to the circulation in response to hormonal stimulation typical for strenuous exercise (e.g. catecholamines, cortisol). During post-exercise recovery, these lymphocytes migrate back to tissues, especially to those generating strong inflammatory/chemotactic signals [Citation53]. This hypothesis seems to be supported by the post-exercise increase in Treg count and subsequent post-recovery normalization of this parameter, observed during the 2nd examination in the placebo group (Fig. ).
ANOVA did not demonstrate a significant main effect of supplementation on Treg count; nevertheless, athletes from the supplemented group did not show a significant post-exercise increase in this parameter during the 2nd examination (Fig. ). Therefore, it can be assumed that supplementation with SPR played a role in the maintenance of lower Treg counts in tissues, preventing immunosuppressive effect of these cells and restoring an immune balance.
During the 2nd examination, athletes from the supplemented group presented with significantly lower pre-exercise and post-exercise values of Treg/CTL ratio compared to subjects from the placebo group (Fig. ). Lower values of Treg/CTL ratio in the supplemented group might also reflect a beneficial effect of SPR supplementation. The decrease in this parameter implies that SPR might mitigate the inhibitory effect of Tregs on CTLs. Due to lesser immune deficit, athletes from the supplemented group might have been better protected against opportunistic infections and reactivation of latent viral infections (e.g., with CMV, EBV and HSV-1). In turn, higher values of Treg/CTL ratio in the placebo group might reflect an unfavorable shift in the “overtraining threshold” associated with a radical deterioration of immunity.
Irrespective of the examination term and the study group, we observed a significant post-recovery increase in Treg/Tδγ ratio (Fig. ). This effect was modulated solely by exercise. Nevertheless, during the 2nd examination we observed a significant post-recovery decrease in Tδγ cell count in the placebo group, but not in the supplemented group (Fig. ). Due to presumable protective effect of SPR, preventing the post-recovery decrease in Tδγ cell count, strenuous exercise had probably less detrimental effect on the immune function in supplemented athletes. Tδγ cells play a key role in antibacterial, antiviral and antitumor immunity. Published evidence suggests that the number of Tδγ cells is inter alia modulated by exercise intensity. Anane et al. [Citation1] demonstrated that either high- or low-intensity exercise stimulated an increase in the number of circulating Tδγ cells in previously untrained persons. However, another study showed that escalation of training loads during a winter training season resulted in a decrease in Tδγ cell counts in peripheral blood of elite swimmers [Citation54].
Published data imply that SPR may exert a substantial effect on the activation of NK cells and their cytotoxic potential [Citation35, Citation37, Citation43]. We did not observe a significant main effect of supplementation on either NK cell count (Fig. ) or Treg/NK ratio (Fig. ); however, irrespective of the examination term, strenuous exercise stimulated a significant increase in the number of NK cells (Fig. ).
According to a widely accepted concept, strenuous exercise stimulates migration of tissue lymphocytes to peripheral blood in a catecholamine-dependent mechanism. This hypothesis is inter alia supported by an increase in circulating lymphocyte count observed after infusion of epinephrine [Citation55, Citation56]. However, the fact that the post-exercise increase in lymphocyte count in elite athletes was less evident than in untrained persons, implies that immune response of the former group to strenuous exercise may be weaker [Citation57, Citation58].
Post-exercise changes in lymphocyte count in intensively trained athletes were markedly less evident than in untrained persons, and less pronounced than it could be expected based on epinephrine and cortisol concentrations. However, published evidence suggests that changes in hormonal parameters do not necessarily correlate with lesser mobilization of tissue lymphocytes and their migration to peripheral blood [Citation8, Citation10]. Therefore, lesser responsiveness of tissue lymphocytes from elite athletes to strenuous exercise was interpreted as a consequence of training-induced changes, such as a decrease in the reactivity of ß2-adrenergic receptors on these cells, altered expression of adhesive molecules or their ligands [Citation57, Citation58].
