
Abstract
Middle East respiratory syndrome (MERS) is a newly emerging infectious disease caused by a novel coronavirus, MERS-coronavirus (MERS-CoV), a new member in the lineage C of β-coronavirus (β-CoV). The increased human cases and high mortality rate of MERS-CoV infection make it essential to develop safe and effective vaccines. In this review, the current advancements and potential strategies in the development of MERS vaccines, particularly subunit vaccines based on MERS-CoV spike (S) protein and its receptor-binding domain (RBD), are discussed. How to improve the efficacy of subunit vaccines through novel adjuvant formulations and routes of administration as well as currently available animal models for evaluating the in vivo efficacy of MERS-CoV vaccines are also addressed. Overall, these strategies may have important implications for the development of effective and safe vaccines for MERS-CoV in the future.
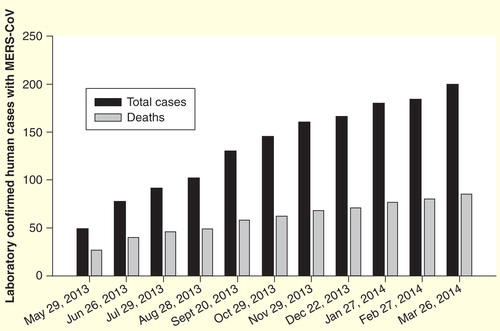
A novel β-coronavirus (β-CoV) named Middle East respiratory syndrome (MERS) coronavirus (MERS-CoV) was first identified in humans in the Middle East in June 2012. The virus was isolated from a 60-year-old Saudi man who was diagnosed with progressive community-acquired pneumonia and subsequent acute renal failure with a fatal outcome Citation[1]. Increasing numbers of cases have resulted from MERS-CoV infection as shown in Citation[2]. As of 26 March 2014, a total of 200 human MERS-CoV infections with 85 deaths had been reported in the Middle East, North Africa and several European countries by WHO Citation[2].
Figure 1. Laboratory confirmed cases of human infection with MERS-coronavirus. The black columns indicate the total laboratory confirmed human cases infected with MERS-CoV, while the gray columns demonstrate the total deaths caused by MERS-CoV infection from September 2012 to the indicated date.
While many animals are considered reservoirs for CoVs, the origin of MERS-CoV and mode of disease transmission still remain unknown. Based on a sequence fragment identical to MERS-CoV found in a Taphozous perforates bat captured in Saudi Arabia, it has been inferred that bats are the most likely reservoir for MERS-CoV Citation[3]. The identification of MERS-CoV in dromedary camels and the detection of neutralizing antibodies in the sera of these animals suggest that camels could be the intermediate host of MERS-CoV Citation[4,5]. It is thus suspected that MERS-CoV emerged as interspecies transmission from bats to other animals, such as camels, and then to humans Citation[6].
Several family clusters with MERS-CoV infection have been reported Citation[7,8], suggesting human-to-human transmission of the disease. Most people who were infected with MERS-CoV developed severe acute respiratory illness with symptoms like fever, cough and shortness of breath. The virus has an estimated incubation period of 12 days Citation[9]. The transmissibility of MERS-CoV between humans is currently less efficient than that of severe acute respiratory syndrome CoV (SARS-CoV), which emerged in Guangdong Province in China in 2002 and re-emerged in 2004, with over 8000 cases and more than 700 deaths in 29 countries throughout the world Citation[10]. Nonetheless, the mortality rate for MERS (∼43%) is much higher than that for SARS (∼10%) Citation[11]. It has been reported that MERS-CoV, but not SARS-CoV, could replicate in cell lines from four major chiropteran families, implying the ability of MERS-CoV in crossing species barrier between humans and bats Citation[12]. With increased human cases and high mortality rate, MERS-CoV infection has raised serious global concerns, although it is still too early to implicate any impending pandemic. Vaccination is considered one of the most effective strategies to prevent virus infection. Thus, development of effective vaccines is essential to fight against this infectious disease.
CoV classification & MERS-CoV characterization
As proposed by the Coronavirus Study Group of the International Committee for Taxonomy of Viruses (ICTV), three groups, 1, 2 and 3, of the Coronaviridae family have been commonly accepted as α-CoV, β-CoV and γ-CoV, respectively, where the genus β-CoV consists of four lineages, namely, A, B, C and D Citation[13]. A novel genus, δ-CoV, including HKU11, HKU12 and HKU13 identified in birds in 2005, was added in the CoV genera Citation[13].
Unlike any other previously identified CoVs, such as α-CoV NL63, which was first reported in 2004 Citation[14], as well as β-CoV HKU1, which was first discovered in Hong Kong in 2005 Citation[15,16] and SARS-CoV, which caused a worldwide outbreak in 2003 Citation[17], MERS-CoV belongs to lineage C of β-CoV, and is the first known lineage C β-CoV associated with human infections Citation[18]. Phylogenetically, MERS-CoV is closely related to bat CoVs HKU4 and HKU5 Citation[19–21].
MERS-CoV is a positive-sense, single-stranded RNA virus. Its genome is about 30 kilobase (kb) pairs in length, consisting of 11 functional open reading frames (ORFs) . The viral genome encodes five unique proteins, known as accessory proteins (3, 4a, 4b, 5, 8b), which serve different functions with no known homologs to any other CoV proteins. Accessory protein 4a, for example, was recently proven to block the production of host interferon (IFN) Citation[22]. Although these accessory proteins are not essential for virus replication, recent studies by reverse genetics demonstrated that the absence of the genes encoding these proteins as a group may attenuate viral titers Citation[23]. Additionally, the genome of MERS-CoV encodes six proteins, which are homologous to those of the known CoVs, including two replicase proteins, ORFs 1a and 1b and four major structural proteins, such as spike (S), envelope (E), membrane (M) and nucleocapsid (N) proteins Citation[11]. The E protein is a transmembrane protein forming an ion channel on the viral surface. The CoV N protein plays an important role in encapsidating the genomic RNA and interacting with the M protein and other N molecules Citation[24].
Figure 2. Genomic and schematic diagram of MERS-coronavirus structure. (A) MERS-CoV genomic structure. Viral genes, including ORF 1a, ORF 1b, S, 3, 4a, 4b, 5, E, M, 8b and N (GenBank accession number: JX869059) and their respective lengths, are indicated by rectangular boxes in the scheme. (B) Schematic diagram of MERS-CoV structure. MERS-CoV contains four structural proteins. The S protein is a type I transmembrane glycoprotein displayed on the surface of viral membrane as an oligomer. The E protein is also a transmembrane protein that forms an ion channel on the viral surface. The N protein plays an important role in encapsidating the genomic RNA and interacting with the membrane M protein and other N molecules.
