
Abstract
Background
Altitude training is a common practice among middle-distance and marathon runners. During acclimatization, sympathetic drive may increase resting metabolic rate (RMR), therefore implementation of targeted nutritional interventions based on training demands and environmental conditions becomes paramount. This single case study represents the first nutritional intervention performed under hypobaric hypoxic conditions (3900 m) in Paralympic sport. These results may elucidate the unique nutritional requirements of upper body endurance athletes training at altitude.
Case presentation
This case study examined the effects of a nutritional intervention on the body mass of a 36-year-old professional wheelchair athlete (silver medalist at the Paralympic Games and 106 victories in assorted road events) during a five-week altitude training camp, divided into pre-altitude at sea level (BN), acclimatization to altitude (Puno, 3860 m) (BH), specific training (W1,2,3,4) and return to sea level (Post) phases. Energy intake (kcal) and body mass (kg) were recorded daily. Results demonstrated significant decrease in body mass between BN and BH (52.6 ± 0.4 vs 50.7 ± 0.5 kg, P < 0.001) which returned to pre-altitude values, upon returning to sea level at Post (52.1 ± 0.5 kg). A greater daily intake was observed during BH (2899 ± 670 kcal) and W1,2,3 (3037 ± 490; 3116 ± 170; 3101 ± 385 kcal) compared to BN (2397 ± 242 kcal, P < 0.01) and Post (2411 ± 137 kcal, P < 0.01). No differences were reported between W4 (2786 ± 375 kcal), BN and Post. The amount of carbohydrates ingested (g · kg− 1) was greater in W1,2,3, (9.6 ± 2.1; 9.9 ± 1.2; 9.6 ± 1.2) than in BN (7.1 ± 1.2) and Post (6.3 ± 0.8, P < 0.001). Effect sizes (Cohen’s d) for all variables relative to BN (all time points) exceed a large effect (d > 0.80).
Conclusions
These results suggest an elite wheelchair marathoner training at 3860 m required increased nutrient requirements as well as the systematic control needed to re-adapt a nutritional program. Moreover, our findings highlight training and nutritional prescription optimization of elite wheelchair athletes, under challenging environmental conditions.
Background
In recent years, there has been emerging interest in the optimization of nutritional strategies to help athletes reach their fitness goals during hypoxic training conditions [Citation1]. However, nutritional guidelines for athletes training at 4000 m altitude remain unclear as most nutritional and exercise metabolism studies have been completed at lower altitudes [Citation1, Citation2], and the data reflects athletes participating in activities less than marathon distances [Citation3–Citation9]. For example, in distance running only one study has examined well-trained runners at an altitude of 4000 m [Citation10] and, recently a case study reported physiological data on an elite wheelchair marathoner training at 3900 m altitude [Citation11]. Loss of body fat and fat free mass have been reported during high altitude sojourns in people eating ad libitum [Citation12–Citation15], suggesting that strict altitude imposed dietary controls can attenuate daily energy deficits and partially mitigate weight loss [Citation16]. Loss of fat free mass at high altitude increases the risk of illness and injury in extreme environments [Citation5, Citation17–Citation19]. During acclimatization there is a reduction of intra and extracellular water combined with a decrease in plasma volume [Citation6, Citation20], which can result in body mass loss up to 2 kg [Citation14]. Furthermore, during acute phase exposure, total exogenous glucose oxidation appears to be lower than at sea level, and after 21 days of initial exposure at 4300 m not reaching sea level, suggesting oxidation rates under hypoxic conditions do not cover the energy demands of athletes at altitude [Citation9]. Alternatively, other studies suggest individuals have an increased dependence on glucose as a fuel source at high altitude, especially during exercise [Citation3, Citation7, Citation8].
Increased resting metabolic rate (RMR) has also been observed at altitude, which could be due to increased sympathetic drive and subsequent rise in adrenaline levels [Citation21]. Recent research found that RMR in elite middle-distance runners increased by ≈ 19% at a moderate altitude (2100 m) compared to sea-level conditions [Citation2] and 10% at high altitude (3800 m) [Citation22]. In contrast, a small decrease in RMR was reported in a group of Olympic rowers training at 1800 m [Citation23]. Moreover, RMR is more pronounced over the first 2–3 days after arrival [Citation16, Citation24]. However, elevated RMR (≥ 17%) can persist for up to 21 d after initial high altitude exposure [Citation17]. Ultimately, energy expenditure which is elevated at altitude may be equivalent to high intensity exercise conducted at sea level [Citation25].
Due to the aforementioned factors, one of the primary nutritional goals for managing a successful altitude training camp involves matching the energy intake to the daily expenditure in order to minimize body mass loss [Citation26]. In fact, it was reported that, a total of 7.6 g · kg− 1 body mass of carbohydrates (CHO) per day did not cover the energetic demands of cyclists living and training at 4300 m [Citation4]. Importantly, up to 70% of the chronic altitude exposure-related weight loss is said to be due to reductions in muscle mass itself [Citation27]. To consider, D’Hulst & Deldique [Citation28] recently suggested that based on the hypoxic dose theory [Citation29], an exposure of 5000 km · h− 1 is the cutoff point above which muscle loss starts to occur. However, at altitude the stimulation of protein synthesis after exercise might be blunted by hypoxia, as it was shown that increase in muscle protein synthesis following walking at 4559 m [Citation30] was much lower than a comparable study with exercise performed at sea level [Citation31]. Interestingly, in a separate study, body mass was maintained in ski mountaineers following an isocaloric diet of 4000 kcal · d− 1, supplemented with 1.5 g or 2.5 g · kg body mass casein protein per day during seven days at 2500–3800 m [Citation32]. Moreover Bigard and colleagues examined the effects of branch chain amino acids (BCAA) (7.8 g leucine, 3.4 g isoleucine, 11.2 g valine; 1.44 g protein · kg · d) compared to carbohydrate supplementation on body composition following six days of ski mountaineering at 2500–3800 m. Body composition and muscular performance were unaffected by BCAA. However, significant weight loss only occurred in the carbohydrate-supplemented group (− 1.55 vs. -0.8 kg) [Citation32].