However, in our opinion, also other potential mechanisms contributing to reduced mobilization of tissue lymphocytes after strenuous exercise need to be considered. According to a widely accepted and empirically verified hypothesis, various stimuli (among them physical exercise and infections) may induce bidirectional flow of lymphocytes from/to tissues (spleen, muscles, lungs, bone marrow) [Citation9]. Adams et al. [Citation9] showed that exercise promotes rapid migration of tissue lymphocytes (primarily from the spleen and lungs) to the circulation, along with markedly less evident shift of these cells from muscles to blood. Although this observation originates from a study in rats, the post-exercise increase in circulating lymphocyte counts in humans may also, to a certain degree, reflect higher absolute number of these cells in the spleen and lungs (and to a lesser extent, in muscles). Less evident post-exercise migration of lymphocytes to the peripheral blood of well-trained athletes may be associated with lower number of cytotoxic cells in their spleen and lungs, and/or with higher number of tissue Tregs. The latter hypothesis is also supported by the results of our study, which demonstrated a significant post-exercise increase in Treg count in the placebo group during the 2nd examination. However, also lymphopenia, observed few hours post-exercise as a consequence of the recirculation of blood lymphocytes to tissues (only < 10% of circulating lymphocytes undergo apoptosis), is a well-established phenomenon [Citation10]. During post-exercise recovery of well-trained athletes, their lymphocytes may primarily recirculate to the muscles; due to multiple training-induced microinjuries, muscle tissue synthesizes large volumes of cytokines which act as a chemoattractant for peripheral lymphocytes [Citation59, Citation60].
According to Adams et al. [Citation9], recirculation of lymphocytes from muscles to the blood is less pronounced and slower than between spleen or lungs and peripheral circulation. Probably, lymphocytes of well-trained athletes may remain anchored in muscles for a longer time, which makes them less prone to recirculation and more susceptible to apoptosis. This may result in permanent loss of some cytotoxic lymphocytes from the spleen and lungs. However, the exact reason behind a post-exercise increase in Treg count in well-trained athletes is still unclear. Perhaps, this phenomenon is associated with an increase in IL-2 (synthesized by T cells in response to repeated strenuous exercise), stabilization of Foxp3, longer survival of mature Tregs, and resultant increase in their number in some tissues [Citation61, Citation62].
Our findings suggest that another, yet unidentified mechanism may exist behind the post-exercise immune impairment observed in elite athletes subjected to heavy training loads. Higher number of circulating Tregs may reflect their increased counts in immunologically active organs, presumably in the spleen and lungs. The number of these tissue Tregs that migrate to the circulation during strenuous exercise may be higher than the number of migratory cytotoxic T lymphocytes (other than NK cells). Furthermore, it cannot be excluded that immunologically active organs are not only abundant in Tregs, but also contain less cytotoxic lymphocytes (Tδγ, CTL). During strenuous exercise, these sparse cytotoxic cells may migrate to other tissues (primarily muscles) whereby they undergo apoptosis, which probably also contributes to a post-exercise immunity impairment.
Our present study showed, for the first time, that supplementation with SPR may modulate some components of the immune system in athletes exposed to repeated strenuous exercise. The fact that rowers from the supplemented group did not show a post-exercise increase in Treg count (Fig. ) implies that SPR may play a role in maintaining normal tissue level of these cells during strenuous exercise, thus preventing immunosuppression. Moreover, SPR seemed to attenuate a suppressive effect of Tregs on CTLs, since during the 2nd examination, athletes from the supplemented group presented with significantly lower pre-exercise and post-exercise values of Treg/CTL ratio than subjects from the placebo group. Finally, strenuous physical exercise did not exert a significant effect on Tδγ cell count in the supplemented group, whereas athletes from the placebo group showed a post-recovery decrease in this parameter. Altogether, these findings suggest that supplementation with SPR may exert a beneficial effect on selected components of the immune system in athletes exposed to heavy training loads.
Future studies should center around better understanding of the mechanisms of immune impairment activated during and after strenuous exercise. Another direction of future research should be the identification of factors that may counterbalance the unfavorable consequences of exposure to maximal training loads.
Conclusions
The results of this study imply that supplementation with SPR extract might prevent a post-exercise increase in Treg counts and a post-recovery decrease in the number of Tδγ cells, which contributed to lower Treg/CTL pre-exercise and post-exercise values ratio during the 2nd examination. Altogether, these findings suggest that supplementation with SPR extract may protect athletes against a deficit in immune function (especially, anti-infectious function) associated with strenuous exercise, and may cause a beneficial shift in an “overtraining threshold” that prevents a radical deterioration of immunity.
Authors’ contributions
The present study was designed by ASS and AJ; data were collected by PB, JT, KŁ; data interpretation and manuscript preparation were undertaken by AJ, ASS, EP and BM. All authors approved the final version of the paper.
