
Abstract
miRNAs are short, non-coding, regulatory RNAs that exert cell type-dependent, context-dependent, transcriptome-wide gene expression control under physiological and pathological conditions. Tissue slide-based assays provide qualitative (tumor compartment) and semi-quantitative (expression levels) information about altered miRNA expression at single-cell resolution in clinical tumor specimens. Reviewed here are key technological advances in the last 5 years that have led to implementation of fully automated, robust and reproducible tissue slide-based assays for in situ miRNA detection on US FDA-approved instruments; recent tissue slide-based discovery studies that suggest potential clinical applications of specific miRNAs in cancer medicine are highlighted; and the challenges in bringing tissue slide-based miRNA assays into the clinic are discussed, including clinical validation, biomarker performance, biomarker space and integration with other biomarkers.
MicroRNAs (miRNAs) are an evolutionarily conserved class of short, non-coding, regulatory RNAs that generally control gene expression at the post-transcriptional level Citation[1–5]. A single miRNA can interact with hundreds of target messenger RNAs (mRNAs) and affect the protein output of a large number of target genes at once Citation[6–8]. In some instances, specific miRNAs can coordinately regulate the expression of multiple protein components of the same signaling pathway or biological process. Thus, deregulation of miRNA activity can have broad and deleterious consequences for cell homeostasis and can contribute to human disease Citation[9–14]. Over the last decade, rapid technological advances in platforms for high-throughput miRNA profiling in tissue and blood samples have generated lists and signatures of miRNAs associated with various cancer types, neurodegenerative conditions and cardiovascular disease Citation[9,10,15,16]. Several quantitative real-time PCR-based assays have been commercialized as laboratory-developed tests (LDTs) for use in tissue samples to differentially diagnose pancreatic ductal adenocarcinoma, to diagnose the subtypes of kidney and lung cancer, and to identify the organ site of cancers of unknown primary origin Citation[17–23]; a handful of these tests are already being reimbursed for by health insurance companies Citation[24,25]. Some of the disease-associated miRNAs have also been functionally characterized in preclinical models and some front-runner compounds are already being evaluated in clinical trials for anti-viral and oncology applications Citation[26,27].
Recently, more attention has been paid to the cell types in which miRNA expression is altered in tumor tissues. Tumors are complex quasi-organs that contain cancer cells and several other supportive and reactive cell types in the tumor microenvironment. Intra- and inter-tumoral heterogeneity at the molecular and cellular levels Citation[28–37] can confound and mislead the interpretation of miRNA expression analyses that use RNA extracted from tumor tissue or blood Citation[15,16,38,39]. Over the last 5 years, various groups have developed robust and reproducible in situ hybridization (ISH) methods to visualize miRNA expression within individual cells in formalin-fixed paraffin-embedded clinical tissue specimens, because of the clinical implications of the cell type (tumor compartment) that harbors altered miRNA expression.
This review focuses on the implementation of tissue slide-based assays for miRNA ISH detection in cancer and the implications for cancer medicine. It provides a brief historical perspective of the origins of these tissue slide-based assays and describes technological advances that rapidly led from manual single-marker assays to fully automated multiplex assays for the co-detection of miRNA and protein markers. These technological advances are intimately intertwined with the potential clinical applications of miRNA biomarkers. These tissue slide-based assays provide powerful investigational tools to better understand the etiological relevance of miRNAs and, eventually, could provide diagnostic tools to guide treatment decisions. The review highlights recent tissue slide-based discovery studies that assess the diagnostic and prognostic value of miRNAs in common cancer types including breast, colon, lung, pancreas and skin. The final topic deals with considerations and challenges to further implement lead miRNA indicators into useful biomarkers that deliver solutions for unmet clinical needs in cancer medicine.
High-throughput expression profiling of miRNAs in cancer tissues
The first link between aberrant miRNA expression and function in human cancer was found in 2002, when chromosomal deletion of the mir-15/mir-16 locus and/or altered expression of miR-15/miR-16 were associated with chronic lymphocytic leukemia Citation[40]. Subsequently, miR-15/miR-16 were functionally linked to disease etiology via regulation of a key target gene, BCL2 Citation[41]. Soon after, various groups developed different high-throughput detection platforms, including bead-based hybridization and microarray chips, to profile the expression of known miRNAs in panels of different hematologic and solid cancer types Citation[42–51]. Over the years, improvements in these platforms as well as newer technologies such as multiplex RT-PCR plates, RNA-sequencing and Nanostring have enabled an in-depth characterization of microRNome changes in many cancer types Citation[52–55].
Validation of miRNA expression profiling signatures
Technical validation of differentially expressed miRNAs identified via high-throughput platforms typically consists of single miRNA detection by a stem-loop RT-PCR assay Citation[56] or a similar approach. While there are technical discrepancies and different biases of each detection method Citation[54,55], miRNA expression signatures have been technically validated in multiple studies. However, none of these detection methods enable us to identify cellular source(s) of altered miRNA expression. Most studies have used RNA extracted from whole tumor tissues or normal reference controls. It has often been assumed that altered miRNA expression in whole-tumor tissue biopsies or in blood reflects molecular changes within cancer cells. Accordingly, functional studies have been carried out in cancer cell lines to understand the molecular mechanism(s) behind deregulated miRNA expression, which may lead to new therapeutic opportunities Citation[13,14]. A predominant deregulation of miRNA activity in cell types other than cancer cells would put into question the interpretation of functional assays with cancer cell lines and would force us to reconsider the etiological contribution of particular miRNAs to carcinogenesis. Thus, the confirmation of altered miRNA expression within individual cells in the native context of a complex tumor lesion is required to unequivocally assign miRNA dysregulation to cancer cells or other cells of the tumor microenvironment.
Some studies have enriched tissue samples for specific cellular content (cancer cells vs stroma) using laser capture microdissetion Citation[57–60], a tissue core punch extractor Citation[61] or tissue macrodissection Citation[62] before miRNA detection with high-throughput and sensitive methods. However, these approaches for enrichment are labor-intensive and technically demanding, but still do not enable miRNA detection within individual cells. ISH is an ideal assay for validating miRNA expression at single-cell resolution using a limited amount of a clinical tissue specimen. Moreover, miRNA detection by ISH follows a procedure very similar to tissue slide-based morphology-driven clinical assays for protein detection by immunohistochemistry (IHC) and DNA detection by fluorescence ISH. The principles and clinical applications of miRNA detection by ISH are presented in following sections.
Tissue slide-base assays for in situ miRNA detection
In situ detection of miRNA expression was first achieved in vertebrate embryos using whole-mount ISH protocols Citation[63,64]. The key innovation was the incorporation of locked nucleic acid (LNA)-modified antisense probes. LNAs are high-affinity RNA analogs that have unprecedentedly strong and specific binding properties Citation[65,66] to complementary short RNA sequences such as those of mature miRNAs. Whole-mount ISH results confirmed expression profiling findings of mammalian organ- and cell-type-specific miRNA expression patterns Citation[67,68]. Soon after that key innovation, various groups implemented chromogenic and fluorescent-based ISH assays using LNA-modified probes to detect miRNA expression in frozen and fresh clinical tumor specimens to validate expression profiling signatures Citation[51,69–74]. Nelson et al. Citation[51] used single digoxigenin-tagged LNA-modified probes against miRNAs (miR-9, miR-124, miR-125b) differentially expressed in brain cancer; the probes were detected using anti-digoxigenin antibody conjugated to alkaline phosphatase and chromogenic development of the 4-nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate substrate. Sempere et al. Citation[69] used single fluorescein (fluorescein isothiocyanate [FITC])-tagged LNA-modified probes against miRNAs (let-7a, miR-21, miR-145, miR-125b miR-205, miR-451) in breast cancer; the probes were detected with anti-FITC antibody conjugated to horseradish peroxidase (HRP) and deposition of reactive tyramide substrate conjugated to FITC. Both studies demonstrated that altered miRNA expression in tumors was complex and involved distinct normal cell types and tumor compartments for different miRNAs; some expression changes suggested an etiological contribution of specific miRNAs, whereas others did not. These early studies set the basis for a workable assay.