The purpose of this study was to examine the effects of a nutritional intervention on the body mass of an elite wheelchair marathoner during a five-week training camp performed between sea level and 3900 m altitude. The intervention was designed to anticipate increases in RMR due to the combined effects of both environmentally induced hypoxia and the demands of marathon training.
Case presentation
The study athlete was a 36-year-old, elite wheelchair marathoner, functional class T52 (upper limb involvement category). Some of his accolades include winning a silver medal at the Paralympic Games and 106 victories in assorted road events, including a win at the 2016 Boston Marathon, ten weeks after returning to sea level from Los Andes (Peruvian Altiplano). Our participant’s height = 1.76 m; body mass = 52.6 ± 0.4 kg; power output at second ventilatory threshold = 62 W; training 8000 km per year; former world record holder in the T52 division in 800 m (1 min:56 s); 1500 m (3 min:36 s); world record holder in 5000 m (12 min:37 s); half marathon (50 min:28 s) and fourth best ever time in marathon (1 h:42 min:05 s). Additionally, he has more than ten years of altitude training experience, with training camps performed in Boulder, CO (1655 m), Navacerrada, Spain (1858 m), Flagstaff, AZ (2106 m), Sierra Nevada, Spain (2320 m), Keystone, CO (2796 m) and Breckenridge, CO (2926 m), performing both altitude models: Live-High-Train-High (LHTH) and Live-High-Train-Low (LHTL) and has been exposed to more than 8000 h of normobaric-hypoxia. For the last five seasons prior to the current study, the athletes trained at moderate altitudes (1655 up to 2926 m) for: 78, 82, 101, 79 and 62 days.
The athlete requested advice for the development of an individualized nutritional program based on training loads to prepare for his upcoming season. Therefore, after consultation with laboratory members a nutrition program was designed, according to his training load (Table ).
Table 1 Main meals designed for each type of session under altitude conditions
The research participant provided written consent prior to participation in the current study and read the manuscript before submission. Research was approved by the Ethics Research Committee of the University Miguel Hernandez.
Training protocol
Both pre-altitude (BN), at 16 m and acclimatization (BH) at 3900 m incorporated identical training loads (128 km of mileage each). However, the first two days of BH incorporated no training to minimize the effects of jet-lag, and acute mountain symptoms (AMS), like headache [Citation33]. Two daily training sessions were performed from Wednesday to Friday under the first ventilatory threshold (<VT1). The morning session involved 20 km of distance training and the afternoon session 16 km. A 20 km workout was performed on Saturday <VT1. Sunday was a rest day. Specific training weeks “W1, W2, W3 & W4” were based on a day-to-day basis periodization, according to level of heart rate variability (HRV) [Citation34]. When the HRV reached a reference value (RV), the subject completed a specific session in the morning, followed by an evening off. If the RV was not reached, two workouts <VT1 were performed: 20 km in the morning and 16 km in the afternoon. On three days the training was fixed; On Mondays and Thursdays the AM sessions were 16 km < VT1, while the PM sessions involved resistance training and Sundays were off. The specific sessions were known as: A (20 × 400 m at ~ second ventilatory threshold (VT2) in a plateau at 4090 m altitude; recovery reps: 75 s); B (30 km ~ VT1) and C (6 × 2000 m ~ VT2 in a plateau at 4090 m altitude; recovery reps: 120 s).
As a way to induce muscle hypertrophy, resistance sessions were performed at 80% of 1 RM [Citation35] with 4 sets of 8 reps with 150 s recovery, aimed at avoiding loss of muscle mass induced by chronic hypoxia. RM test was not performed under altitude conditions due to high risk of injury, so it was done four days before flying to Peru. More details on the experimental design have been reported previously [Citation11].
Daily recording
Throughout the experiment, basal body mass was recorded in fasting conditions, naked, after waking up, with a digital scale (Tanita BC-601®, TANITA Corporation, Tokyo, Japan). Utilizing a food recording system previously reported [Citation1], a nutritional diary was maintained by the subject to record daily intake, which included main meals (breakfast, lunch and dinner), two small snacks and all training activities that occurred post-intake (Figs. and ).
Fig. 1 Timing of Daily Food and Fluid Intake during Altitude, based on different training routines
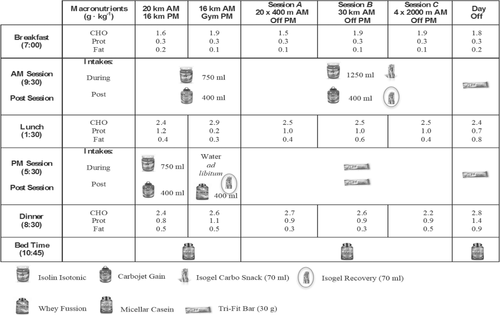
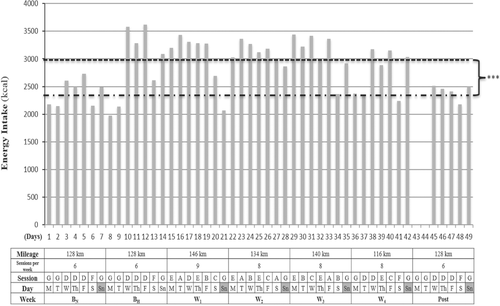
Fig. 2 Training program and energy intake during BN, BH, W1,2,3,4 and Post. BN, baseline in normoxia; BH, baseline in hypoxia; W1,2,3,4, specific training weeks in hypoxia; Post, returning sea level week. Session A: performed on a plateau at 4090 m; 8 km + technique drills + 5 × 80 m accelerations + 20 × 400 m ~ VT2 + 2 km. Recovery reps 75 s. Session B: 2 h ~ VT1. Session C: performed on a plateau at 4090 m; 8 km + technique drills + 5 × 80 m accelerations + 6 × 2000 m ~ VT2 + 2 km. Recovery reps 120 s. Session D: 20 km < VT1 in the morning + 16 km < VT1 in the afternoon. Session E: 16 km < VT1 in the morning + gym session in the afternoon (4 sets × 8 reps recovery sets 150 s at 80% RM). Exercises for resistance session: press bench, close grip, dumbbell press, seated military press and seated cable row). Session F: 20 km < VT1 in the morning + resting afternoon. Session G: Day off. Dash line: Represent mean energy intake (2423 kcal) in normoxic conditions at sea level. Round dot line: Represent mean energy intake (3017 kcal) in hypoxic conditions at 3900 m altitude. Differences from mean energy intake under hypoxic conditions: *** P < 0.001
Total energy (kcal), carbohydrates, proteins and fats (g · kg− 1 body mass) were estimated according to nutritional composition database supported by the Spanish Ministry of Science and Innovation [Citation36].