Ethics approval and consent to participate
The experimental procedures and potential risks were explained to the participants and a written informed consent was provided and signed prior to inclusion in the study. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of the Poznan University of Medical Sciences (act 769/13 archived number 10/October/2013).
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest, financial or otherwise. The authors declare that the results of the study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
| CD | = | Cluster of differentiation |
| CMV | = | Cytomegalovirus |
| CTL | = | Cytotoxic T lymphocyte |
| DCs | = | Dendritic cells |
| EBV | = | Epstein-Barr virus |
| FOXP3 | = | Forkhead box P3 |
| HIV | = | Human immunodeficiency virus |
| HSV | = | Herpes simplex viruses |
| IL | = | Interleukin |
| INT-γ | = | Interferon gamma |
| LA | = | Lactic acid |
| NK | = | Natural killer |
| RDA | = | Recommended dietary allowance |
| SPR | = | Spirulina platensis |
| TNF alpha | = | Tumor necrosis factor alpha |
| Treg | = | Regulatory T cells |
| Tδγ | = | T cells gamma delta |
Acknowledgments
We thank all subjects for cooperation with the research.
Funding
The experiment was founded by knowledge grant of Polish Ministry of Science and Higher Education No 0034/RS3/2015/53.
Availability of data and materials
The datasets used and/or analyzed during the current study is available from the corresponding author on reasonable request.
References
- AnaneLHEdwardsKMBurnsVEDraysonMTRiddellNEvan ZantenJVWallaceGRMillsPJBoschJAMobilization of gammadelta T lymphocytes in response to psychological stress, exercise, and beta-agonist infusionBrain Behav Immun200923 6 823 829 1:CAS:528:DC%2BD1MXosVGhtbw%3D https://doi.org/10.1016/j.bbi.2009.03.003 19318122
- PedersenBKHoffman-GoetzLExercise and the immune system: regulation, integration, and adaptationPhysiol Rev200080 3 1055 1081 1:CAS:528:DC%2BD3cXlsFWjsLc%3D https://doi.org/10.1152/physrev.2000.80.3.1055 10893431
- WalshNPGleesonMPyneDBNiemanDCDhabharFSShephardRJOliverSJBermonSKajenieneAPosition statement. Part two: maintaining immune healthExerc Immunol Rev201117 64 103 21446353
- NiemanDCIs infection risk linked to exercise workload?Med Sci Sports Exerc200032 7 Suppl S406 S411 1:STN:280:DC%2BD3M%2FlvVOrsw%3D%3D https://doi.org/10.1097/00005768-200007001-00005 10910297
- GleesonMPyneDBRespiratory inflammation and infections in high-performance athletesImmunol Cell Biol201694 2 124 131 1:CAS:528:DC%2BC2MXitVymtbzF https://doi.org/10.1038/icb.2015.100 26568028
- GleesonMBishopNOliveiraMMcCauleyTTaulerPMuhamadASRespiratory infection risk in athletes: association with antigen-stimulated IL-10 production and salivary IgA secretionScand J Med Sci Sports201222 3 410 417 1:STN:280:DC%2BC38zjvVSgug%3D%3D https://doi.org/10.1111/j.1600-0838.2010.01272.x 21385218
- PeakeJMNeubauerODella GattaPANosakaKMuscle damage and inflammation during recovery from exerciseJ Appl Physiol2016122 3 559 570 https://doi.org/10.1152/japplphysiol.00971.2016 28035017