Methodological implementation & protocol variations
Over the last 5 years, several groups have considerably improved these chromogenic and fluorescent-based ISH assays for miRNA detection in formalin-fixed paraffin-embedded tissue specimens . The limit of detection of the ISH assay is low compared with exponential amplification methods such as quantitative real-time PCR; thus, detection of miRNAs expressed at low copy number is technically difficult. The author describes below general strategies and variants that have been implemented for optimal in situ detection of miRNA expression . The author highlights some critical parameters; more thorough technical details and general troubleshooting tips have been presented elsewhere Citation[75–79].
Table 1. Variations and evolution of tissue slide-based assays for in situ detection of miRNAs†.
Preanalytical sample preparation
Tissue fixation in formalin for less than 24 h or more than 72 h before for paraffin embedding and other sample preparation steps such as time from operating room to tissue processing can affect miRNA detection Citation[38,80]. These can be confounding factors when analyzing miRNA expression from tissue specimens collected in different institutions and at different times, especially in historical archives dating back to the 1980s and 90s, before standardization of tissue processing procedures. A post-fixation step with 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) Citation[79,81] can be also used to increase the retention of miRNA molecules. However, EDC fixation may limit the multiplex capability of miRNA and protein co-detection because EDC cross-links can cause epitope masking Citation[79].
Probe design
LNA-modified nucleotides are typically incorporated every 2–3 nucleotides to increase the melting temperature (Tm) by 10–15°C. Ideally, the Tm for a probe should be about 75°C, but Tm prediction formulae are not always accurate. Empirical determination of Tm for optimal probe detection includes variables such as probe concentration, ionic strength and formamide concentration in the hybridization solution, time and temperature of hybridization and temperature and stringency of saline sodium citrate washes. Reticular nuclear and/or nucleolar stains typically indicate spurious hybridization of the probe against ribosomal or other abundant RNAs. Thus, it is important to have adequate control tissues and/or reference cell lines to assess the specificity of probe binding.
Probe tags
The addition of multiple hapten tags – most commonly two terminal tags (e.g., DIG2X, FITC2X) Citation[75–78,82–98], but at times up to six (FITC6X, Bio2X) Citation[81] – increases probe detection.
Probe access to fixed miRNA
Enzymatic pretreatment with proteinase K is commonly used to digest tissue and break down protein–RNA crosslinks to facilitate probe penetration and miRNA exposure. Changes in time, temperature and concentration of proteinase K between samples (intra-experimental) and between experiments (inter-experimental) affect stains by failing to sufficiently open up the tissue or by causing too much tissue damage Citation[38,78]. Heat-induced retrieval with citrate buffer can be used Citation[99,100] instead of proteinase K. This pretreatment may be beneficial for co-detection of miRNAs and proteins if concerns exist that proteinase K may destroy protein epitopes. However, various groups have reported multiplex co-detection of miRNAs and a score of proteins after proteinase K pretreatment Citation[75,76,80,92–96].
Detection methods
Chromogenic or fluorescent-based stains have been used to detect miRNA probes. Chromogenic staining can be achieved by alkaline phosphatase-mediated development of 4-nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate substrate and/or HRP-mediated development of 3,3′-diaminobenzidine substrate. Fluorescent staining can be achieved by HRP-mediated deposition of reactive substrates conjugated to a variety of fluorochromes (FITC, rhodamine, Dyglight dye series, Alexa dye series, Cy dye series). The use of antibody sandwich amplification (e.g., anti-tag primary antibody followed by secondary antibody-conjugated to HRP) Citation[38,75,76] and/or enzymatic polymers (e.g., poly-HRP) Citation[76,80] increases the number of functional units (e.g., HRP units) per bound probe molecule and thereby increases label deposition. The use of fluorescent stains (enzyme-reactive substrates conjugated to fluorochromes) can increase signal sensitivity and dynamic range of detection and can also enable a variety of multiplexing applications for high-content marker analysis Citation[16,81,93–95,101].
Several research groups have already implemented these assays in fully automated staining stations Citation[76,80,97,98], some of which are US FDA-approved for diagnostic IHC assays. Automation enables precise and consistent performance of time-sensitive steps such as proteinase K pretreatment and substrate incubation for signal development. These robust and reproducible ISH assays now meet technical standards of, use same instrumentation as and are compatible with diagnostic IHC assays.
High-content image analysis
The scoring of miRNA expression takes into account the tumor compartment and the intensity of the stain. Some studies rely on visual examination by independent viewers, who report intensity of staining on a 0–3 scale and score cases as low or high, noting the tumor compartment (cancer vs stroma). Some studies have used matched normals as a reference for intensity changes in the tumor. This reporting system is very similar to what is used for protein stains in the clinic. As with protein stains, it is assumed that tissues are handled, processed and preserved under identical conditions, so that any difference in stain intensity reflects expression changes. This may not be always the case. Some studies have used expression of small nuclear RNA U6 or ribosomal RNA 18S as a control for RNA quality and integrity Citation[38,95,101,102]. However, the expression of these control RNA markers varies depending of disease stage. A universal RNA control for miRNA normalization in tissue slide-based assays or other detection methods has not yet been established. Notwithstanding this limitation, various groups have demonstrated that ISH-based stainings are reproducible and semi-quantitative Citation[95,97,101,103]. Direct comparison on the same tissue samples indicate that ISH assays can discriminate a similar range of fold changes in expression as RT-PCR assays Citation[97,101,103].
To further standardize and objectify staining scores, several groups have begun to employ computer-assisted image analysis tools Citation[38,76,95,97,98,101–105]. For example, fluorescent staining of let-7a and miR-205 in the cancer cell compartment of breast cancer tissue core sections was analyzed with an Ariol automated microscopic image capture system Citation[103]. Chromogenic staining of miR-21 in the stromal compartment of colon cancer whole-tissue sections was analyzed with a Visiopharm integrated microscope and software module; morphological characteristics indicated that miR-21 was predominantly expressed in tumor-associated fibroblasts Citation[97]. Chromogenic staining of miR-126 in blood vessels of lung cancer whole-tissue sections was also analyzed with the Visiopharm system Citation[104]. In addition to morphological characteristics, co-stains with cell type-specific proteins in multiplex assays can more unambiguously identify individual or a small nest of cancer cells (e.g., cytokeratin markers for carcinomas), tumor-associated fibroblasts (e.g., smooth muscle actin), endothelial cells (e.g., CD31) and leukocyte lineages and subpopulations (e.g., CD3 for T-cell lymphocytes, CD11b for myeloid cells, CD68 for macrophages) Citation[38,76]. Fluorescent staining of miR-221 in the cancer cell compartment of breast cancer tissue core sections was analyzed with Automated Quantitative Analysis technology; co-staining with pan-cytokeratin marker was used to define cancer cell compartment Citation[101]. Fluorescent staining of miR-21 in the stromal compartment of whole-tissue sections of breast and colon cancer cases was analyzed with Image-Pro Plus; co-staining with vimentin and smooth muscle actin markers confirmed that miR-21 was predominantly expressed in tumor-associated fibroblasts Citation[38].