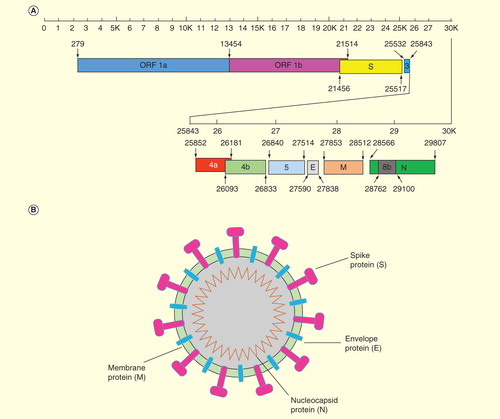
Table 1. MERS-coronavirus RNA genome and protein coding assignments.
The S protein of CoVs is a type I transmembrane glycoprotein displayed as an oligomer on the surface of the viral membrane. The precursor S protein contains a cleavage site at which the protein could be cleaved into two non-covalently associated subunits: the distal subunit S1 and the membrane-anchored subunit S2 Citation[25]. The S1 subunit contains the cellular receptor-binding domain (RBD) Citation[26], while the S2 subunit contains a putative fusion peptide, transmembrane domain and two heptad repeat regions: the heptad repeat 1 and 2 (HR1 and HR2) Citation[27]. lists the representative structures of S proteins of α- and β-CoV NL63-CoV, mouse hepatitis CoV (MHV-CoV) and SARS-CoV Citation[28–30], with the RBDs of these CoV S proteins consisting of respective core and receptor-binding subdomains. For example, the receptor-binding motif of SARS-CoV contains residues from 424 to 494, which form a long extended loop with two short antiparallel β-sheets and one disulfide bond between C467 and C474 Citation[31].
Figure 3. Schematic diagrams of spike protein structures of representative coronaviruses. Schematic diagrams of the structures of S proteins of the previously identified coronaviruses NL63-CoV, MHV-CoV, SARS-CoV (A), and the newly emerged coronavirus MERS-CoV (B) are indicated. The S proteins contain S1 and S2 subunits, respectively, with RBDs located in the S1 region. RBMs are functional regions within RBDs for receptor recognition. Other functional domains, such as fusion peptide, HR1, HR2, transmembrane and cytoplasmic tail, contain variant lengths within different coronaviruses.
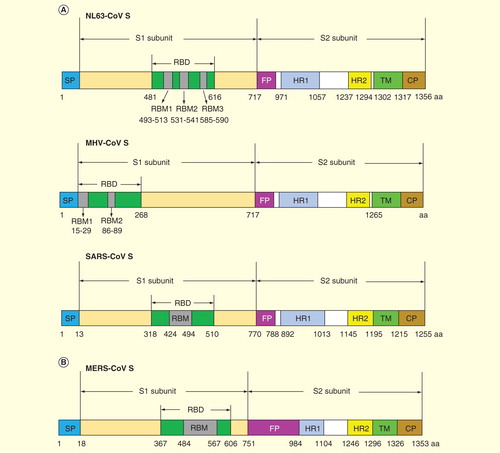
Similar to the S proteins of other CoVs, the S protein of MERS-CoV also contains S1 and S2 subunits, with S1 being responsible for receptor binding and S2 for membrane fusion Citation[32]. Unlike CoVs SARS-CoV and NL63-CoV, which use human angiotensin-converting enzyme 2 as their receptors Citation[33,34], MERS-CoV utilizes human dipeptidyl peptidase 4 (DPP4, also known as CD26) as its cellular receptor. DPP4 is expressed on primary bronchiolar epithelial cells and regulates the activity of hormones and chemokines through proteolytic cleavage Citation[12,35]. Recent studies have solved the crystal structures of MERS-CoV S protein alone, or complexed with its receptor DPP4, and mapped the RBD of MERS-CoV to the residues covering 367–588 or 367–606, respectively Citation[32,36,37]. According to the crystal structures, the RBD of MERS-CoV also contains the core and receptor-binding subdomains, with the receptor-binding motif consisting of 84 amino acids (from V484 to L567) that form a four-stranded antiparallel β-sheet Citation[32,36]. Similar to other CoVs, the HR1 and HR2 of MERS-CoV form a 6-helix bundle believed to drive virion-cell membrane fusion, with three HR1 helices forming the central coiled-coil core and three HR2 chains surrounding the core in the HR1 side grooves Citation[23,38,39]. describes the crystal structures of SARS-CoV and MERS-CoV RBD complexed with their respective receptors, and the major neutralizing epitopes on SARS-CoV RBD for the neutralizing monoclonal antibody against SARS-CoV Citation[36,40–42]. Understanding of these neutralizing epitopes in RBD of SARS-CoV will provide useful guidance for the identification of the neutralizing epitopes in MERS-CoV RBD and, hence designing RBD-based, neutralizing epitope-dependent vaccines against MERS-CoV.
Figure 4. Crystal structures of SARS-CoV and MERS-CoV RBD and neutralizing epitopes on SARS-CoV RBD. (A) Crystal structure of SARS-CoV RBD complexed with its receptor ACE2 (blue) (PDB ID: 2AJF) Citation[31]. (B) Neutralizing epitope for mAb m396 Fab on SARS-CoV RBD (PDB ID: 2DD8) Citation[41]. SARS-CoV RBD is in black with RBM region in yellow. (C) Neutralizing epitope for mAb F26G19 Fab on SARS-CoV RBD (PDB ID: 3BGF) Citation[40]. (D) Neutralizing epitope for mAb 80R scFv on SARS-CoV RBD (PDB ID: 2GHW) Citation[42]. (E) Crystal structure of MERS-CoV RBD. The RBM is in yellow. (PDB ID: 4KQZ) Citation[36]. (F) Crystal structure of MERS-CoV RBD (black) complexed with its receptor human DPP4 (blue) (PDB ID: 4KR0) Citation[37].
![Figure 4. Crystal structures of SARS-CoV and MERS-CoV RBD and neutralizing epitopes on SARS-CoV RBD. (A) Crystal structure of SARS-CoV RBD complexed with its receptor ACE2 (blue) (PDB ID: 2AJF) Citation[31]. (B) Neutralizing epitope for mAb m396 Fab on SARS-CoV RBD (PDB ID: 2DD8) Citation[41]. SARS-CoV RBD is in black with RBM region in yellow. (C) Neutralizing epitope for mAb F26G19 Fab on SARS-CoV RBD (PDB ID: 3BGF) Citation[40]. (D) Neutralizing epitope for mAb 80R scFv on SARS-CoV RBD (PDB ID: 2GHW) Citation[42]. (E) Crystal structure of MERS-CoV RBD. The RBM is in yellow. (PDB ID: 4KQZ) Citation[36]. (F) Crystal structure of MERS-CoV RBD (black) complexed with its receptor human DPP4 (blue) (PDB ID: 4KR0) Citation[37].](/cms/asset/3a846431-04d9-4014-80a7-f583744ba9a4/ierv_a_912134_f0004_b.jpg)
Previous reviews on SARS have summarized the approaches for the development of effective CoV vaccines, pointed out the importance of SARS-CoV S protein as a target for vaccine and therapeutic development and outlined a roadmap for the product development of a SARS-CoV RBD protein-based subunit vaccine with manufacturing for future clinical testing Citation[43–46]. In this review, we will briefly discuss current stages of MERS vaccine development, and provide potential strategies for developing MERS vaccines based on the experience from development of SARS vaccines, with the focus on subunit vaccines and how to improve the efficacy of MERS subunit vaccines. In addition, we also summarize current animal models for MERS-CoV and emphasize the importance for evaluation of efficacy of MERS vaccine candidates in effective animal models.