Nutritional program
The athlete was instructed by a nutritionist to prepare all meals which included weighing both ingredients prior to cooking and left overs prior to disposal. On days when the athlete ate at restaurants, which occurred on four occasions, he was instructed to send pictures of these meals to the research team [Citation37]. A personal chef was contacted to buy and cook all foods/ingredients for the athlete on a daily basis according to athlete instructions while the weighing and cooking process occurred under the athlete’s supervision. Additionally, the athlete was instructed to prepare all training drinks and post-training recovery solutions. To prevent contamination, the athlete did not eat raw foods or unpeeled fruits or vegetables and no water from the tap was consumed [Citation38]. At sea level the athlete cooked all meals at home.
Daily energy intake was increased ~ 20% from pre-altitude (BN), to arrival at altitude (BH) to avoid body mass loss from increased RMR which is common while living and training at higher altitudes [Citation2, Citation22]. Moreover, main meals were designed according to the type of training session performed (Fig. ), as we have recently reported that during specific training weeks (W1,2,3,4) number of A,B,C, sessions differed between specific training weeks, according to a training program based in HRV [Citation11], which explains why at W2 the greatest amount of CHO was ingested (9.9 ± 1.2 g · kg− 1 body mass), and why during BH and W4 the total amount of CHO tended to be lower than W1,2,3 ( ). Moreover, main meals were accompanied by two rich-carbohydrate snacks, based on reports that the inclusion of several rich carbohydrate snacks, more optimally covers increased energy requirements than three standalone main meals [Citation38]. Furthermore, regarding proteins, a minimum intake of 2.4 g · kg− 1 body mass was targeted in the current nutritional design to avoid loss of lean mass [Citation39]. To avoid gastrointestinal issues (GI) and fullness [Citation40], a low protein/fat intake was provided for breakfast and PM sessions, however the percentage of lipids at lunch was lower than dinner. Protein intake at lunch and dinner were ≈ 1 g · kg− 1, given that specific and, more demanding sessions (A,B,C) were performed in the morning, and muscle tissue repair is a main meal target. The ingestion of lipids was set at a minimum of 1 g · kg− 1 body mass throughout the sea level and altitude camps, as fat cells increase their sensitivity to hormonal stimulation after training, resulting in a greater mobilization of fatty acids [Citation41]. Moreover, an Iso-Lyn Isotonic (AMIX) sports drink was used for workouts < VT1 shorter than 65 min (20 and 16 km). The athlete was instructed to drink a solution with 750 ml of water and 56.4 g of CHO, while a solution of 1250 ml with 80 g of CHO was recommended for specific sessions. The CHO rate was 0.5 to 1 g · kg− 1 body mass per hour [Citation42]. Despite these recommendations, the athlete and team elected to preserve his natural drinking habits that involved consuming drinks every 10 min. This decision was made because fluid consumption for a wheelchair racer can be precarious during propulsion, as they must come out of their natural prone/kneeling body position to drink. This action can force loss of vision, which increases the risk of collision or crashing. Because our participant never experienced GI in his career with the use of carb gels [Citation43], he drank a 42 g CHO (Glucose + Fructose) Iso-Gel carbo snack (AMIX) during specific sessions workouts [Citation44]. Gels were consumed in the A session after fourteen 400 m rep, in the B session 90 min after starting, and in the C session after four 2000 m rep. Both types of carbs used in the solution and gels were multiple transportable carbohydrates, as directed by Jeukendrup [Citation45].
During gym sessions water was consumed ad libitum and immediately after gym sessions the athlete co-ingested a rich leucine whey protein (23.6 g) (Whey Fussion, AMIX) dissolved in 400 ml of water and a carbohydrate gel (Iso-Gel Recovery, AMIX) (37.6 g maltodextrin + fructose + Vitargo®) as directed for speeding up to 25% glycogen synthesis [Citation46]. For refueling purposes carbohydrate guidelines [Citation42], suggest aiming for post-exercise rapid recovery of muscle glycogen deposits, with 1 g · kg− 1 body mass of CHO, repeated every 2–3 h. After specific sessions, a carbohydrate shake was taken with a carbohydrate gel, providing 1.4 g · kg− 1 body mass. In the hour immediately after 16 km and 20 km < VT1, the subject drank a carbohydrate solution (Carbojet Gain, AMIX) (34 g CHO, 7.5 g prot, 1.8 g fat) dissolved in 400 ml of water, and after specific sessions he ingested a combination of the same drink plus Iso-Gel Recovery. To consider, 2.4 g · kg− 1 body mass, CHO were consumed (Fig. ) at lunch which occurred approximately two hours post-exercise meal, in order to achieve 3.1 g · kg− 1 body mass of CHO 3 h post-exercise for our athlete vs. 3 g · kg− 1 body mass as suggested by Burke and colleagues [Citation42].