Discovery studies in cancer using tissue slide-based assays for in situ miRNA detection
Recent tissue slide-based discovery studies suggest potential clinical applications of specific miRNAs in variety of cancers, including breast, colon, lung, pancreas and skin . Most of the studied miRNAs had been identified as cancer-associated by high-throughput expression profiling analyses. These tissue slide-based studies for the first time evaluated the effect of the tumor compartment on the diagnostic and prognostic value of altered miRNA expression. In one such study, the prognostic value of miR-150 was assessed in colon cancer by RT-PCR and ISH assays Citation[106]. Although expression analysis was determined in two independent patient cohorts, it is worth noting that the ISH-based miR-150 score was more powerful in stratifying patients who had better clinical outcomes Citation[106]. The author highlights below the main findings of some other tissue-slide studies that reinforce the importance of contextual miRNA expression analysis.
Table 2. Potential clinical applications of miRNA-based in situ hybridization assays†.
Altered miRNA expression in cancer cells
High let-7g and miR-221 expression and low expression of miR-27 in the cancer cell compartment of breast cancer cases are associated with better clinical outcome Citation[101,107,108]. miR-205 is expressed at high levels in myoepithelial and basal epithelial cells in normal breast and skin tissue, respectively; retention of miR-205 expression in cancer cells is associated with better clinical outcomes in both breast cancer and melanoma cases Citation[69,102,103]. High miR-182 expression and low miR-126 and miR-146 expression in the cancer cell compartment are associated with better outcomes in lung cancer cases Citation[83,84,87]. The cancer cell expression of miR-126 is intriguing, because miR-126 is a well-characterized miRNA for its role in endothelial cell biology and has been shown to have expression confined to endothelial cells Citation[38,109–115]. Future studies are needed to validate the prognostic value of miR-126 expression in lung cancer cells.
Altered miRNA expression in stromal cells
miR-21 is expressed in cancer cells and stromal cells (mainly tumor-associated fibroblasts) in several solid cancers Citation[38,69–71,90,91,97,98,116]. In colon and pancreas cancer, most cases predominantly express miR-21 in the stroma, and this is the only tumor compartment with prognostic value Citation[90,97,98]. In vitro co-culture studies of cancer cells and tumor-associated fibroblasts isolated from colon and pancreatic cancer patients have revealed fibrogenic roles of miR-21, suggesting a functional etiological contribution of miR-21 to support cancer cell growth and enhance malignancy Citation[90,117]. In lung cancer, it appears that high expression of miR-21 in stromal cells is a negative prognostic indicator, whereas high expression in the cancer cells of node-positive cases is protective Citation[85]. In contrast, high expression of miR-21 in cancer cells and tumor-associated blood vessels in glioma cases are associated with poor clinical outcome Citation[105]. The difference between expression patterns of miR-21 in carcinomas and gliomas may reflect the different idiosyncrasies of cancers of epithelial and neural origin, respectively. In an interesting study, high levels of miR-126 in endothelial cells in primary tumor tissues were associated with treatment response to first-line capecitabine and oxaliplatin chemotherapy and with better clinical outcome in advanced colorectal cancer cases with metastatic disease Citation[104]. On a separate study, expression of hypoxia-induced miR-210 was detected in cancer and stromal cells in lung cancer cases, but only high levels of miR-210 in stromal cells were significantly associated with a better outcome Citation[118].
Study design limitations
These tissue slide-based discovery studies reported statistically significant prognostic indications for compartment-specific altered miRNA expression in multivariate analyses that adjusted for standard clinicopathologic prognostic factors . These studies suggest that prognostic information obtained with miRNA expression is independent of standard prognostic factors. However, the sample size and/or limited molecular characterization of tumor specimens precluded a more in-depth evaluation of specific miRNAs in the context of known molecular biomarkers that impact clinical decisions (e.g., estrogen receptor [ER]/progesterone receptor [PR]/human epidermal growth factor-like receptor 2 [HER2] status in breast cancer; K-Ras and EGFR mutation status in colon and lung cancer). Nonetheless, a handful of adequately powered studies with larger case sets (n > 500) have started to find links to specific molecular alterations. In breast cancer, the status of the ER, PR and HER2 are used for diagnostic, prognostic and treatment decisions. Within major subtypes defined by the combined status of ER/PR/HER2, patients still exhibit heterogeneous responses to treatment and clinical outcome Citation[119]. Low expression of tumor suppressive let-7b in cancer cells from ER+ and/or PR+ (with or without HER2+) cases was associated with a better clinical outcome Citation[103]. High expression of oncogenic miR-21 in cancer cells from ER+ and/or PR+ HER2– cases and in tumor-associated fibroblasts from ER–, PR–, HER2– cases was associated with a poorer clinical outcome [Mackenzie TA, Schwartz G, Calderone H, et al. Stromal Expression Of miR-21 Identifies High-Risk Group In Triple Negative Breast Cancer, Submitted]. Although more studies and clinical validation of let-7b and miR-21 indications are necessary, these early works suggest ways in which miRNA biomarkers could complement current clinically used protein markers to improve prognostic assessment and to guide treatment decisions .
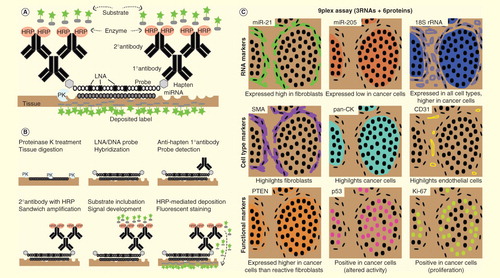
Figure 1. Automated platform for high-content marker analysis of tumor tissues. (A) Key components of in situ miRNA detection assay. (B) Purposes of key components and the steps they take part in, from tissue digestion to covalently linked deposition of fluorescent labels. (C) Virtual staining rendition of a 9plex assay that co-detects three RNA markers (two miRNAs + control rRNA) and six protein markers (three cell type-specific + three functional/prognostic markers); black dots indicate nuclei revealed by counterstaining with DAPI. Top left, representative miR-21 stain (green) on an ER−PR−HER2− breast cancer case in the first round of HRP-mediated deposition of reactive substrate conjugated with a fluorochrome. Sequential rounds of HRP-mediated deposition of substrates conjugated with spectrally unique fluorochromes are carried out to detect probes against other RNA markers with appropriate anti-hapten primary and secondary antibody combinations or protein markers with appropriate primary and secondary antibody combinations. Each new staining round requires a peroxide block step to inactive HRP from the preceding round.
Expert commentary
Cancer cell clonal diversity, tissue heterogeneity and the tumor microenvironment influence the efficacy of current treatments of solid tumors. The detailed cellular and molecular characterization of tumors presents a unique opportunity to translate scientific knowledge into actionable clinical information. Over the last 5 years, several groups have overcome technical challenges of detecting miRNA using ISH assays in fixed tumor tissues. These tissue slide-based assays enable the characterization of miRNA expression in individual cells in the tumor microenvironment and the determination of which tumor compartment(s) harbors this altered expression. This type of assay enhances the precision and accuracy of informative miRNA biomarkers for which context and morphological features are important. However, the use of tissue slide-based miRNAs in clinical application is still in an early stage of development relative to RT-PCR-based assays, which have already entered the clinic as LDTs. Thus, it is important to consider the advantages and limitations of each platform from the technical, instrumental and diagnostic vantage points Citation[15,16], so as to better focus efforts on the development of tissue slide-based assays that can uniquely shed light on knowledge gaps and address unmet clinical needs in cancer medicine.