- KrügerKLechtermannAFobker M VölkerKMoorenFCExercise-induced redistribution of T lymphocytes is regulated by adrenergic mechanismsBrain Behav Immun200822 3 324 338 https://doi.org/10.1016/j.bbi.2007.08.008 17910910
- AdamsGRZaldivarFPNanceDMKodeshERadom-AizikSCooperDMExercise and leukocyte interchange among central circulation, lung, spleen, and muscleBrain Behav Immun201125 4 658 666 https://doi.org/10.1016/j.bbi.2011.01.002 21238578 4666294
- WitardOCTurnerJEJackmanSRTiptonKDJeukendrupAEKiesAKBoschJAHigh-intensity training reduces CD8+ T-cell redistribution in response to exerciseMed Sci Sports Exerc201244 9 1689 1697 1:CAS:528:DC%2BC38Xht1emsr%2FK https://doi.org/10.1249/MSS.0b013e318257d2db 22525761
- BeyerMClassenSEndlEKochanekMWeihrauchMRDebey-PascherSKnollePASchultzeJLComparative approach to define increased regulatory T cells in different cancer subtypes by combined assessment of CD127 and FOXP3Clin Dev Immunol20112011 734036 https://doi.org/10.1155/2011/734036 21904560 3166761
- BuiJDUppaluriRHsiehCSchreiberRDComparative analysis of regulatory and effector T cells in progressively growing versus rejecting tumors of similar originsCancer Res200666 14 7301 7309 1:CAS:528:DC%2BD28XmvFGkur4%3D https://doi.org/10.1158/0008-5472.CAN-06-0556 16849580
- ChattopadhyaySMehrotraSChhabraAHegdeUMukherjiBChakrabortyNEffect of CD4+CD25+ and CD4+CD25- T regulatory cells on the generation of cytolytic T cell response to a self but human tumor-associated epitope in vitroJ Immunol2006176 2 984 990 1:CAS:528:DC%2BD28Xht1Giuw%3D%3D https://doi.org/10.4049/jimmunol.176.2.984 16393984
- KukrejaACostGMarkerJZhangCSunZLin-SuKTenSSanzMExleyMWilsonBPorcelliSMaclarenNMultiple immuno-regulatory defects in type-1 diabetesJ Clin Invest2002109 1 131 140 1:CAS:528:DC%2BD38Xjtlartg%3D%3D https://doi.org/10.1172/JCI0213605 11781358 150819
- MillsKGRegulatory T cells: friend or foe in immunity to infection?Nat Rev Immunol.20044 11 841 855 1:CAS:528:DC%2BD2cXptFegtLc%3D https://doi.org/10.1038/nri1485 15516964
- MiyaraMAmouraZParizotCBadoualCDorghamKTradSKambouchnerMValeyreDChapelon-AbricCDebréPPietteJCGorochovGThe immune paradox of sarcoidosis and regulatory T cellsJ Exp Med2006203 2 359 370 https://doi.org/10.1084/jem.20050648 16432251 2118208
- SatoEOlsonSHAhnJBundyBNishikawaHQianFJungbluthAAFrosinaDGnjaticSAmbrosoneCKepnerJOdunsiTRitterGLeleSOdunsiKIntraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancerProc Natl Acad Sci U S A2005102 51 18538 18543 1:CAS:528:DC%2BD28Xpt1Om https://doi.org/10.1073/pnas.0509182102 16344461 1311741
- SmythMJTengMLSwannJKyparissoudisKGodfreyDIHayakawaYCD4+CD25+ T regulatory cells suppress NK cell-mediated immunotherapy of cancerJ Immunol2006176 3 1582 1587 1:CAS:528:DC%2BD28XlsFynsw%3D%3D https://doi.org/10.4049/jimmunol.176.3.1582 16424187
- ZouWRegulatory T cells, tumor immunity and immunotherapyNat Rev Immunol20066 4 295 307 1:CAS:528:DC%2BD28XivVyru7k%3D https://doi.org/10.1038/nri1806 16557261
- Spychalski Ł. [Regulatory T lymphocytes in a microenvironment of pleural effusion in lung cancer. Doctoral dissertation from the Department of Pulmonology, Allergology and Pulmonological Oncology]. Poznan 2010.