On specific session days, rest was provided in the evenings along with a snack at 5:30 PM, to meet increased energy requirements [Citation38]. This snack included two 30 g cereal bars (Tri-Fit Bar, AMIX) (34.9 g CHO, 3.9 g prot, and 10.1 g fat).
In a manner to avoid loss of body mass [Citation32] and enhance muscle protein synthesis [Citation47] the athlete consumed 2.5 g leucine, 1.5 g isoleucine, and 1.5 g valine) immediately after each session (BCAA Elite Rate, AMIX). Before bedtime, 30 g of casein protein (Micellar Casein, AMIX) (1.7 g CHO, 24 g prot, 0.6 g fat) was ingested as suggested by Snijders and colleagues [Citation48].
Finally, the athlete maintained iron levels through a daily intake of 105 mg of ferrous sulphate (Ferogradumet®, Ross, Abbott Científica), as ferrous sulphate intake has been related to the production of Hemoglobin and red cells [Citation49, Citation50]. To comply with World Anti-Doping Agency (WADA) regulations, none of the aforementioned supplements contain prohibited substance.
For a description of the macronutrients intake during main meals in each session see Fig. .
Statistical analysis
All data are presented as mean ± SD. A repeated-measures ANOVA was carried out for all the variables including the factor TIME with levels BN, BH, W1, W2, W3, W4 and Post. A post hoc least significance difference (LSD) multiple-range test was performed to determine differences between the factor levels. Effect size (d) associated with change in body mass was calculated using Cohen’s d (difference in mean scores over time divided by pooled SD) with its 95% confidence limits (CL) [Citation51] and were interpreted as trivial (≤ 0.19), small (0.20–0.49), medium (0.50–0.79), and large (≥ 0.80) [Citation52]. An alpha level of 0.05 was stated for statistical significance. Statistical analyses were performed using the SPSS version 22.0 (SPSS, Inc., Chicago, IL, USA) software.
Results
Our nutritional intervention results can be found in Table .
Table 2 Body mass and nutritional parameters during sea level and altitude
Body mass
A significant decrease in body mass was observed from BN to BH [P < 0.001; d = 4.16, 95% CL (2.02; 5.71)] but returned to near baseline levels during Post. There were no significant effect for time during the W1,2,3 period, however we observed a significant increase in body mass from W1 to W4 [P < 0.001; d = 2.35, 95% CL (0.86; 3.51)].
Energy intake
Results show a greater amount of kcal in BH [P < 0.01; d = 0.96, 95% CL (− 0.25; 2.04)] and W1 [P < .01; d = 1.61, 95% CL (0.27; 2.73)], W2 (P < 0.01; d = 3.49, 95% CL (1.59; 4.91)], W3 [P < 0.01; d = 2.15, 95% CL (− 0.66; 3.33)] than in BN. Same differences were observed within BH [P < 0.01; d = 0.97, 95% CL (− 0.24; 2.05)], W1 [P < 0.01; d = 1.68, 95% CL (0.31; 2.80)], W2 [P < 0.01; d = 4.52, 95% CL (2.26; 6.16)], W3 [P < 0.01; d = 2.31, 95% CL (0.78; 3.51)] and Post. No differences were reported between W4, BN and Post.
Carbohydrates
The amount of CHO ingested (g · kg− 1 body mass) was greater in W1 [P < 0.001; d = 1.43, 95% CL (0.12; 2.53)], W2 [P < 0.001; d = 2.33, 95% CL (0.80; 3.54)], W3 [P < 0.001; d = 2.08, 95% CL (0.62; 3.26)] than in BN. Differences were observed within W1 [v0.01; d = 2.01, 95% CL (0.56; 3.17)], W2 [P < 0.01; d = 3.47, 95% CL (1.58; 4.88)], W3 [P < 0.01; d = 3.18, 95% CL (1.38; 4.53)] and Post.
Proteins
Protein intake (g · kg− 1 body mass) was greater in BH (P < 0.001; d = 2.54, 95% CL (0.95; 3.79)] and W1 (P < 0.001; d = 2.03, 95% CL (0.58; 3.20)], W2 (P < 0.001; d = 2.16, 95% CL (0.67; 3.34)], W3 (P < 0.001; d = 2.03, 95% CL (0.58; 3.20)], W4 (P < 0.001; d = 2.31, 95% CL (0.78; 3.52)] than in BN. Same differences were found within BH (P < 0.01; d = 2.38, 95% CL (0.83; 3.59)], W1 (P < 0.01; d = 1.90, 95% CL (0.48; 3.05)], W2 (P < 0.01; d = 1.96, 95% CL (0.52; 3.11)], W3 (P < 0.01; d = 1.90, 95% CL (0.48; 3.05)], W4 (P < 0.01; d = 2.00, 95% CL (0.56; 3.16)] and Post.
Lipids
No differences were found in lipids intake (g · kg− 1 body mass) within any period.
Discussion
The aim of this case study was to assess the effectiveness of an evidence based individualized nutrition program applied to an elite wheelchair marathoner during a five-week altitude training camp, carried out in the Peruvian Altiplano (Puno, Peru) at 3900 m. The program was designed based on existing literature for its ability to sustain the athlete’s body mass and meet the energetic demands of intense training, while promoting substrate availability, nutrient recovery, and muscle tissue repair. Interestingly, the designed nutritional intervention helped to: 1) maintain the athlete’s body mass throughout the altitude camp, 2) minimize performance deficits during intense training at altitude compared to sea level (~ 20 to ~ 24% in 1609 m and 3218 m reps respectively) [Citation10], as evidence by recently reported data demonstrating a ~ 3% reduction in reps (2000 m) [Citation11], 3) facilitate intra-sessions recovery through faster glycogen restoration, helping the athlete to perform during physiological demanding sessions (~ VT2) when completed consecutively, or until two sessions of ~ 2 h at ~ VT1 at W2 [Citation11], and 4) maintain quality training sessions at altitude as evidence by: a) improved power output, 11-d post-altitude compared to 4-d pre-altitude (44 W vs 50 W), b) time reductions during 3000 m races 12-d post-altitude compared to 3-d pre-altitude (472 s vs 456 s) [Citation11].