Tissue analysis using tissue slide-based assays provides a direct evaluation of disease state based on miRNA expression (or other biomarkers) for diagnostic and prognostic purposes, but it is not generally an appropriate biological material for early disease detection or disease monitoring. For such applications, RT-PCR-based miRNA detection in blood (serum, plasma, cellular component, exosome fraction) or other biological fluids, which can be collected by non-invasive means at frequent intervals, is being actively pursued by many research laboratory and diagnostic companies Citation[120].
Tissue slide-based assays can be more cost–effective in screening the expression of selected miRNAs (n < 10) in a large number of tissue samples (n > 500) using tissue microarrays, whereas RT-PCR assays provide high throughput in terms of the number of independent miRNAs (n > 100) that can be detected in a single tube reaction.
Tissue slide-based assays can more precisely and more readily provide contextual information for abundantly expressed miRNAs, whereas RT-PCR assays are more sensitive and can be used to detect all known miRNAs.
Tissue slide-based assays enable co-detection of miRNAs and other bioanalytes (DNA, mRNA, lncRNAs, rRNAs, snRNAs, tRNAs) in their native tissue, cellular and subcellular context on the same tissue section. Integrative approaches and multi-marker contextual signatures could considerably enhance the diagnostic power of individual components.
Tissue slide-based assays enable co-detection of miRNAs and their cognate target mRNAs or target gene products (protein). These assays can be used to validate miRNA and target gene interactions at the mRNA (multiplex ISH) or protein (combined ISH/IHCplex) levels and to assess whether miRNA regulatory networks are engaged and functional in individual tumor cases.
For some specific questions and applications, tissue slide-based assays could outperform RT-PCR assays. This point was illustrated in previous section by the cancer cell expression of miR-150 Citation[106], the endothelial cell expression of miR-126 in colon cancer Citation[104] and the stromal cell expression of miR-21 in breast, colon and pancreatic cancer Citation[90,91,97] [Mackenzie TA, Schwartz G, Calderone H, et al. Stromal Expression Of miR-21 Identifies High-Risk Group In Triple Negative Breast Cancer, Submitted]. Future tissue slide-based discovery and validation studies may identify a clinical value of miRNAs that did not pass quality or statistical filters in previous expression profiling studies using mixed RNA pools from whole tissue samples. In addition, there may be viable business development opportunities to build on and eventually replace RT-PCR assays. The Asuragen miRInform® PANCREAS is a multiplex RT-PCR assay that measure levels of two miRNAs to improve the detection and diagnosis of pancreatic ductal adenocarcinoma Citation[24]. This and other LDTs, including the Rosetta lung cancer test Citation[25], use fewer than 10 miRNAs to establish informative test results. Thus, these tests would be amenable to adaptation for tissue slide-based assays.
For other clinical applications, in which a miRNA signature of 10 or more is required to establish informative test results, RT-PCR-based assays are a more appropriate platform. The Rosetta cancer origin test is an example of such a multiplex RT-PCR-based assay, which measures expression of a large miRNA signature (64 miRNAs) to identify likely primary tumor types using primary or metastatic tumors Citation[25]. This test may be particularly helpful for cancers having an unknown primary, as correct diagnosis would allow treatment using the standard of care for that particular cancer type. Currently, IHC assays with a battery of cell type-specific markers are needed to identify or confirm origin (after the Rosetta cancer origin test). ISH detection of informative miRNAs by the Rosetta cancer origin test could be used to analytically confirm test results before or in conjunction with IHC markers. Together, these RT-PCR-based LDTs offer successful examples that could guide a business development plan for tissue slide-based miRNA diagnostics and inform regulatory procedures required by FDA and other regulatory agencies.
Clinical implications of tissue slide-based assays
Besides the potential of tissue slide-based assays to be more informative than RT-PCR assays, a question with broader implications is whether miRNAs can be more informative, advantageous or versatile than other biomarkers for tissue analysis. RT-PCR-based detection of miR-205 for differential diagnosis of non-small-cell lung cancer adenocarcinomas versus squamous cell carcinomas is a case in point Citation[121–125]. Would ISH miR-205 detection provide any additional value to the current multi-protein panels (e.g., TTF-1, napsin A, p63, cytokeratin 5/6) Citation[126,127] in making this determination? While it seems unlikely that miR-205 could displace such well-established panels, miR-205 may also have prognostic value in squamous cell carcinomas Citation[128]; thus, a combined ISH/IHC of this miRNA and protein could expand current application of individual markers. In general, it would be important to consider the availability and performance of known biomarkers to identify areas in which miRNAs can be particularly useful. For example, it would seem impractical to use miRNA expression to obtain information similar to the well-established ER/PR/HER2 status in breast cancer, but miRNA expression could be of use in cancer cells or other tumor compartments to refine the characterization and prognostic indications based on ER/PR/HER2 status. When useful IHC markers are not available, developing a miRNA biomarker program could have unique advantages. Since specific probes and other reagents to detect miRNA expression can be readily synthesized based on Watson–Crick sequence complementarity rules, tissue slide-based assays of lead miRNA biomarkers could be expeditiously implemented for disease assessment, such as cancer cell-expression of miR-34 for recurrence risk stratification in bladder cancer [Andrew AS, Cowper R, Marsit CJ, et al. Expression of Tumor Suppressive microRNA-34a Is Associated With A Reduced Risk Of Bladder Cancer Recurrence, Submitted].
Five-year view
Automated pipelines exist for whole-slide image acquisition and computer-assisted image analysis Citation[129], as mentioned above. Some of these platforms integrate multispectral analysis capabilities which could enable detection of as many as 10–12 markers in a single tissue slide using spectrally distinct fluorochromes. As the field of digital pathology and associated quantitative image analysis tools mature Citation[129,130], it is foreseeable that these technologies could be applied for high-content analysis of miRNA expression in translational pathology research laboratories as well as in clinical laboratories. High-content multi-panel assays would contain miRNAs, reference RNAs and cell type-specific and prognostic protein markers to maximize contextual information per tissue slide for diagnostic and prognostic applications. A contextual signature of 20–36 miRNA and protein markers (containing up to 9 miRNAs) could be generated using 2–3 consecutive tissue sections, which would be in a similar range of marker numbers as commercially available prognostic gene expression signatures (e.g., Oncotype Dx, 21 mRNAs; PAM50/Prosigna, 50 mRNAs) Citation[131]. The ancillary protein markers would refine cell identity and the complementary prognostic markers would dissect the molecular status of miRNA-expressing cells in cancer cells, reactive stromal elements such as tumor-associated fibroblasts and immune cells such as T cells and macrophages . Whole-slide image analysis would also enable a deeper understanding of tumor tissue composition and heterogeneity, namely, how local and regional distribution in response to hypoxia, inflammation and pH changes may affect cells at the molecular level and in cell–cell interactions. Thus, it could be technologically feasible to integrate ISH/IHC multiplex assays for high-content morphology-driven analysis of informative miRNA and protein biomarkers in clinical settings .