- HoffmannPErmannJEdingerMFathmanCGStroberSDonor-type CD4(+)CD25(+) regulatory T cells suppress lethal acute graft-versus-host disease after allogeneic bone marrow transplantationJ Exp Med2002196 3 389 399 1:CAS:528:DC%2BD38XmtVOhtLY%3D https://doi.org/10.1084/jem.20020399 12163567 2193938
- PiccirilloCAShevachEMCutting edge: control of CD8+ T cell activation by CD4+CD25+ immunoregulatory cellsJ Immunol2001167 3 1137 1140 1:CAS:528:DC%2BD3MXls1WktL0%3D https://doi.org/10.4049/jimmunol.167.3.1137 11466326
- TrzonkowskiPSzmitEMyśliwskaJDobyszukAMyśliwskiACD4+CD25+ T regulatory cells inhibit cytotoxic activity of T CD8+ and NK lymphocytes in the direct cell-to-cell interactionClin Immunol2004112 3 258 267 1:CAS:528:DC%2BD2cXmslOgsro%3D https://doi.org/10.1016/j.clim.2004.04.003 15308119
- GhiringhelliFMénardCTermeMFlamentCTaiebJChaputNPuigPENovaultSEscudierBVivierELecesneARobertCBlayJYBernardJCaillat-ZucmanSFreitasATurszTWagner-BallonOCapronCVainchenckerWMartinFZitvogelLCD4+CD25+ regulatory T cells inhibit natural killer cell functions in a transforming growth factor-beta-dependent mannerJ Exp Med2005202 8 1075 1085 1:CAS:528:DC%2BD2MXhtFelu7rL https://doi.org/10.1084/jem.20051511 16230475 2213209
- SakaguchiSNaturally arising CD4 + regulatory T cells for immunolgic self-tolerance and negative control of immune responsesAnn Rev Immunol200422 1 531 562 1:CAS:528:DC%2BD2cXktlOgsLo%3D https://doi.org/10.1146/annurev.immunol.21.120601.141122
- KunzmannVKimmelBHerrmannTEinseleHWilhelmMInhibition of phosphoantigen-mediated gammadelta T-cell proliferation by CD4+ CD25+ FoxP3+ regulatory T cellsImmunol2009126 2 256 267 1:CAS:528:DC%2BD1MXhsVCitrk%3D https://doi.org/10.1111/j.1365-2567.2008.02894.x
- HandzlikMKShawAJDungeyMBishopNCGleesonMThe influence of exercise training status on antigen-stimulated IL-10 production in whole blood culture and numbers of circulating regulatory T cellsEur J Appl Physiol2013113 7 1839 1848 1:CAS:528:DC%2BC3sXpsVOrsrw%3D https://doi.org/10.1007/s00421-013-2614-y 23435500
- GleesonMBishopNCThe T cell and NK cell immune response to exerciseAnn Transplant200510 4 43 48 17037088
- SteensbergAToftADBruunsgaardHSandmandMHalkjaer-KristensenJPedersenBKStrenuous exercise decreases the percentage of type 1 T cells in the circulationJ Appl Physiol200191 4 1708 1712 1:CAS:528:DC%2BD3MXnsFSls7g%3D https://doi.org/10.1152/jappl.2001.91.4.1708 11568154
- WangJSongHTangXYangYVieiraVJNiuYMaYEffect of exercise training intensity on murine T-regulatory cells and vaccination responseScan J Med Sci Sports201222 5 643 652 1:STN:280:DC%2BC38zjvVGgsg%3D%3D https://doi.org/10.1111/j.1600-0838.2010.01288.x
- WilsonLDZaldivarFPSchwindtCDWang-RodriguezJCooperDMCirculating T-regulatory cells, exercise and the elite adolescent swimmerPediatr Exerc Sci200921 3 305 317 https://doi.org/10.1123/pes.21.3.305 19827454 4822516
- PerryCPickMBdolachNHazan-HaleviIKaySBerrIRechesAHarishanuYGrisaruDEndurance exercise diverts the balance between Th17 cells and regulatory T cellsPLoS One20138 10 e74722 1:CAS:528:DC%2BC3sXhs1egsrfI https://doi.org/10.1371/journal.pone.0074722 24130669 3793976
- BorowitzkaMMEGershwinASpirulina in human nutrition and healthJ Appl Phycol200921 6 747 748 https://doi.org/10.1007/s10811-009-9467-0
- WuQLiuLMironAKlímováBWanDKučaKThe antioxidant, immunomodulatory, and anti-inflammatory activities of spirulina: an overviewArchiv Toxicol201690 8 1817 1840 1:CAS:528:DC%2BC28XptlCntbw%3D https://doi.org/10.1007/s00204-016-1744-5