At 4300 m there can be an increase in respiratory water loss, due to greater ventilation and an increase in urinary water loss that can increase up to 500 ml per day [Citation17]. This could explain the nearly 2 kg weight loss observed from baseline (BN) to acclimatization phase (BH) and the return to pre-altitude levels in post (Table ). It should be noted that there was an increment of energy intake of ≈ 500 kcal in hypoxic conditions compared to normoxic conditions (P = 0.001) and same training was done in BN and BH (Fig. ). Of note, all effect sizes associated with statistically significant changes in body mass far exceeded Cohen’s convention for a large effect.
Increased RMR has been reported in athletes who live and train at altitude [Citation2]. For this reason, to maintain body mass in the current study, there was a significant increase in the amount of carbohydrates per kilogram of body mass and proteins per kilogram of body mass provided at altitude compared to sea level. We suspect that the slight increase in body mass observed in W4 was induced by the different number of specific sessions performed from W1 to W4; 2 in W1, 3 in W2, 2 in W3 and 1 in W4 [Citation11]. To increase energy supply, as a result of a greater energy demands and to avoid GI, six meals (breakfast, post-training AM, lunch, snack or post-training PM, dinner and bedtime) were projected in an elapsed time within three hours each one (Fig. ), as it has been recommended to include several rich carbohydrate snacks, rather than three main meals [Citation38]. We did not find differences in energy intake between acclimatization (BH) and specific training weeks (W1 to W4) however this could be due to the fact that when the athlete performed a specific session in the morning, a rest afternoon was followed, in spite of two sessions performed daily during acclimatization with 36 km volume (Fig. ). Furthermore, we did not consider a slightly lower exogenous glucose oxidation rate during acclimatization and chronic altitude [Citation53], as it has been reported that such observations should be contrasted with fully fed individuals, although evidence exists to the contrary [Citation3, Citation8]. Three hours before training sessions, a rich CHO meal was consumed, as it has been shown to increase glycogen availability [Citation42]. We recommended that the athlete change from cereals to a lower fiber food like white bread to avoid GI distress however because of disability imposed manual dexterity deficits which prevent cutting bread slices and spreading fruit jam, he decided to use cereals. The research team also had to consider that the athlete ate breakfast by seven in the morning, which was nearly two and a half hours before training sessions. However, the athlete commonly practiced training in a fasted state like this during training sessions at home, to minimize GI. Despite the athlete’s comfortability with this practice, it was discarded in Puno because temperatures were extremely cold by 7 am (~ 0 °C) and he trained barefoot.
To avoid a loss of muscle mass, high-protein foods were spread out across all meals (Fig. ), while whey and casein protein training products were consumed to ensure minimum requirements of 2.4 g · kg− 1 body mass were achieved [Citation39]. However, we have to consider that the hypoxic dose [Citation29] of this training camp was 3300 km · h− 1, not reaching the cut off point, where muscle loss begins [Citation28]. Due to personal preferences, protein delivery by meat was introduced at lunch, while fish was eaten at dinner. No eggs were eaten while training however the athlete ate an omelet for lunch during rest days (Table ).
Limitations
Main limitations of this study are evident in the absence of outcomes like upper body skinfolds, and upper arm circumference measurements, which could help us to know if body fat percentage and loss of muscle mass occurred in our athlete which was reported previously in subjects eating ad libitum under hypoxic conditions [Citation12–Citation15]. Moreover, RMR was not assessed, as recently reported [Citation23] in Olympic rowers training at 1800 m who did not show an increase in RMR. However, our athlete was exposed to more intense hypoxic conditions, so sympathoexcitation may have occurred [Citation54] leading to elevated adrenaline levels and subsequent greater energetic demands. Another limitation was evident in the use of a self-reported intake diary conducted without supervision from a nutritionist, however the athlete was provide instructions for meal preparation as described previously. Importantly, similar self reported nutritional tools have been validated for estimating energy and nutrient intake [Citation37]. Also, the use of pictures on four occasions to record restaurant meal consumption must be considered as a limitation. However, this methodology has been supported by exercise nutritionists as a useful strategy, particularly when research teams are not present [Citation1]. Finally, the absence of muscular biopsies did not allow us to measure glycogen and protein muscle content.
Conclusions
The aim of the daily meal distributions (Fig. ) was to cover the energetic demands of training sessions and to ensure substrate availability, nutrients recovery, and muscle tissue repair according to literature recommendations.
This paper can help us to better understand the unique nutritional requirements of upper body endurance athletes during altitude training conditions where nutritional strategies may differ from able-bodied athletes. Importantly, to confirm and expand on the current findings specific to the aforementioned differences between able bodied and upper limb athletes, more research is needed on both populations. However, analogous studies are scarce in able bodied athletes and nonexistent in upper limb athletes. For example, only one study, published in 1967 examined well-trained athletes at 4000 m [Citation10], while others have investigated nutritional interventions or exercise metabolism at moderate altitudes only (2150 m) [Citation1, Citation2]. To date, the only other studies conducted at altitudes similar to ours involved either dissimilar sports disciplines [Citation4], lacked a nutritional component [Citation10], or utilized none elite athletes [Citation53]. Ultimately, this study represents the first nutritional intervention conducted on an elite wheelchair marathoner under altitude conditions. Since no specific nutritional interventions have been performed on able-bodied marathon runners or wheelchair athletes at 4000 m altitude, all nutritional guidelines were reflective of the literature pertaining to able-bodied athletes training at lower altitudes.