While miRNA biomarkers continue to generate great interest and hold great potential, they need to be subject to same scrutiny and achieve the same benchmark performance as other bioanalytes for clinical implementation Citation[132–134]. So far altered miRNA expression has an exploratory biomarker status. For most tissue slide-based in situ miRNA detection, analytical and clinical validation is still required. A hurdle for clinical validation is to have access to high-quality tissue specimens from large cohorts of patients with well-annotated clinicopathologic features and outcome data. National programs exist, such as the Cancer Therapy Evaluation Program Citation[135] from the National Cancer Institute in the USA, which provide valuable biospecimen resources for retrospective and prospective miRNA biomarker studies. A large number of tissue specimens could be analyzed, first in screening tissue microarrays to cover the entire patient cohorts in an expedited and economic study and then in follow-up analysis with individual whole tissue sections for miRNAs that warrant further evaluation as diagnostic, prognostic or predictive biomarkers.
Purely prognostic miRNA biomarkers may inform tumor biology and provide lead candidates for therapeutic intervention in the translational cancer research setting. However, their immediate impact on clinical decisions may be limited as they would not inform specific course of treatment, other than perhaps suggest a more aggressive intervention plan. Predictive miRNA biomarkers that indicate a specific treatment approach or selection may be more desirable as it may have a more direct impact on clinical decisions. Further development as predictive or integrated biomarkers for specific treatment recommendation in tissue specimens collected during multi-arm clinical trials evaluating different treatment combinations is now possible thanks to Cancer Therapy Evaluation Program and similar national programs. For example, tissue specimens from several breast cancer clinical trials are available for researchers to evaluate the predictive value of compartment-specific miR-21 expression in specific subtype breast cancer cases treated with different combination of hormonal and/or chemotherapy. In this context, the predictive value of miRNA would not likely reflect a direct effect of the drug treatment on miRNA activity as these drugs do not target the miRNA. In contrast, expression analysis of a miRNA that is the intended target of a drug could serve as an integral predictive biomarker in miRNA-based therapy trials. miR-34-based restoration therapy using a synthetic compound that mimics pre-miR-34 is in a Phase I clinical trial for cancer patients with liver involvement, either primary or metastatic tumors Citation[26]. Restoration therapies of tumor suppressive let-7 Citation[136,137] and immunomodulatory miR-155 activities Citation[138] as well as inhibition therapies of tumor promoting and fibrogenic miR-21 activities Citation[90,117,139,140] have been recently proposed and may be clinically evaluated in the near future Citation[141]. Tissue slide-based determination of expression levels of targeted miRNA, which closely correlate with miRNA activity, would provide personalized and precise selection of patients who would more likely benefit from miRNA-based therapeutic interventions.
In conclusion, the technology and resources are now available to generate high-content contextual signatures that integrate miRNA biomarker information, but is the biology relevant and robust enough to make a clinical impact? Clinical validation of lead miRNA biomarkers will be the first step toward clinical implementation and commercialization of tissue slide-based miRNA detection assays.
Key issues
The use of altered miRNAs as new biomarkers for diagnostic, prognostic and therapeutic applications in cancer medicine is a rapidly growing field. Various detection methods exist to measure changes of miRNA levels.
Tissue slide-based assays for in situ miRNA detection on tumor tissues offer unique advantages over other methods of determining miRNA expression in individual cells (tumor compartment) and of registering the location and distribution (intratumoral heterogeneity) of the miRNA-expressing cells.
Tissue slide-based assays characterize miRNA expression in its intact and native tumor context, in contrast to free miRNA in solution that is used for quantitative real-time PCR or other detection methods.
Tissue slide-based assays can provide more precise and accurate diagnostic information because they provide qualitative and semiquantitative information of altered miRNA expression at single-cell resolution.
Recent tissue slide-based discovery studies suggest the diagnostic and prognostic potential of altered miRNAs in specific tumor compartments. Cancer cells are not the only tumor compartment in which altered miRNA expression can be informative. Indeed, altered expression of miR-21 in the stromal compartment (mainly tumor-associated fibroblasts) provides prognostic information in breast, colon and pancreas cancer.
These tissue slide-based in situ hybridization assays are compatible and can be combined with standard clinical immunohistochemistry assays. miRNA and protein diagnostic information can be integrated in a single tissue section.
Recent advances in multispectral analysis and digital pathology enable whole-slide high-content image analysis. Multiplexing of up to 12 markers is feasible using work flow that is comparable to the routine/standard procedures in translational pathology research and clinical laboratories.
Clinical validation of lead miRNA biomarkers in large prospective and longitudinal studies is required to ignite the interest of clinicians to consider using miRNA in their practice and of industry partners in developing business plans for miRNA-based clinical tools.
Acknowledgements
The author thanks H Calderone, G Hostetter and J Westerhuis for critical reading and suggestions during preparation of this manuscript, and K Habermehl and D Nadziejka for technical editing.
Financial & competing interests disclosure
The author has no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending or royalties.
No writing assistance was utilized in the production of this manuscript.
Notes
Related Research Data
References
- Lee RC, Ambros V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001;294:862-4
- Lagos-Quintana M, Rauhut R, Lendeckel W, et al. Identification of novel genes coding for small expressed RNAs. Science 2001;294:853-8
- Lau NC, Lim LP, Weinstein EG, et al. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 2001;294:858-62
- Sempere LF, Cole CN, McPeek MA, et al. The Phylogenetic Distribution of Metazoan microRNAs: insights into Evolutionary Complexity and Constraint. J Exp Zool B Mol Dev Evol 2006;306:575-88
- Bartel DP, Chen CZ. Micromanagers of gene expression: the potentially widespread influence of metazoan microRNAs. Nat Rev Genet 2004;5:396-400
- Selbach M, Schwanhausser B, Thierfelder N, et al. Widespread changes in protein synthesis induced by microRNAs. Nature 2008;455:58-63
- Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009;136:215-33
- Baek D, Villen J, Shin C, et al. The impact of microRNAs on protein output. Nature 2008;455:64-71
- Mendell JT, Olson EN. MicroRNAs in stress signaling and human disease. Cell 2012;148:1172-87
- Sempere LF, Kauppinen S. Translational implications of MicroRNAs in clinical diagnostics and therapeutics. In: . and Bradshaw RADennis EA, editors. Handbook of cell signaling. 2nd volume. Academic Press; Oxford, UK: 2009. p. 2965-81
- Ventura A, Jacks T. MicroRNAs and cancer: short RNAs go a long way. Cell 2009;136:586-91
- Garzon R, Marcucci G, Croce CM. Targeting microRNAs in cancer: rationale, strategies and challenges. Nat Rev Drug Discov 2010;9:775-89
- Fabbri M, Calore F, Paone A, et al. Epigenetic regulation of miRNAs in cancer. Adv Exp Med Biol 2013;754:137-48
- Nana-Sinkam SP, Croce CM. Clinical applications for microRNAs in cancer. Clin Pharmacol Ther 2013;93:98-104
- Zhang X, Lu X, Lopez-Berestein G, et al. In situ hybridization-based detection of microRNAs in human diseases. microRNA. Diagn Ther 2013;1:12-23
- Sempere LF. Integrating contextual miRNA and protein signatures for diagnostic and treatment decisions in cancer. Expert Rev Mol Diagn 2011;11:813-27
- Spector Y, Fridman E, Rosenwald S, et al. Development and validation of a microRNA-based diagnostic assay for classification of renal cell carcinomas. Mol Oncol 2013;7:732-8
- Gilad S, Lithwick-Yanai G, Barshack I, et al. Classification of the four main types of lung cancer using a microRNA-based diagnostic assay. J Mol Diagn 2012;14:510-17
- Meiri E, Mueller WC, Rosenwald S, et al. A second-generation microRNA-based assay for diagnosing tumor tissue origin. Oncologist 2012;17:801-12
- Varadhachary GR, Spector Y, Abbruzzese JL, et al. Prospective gene signature study using microRNA to identify the tissue of origin in patients with carcinoma of unknown primary. Clin Cancer Res 2011;17:4063-70
- Benjamin H, Lebanony D, Rosenwald S, et al. A diagnostic assay based on microRNA expression accurately identifies malignant pleural mesothelioma. J Mol Diagn 2010;12:771-9
- Rosenwald S, Gilad S, Benjamin S, et al. Validation of a microRNA-based qRT-PCR test for accurate identification of tumor tissue origin. Mod Pathol 2010;23:814-23
- Szafranska-Schwarzbach AE, Adai AT, Lee LS, et al. Development of a miRNA-based diagnostic assay for pancreatic ductal adenocarcinoma. Expert Rev Mol Diagn 2011;11:249-57
- Asuragen. Available from: http://asuragen.com/products-and-services/clinical-lab/mirinform-pancreas/
- Rosetta genomics. Available from: www.rosettagenomics.com/micrornas-in-diagnostics
- Ling H, Fabbri M, Calin GA. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat Rev Drug Discov 2013;12:847-65
- Janssen HL, Reesink HW, Lawitz EJ, et al. Treatment of HCV infection by targeting microRNA. N Engl J Med 2013;368:1685-94
- Heppner GH. Tumor heterogeneity. Cancer Res 1984;44:2259-65
- Martinez P, Birkbak NJ, Gerlinger M, et al. Parallel evolution of tumour subclones mimics diversity between tumours. J Pathol 2013;230:356-64
- Gerlinger M, Rowan AJ, Horswell S, et al. Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. N Engl J Med 2012;366:883-92
- Klein CA. Selection and adaptation during metastatic cancer progression. Nature 2013;501:365-72
- Bedard PL, Hansen AR, Ratain MJ, et al. Tumour heterogeneity in the clinic. Nature 2013;501:355-64
- Junttila MR, de Sauvage FJ. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 2013;501:346-54
- Burrell RA, McGranahan N, Bartek J, et al. The causes and consequences of genetic heterogeneity in cancer evolution. Nature 2013;501:338-45
- Meacham CE, Morrison SJ. Tumour heterogeneity and cancer cell plasticity. Nature 2013;501:328-37
- Marte B. Tumour heterogeneity. Nature 2013;501:327
- Almendro V, Marusyk A, Polyak K. Cellular heterogeneity and molecular evolution in cancer. Annu Rev Pathol 2013;8:277-302
- Sempere LF, Preis M, Yezefski T, et al. Fluorescence-based codetection with protein markers reveals distinct cellular compartments for altered MicroRNA expression in solid tumors. Clin Cancer Res 2010;16:4246-55
- Nelson PT, Wilfred BR. In situ hybridization is a necessary experimental complement to microRNA (miRNA) expression profiling in the human brain. Neurosci Lett 2009;466:69-72
- Calin GA, Dumitru CD, Shimizu M, et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA 2002;99:15524-9
- Cimmino A, Calin GA, Fabbri M, et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci USA 2005;102:13944-9
- Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature 2005;435:834-8
- Volinia S, Calin GA, Liu CG, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci USA 2006;103:2257-61
- Yanaihara N, Caplen N, Bowman E, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006;9:189-98
- Calin GA, Ferracin M, Cimmino A, et al. A MicroRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N Engl J Med 2005;353:1793-801
- Ciafre SA, Galardi S, Mangiola A, et al. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem Biophys Res Commun 2005;334:1351-8
- Iorio MV, Ferracin M, Liu CG, et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res 2005;65:7065-70
- Liu CG, Calin GA, Meloon B, et al. An oligonucleotide microchip for genome-wide microRNA profiling in human and mouse tissues. Proc Natl Acad Sci USA 2004;101:9740-4
- Calin GA, Liu CG, Sevignani C, et al. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc Natl Acad Sci USA 2004;101(32):11755-60
- Hu Z, Fan C, Oh DS, et al. The molecular portraits of breast tumors are conserved across microarray platforms. BMC.Genomics 2006;7:96
- Nelson PT, Baldwin DA, Kloosterman WP, et al. RAKE and LNA-ISH reveal microRNA expression and localization in archival human brain. RNA 2006;12:187-91
- de Planell-Saguer M, Rodicio MC. Detection methods for microRNAs in clinic practice. Clin Biochem 2013;46:869-78
- Pritchard CC, Cheng HH, Tewari M. MicroRNA profiling: approaches and considerations. Nat Rev Genet 2012;13:358-69
- Kolbert CP, Feddersen RM, Rakhshan F, et al. Multi-platform analysis of microRNA expression measurements in RNA from fresh frozen and FFPE tissues. PLoS One 2013;8:e52517
- Git A, Dvinge H, Salmon-Divon M, et al. Systematic comparison of microarray profiling, real-time PCR, and next-generation sequencing technologies for measuring differential microRNA expression. RNA 2010;16:991-1006
- Chen C, Ridzon DA, Broomer AJ, et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic acids Res 2005;33:e179
- du Rieu MC, Torrisani J, Selves J, et al. MicroRNA-21 is induced early in pancreatic ductal adenocarcinoma precursor lesions. Clin Chem 2010;56:603-12
- Hannafon BN, Sebastiani P, de las MA, et al. Expression of microRNA and their gene targets are dysregulated in preinvasive breast cancer. Breast Cancer Res 2011;13:R24
- Grutzmann R, Foerder M, Alldinger I, et al. Gene expression profiles of microdissected pancreatic ductal adenocarcinoma. Virchows Arch 2003;443:508-17