Current advancements on MERS vaccine development
No currently licensed MERS vaccines are available for use. However, researchers are working to develop potential vaccines, and recent successes in the engineering of a replication-competent, propagation-defective MERS-CoV using reverse genetics provide the possibility of developing attenuated viruses as MERS vaccine candidates Citation[47].
SARS studies have demonstrated that vaccines inducing strong neutralizing antibodies completely protected immunized subjects against SARS-CoV infection. Moreover, recovered patients infected with SARS-CoV could maintain potent and persistent RBD-specific antibody responses with neutralizing activity Citation[48], revealing the significance of neutralizing antibodies in the prevention of SARS. Like the case of SARS-CoV vaccines, although vaccines eliciting favorable cellular immune responses may play a role in the clearance of virus infection, vaccines inducing neutralizing antibodies will be sufficient for protection from MERS-CoV infection. Thus, the ability of eliciting neutralizing antibodies is expected to be one of the important criteria for evaluating the efficacy of MERS vaccines.
Previous studies of SARS-CoV have shown that the S protein, particularly the RBD, may induce highly potent neutralizing antibodies that protected vaccinated animals against SARS-CoV challenge Citation[43,49–51]. It is thus expected that the S protein in MERS-CoV will also be important for the development of vaccines. Song et al. showed that vaccination with a recombinant-modified vaccinia virus Ankara (MVA) expressing the full-length S protein of MERS-CoV induced high levels of neutralizing antibodies in vaccinated mice Citation[52]. However, Cao and colleagues have reported that although the MVA-based, full-length S protein-dependent SARS vaccine could induce neutralizing antibody responses in ferrets, it failed to protect the vaccinated animals from SARS-CoV challenge. Even worse, this vaccine was associated with enhanced inflammatory and immunopathological effects, resulting in serious liver damage in the vaccinated ferrets after viral challenge Citation[53]. Therefore, caution should be taken when the full-length S protein of MERS-CoV is used as a vaccine antigen since the non-neutralizing epitopes in S protein may induce antibody-mediated disease enhancement (ADE) effects, as those elicited by the S protein of feline infectious peritonitis virus (FIPV, also a CoV) Citation[54].
Therefore, it is favorable to develop vaccines based on S protein fragments, particularly RBD, of MERS-CoV, and such strategy has been reported to be effective in the development of MERS vaccines. Studies have shown that neutralization against MERS-CoV infection could be elicited in vaccinated rabbits by a fragment containing residues 358–588 of MERS-CoV S protein Citation[55]. The induction of neutralizing activity by a recombinant protein fragment containing residues 377–662 of MERS-CoV RBD has also been demonstrated. Particularly, a truncated 212-amino acid RBD fragment containing residues 377–588 (inside RBD: residues 367–606) of MERS-CoV S protein induced strong MERS-CoV S-specific antibodies in vaccinated mice, blocking the binding of MERS-CoV RBD to viral receptor DPP4 and effectively neutralizing MERS-CoV infection, thus providing promise for further development as an effective and safe MERS vaccine Citation[56,57].
In addition to the S protein, other MERS-CoV viral proteins might play a role in the development of MERS candidate vaccines. However, few studies have addressed this possibility, in particular because MERS has only recently emerged as a novel human disease, and vaccine development is a time-consuming process. A recent study reveals that structural and accessory proteins, such as M, ORF 4a, ORF 4b and ORF 5, of MERS-CoV may function as potent IFN antagonists Citation[58], with ORF 4a protein showing the greatest potential for counteracting the antiviral effects of IFN by inhibiting IFN production and IFN-sensitive response element signaling pathways. Therefore, these proteins may also serve as targets for developing MERS vaccines.
Potential strategies for developing MERS vaccines
Possibility for developing MERS vaccines with various types
Traditionally, inactivated and live-attenuated virus vaccines have been the most commonly developed and used vaccine types against viral pathogens, including SARS-CoV. Other types of vaccines, such as those based on DNA, viral vectors, virus-like particles (VLPs) and recombinant proteins, have also been discussed in terms of their potential application as MERS vaccine candidates. Because only a few papers related to the MERS-CoV vaccine development have been published so far, the rest of the review will discuss the development of SARS-CoV vaccines, from which we can learn what the best strategy is for developing MERS-CoV vaccine and how to move it forward more efficiently. The main advantages and disadvantages of different strategies for CoV vaccine development are listed in .
Table 2. The potential strengths and weaknesses of different types of coronavirus vaccines.
Inactivated and live-attenuated virus vaccines
The inactivated and live-attenuated SARS-CoV has been tested as vaccine candidates, some of which are proven to be effective to provide protective immunity against SARS-CoV infection Citation[59–61]. It was previously reported that a β-propiolactone-inactivated SARS-CoV vaccine administered subcutaneously (s.c.) to mice was more effective in inhibiting virus replication in the murine respiratory tract than the adenovirus-based N and S vaccine given either intranasally (i.n.) or intramuscularly (i.m.) Citation[62]. In addition, a live, impaired-fidelity SARS-CoV vaccine with engineered inactivation of exonuclease activity protects aged and immunocompromised mice against lethal virus challenge Citation[63]. Apparently, both local and serum antibody responses can be induced in inactivated SARS-CoV vaccinated mice Citation[64]. Like SARS-CoV, it might be possible to attenuate MERS-CoV by deleting the envelope E protein to diminish virus growth in tissue culture, while abrogating virulence in animals Citation[65,66], or by inactivating exonuclease activity. In spite of the possibility of inducing highly potent immune responses and protection, vaccine candidates based on the inactivated and live-attenuated MERS-CoV might have the potential of recovering virulence, raising safety concerns; as such, this vaccine type may be inappropriate for use by highly immunosuppressed individuals.
DNA vaccines
DNA vaccines consist of a plasmid DNA, which has been genetically engineered to produce immunological response to protect organisms against diseases. DNA is usually administered through two routes, i.m. delivery and bombardment of the skin using a gene gun Citation[67]. Currently, two popular theories of DNA uptake are used, including non-specific phagocytosis or pinocytosis and antigen-specific receptor-mediated uptake Citation[68,69]. Containing non-replicating and non-infectious components and only encoding the protein of interest, DNA vaccines are safe for use, minimizing undesirable side effects Citation[67]. In addition, vaccines based on DNAs are easy to be scaled up by the fermentation technique Citation[70–72] and are generally stable and inexpensive.