Ultimately, our nutritional intervention targeted body mass maintenance to sufficiently anticipate increases in RMR due to the combined effects of environmentally induced hypoxia and the demands of marathon training. Moreover, the intervention helped minimize performance perturbations, facilitated overall recovery, and enhanced athlete performance post-altitude. Future related studies should be designed based on considerations from the current study, however with more specificity therefore utilizing deeper assessment tools like biological samples. For example biopsies could be applied to determine the protein and glycogen synthesis-breakdown cycle of athletes during periods of intense training.
Authors’ contributions
RLG, MMR and GBC conceived the study. RLG, MMR, GBC and SSQ participated in the design of the study. Data were collected by SSQ and analyzed by SSQ, IR, RLG, MMR, TU and GBC. Data interpretation and manuscript preparation were undertaken by SSQ, IR, RLG, MMR, TU and GBC. All authors approved the final manuscript.
Ethics approval and consent to participate
This investigation had prior ethical approval by the Ethics Research Committee of the University Miguel Hernandez.
Consent for publication
The athlete provided consent for publication, after reading the last version of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
| AMS | = | Acute mountain symptoms |
| BCAA | = | Branch chain amino acids |
| BH | = | Altitude acclimatization |
| BN | = | Pre-altitude |
| CHO | = | Carbohydrates |
| CL | = | Confidence limits |
| GI | = | Gastrointestinal issues |
| HRV | = | Heart rate variability |
| LHTH | = | Live-High-Train-High |
| LHTL | = | Live-High-Train-Low |
| RMR | = | Resting metabolic rate |
| RV | = | Reference value |
| SD | = | Standard Deviation |
| VT1 | = | First ventilatory threshold |
| VT2 | = | Second ventilatory threshold |
| W1,W2, W3, W4 | = | Specific training weeks at altitude |
Acknowledgements
The authors wish to thank the athlete who volunteered for this case study.
Funding
No funding was received for this study.
Availability of data and materials
Please contact authors for data requests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- HeikuraIABurkeLMBerglandDUusitaloALTMeroAAStellingwerffTImpact of energy availability, health and sex on hemoglobin-mass responses following live-high-train-high altitude training in elite female and male distance athletesInt J Sports Physiol Perform201813 1090 1096 29431548 https://doi.org/10.1123/ijspp.2017-0547
- WoodsALSharmaAPGarvican-LewisLASaundersPRiceTThompsonKGFour weeks of classical altitude training increases resting metabolic rate in highly trained middle-distance runnersInt J Sport Nutr Exerc Metab201627 83 90 27459673 https://doi.org/10.1123/ijsnem.2016-0116 1:CAS:528:DC%2BC1cXhsFKntLbI
- BrooksGAButterfieldGEWolfeRRGrovesBMMazzeoRSSuttonJRWolfelEEReevesJTIncreased dependence on blood glucose after acclimatization to 4300 mJ Appl Physiol199170 919 927 1:CAS:528:DyaK3MXhs1WlsL0%3D 2022585 https://doi.org/10.1152/jappl.1991.70.2.919 2022585
- FulcoCSKambisKWFriedlanderALRockPBMuzaSRCymermanACarbohydrate supplementation improves time-trial cycle performance during energy deficit at 4300-m altitudeJ Appl Physiol200599 867 876 1:CAS:528:DC%2BD2MXhtVChu7vE 15879171 https://doi.org/10.1152/japplphysiol.00019.2005 15879171
- SergiGImoscopiASartiSPerissinottoECoinAInelmenEMZambonSBusettoLSeresinCManzatoEChanges in total body and limb composition and muscle strength after a 6-8 weeks sojourn at extreme altitude (5000-8000m)J Sports Med Phys Fitness201050 450 455 1:STN:280:DC%2BC3M%2FmtFyjsw%3D%3D 21178932 21178932
- HoytRWDurkotMJKamimoriGHSchoellerDACymermanA SuttonJRCoatsGHoustonCSChronic altitude exposure (4300 m) decreases intracellular and total body water in humansHypoxia and mountain medicine1992 Burlington Queen City printers 306
- RobertsACButterfieldGECymermanAReevesJTWolfelEEBrooksGAAcclimatization to 4300-m altitude decreases reliance on fat as a substrateJ Appl Physiol199681 1762 1771 1:STN:280:DyaK2s%2FmtVGmsg%3D%3D 8904597 https://doi.org/10.1152/jappl.1996.81.4.1762 8904597
- RobertsACReevesJTButterfieldGEMazzeoRSSuttonJRWolfelEEBrooksGAAltitude and beta-blockade augment glucose utilization during submaximal exerciseJ Appl Physiol199680 605 615 1:CAS:528:DyaK28Xhslylt74%3D 8929605 https://doi.org/10.1152/jappl.1996.80.2.605 8929605
- YoungAJBerrymanCEKenefickRWDerosierENMargolisLMWilsonMACarriganCTMurphyNECarboneJWRoodJCPasiakosSMAltitude acclimatization alleviates the hypoxia-induced suppression of exogenous glucose oxidation during steady-state aerobic exerciseFront Physiol20189 830 30038576 6046468 https://doi.org/10.3389/fphys.2018.00830
- BuskirkERKolliasJAkersRFProkopEKReateguiEPMaximal performance at altitude and on return from altitude in conditioned runnersJ Appl Physiol196723 259 266 1:STN:280:DyaF2s3gvVKqtQ%3D%3D 6033527 https://doi.org/10.1152/jappl.1967.23.2.259 6033527
- Sanz-Quinto S, López-Grueso R, Brizuela G, Flatt AA, Moya-Ramón M. Influence of training models at 3900 m altitude on the physiological response and performance of a professional wheelchair athlete: a case study. J Strength Cond Res. 2019;33:1714–22.