- Zhu L, Yan W, Rodriguez-Canales J, et al. MicroRNA analysis of microdissected normal squamous esophageal epithelium and tumor cells. Am J Cancer Res 2011;1:574-84
- Bloomston M, Frankel WL, Petrocca F, et al. MicroRNA expression patterns to differentiate pancreatic adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA 2007;297:1901-8
- Munding JB, Adai AT, Maghnouj A, et al. Global microRNA expression profiling of microdissected tissues identifies miR-135b as a novel biomarker for pancreatic ductal adenocarcinoma. Int J Cancer 2012;131:E86-95
- Kloosterman WP, Wienholds E, de BE, et al. In situ detection of miRNAs in animal embryos using LNA-modified oligonucleotide probes. Nat Methods 2006;3:27-9
- Wienholds E, Kloosterman WP, Miska E, et al. MicroRNA Expression in Zebrafish Embryonic Development. Science 2005;309:310-11
- Valoczi A, Hornyik C, Varga N, et al. Sensitive and specific detection of microRNAs by northern blot analysis using LNA-modified oligonucleotide probes. Nucleic Acids Res 2004;32:e175
- Kauppinen S, Vester B, Wengel J. Locked nucleic acid: high-affinity targeting of complementary RNA for RNomics. Handb Exp Pharmacol 2006;405-22
- Sempere LF, Freemantle S, Pitha-Rowe I, et al. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol 2004;5:R13
- Lagos-Quintana M, Rauhut R, Yalcin A, et al. Identification of tissue-specific microRNAs from mouse. Curr Biol 2002;12:735-9
- Sempere LF, Christensen M, Silahtaroglu A, et al. Altered MicroRNA expression confined to specific epithelial cell subpopulations in breast cancer. Cancer Res 2007;67:11612-20
- Dillhoff M, Liu J, Frankel W, et al. MicroRNA-21 is Overexpressed in Pancreatic Cancer and a Potential Predictor of Survival. J Gastrointest Surg 2008;12:2171-6
- Yamamichi N, Shimomura R, Inada K, et al. Locked nucleic acid in situ hybridization analysis of miR-21 expression during colorectal cancer development. Clin Cancer Res 2009;15:4009-16
- Habbe N, Koorstra JB, Mendell JT, et al. MicroRNA miR-155 is a biomarker of early pancreatic neoplasia. Cancer Biol Ther 2009;8:340-6
- Dyrskjot L, Ostenfeld MS, Bramsen JB, et al. Genomic profiling of microRNAs in bladder cancer: miR-129 is associated with poor outcome and promotes cell death in vitro. Cancer Res 2009;69:4851-60
- Silahtaroglu AN, Nolting D, Dyrskjot L, et al. Detection of microRNAs in frozen tissue sections by fluorescence in situ hybridization using locked nucleic acid probes and tyramide signal amplification. Nat Protoc 2007;2:2520-8
- Sempere LF, Korc M. A method for conducting highly sensitive microRNA in situ hybridization and immunohistochemical analysis in pancreatic cancer. Methods Mol Biol 2013;980:43-59
- Sempere LF. Fully automated fluorescence-based 4-color multiplex assay for co-detection of microRNA and protein biomarkers in clinical tissue specimens. Methods Mol Med 2014; In press
- Nielsen BS. MicroRNA in situ hybridization. Methods Mol Biol 2012;822:67-84
- Jorgensen S, Baker A, Moller S, et al. Robust one-day in situ hybridization protocol for detection of microRNAs in paraffin samples using LNA probes. Methods 2010;52:375-81
- Pena JT, Sohn-Lee C, Rouhanifard SH, et al. miRNA in situ hybridization in formaldehyde and EDC-fixed tissues. Nat Methods 2009;6:139-41
- Singh U, Keirstead N, Wolujczyk A, et al. General principles and methods for routine automated microRNA in situ hybridization and double labeling with immunohistochemistry. Biotech Histochem 2013;89:259-66
- Renwick N, Cekan P, Masry PA, et al. Multicolor microRNA FISH effectively differentiates tumor types. J Clin Invest 2013;123:2694-702
- Donnem T, Eklo K, Berg T, et al. Prognostic impact of MiR-155 in non-small cell lung cancer evaluated by in situ hybridization. J Transl Med 2011;9:6
- Donnem T, Lonvik K, Eklo K, et al. Independent and tissue-specific prognostic impact of miR-126 in nonsmall cell lung cancer: coexpression with vascular endothelial growth factor-A predicts poor survival. Cancer 2011;117:3193-200
- Stenvold H, Donnem T, Andersen S, et al. Stage and tissue-specific prognostic impact of miR-182 in NSCLC. BMC Cancer 2014;14:138
- Stenvold H, Donnem T, Andersen S, et al. High tumor cell expression of microRNA-21 in node positive non-small cell lung cancer predicts a favorable clinical outcome. BMC Clin Pathol 2014;14:9
- Eilertsen M, Andersen S, Al-Saad S, et al. Positive prognostic impact of miR-210 in non-small cell lung cancer. Lung Cancer 2014;83:272-8
- Li J, Yang H, Li Y, et al. microRNA-146 up-regulation predicts the prognosis of non-small cell lung cancer by miRNA in situ hybridization. Exp Mol Pathol 2014;96:195-9
- Li J, Li X, Li Y, et al. Cell-specific detection of miR-375 downregulation for predicting the prognosis of esophageal squamous cell carcinoma by miRNA in situ hybridization. PLoS One 2013;8:e53582
- Jang K, Ahn H, Sim J, et al. Loss of microRNA-200a expression correlates with tumor progression in breast cancer. Transl Res 2014;163:242-51
- Kadera BE, Li L, Toste PA, et al. MicroRNA-21 in pancreatic ductal adenocarcinoma tumor-associated fibroblasts promotes metastasis. PLoS One 2013;8:e71978
- Rask L, Balslev E, Jorgensen S, et al. High expression of miR-21 in tumor stroma correlates with increased cancer cell proliferation in human breast cancer. APMIS 2011;119:663-73
- Hagman Z, Haflidadottir BS, Ceder JA, et al. miR-205 negatively regulates the androgen receptor and is associated with adverse outcome of prostate cancer patients. Br J Cancer 2013;108:1668-76
- Schneider M, Andersen DC, Silahtaroglu A, et al. Cell-specific detection of microRNA expression during cardiomyogenesis by combined in situ hybridization and immunohistochemistry. J Mol Histol 2011;42:289-99
- Nielsen BS, Holmstrom K. Combined microRNA in situ hybridization and immunohistochemical detection of protein markers. Methods Mol Biol 2013;986:353-65
- Preis M, Gardner TB, Gordon SR, et al. microRNA-10b expression correlates with response to neoadjuvant therapy and survival in pancreatic ductal adenocarcinoma. Clin Cancer Res 2011;17:5812-21
- Liu X, Sempere LF, Guo Y, et al. Involvement of microRNAs in lung cancer biology and therapy. Transl Res 2011;157:200-8
- Nielsen BS, Jorgensen S, Fog JU, et al. High levels of microRNA-21 in the stroma of colorectal cancers predict short disease-free survival in stage II colon cancer patients. Clin Exp Metastasis 2011;28:27-38
- Kjaer-Frifeldt S, Hansen TF, Nielsen BS, et al. The prognostic importance of miR-21 in stage II colon cancer: a population-based study. Br J Cancer 2012;107:1169-74
- de Planell-Saguer M, Rodicio MC, Mourelatos Z. Rapid in situ codetection of noncoding RNAs and proteins in cells and formalin-fixed paraffin-embedded tissue sections without protease treatment. Nat Protoc 2010;5:1061-73
- Chaudhuri AD, Yelamanchili SV, Fox HS. Combined fluorescent in situ hybridization for detection of microRNAs and immunofluorescent labeling for cell-type markers. Front Cell Neurosci 2013;7:160
- Hanna JA, Wimberly H, Kumar S, et al. Quantitative analysis of microRNAs in tissue microarrays by in situ hybridization. Biotechniques 2012;52:235-45