The efficacy of DNA-based vaccines against SARS-CoV infection has been widely reviewed. It is revealed that a DNA vaccine encoding SARS-CoV S protein induces neutralizing antibody as well as T-cell responses that protect vaccinated mice from SARS-CoV challenge Citation[73]. In addition, IgG antibody and T-cell-mediated immune responses are induced by DNA vaccines encoding S1 of SARS-CoV Citation[74]. Interestingly, expression of M protein of SARS-CoV can augment immune responses induced by NP protein of a SARS-CoV DNA vaccine Citation[75]. Nevertheless, compared with other vaccine types, such as inactivated and live-attenuated virus vaccines, DNA-based vaccines usually have lower immunogenicity Citation[76], indicating the need to further improve their immune efficacy. Although it is possible to use similar strategies to develop DNA-based vaccines against MERS-CoV infection, suitable measures such as priming-boost strategy or combination of DNA vaccines with other vaccination types should be considered to improve the immunogenicity of DNA-based MERS vaccines Citation[77].
Viral vector-based vaccines
Viral vectors have been applied as an important delivery vehicle for developing candidate vaccines against SARS-CoV infection. Currently, reported viral vectors for SARS vaccines include MVA, vesicular stomatitis virus, rhabdovirus, adenovirus, adeno-associated virus and attenuated parainfluenza virus Citation[78–80]. Previous reports have indicated the ability of an MVA-based vaccine expressing SARS-CoV S protein in the induction of neutralizing antibody with protection of vaccinated mice against SARS-CoV challenge Citation[78]. In addition, a combination of adenoviral-based SARS vaccines encoding S1, M and NP protein, respectively, induced strong neutralizing antibodies and T-cell immune responses, protecting immunized monkeys from challenge of SARS-CoV Citation[80]. The induction of sufficient cellular and humoral immune responses has also been demonstrated with neutralizing activity by an adeno-associated virus-based vaccine containing RBD of SARS-CoV in the protection of SARS-CoV challenge Citation[81,82].
The above approaches for developing viral vector-based SARS vaccines can be adopted for the development of MERS vaccines. Indeed, MERS-CoV S protein delivered by an MVA vector induces in the vaccinated mice antibody responses with neutralizing activity Citation[52], bringing some hopes for the development of viral vector-based vaccines to prevent MERS-CoV infection. However, it should be noted that viral vector-based vaccines do have some disadvantages. For example, some vectors, such as MVA or adenovirus, have pre-existing immunity, or cause harmful immune responses Citation[83,84] and inflammation Citation[85]. Thus, caution should be taken when developing MERS vaccines using viral vectors.
VLPs as vaccines
VLP-based vaccines have been developed as a new generation of non-egg-based, cell culture-derived vaccine candidates against virus infection, and thus have gained much more attention for their potential promise in developing effective and safe vaccines against SARS-CoV Citation[86]. VLPs can be produced in several expression systems, such as mammalian cells (e.g., 293T, CHO), yeast, recombinant vaccinia virus and baculovirus expression systems Citation[87,88]. It is worth noting that the minimal components essential for VLP formation and the immunogenicity induced by VLPs need to be considered. Although M and E proteins of SARS-CoV are sufficient for the efficient formation of VLPs Citation[89], the immunogenicity induced by VLPs without S protein might be greatly reduced. By comparison, chimeric VLPs carrying SARS-CoV S protein, and E, M and N proteins of MHV-CoV elicited high levels of SARS-CoV-specific neutralizing antibodies, protecting vaccinated mice from SARS-CoV challenge Citation[88].
The above findings on SARS-CoV VLPs may be applied to constructing VLPs for MERS-CoV. Compared with inactivated and live-attenuated virus vaccines, VLPs do not contain the infectious materials, making VLP technology a promising alternative for MERS vaccine development. However, the immunogenicity of VLPs is relatively lower than that of inactivated and live-attenuated virus vaccines. Several strategies may be explored to increase the immunogenicity of VLPs. For example, dendritic cell-stimulating molecules or Toll-like receptor 5 ligand flagellin can be genetically engineered for incorporation into MERS-CoV VLPs to increase immunogenicity and enhance protective efficacy against further heterosubtypic virus challenge Citation[90,91].
Subunit vaccines
Compared with other types of vaccines, subunit vaccines can claim several unique advantages. The most important characteristic of subunit vaccines is their safety in that the components only contain synthetic peptides or recombinant proteins expressing specific immunogenic fragments of a pathogen, without the involvement of infectious viruses. Additionally, this type of vaccine is less likely to induce side effects at the injection sites, and consistent production can be attained for subunit vaccines because of constant conditions and well-defined pathogenic fragments. The above features have made subunit vaccines an attractive vaccine candidate.
Like SARS-CoV S protein, the S protein of MERS-CoV is also considered an important target for development of subunit vaccines, because it is the main antigenic component responsible for inducing host immune responses. Since the S protein contains regions for receptor binding and membrane fusion, S protein-based vaccines are expected to induce antibodies to block virus binding and subsequent membrane fusion or neutralize viral infection Citation[43]. However, the S protein of SARS-CoV also contains some non-neutralizing epitopes in the immmunopredominant domain Citation[92], which may induce harmful immune responses Citation[85]. Therefore, it is essential to identify and exclude the epitopes in the S protein that may induce harmful or ADE responses from the S protein-based MERS-CoV vaccines.
Experience from developing RBD-based SARS subunit vaccines provides important guidance on the rational design of RBD protein-based MERS vaccines. Since the RBD of SARS-CoV contains multiple neutralizing epitopes, RBD-based SARS subunit vaccines covering these neutralizing epitopes are able to induce potent neutralizing antibodies completely preventing SARS-CoV challenge Citation[50,51]. Similarly, it has been shown that the RBD of MERS-CoV induced strong neutralizing antibody responses against infection of MERS-CoV Citation[56,57] and that the MERS-CoV RBD does, indeed, contain several important neutralizing epitopes Citation[93]. Accordingly, further optimization of the RBD and development of optimized RBD-based MERS subunit vaccines containing multiple neutralizing epitopes would be desirable and feasible to improve the vaccine efficacy.
Both MERS-CoV and SARS-CoV RBDs consist of a core and a receptor-binding subdomain. They share a high degree of structural similarity in the core subdomains, but the receptor-binding subdomains are notably divergent Citation[32]. Because of variation of the receptor-binding subdomain region within subgroups of the same CoV or across different CoV groups, it is advisable to rationally design MERS subunit vaccines using chimeric S protein containing several neutralizing epitopes from divergent subgroups. This approach will provide a strategic platform for the rational design of subunit vaccines against future emerging CoVs by focusing on the chimeric S protein containing neutralizing epitopes from multiple virus strains across different subgroups Citation[94].
To summarize, although the inactivated and live-attenuated virus and viral vector-based vaccines are able to induce strong immune responses, their tendency to recover virulence or cause harmful immune responses raises safety concerns for the development of MERS vaccines. While DNA-based vaccines have a high safety profile, they may be unable to induce highly potent immune response and protection in humans and might cause toxic effect at the injection sites with the repeated vaccinations. By comparison, development of recombinant RBD protein-based MERS subunit vaccines remains a high priority, due to a variety of advantages noted above.