- BoyerSJBlumeFDWeight loss and changes in body composition at high altitudeJ Appl Physiol198457 1580 1585 1:STN:280:DyaL2M7gsVCktA%3D%3D 6520055 https://doi.org/10.1152/jappl.1984.57.5.1580 6520055
- ConsolazioCFMatoushLOJohnsonHLKrzywickiHJIsaacGJWittNFMetabolic aspects of calorie restriction: Hypohydration effects on body weight and blood parametersAm J Clin Nutr196821 793 802 1:STN:280:DyaF1czmtlyqtg%3D%3D 5667919 https://doi.org/10.1093/ajcn/21.8.793 5667919
- KayserBAchesonKDecombazJFernECerretelliPProtein absorption and energy digestibility at high altitudeJ Appl Physiol199273 2425 2431 1:STN:280:DyaK3s7ks12qsg%3D%3D 1490954 https://doi.org/10.1152/jappl.1992.73.6.2425 1490954
- SurksMIChinnKSMatoushLRAlterations in body composition in man after acute exposure to high altitudeJ Appl Physiol196621 1741 1746 1:STN:280:DyaF2s%2FlsFekuw%3D%3D 5929298 https://doi.org/10.1152/jappl.1966.21.6.1741 5929298
- ButterfieldGEGatesJFlemingSBrooksGASuttonJRReevesJTIncreased energy intake minimizes weight loss in men at high altitudeJ Appl Physiol199272 1741 1748 1:STN:280:DyaK383ptV2mtg%3D%3D 1601781 https://doi.org/10.1152/jappl.1992.72.5.1741 1601781
- HoppelerHKleinertESchlegelCClaassenHHowaldHKayarSRCerretelliPMorphological adaptations of human skeletal muscle to chronic hypoxiaInt J Sports Med199011 S3 S9 2323861 https://doi.org/10.1055/s-2007-1024846 2323861
- FulcoCSRockPBCymermanAMaximal and submaximal exercise performance at altitudeAviat Space Environ Med199869 793 801 1:STN:280:DyaK1cznvFyhug%3D%3D 9715971 9715971
- MurdochDRSymptoms of infection and altitude illness among hikers in the Mount Everest region of NepalAviat Space Environ Med199566 148 151 1:STN:280:DyaK2M3ktlyksg%3D%3D 7726779 7726779
- MilledgeJSSalt and water control at altitudeInt J Sports Med199213 S61 S63 1483795 https://doi.org/10.1055/s-2007-1024596 1483795
- CalbetJAChronic hypoxia increases blood pressure and noradrenaline spillover in healthy humansJ Physiol2003551 379 386 1:CAS:528:DC%2BD3sXns1Srs7w%3D 12844510 2343162 https://doi.org/10.1113/jphysiol.2003.045112
- StockMJNorganNGFerro-LuzziAEvansEEffect of altitude on dietary-induced thermogenesis at rest and during light exercise in manJ Appl Physiol197845 345 349 1:STN:280:DyaE1M%2Fis12isw%3D%3D 701118 https://doi.org/10.1152/jappl.1978.45.3.345 701118
- WoodsALGarvican-LewisLARiceAThompsonKG12 days of altitude exposure at 1800 m does not increase resting metabolic rate in elite rowersAppl Physiol Nutr Metab201742 672 676 1:CAS:528:DC%2BC2sXlslCrurs%3D 28278387 https://doi.org/10.1139/apnm-2016-0693 28278387
- GroverRFBasal oxygen uptake of man at high altitudeJ Appl Physiol196318 909 912 1:STN:280:DyaF2c%2FitVyrtQ%3D%3D 14063259 https://doi.org/10.1152/jappl.1963.18.5.909 14063259
- WesterterpKRKayserBBrounsFHerryJPSarisWHEnergy expenditure climbing MtEverest J Appl Physiol199273 1815 1819 1:STN:280:DyaK3s7hsFWgsQ%3D%3D 1474056 https://doi.org/10.1152/jappl.1992.73.5.1815 1474056
- KayserBNariciMVCibellaF SuttonJRHoustonCSCoatesGFatigue and performance at high altitudeHypoxia and molecular medicine1993 Burlington, NJ Queen City press 222 234
- MacDougallJDGreenHJSuttonJRCoatesGCymermanAYoungPHoustonCSOperation Everest II: structural adaptations in skeletal muscle in response to extreme simulated altitudeActa Physiol Scand1991142 421 427 1:STN:280:DyaK38%2FisFalsQ%3D%3D 1927554 https://doi.org/10.1111/j.1748-1716.1991.tb09176.x 1927554
- D’HulstGDeldicqueLHuman skeletal muscle wasting in hypoxia: a matter of hypoxic dose?J Appl Physiol2017122 406 408 27742801 https://doi.org/10.1152/japplphysiol.00264.2016 1:CAS:528:DC%2BC1cXjt1CmsLc%3D 27742801
- Garvican-LewisLASharpeKGoreCJTime for a new metric for hypoxic dose?J Appl Physiol2016121 352 355 26917695 https://doi.org/10.1152/japplphysiol.00579.2015 26917695
- ImoberdorfRGarlickPJMcNurlanMACasellaGAMariniJCTurgayMBärtschPBallmerPESkeletal muscle protein synthesis after active or passive ascent to high altitudeMed Sci Sports Exerc200638 1082 1087 1:CAS:528:DC%2BD28XlsFOmtb4%3D 16775549 https://doi.org/10.1249/01.mss.0000222836.66391.35
- Sheffield-MooreMYeckelCWVolpiEWolfSEMorioBChinkesDLPaddon-JonesDWolfeRRPostexercise protein metabolism in older and younger men following moderate-intensity aerobic exerciseAm J Physiol Endocrinol Metab2004287 E513 E522 1:CAS:528:DC%2BD2cXnvVejurk%3D 15149953 https://doi.org/10.1152/ajpendo.00334.2003
- BigardAXSatabinPLavierPCanonFTaillandierDGuezennecCYEffects of protein supplementation during prolonged exercise at moderate altitude on performance and plasma amino acid patternEur J Appl Physiol Occup Physiol199366 5 10 1:STN:280:DyaK3s7ksF2htA%3D%3D 8425512 https://doi.org/10.1007/BF00863392
- SchneiderMBärtschPCharacteristics of headache and relationship to acute mountain sickness at 4559 metersHigh Alt Med Biol201819 321 328 30067102 https://doi.org/10.1089/ham.2018.0025
- VesterinenVNummelaAHeikuraILaineTHynynenEBotellaJHäkkinenKIndividual endurance training prescription with heart rate variabilityMed Sci Sport Exer201648 1347 1355 https://doi.org/10.1249/MSS.0000000000000910
- CamposGELueckeTJWendelnHKTomaKHagermanFCMurrayTFRaggKERatamessNAKraemerWJStaronRSMuscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zonesEur J Appl Physiol200288 50 60 12436270 https://doi.org/10.1007/s00421-002-0681-6
- Spanish Ministry of Science and Innovation. Base de Datos Española de Composición de Alimentos (BEDCA). http://www.bedca.net/bdpub/index.php. Accessed 20 Jan 2017.