- Hanna JA, Hahn L, Agarwal S, et al. In situ measurement of miR-205 in malignant melanoma tissue supports its role as a tumor suppressor microRNA. Lab Invest 2012;92:1390-7
- Quesne JL, Jones J, Warren J, et al. Biological and prognostic associations of miR-205 and let-7b in breast cancer revealed by in situ hybridization analysis of micro-RNA expression in arrays of archival tumour tissue. J Pathol 2012;227:306-14
- Hansen TF, Sorensen FB, Lindebjerg J, et al. The predictive value of microRNA-126 in relation to first line treatment with capecitabine and oxaliplatin in patients with metastatic colorectal cancer. BMC Cancer 2012;12:83
- Hermansen SK, Dahlrot RH, Nielsen BS, et al. MiR-21 expression in the tumor cell compartment holds unfavorable prognostic value in gliomas. J Neurooncol 2013;111:71-81
- Ma Y, Zhang P, Wang F, et al. miR-150 as a potential biomarker associated with prognosis and therapeutic outcome in colorectal cancer. Gut 2012;61:1447-53
- Qian P, Zuo Z, Wu Z, et al. Pivotal role of reduced let-7g expression in breast cancer invasion and metastasis. Cancer Res 2011;71:6463-74
- Tang W, Zhu J, Su S, et al. MiR-27 as a prognostic marker for breast cancer progression and patient survival. PLoS One 2012;7:e51702
- Fish JE, Santoro MM, Morton SU, et al. miR-126 regulates angiogenic signaling and vascular integrity. Dev Cell 2008;15:272-84
- Meister J, Schmidt MH. miR-126 and miR-126*: new players in cancer. ScientificWorldJournal 2010;10:2090-100
- Metzinger-Le Meuth V, Andrianome S, Chillon JM, et al. microRNAs are dysregulated in the cerebral microvasculature of CKD mice. Front Biosci 2014;6:80-8
- Nikolic I, Plate KH, Schmidt MH. EGFL7 meets miRNA-126: an angiogenesis alliance. J Angiogenes Res 2010;2:9
- Schober A, Nazari-Jahantigh M, Wei Y, et al. MicroRNA-126-5p promotes endothelial proliferation and limits atherosclerosis by suppressing Dlk1. Nat Med 2014;20:368-76
- Urbich C, Kuehbacher A, Dimmeler S. Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovasc Res 2008;79:581-8
- Wang S, Aurora AB, Johnson BA, et al. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev Cell 2008;15:261-71
- Anastasov N, Hofig I, Vasconcellos IG, et al. Radiation resistance due to high expression of miR-21 and G2/M checkpoint arrest in breast cancer cells. Radiat Oncol 2012;7:206
- Bullock MD, Pickard KM, Nielsen BS, et al. Pleiotropic actions of miR-21 highlight the critical role of deregulated stromal microRNAs during colorectal cancer progression. Cell Death Dis 2013;4:e684
- Wang M, Yang W, Li M, et al. Low expression of miR-150 in pediatric intestinal Burkitt lymphoma. Exp Mol Pathol 2014;96:261-6
- Curtis C, Shah SP, Chin SF, et al. The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups. Nature 2012;486:346-52
- Schwarzenbach H, Nishida N, Calin GA, et al. Clinical relevance of circulating cell-free microRNAs in cancer. Nat Rev Clin Oncol 2014;11:145-56
- Del VV, Cantaloni C, Cucino A, et al. miR-205 Expression levels in nonsmall cell lung cancer do not always distinguish adenocarcinomas from squamous cell carcinomas. Am J Surg Pathol 2011;35:268-75
- Fassina A, Cappellesso R, Fassan M. Classification of non-small cell lung carcinoma in trans-thoracic needle specimens using microRNA expression Profiling. Chest 2011;140:1305-11
- Bishop JA, Benjamin H, Cholakh H, et al. Accurate classification of non-small cell lung carcinoma using a novel microRNA-based approach. Clin Cancer Res 2010;16:610-19
- Xing L, Todd NW, Yu L, et al. Early detection of squamous cell lung cancer in sputum by a panel of microRNA markers. Mod Pathol 2010;23:1157-64
- Lebanony D, Benjamin H, Gilad S, et al. Diagnostic assay based on hsa-miR-205 expression distinguishes squamous from nonsquamous non-small-cell lung carcinoma. J Clin Oncol 2009;27:2030-7
- Noh S, Shim H. Optimal combination of immunohistochemical markers for subclassification of non-small cell lung carcinomas: a tissue microarray study of poorly differentiated areas. Lung Cancer 2012;76:51-5
- Mukhopadhyay S, Katzenstein AL. Subclassification of non-small cell lung carcinomas lacking morphologic differentiation on biopsy specimens: utility of an immunohistochemical panel containing TTF-1, napsin A, p63, and CK5/6. Am J Surg Pathol 2011;35:15-25
- Markou A, Tsaroucha EG, Kaklamanis L, et al. Prognostic value of mature microRNA-21 and microRNA-205 overexpression in non-small cell lung cancer by quantitative real-time RT-PCR. Clin Chem 2008;54:1696-704
- Romero Lauro G, Cable W, Lesniak A, et al. Digital pathology consultations-a new era in digital imaging, challenges and practical applications. J Digit Imaging 2013;26:668-77
- Ghaznavi F, Evans A, Madabhushi A, et al. Digital imaging in pathology: whole-slide imaging and beyond. Annu Rev Pathol 2013;8:331-59
- Sabatier R, Goncalves A, Bertucci F. Personalized medicine: present and future of breast cancer management. Crit Rev Oncol Hematol 2014. [Epub ahead of print]
- Jennings L, Van Deerlin VM, Gulley ML, et al. Recommended principles and practices for validating clinical molecular pathology tests. Arch Pathol Lab Med 2009;133:743-55
- O’Hurley G, Sjostedt E, Rahman A, et al. Garbage in, garbage out: a critical evaluation of strategies used for validation of immunohistochemical biomarkers. Mol Oncol 2014;8(4):783-98
- Dancey JE, Dobbin KK, Groshen S, et al. Guidelines for the development and incorporation of biomarker studies in early clinical trials of novel agents. Clin Cancer Res 2010;16:1745-55
- Cancer therapy evaluation program. Available from: http://ctep.cancer.gov/
- Trang P, Wiggins JF, Daige CL, et al. Systemic delivery of tumor suppressor microRNA mimics using a neutral lipid emulsion inhibits lung tumors in mice. Mol Ther 2011;19:116-1122
- Trang P, Medina PP, Wiggins JF, et al. Regression of murine lung tumors by the let-7 microRNA. Oncogene 2010;29:1580-7
- Cubillos-Ruiz JR, Baird JR, Tesone AJ, et al. Reprogramming tumor-associated dendritic cells in vivo using miRNA mimetics triggers protective immunity against ovarian cancer. Cancer Res 2012;72:1683-93
- Wei J, Feng L, Li Z, et al. MicroRNA-21 activates hepatic stellate cells via PTEN/Akt signaling. Biomed Pharmacother 2013;67:387-92
- Han M, Wang Y, Liu M, et al. MiR-21 regulates epithelial-mesenchymal transition phenotype and hypoxia-inducible factor-1alpha expression in third-sphere forming breast cancer stem cell-like cells. Cancer Sci 2012;103:1058-64
- Gene silencing news. Available from: www.genomeweb.com/rnai/regulus-considers-mir-21-drug-next-clinical-candidate-amid-positive-alport-data
- Liu X, Sempere LF, Galimberti F, et al. Uncovering growth-suppressive MicroRNAs in lung cancer. Clin Cancer Res 2009;15:1177-83
- Liu X, Sempere LF, Ouyang H, et al. MicroRNA-31 functions as an oncogenic microRNA in mouse and human lung cancer cells by repressing specific tumor suppressors. J Clin Invest 2010;120:1298-309
- Li T, Li RS, Li YH, et al. miR-21 as an independent biochemical recurrence predictor and potential therapeutic target for prostate cancer. J Urol 2012;187:1466-72