- AkaoYEbiharaTMasudaHSaekiYAkazawaTHazekiKMatsumotoMSeyaTEnhancement of antitumor natural killer cell activation by orally administered spirulina extract in miceCancer Sci2009100 8 1494 1501 1:CAS:528:DC%2BD1MXhtVWnsLvE https://doi.org/10.1111/j.1349-7006.2009.01188.x 19432881
- TornabeneTBourneTRaziuddinSBen-AmotzALipid and lipopolysaccharide constituents of cyanobacterium Spirulina Platensis (Cyanophyceae, Nostocales)Mar Ecol Prog Ser198522 121 125 1:CAS:528:DyaL2MXitVSrs78%3D https://doi.org/10.3354/meps022121
- HirahashiTMatsumotoMHazekiKSaekiYUiMSeyaTActivation of the human innate immune system by spirulina: augmentation of interferon production and NK cytotoxicity by oral administration of hot water extract of Spirulina PlatensisInt Immunopharmacol20022 4 423 434 1:CAS:528:DC%2BD38XivFylt7g%3D https://doi.org/10.1016/S1567-5769(01)00166-7 11962722
- AyehunieSBelayABabaTWRuprechtRMInhibition of HIV-1 replication by an aqueous extract of Spirulina Platensis (Arthrospira Platensis)J Acquir Immune Defic Syndr199818 1 7 12 1:STN:280:DyaK1c3lsVartg%3D%3D https://doi.org/10.1097/00042560-199805010-00002
- HayashiKHayashiTKojimaIA natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina Platensis: in vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus activitiesAIDS Res Hum Retrovir199612 15 1463 1471 1:CAS:528:DyaK28XmsFOksL4%3D https://doi.org/10.1089/aid.1996.12.1463 8893054
- LøbnerMWalstedALarsenRBendtzenKNielsenCHEnhancement of human adaptive immune responses by administration of a high-molecular-weight polysaccharide extract from the cyanobacterium Arthrospira PlatensisJ Med Food200811 2 313 322 https://doi.org/10.1089/jmf.2007.564 18598175
- MaertenPShenCBullensDAVan AsscheGVan GoolSGeboesKRutgeertsPCeuppensJLEffects of interleukin 4 on CD25+CD4+ regulatory T cell functionJ Autoimmun200525 2 112 120 1:CAS:528:DC%2BD2MXht1ars7%2FE https://doi.org/10.1016/j.jaut.2005.04.001 16051465
- MaoTKVan de WaterJGershwinMEEffects of a spirulina-based dietary supplement on cytokine production from allergic rhinitis patientsJ Med Food20058 1 27 30 1:CAS:528:DC%2BD2MXjsFCgurY%3D https://doi.org/10.1089/jmf.2005.8.27 15857205
- NielsenCHBalachandranPChristensenOPughNDTamtaHSufkaKJWuXWalstedASchjørring-ThyssenMEnevoldCPascoDSEnhancement of natural killer cell activity in healthy subjects by Immulina®, a spirulina extract enriched for Braun-type lipoproteinsPlanta Med201076 16 802 808 https://doi.org/10.1055/s-0030-1250043
- GaoQQiuSFanJZhouJWangXXiaoYXuYLiYTangZIntratumoral balance of regulatory and cytotoxic T cells is associated with prognosis of hepatocellular carcinoma after resectionJ Clin Oncol200725 18 2586 2593 https://doi.org/10.1200/JCO.2006.09.4565 17577038
- JaroszM Nutrition standards for the polish population - amendment2012 Warszawa Copyright by Instytut Żywności i Żywienia
- HuehnJSiegmundKLehmannJUSiewertCHauboldUFeuererMDebesGFLauberJFreyOPrzybylskiGKNiesnerUde la RosaMSchmidtCABräuerRBuerJScheffoldAHamannADevelopmental stage, phenotype, and migration distinguish naive- and effector/memory-like CD4+ regulatory T cellsJ Exp Med2004199 3 303 313 1:CAS:528:DC%2BD2cXhtVCitL8%3D https://doi.org/10.1084/jem.20031562 14757740 2211798
- MenningAHöpkenUESiegmundKLippMHamannAHuehnJDistinctive role of CCR7 in migration and functional activity of naive- and effector/memory-like Treg subsetsEur J Immunol200737 6 1575 1583 1:CAS:528:DC%2BD2sXntVSgs7g%3D https://doi.org/10.1002/eji.200737201 17474155
- ZhangNSchröppelBLalGJakubzickCMaoXChenDYinNJessbergerROchandoJCDingYBrombergJSRegulatory T cells sequentially migrate from inflamed tissues to draining lymph nodes to suppress the alloimmune responseImmunity200930 3 458 469 1:CAS:528:DC%2BD1MXltFSmt7c%3D https://doi.org/10.1016/j.immuni.2008.12.022 19303390 2737741