- MartinCKCorreaJBHanHAllenHRRoodJCChampagneCMGunturkBKBrayGAValidity of the remote food photography method (RFPM) for estimating energy and nutrient intake in near real-timeObesity.201220 891 899 1:CAS:528:DC%2BC38XkvVemu70%3D 22134199 https://doi.org/10.1038/oby.2011.344
- BurkeLPractical issues in nutrition for athletesJ Sports Sci199513 S83 S90 8897324 https://doi.org/10.1080/02640419508732281
- MortonRWMcGloryCPhillipsSMNutritional interventions to augment resistance training-induced skeletal muscle hypertrophyFront Physiol20156 245 26388782 4558471 https://doi.org/10.3389/fphys.2015.00245
- DhillonJCraigBALeidyHJAmankwaahAFOsei-Boadi AnguahKJacobsAJonesBLJonesJBKeelerCLMcCroryMARiveraRLSlebodnikMMattesRDTuckerRMThe effects of increased protein intake on fullness: a meta-analysis and its limitationsJ Acad Nutr Diet2016116 968 983 26947338 https://doi.org/10.1016/j.jand.2016.01.003 26947338
- BjorntorpPImportance of fat as a support nutrient for energy: metabolism of athletesJ Sports Sci19919 71 76 1895364 https://doi.org/10.1080/02640419108729867 1895364
- BurkeLMCoxGRCummingsNKDesbrowBGuidelines for daily carbohydrate intakeInt J Sports Med200131 267 299 1:STN:280:DC%2BD3M3hvVyhtw%3D%3D https://doi.org/10.2165/00007256-200131040-00003
- PfeifferBCotterillAGrathwohlDStellingwerffTJeukendrupAEThe effect of carbohydrate gels on gastrointestinal tolerance during a 16-km runInt J Sport Nutr Exerc Metab.200919 485 503 1:CAS:528:DC%2BD1MXhtlCju7vM 19910651 https://doi.org/10.1123/ijsnem.19.5.485 19910651
- JentjensRLMoseleyLWaringRHHardingLKJeukendrupAEOxidation of combined ingestion of glucose and fructose during exerciseJ Appl Physiol200496 1277 1284 1:CAS:528:DC%2BD2cXjtlGiu74%3D 14657042 https://doi.org/10.1152/japplphysiol.00974.2003 14657042
- JeukendrupAECarbohydrate feeding during exerciseEur J Sport Sci20088 77 86 https://doi.org/10.1080/17461390801918971
- BurkeLMHawleyJAWongSHJeukendrupAECarbohydrates for training and competitionJ Sports Sci201129 S17 S27 21660838 https://doi.org/10.1080/02640414.2011.585473 21660838
- NortonLayne EWilsonGabriel JLaymanDonald KMoultonChristopher JGarlickPeter JLeucine content of dietary proteins is a determinant of postprandial skeletal muscle protein synthesis in adult ratsNutrition & Metabolism20129 1 67 1:CAS:528:DC%2BC38XhvVSiu7%2FF https://doi.org/10.1186/1743-7075-9-67
- SnijdersTSmeetsJSvan VlietSvan KranenburgJMaaseKKiesAKVerdijkLBvan LoonLJProtein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young menJ Nutr2015145 1178 1184 1:CAS:528:DC%2BC2MXhtVOgsbjM 25926415 https://doi.org/10.3945/jn.114.208371 25926415
- Stray-GundersenJMordecaiNLevineBDO2 transport response to altitude training in runnersMed Sci Sports Exerc199527 S202 https://doi.org/10.1249/00005768-199505001-01131
- Garvican-LewisLAVuongVLGovusADPeelingPJungGNemethEHughesDLovellGEichnerDGoreCJIntravenous iron does not augment the hemoglobin mass response to simulated hypoxiaMed Sci Sports Exerc201850 1669 1678 1:CAS:528:DC%2BC1cXhtlWrtrnJ 29538179 https://doi.org/10.1249/MSS.0000000000001608 29538179
- CummingGFinchSA primer on the understanding, use, and calculation of confidence intervals that are based on central and noncentral distributionsEduc Psychol Meas200161 530 572 https://doi.org/10.1177/0013164401614002
- CohenJA power primerPsychol Bull1992112 155 159 1:STN:280:DC%2BD1MvlsV2mug%3D%3D 19565683 https://doi.org/10.1037/0033-2909.112.1.155 19565683
- PasiakosSMBerrymanCECarriganCTYoungAJCarboneJWMuscle protein turnover and the molecular regulation of muscle mass during hypoxiaMed Sci Sports Exerc201749 1340 1350 1:CAS:528:DC%2BC2sXhtVSqurrJ 28166119 https://doi.org/10.1249/MSS.0000000000001228 28166119
- HansenJSanderMSympathetic neural overactivity in healthy humans after prolonged exposure to hypobaric hypoxiaJ Physiol2003546 921 929 1:CAS:528:DC%2BD3sXitlSmt7w%3D 12563015 https://doi.org/10.1113/jphysiol.2002.031765 12563015