Potential strategies for improving the immunogenicity of MERS subunit vaccines
In spite of a number of merits, protein-based subunit vaccines might have relatively lower immunogenicity as compared with inactivated and live-attenuated vaccines Citation[95], necessitating continued improvement in the immunogenicity of subunit vaccines. Two possibilities are discussed as potential strategies to improve the immunogenicity of MERS vaccine candidates.
Formulation of subunit vaccines with suitable adjuvants
To improve the immunogenicity of MERS candidate vaccines, adjuvants should be selected and formulated on the basis of their mechanisms of action. Normally, vaccines with limited immunogenicity require the addition of specific adjuvants to stimulate a protective and long-lasting immune response Citation[96–98]. Although a variety of novel adjuvants are now emerging and being investigated, only a few have been approved for clinical use. Both aluminum salt-based adjuvants, usually called ‘Alum’ and MF59TM can induce antigen-specific humoral and CTL responses Citation[99,100]; hence, they have the potential for use in MERS-CoV subunit vaccines. Another adjuvant named glucopyranosyl lipid A, a synthetic TLR4 agonist, was also shown to improve the immunogenicity of a protein-based vaccine candidate Citation[101]. Other adjuvants, such as Montanide ISA51, which has been approved for human use in Europe Citation[102,103], is used in making MERS and influenza subunit vaccines, demonstrating a strong ability to promote the induction of highly potent immune responses of these candidate vaccines Citation[56,57,104]. Furthermore, the combination of two adjuvants with distinct mechanisms of action is also feasible to improve the efficacy of designed MERS subunit vaccines. Since Alum alone cannot induce strong Th1 immune responses, it is thus applicable to combine Alum with other adjuvants with potential complementary effects in a hope to improve the efficacy induced by Alum-adjuvanted subunit vaccines. For example, combination of Alum plus glucopyranosyl lipid A will be applied to improve the efficacy of a SARS-CoV RBD protein-based subunit vaccine Citation[45]. Therefore, similar strategies can be utilized in MERS-CoV subunit vaccines to improve their immunogenicity.
Selection of suitable administration routes
Several administration routes, including i.m., intradermal, s.c. and i.n. pathways, have been used for vaccine immunization in both experimental research and clinical use. The route of vaccine administration directly influences both the quality and quantity of vaccine-induced immunity Citation[105–108]. Traditional parenteral administration routes, such as i.m., intradermal and s.c., have been shown to induce favorable immune responses and/or protection against SARS-CoV and MERS-CoV infections Citation[57,109–111]. It is demonstrated that i.m. vaccination with RBD protein of SARS-CoV elicits long-term immune responses with neutralizing activity, protecting immunized mice from SARS-CoV challenge Citation[50]. Additionally, potent neutralizing antibodies and protection against SARS-CoV infection are observed in the mice s.c. vaccinated with SARS-CoV RBD proteins, respectively, expressed in mammalian cell, insect cell and Escherichia coli expression systems Citation[49,112]. Furthermore, strong neutralizing antibody responses are revealed in the mice s.c.-vaccinated MERS-CoV RBD proteins Citation[56].
In addition to inducing systemic immunity, the stimulation of local immunity at mucosal sites has become an important objective for vaccination. Although there are some unfavorable features from the i.n. route, such as the possibility of delivering antigens into the central nervous system Citation[113], as well as the unsuitability for children younger than 2 years and adults older than 50 years, especially those with chronic pulmonary diseases and immunosuppression Citation[114], the advantages of the i.n. route outweigh its disadvantages. Reports have indicated that i.n. vaccination effectively induced both local and systemic immune responses, especially mucosal IgA, against mucosal pathogens, cross-protecting vaccinated animals from infection by homologous and heterologous strains of influenza virus Citation[115], as well as providing long-term protection against SARS-CoV infection Citation[81]. As an important mucosal pathogen infecting humans via the mucosal pathway Citation[9], MERS vaccines delivered via the mucosal route are anticipated to elicit effective mucosal immune responses, as represented by mucosal IgA. Intranasal immunization of a subunit candidate vaccine containing recombinant RBD protein fused with Fc of human IgG has been demonstrated to induce strong anti-RBD- and anti-S1-specific neutralizing antibody responses, particularly IgA mucosal immune responses, against infections from MERS-CoV Citation[93], providing a fundamental basis for developing effective MERS mucosal vaccines.
Therefore, selection of suitable vaccination routes, combined with antigen formulation with appropriate adjuvants, will play important guidance for developing MERS-CoV subunit vaccines and further evaluating their efficacy against MERS-CoV infection.
Animal models for MERS-CoV
Establishment of effective animal models is important for evaluating the efficacy of candidate vaccines against MERS-CoV. Unlike SARS-CoV, which can infect a variety of animals, including non-human primates, and small animal models such as ferrets, hamsters and mice Citation[76,78,116–118], the animal species that could be infected by MERS-CoV are very limited. It has been shown that MERS-CoV is unable to replicate in hamsters, ferrets and mice Citation[119–122], significantly restricting the efficacy evaluation of MERS vaccines in small animal models. Although rhesus macaques can be infected by MERS-CoV, with the infected animals showing clinical signs of disease and virus replication Citation[123,124], the high cost of non-human primate models would prevent the majority of researchers from using this animal model. Therefore, the development of effective small animal models for MERS-CoV infection is urgently needed Citation[125]. It was recently reported that mice transduced with adenoviral vectors expressing MERS-CoV’s receptor DPP4 were susceptible to MERS-CoV infection, with the infected mice developing pneumonia and clinical disease accompanied by histopathological changes Citation[126]. Nevertheless, a better transgenic mouse model with the gene encoding human DPP4 integrated into its genome has been developed by several laboratories Citation[127].
Conclusions
Based on the previous experience in developing SARS-CoV vaccines, current advancements and potential strategies in the development of MERS-CoV vaccines have been discussed in this review. Apart from inactivated and live-attenuated virus vaccines, DNA- and VLP-based vaccines, particularly subunit vaccines containing the RBD of MERS-CoV S protein, are critically important. Additionally, suitable adjuvant formulation and appropriate administration routes may need to be further considered to enhance the immunogenicity of the vaccine candidates. Furthermore, development of effective animal models for MERS-CoV infection is important for evaluating the efficacy of MERS-CoV candidate vaccines. Taken together, the strategies discussed above will have important implications for the development of effective MERS vaccines in the future.
Expert commentary
MERS-CoV, as a newly emerging CoV, has raised worldwide concerns based on its ability to cause increasing number of human infections with limited human-to-human transmission, making it particularly important to explore efficient and safe measures for the control of the disease. Among these, vaccination is considered one of the most effective strategies to prevent virus infection. Different from other structural proteins (E, M and N), S protein of MERS-CoV plays a key role in virus infection via S1 binding to viral receptor, followed by subsequent virus and cell membrane fusion via S2. DPP4 was identified to be the cellular receptor of MERS-CoV. Subsequently, the viral RBD was mapped by crystal structural analysis into residues 367–588 and 367–606 in the S1 subunit of S protein. The truncated RBD containing residues 377–588 induced the highest neutralizing antibody response against virus infection in vaccinated animals and thus demonstrating the importance for developing S-RBD-based vaccines against MERS-CoV. Compared with other vaccine types, subunit vaccines possess a higher safety profile, having the potential to elicit stronger immune responses, including neutralizing antibodies. Suitable choices in adjuvant formulation and immunization routes can further increase the efficacy of such subunit vaccines, highlighting the need to develop MERS-CoV subunit vaccines based on the RBD of viral S protein.