- MorgadoJMRamaLSilvaIde Jesus InácioMHenriquesALaranjeiraPPedreiroSRosadoFAlvesFGleesonMPaisMLPaivaATeixeiraAMCytokine Production by monocytes, neutrophils, and dendritic cells is hampered by long-term intensive training in elite swimmersEur J Appl Physiol2012112 2 471 482 1:CAS:528:DC%2BC38XpslOquw%3D%3D https://doi.org/10.1007/s00421-011-1966-4 21584686
- GabrielHHUrhausenAValetGHeidelbachUKindermannWOvertraining and immune system: a prospective longitudinal study in endurance athletesMed Sci Sports Exerc199830 7 1151 1157 1:CAS:528:DyaK1cXks12htLY%3D https://doi.org/10.1097/00005768-199807000-00021 9662688
- RowbottomDGKeastDGoodmanCMortonARThe haematological, biochemical and immunological profile of athletes suffering from the overtraining syndromeEur J Appl Physiol Occup Physiol199570 6 502 509 1:STN:280:DyaK28%2FivVOhuw%3D%3D https://doi.org/10.1007/BF00634379 7556122
- BlumKSPabstRLymphocyte numbers and subsets in the human bloodDo they mirror the situation in all organs? Immunol Lett2007108 1 45 51 1:CAS:528:DC%2BD2sXht1Sjtw%3D%3D 17129612
- DhabharFSMillerAHSpencerRLStress-induced changes in blood leukocyte distribution. Role of adrenal steroid hormonesJ Immunol1996157 4 1638 1644 1:CAS:528:DyaK28XkvFWhu70%3D 8759750
- TeixeiraAMRamaLCarvalhoHMBorgesGCarvalheiroTGleesonMAlvesFTrindadeHPaivaAChanges in naïve and memory T-cells in elite swimmers during a winter training seasonBrain Behav Immun201439 186 193 1:CAS:528:DC%2BC2cXhtlGls7w%3D https://doi.org/10.1016/j.bbi.2014.01.002 24412212
- DimitrovSLangeTBomJSelective mobilization of cytotoxic leukocytes by epinephrineJ Immunol2010184 l 503 5ll 1:CAS:528:DC%2BD1MXhsFOnsLfJ https://doi.org/10.4049/jimmunol.0902189 19949113
- KaraszewskiJWRederATAnlarBKimWCAmasonBGIncreased lymphocyte beta-adrenergic receptor density in progressive multiple sclerosis is specific for the CD8+, CD28- suppressor cellAnn Neurol199130 l 42 47 1:STN:280:DyaK38%2FislGhtQ%3D%3D https://doi.org/10.1002/ana.410300109 1656847
- HongSJohnsonTAFaragNHGuyHJMatthewsSCZieglerMGMillsPJAttenuation of T-lymphocyte demargination and adhesion molecule expression in response to moderate exercise in physically fit individualsJ Appl Physiol200598 3 1057 1063 1:CAS:528:DC%2BD2MXisVCjsL4%3D https://doi.org/10.1152/japplphysiol.00233.2004 15501922
- MillsPJHongSRedwineLCarterSMChiuAZieglerMGDimsdaleJEMaiselASPhysical fitness attenuates leukocyte-endothelial adhesion in response to acute exerciseJ Appl Physiol2006101 3 785 788 1:CAS:528:DC%2BD28XpvVCkurw%3D https://doi.org/10.1152/japplphysiol.00135.2006 16728524
- PaulsenGCrameriRBenestadHBFjeldJGMørkridLHallénJRaastadTTime course of leukocyte accumulation in human muscle after eccentric exerciseMed Sci Sports Exerc201042 1 75 85 https://doi.org/10.1249/MSS.0b013e3181ac7adb 20010127
- TidballJGVillaltaSARegulatory interactions between muscle and the immune system during muscle regenerationAm J Physiol Regul Integr Comp Physiol2010298 5 R1173 R1187 1:CAS:528:DC%2BC3cXmt1Klsrc%3D https://doi.org/10.1152/ajpregu.00735.2009 20219869 2867520
- ChinenTKannanAKLevineAGFanXKleinUZhengYGasteigerGFengYFontenotJDRudenskyAYAn essential role for the IL-2 receptor in Treg cell functionNat Immunol20167 11 1322 1333 https://doi.org/10.1038/ni.3540
- SetoguchiRHoriSTakahashiTSakaguchiSHomeostatic maintenance of natural 3(+) CD25(+) CD4(+) regulatory T cells by interleukin (IL)-2 and induction of autoimmune disease by IL-2 neutralizationJ Exp Med2005201 5 723 735 1:CAS:528:DC%2BD2MXitl2rt78%3D https://doi.org/10.1084/jem.20041982 15753206 2212841