Five-year view
In the next 5 years, the development of an effective and safe subunit vaccine against MERS can be expected. In particular, the development of a subunit vaccine based on the RBD of MERS-CoV S protein has the greatest potential to contain multiple neutralizing epitopes capable of inducing highly potent immune responses, particularly neutralizing activity, against MERS-CoV infection. The establishment of effective small animal models for MERS infection will allow economical and practical detection of protective efficacy of designed vaccines against challenges from both single and multiple MERS-CoV strains potentially occurring in humans. The application of similar vaccine development strategies against other CoVs that might cause human diseases in the future is also anticipated.
Key issues
First identified in humans in June 2012, Middle East respiratory syndrome (MERS) is a newly emerging infectious disease caused by a novel coronavirus, MERS-coronavirus (MERS-CoV), and is currently causing increased human infections with high mortality.
Belonging to lineage C of β-CoV, phylogenetically related to bat-HKU4 and bat-HKU5, MERS-CoV is the first known lineage C β-CoV associated with human infections.
Human dipeptidyl peptidase 4 is an identified receptor for MERS-CoV, and the receptor-binding domain (RBD) of MERS-CoV has been mapped by crystal structure to the residues covering 367–588 or 367–606, respectively.
A truncated RBD fragment containing a 212-amino acid (residues 377–588) of MERS-CoV S protein induced strong RBD-specific antibodies, blocking MERS-CoV RBD binding to viral receptor dipeptidyl peptidase 4 and effectively neutralizing MERS-CoV infection, providing promise for further development as a MERS candidate vaccine.
MERS-CoV S protein and RBD-based subunit vaccines that contain multiple neutralizing epitopes possess high efficacy to induce strong neutralizing antibody responses and protective immunity against MERS-CoV infection, representing a direction for future design of effective MERS vaccines.
Subunit vaccines formulated with suitable adjuvants and administered with appropriate routes have a high potential to enhance the immunogenicity of MERS candidate vaccines.
Acknowledgements
We thank Yang Yang at the Department of Pharmacology, University of Minnesota Medical School for the structural analysis of SARS-CoV and MERS-CoV RBDs.
Financial and competing interests disclosure
This study was supported by grants from the National Institute of Allergy and Infectious Diseases of the National Institutes of Health (AI109094). The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.
Notes
References
- Zaki AM, van Boheemen S, Bestebroer TM, et al. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 2012;367(19):1814-20
- WHO. Middle East respiratory syndrome coronavirus. (MERS-CoV) - update. Available from: www.who.int/csr/don/2014_03_26/en/ [Last accessed 26 March 2014]
- Memish ZA, Mishra N, Olival K, et al. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg Infect Dis 2013;19(11):1819-23
- Haagmans BL, Al Dhahiry SH, Reusken CB, et al. Middle East respiratory syndrome coronavirus in dromedary camels: an outbreak investigation. Lancet Infect Dis 2013;14(2):140-5
- Reusken CB, Haagmans BL, Müller MA, et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: a comparative serological study. Lancet Infect Dis 2013;13(10):859-66
- Balkhair A, Al Maamari K, Alawi FB. The struggle against MERS-CoV (the novel coronavirus). Oman Med J 2013;28(4):226-7
- Memish ZA, Zumla AI, Al-Hakeem RF, et al. Family cluster of Middle East respiratory syndrome coronavirus infections. N Engl J Med 2013;368:2487-94
- Mailles A, Blanckaert K, Chaud P, et al. First cases of Middle East respiratory syndrome coronavirus (MERS-CoV) infections in France, investigations and implications for the prevention of human-to-human transmission, France, May 2013. Euro Surveill 2013;18(24):pii. 20502
- Guery B, Poissy J, el Mansouf L, et al. Clinical features and viral diagnosis of two cases of infection with Middle East Respiratory Syndrome coronavirus: a report of nosocomial transmission. Lancet 2013;381(9885):2265-72
- Morens DM, Folkers GK, Fauci AS. The challenge of emerging and re-emerging infectious diseases. Nature 2004;430(6996):242-9
- McBride R, Fielding BC. The role of Severe Acute Respiratory Syndrome (SARS)-coronavirus accessory proteins in virus pathogenesis. Viruses 2012;4(11):2902-23
- Müller MA, Raj VS, Muth D, et al. Human coronavirus EMC does not require the SARS-coronavirus receptor and maintains broad replicative capability in mammalian cell lines. MBio 2012;3:6
- Woo PC, Lau SK, Lam CS, et al. Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J Virol 2012;86(7):3995-4008
- Fouchier RA, Hartwig NG, Bestebroer TM, et al. A previously undescribed coronavirus associated with respiratory disease in humans. Proc Natl Acad Sci USA 2004;101(16):6212-16
- Lau SK, Woo PC, Yip CC, et al. Coronavirus HKU1 and other coronavirus infections in Hong Kong. J Clin Microbiol 2006;44(6):2063-71
- Woo PC, Lau SK, Chu CM, et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol 2005;79(2):884-95
- Zhong N, Zheng B, Li Y, et al. Epidemiology and cause of severe acute respiratory syndrome (SARS) in Guangdong, People’s Republic of China, in February, 2003. Lancet 2003;362(9393):1353-8
- Chan JF, Lau SK, Woo PC. The emerging novel Middle East respiratory syndrome coronavirus: the “knowns” and “unknowns”. J Formos Med Assoc 2013;112(7):372-81
- Woo PC, Lau SK, Li KS, et al. Molecular diversity of coronaviruses in bats. Virology 2006;351(1):180-7
- Woo PC, Wang M, Lau SK, et al. Comparative analysis of twelve genomes of three novel group 2c and group 2d coronaviruses reveals unique group and subgroup features. J Virol 2007;81(4):1574-85
- Lau SK, Woo PC, Li KS, et al. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci USA 2005;102(39):14040-5
- Niemeyer D, Zillinger T, Muth D, et al. Middle East respiratory syndrome coronavirus accessory protein 4a is a type I interferon antagonist. J Virol 2013;87(22):12489-95
- Scobey T, Yount BL, Sims AC, et al. Reverse genetics with a full-length infectious cDNA of the Middle East respiratory syndrome coronavirus. Proc Natl Acad Sci USA 2013;110(40):16157-62
- Hurst KR, Ye R, Goebel SJ, et al. An interaction between the nucleocapsid protein and a component of the replicase-transcriptase complex is crucial for the infectivity of coronavirus genomic RNA. J Virol 2010;84(19):10276-88
- Xu Y, Lou Z, Liu Y, et al. Crystal structure of severe acute respiratory syndrome coronavirus spike protein fusion core. J Biol Chem 2004;279(47):49414-19
- Bonavia A, Zelus BD, Wentworth DE, et al. Identification of a receptor-binding domain of the spike glycoprotein of human coronavirus HCoV-229E. J Virol 2003;77(4):2530-8
- Taguchi F. The S2 subunit of the murine coronavirus spike protein is not involved in receptor binding. J Virol 1995;69(11):7260-3
- Wu K, Li W, Peng G, Li F. Crystal structure of NL63 respiratory coronavirus receptor-binding domain complexed with its human receptor. Proc Natl Acad Sci USA 2009;106(47):19970-4
- Peng G, Sun D, Rajashankar KR, et al. Crystal structure of mouse coronavirus receptor-binding domain complexed with its murine receptor. Proc Natl Acad Sci USA 2011;108(26):10696-701
- Zhu X, Liu Q, Du L, et al. Receptor-binding domain as a target for developing SARS vaccines. J Thorac Dis 2013;5(Suppl 2):S142
- Li F, Li W, Farzan M, Harrison SC. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005;309(5742):1864-8
- Wang N, Shi X, Jiang L, et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res 2013;23(8):986-93
- Hofmann H, Pyrc K, van der Hoek L, et al. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci USA 2005;102(22):7988-93
- Li W, Moore MJ, Vasilieva N, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003;426(6965):450-4
- Raj VS, Mou H, Smits SL, et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013;495(7440):251-4
- Chen Y, Rajashankar KR, Yang Y, et al. Crystal structure of the receptor-binding domain from newly emerged Middle East respiratory syndrome coronavirus. J Virol 2013;87(19):10777-83
- Lu G, Hu Y, Wang Q, et al. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 2013;500(7461):227-31
- Gao J, Lu G, Qi J, et al. Structure of the fusion core and inhibition of fusion by a heptad-repeat peptide derived from the S protein of MERS-CoV. J Virol 2013;87(24):13134-40
- Lu L, Liu Q, Zhu Y, et al. Structure-based discovery of Middle East respiratory syndrome coronavirus (MERS-CoV) fusion inhibitor. Nat Commun 2014;5:3067
- Pak JE, Sharon C, Satkunarajah M, et al. Structural insights into immune recognition of the severe acute respiratory syndrome coronavirus S protein receptor binding domain. J Mol Biol 2009;388(4):815-23
- Prabakaran P, Gan J, Feng Y, et al. Structure of severe acute respiratory syndrome coronavirus receptor-binding domain complexed with neutralizing antibody. J Biol Chem 2006;281(23):15829-36
- Hwang WC, Lin Y, Santelli E, et al. Structural basis of neutralization by a human anti-severe acute respiratory syndrome spike protein antibody. J Biol Chem 2006;281(45):34610-16
- Du L, He Y, Zhou Y, et al.The spike protein of SARS-CoV—a target for vaccine and therapeutic development. Nature Rev Microbiol 2009;7(3):226-36
- Graham RL, Donaldson EF, Baric RS. A decade after SARS: strategies for controlling emerging coronaviruses. Nature Rev Microbiol 2013;11(12):836-48
- Jiang S, Bottazzi ME, Du L, et al. Roadmap to developing a recombinant coronavirus S protein receptor-binding domain vaccine for severe acute respiratory syndrome. Expert Rev Vaccines 2012;11(12):1405-13
- Zakhartchouk AN, Sharon C, Satkunarajah M, et al. Immunogenicity of a receptor-binding domain of SARS coronavirus spike protein in mice: implications for a subunit vaccine. Vaccine 2007;25(1):136-43
- Almazán F, DeDiego ML, Sola I, et al. Engineering a replication-competent, propagation-defective Middle East respiratory syndrome coronavirus as a vaccine candidate. mBio 2013;4(5):e00650-13
- Cao Z, Liu L, Du L, et al. Potent and persistent antibody responses against the receptor-binding domain of SARS-CoV spike protein in recovered patients. Virol J 2010;7(1):299
- Du L, Zhao G, Chan C, et al. Recombinant receptor-binding domain of SARS-CoV spike protein expressed in mammalian, insect and E. coli cells elicits potent neutralizing antibody and protective immunity. Virology 2009;393(1):144-50
- Du L, Zhao G, He Y, et al. Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model. Vaccine 2007;25(15):2832-8
- He Y, Lu H, Siddiqui P, et al. Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J Immunol 2005;174(8):4908-15
- Song F, Fux R, Provacia LB, et al. Middle East respiratory syndrome coronavirus spike protein delivered by modified vaccinia virus Ankara efficiently induces virus-neutralizing antibodies. J Virol 2013;87(21):11950-4
- Czub M, Weingartl H, Czub S, et al. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine 2005;23(17):2273-9
- Olsen CW. A review of feline infectious peritonitis virus: molecular biology, immunopathogenesis, clinical aspects, and vaccination. Vet Microbiol 1993;36(1):1-37
- Mou H, Raj VS, van Kuppeveld FJ, et al. The receptor binding domain of the new Middle East respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J Virol 2013;87(16):9379-83
- Du L, Zhao G, Kou Z, et al. Identification of a receptor-binding domain in the S protein of the novel human coronavirus Middle East respiratory syndrome coronavirus as an essential target for vaccine development. J Virol 2013;87(17):9939-42
- Du L, Kou Z, Ma C, et al. A truncated receptor-binding domain of MERS-CoV spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: implication for developing therapeutics and vaccines. PLoS One 2013;8(12):e81587
- Yang Y, Zhang L, Geng H, et al. The structural and accessory proteins M, ORF 4a, ORF 4b, and ORF 5 of Middle East respiratory syndrome coronavirus (MERS-CoV) are potent interferon antagonists. Protein Cell 2013;4(12):951-61
- Zhou J, Wang W, Zhong Q, et al. Immunogenicity, safety, and protective efficacy of an inactivated SARS-associated coronavirus vaccine in rhesus monkeys. Vaccine 2005;23(24):3202-9
- Kong WP, Xu L, Stadler K, et al. Modulation of the immune response to the severe acute respiratory syndrome spike glycoprotein by gene-based and inactivated virus immunization. J Virol 2005;79(22):13915-23
- Lamirande EW, DeDiego ML, Roberts A, et al. A live attenuated severe acute respiratory syndrome coronavirus is immunogenic and efficacious in golden Syrian hamsters. J Virol 2008;82(15):7721-4
- See RH, Zakhartchouk AN, Petric M, et al. Comparative evaluation of two severe acute respiratory syndrome (SARS) vaccine candidates in mice challenged with SARS coronavirus. J Gen Virol 2006;87(3):641-50
- Graham RL, Becker MM, Eckerle LD, et al. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nat Med 2012;18(12):1820-6
- Qu D, Zheng B, Yao X, et al. Intranasal immunization with inactivated SARS-CoV (SARS-associated coronavirus) induced local and serum antibodies in mice. Vaccine 2005;23(7):924-31
- Netland J, DeDiego ML, Zhao J, et al. Immunization with an attenuated severe acute respiratory syndrome coronavirus deleted in E protein protects against lethal respiratory disease. Virology 2010;399(1):120-8
- Fett C, DeDiego ML, Regla-Nava JA, et al. Complete protection against severe acute respiratory syndrome coronavirus-mediated lethal respiratory disease in aged mice by immunization with a mouse-adapted virus lacking E protein. J Virol 2013;87(12):6551-9
- Kim JH, Jacob J. DNA vaccines against influenza viruses. Curr Top Microbiol Immunol 2009;333:197-210
- Khalil IA, Kogure K, Akita H, Harashima H. Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev 2006;58(1):32-45
- Šatkauskas S, Bureau MF, Mahfoudi A, Mir LM. Slow accumulation of plasmid in muscle cells: supporting evidence for a mechanism of DNA uptake by receptor-mediated endocytosis. Mol Ther 2001;4(4):317-23
- Shamlou PA. Scaleable processes for the manufacture of therapeutic quantities of plasmid DNA. Biotechnol Appl Biochem 2003;37(3):207-18
- Prather KJ, Sagar S, Murphy J, Chartrain M. Industrial scale production of plasmid DNA for vaccine and gene therapy: plasmid design, production, and purification. Enzyme Microb Technol 2003;33(7):865-83
- Hoare M, Levy MS, Bracewell DG, et al. Bioprocess engineering issues that would be faced in producing a DNA vaccine at up to 100 m3 fermentation scale for an influenza pandemic. Biotechnol Prog 2005;21(6):1577-92
- Yang ZY, Kong WP, Huang Y, et al. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature 2004;428(6982):561-4
- Zhao B, Jin NY, Wang RL, et al. Immunization of mice with a DNA vaccine based on severe acute respiratory syndrome coronavirus spike protein fragment 1. Viral Immunol 2006;19(3):518-24
- Shi SQ, Peng JP, Li YC, et al. The expression of membrane protein augments the specific responses induced by SARS-CoV nucleocapsid DNA immunization. Mol Immunol 2006;43(11):1791-8
- Roberts A, Lamirande EW, Vogel L, et al. Animal models and vaccines for SARS-CoV infection. Virus Res 2008;133(1):20-32
- Woo PC, Lau SK, Tsoi HW, et al. SARS coronavirus spike polypeptide DNA vaccine priming with recombinant spike polypeptide from Escherichia coli as booster induces high titer of neutralizing antibody against SARS coronavirus. Vaccine 2005;23(42):4959-68
- Bisht H, Roberts A, Vogel L, et al. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc Natl Acad Sci USA 2004;101(17):6641-6
- Bukreyev A, Lamirande EW, Buchholz UJ, et al. Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. Lancet 2004;363(9427):2122-7
- Gao W, Tamin A, Soloff A, et al. Effects of a SARS-associated coronavirus vaccine in monkeys. Lancet 2003;362(9399):1895-6
- Du L, Zhao G, Lin Y, et al. Intranasal vaccination of recombinant adeno-associated virus encoding receptor-binding domain of severe acute respiratory syndrome coronavirus (SARS-CoV) spike protein induces strong mucosal immune responses and provides long-term protection against SARS-CoV infection. J Immunol 2008;180(2):948-56
- Du L, Zhao G, Lin Y, et al. Priming with rAAV encoding RBD of SARS-CoV S protein and boosting with RBD-specific peptides for T cell epitopes elevated humoral and cellular immune responses against SARS-CoV infection. Vaccine 2008;26(13):1644-51
- Pichla-Gollon SL, Lin S-W, Hensley SE, et al. Effect of preexisting immunity on an adenovirus vaccine vector: in vitro neutralization assays fail to predict inhibition by antiviral antibody in vivo. J Virol 2009;83(11):5567-73
- Gao G, Wang Q, Dai Z, et al. Adenovirus-based vaccines generate cytotoxic T lymphocytes to epitopes of NS1 from dengue virus that are present in all major serotypes. Hum Gene Ther 2008;19(9):927-36
- Weingartl H, Czub M, Czub S, et al. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J Virol 2004;78(22):12672-6
- Lu X, Chen Y, Bai B, et al. Immune responses against severe acute respiratory syndrome coronavirus induced by virus‐like particles in mice. Immunology 2007;122(4):496-502
- Liu YV, Massare MJ, Barnard DL, et al. Chimeric severe acute respiratory syndrome coronavirus (SARS-CoV) S glycoprotein and influenza matrix 1 efficiently form virus-like particles (VLPs) that protect mice against challenge with SARS-CoV. Vaccine 2011;29(38):6606-13
- Lokugamage KG, Yoshikawa-Iwata N, Ito N, et al. Chimeric coronavirus-like particles carrying severe acute respiratory syndrome coronavirus (SCoV) S protein protect mice against challenge with SCoV. Vaccine 2008;26(6):797-808
- Ho Y, Lin PH, Liu CY, et al. Assembly of human severe acute respiratory syndrome coronavirus-like particles. Biochem Biophys Res Commun 2004;318(4):833-8
- Sailaja G, Skountzou I, Quan FS, et al. Human immunodeficiency virus-like particles activate multiple types of immune cells. Virology 2007;362(2):331-41
- Wang BZ, Quan FS, Kang SM, et al. Incorporation of membrane-anchored flagellin into influenza virus-like particles enhances the breadth of immune responses. J Virol 2008;82(23):11813-23
- He Y, Zhou Y, Wu H, et al. Identification of immunodominant sites on the spike protein of severe acute respiratory syndrome (SARS) coronavirus: implication for developing SARS diagnostics and vaccines. J Immunol 2004;173(6):4050-7
- Ma C, Li Y, Wang L, et al. Intranasal vaccination with recombinant receptor-binding domain of MERS-CoV spike protein induces much stronger local mucosal immune responses than subcutaneous immunization: implication for designing novel mucosal MERS vaccines. Vaccine 2014;32(18):2100-8
- Agnihothram S, Gopal R, Yount BL, et al. Evaluation of serologic and antigenic relationships between middle eastern respiratory syndrome coronavirus and other coronaviruses to develop vaccine platforms for the rapid response to emerging coronaviruses. J Infect Dis 2014;209(7):995-1006
- Naz RK, Dabir P. Peptide vaccines against cancer, infectious diseases, and conception. Front Biosci 2006;12:1833-44
- Reed SG, Bertholet S, Coler RN, Friede M. New horizons in adjuvants for vaccine development. Trends Immunol 2009;30(1):23-32
- Marciani DJ. Vaccine adjuvants: role and mechanisms of action in vaccine immunogenicity. Drug Discov Today 2003;8(20):934-43
- Lambert SL, Yang CF, Liu Z, et al. Molecular and cellular response profiles induced by the TLR4 agonist-based adjuvant glucopyranosyl Lipid A. PLoS One 2012;7(12):e51618
- Podda A, Del Giudice G. MF59-adjuvanted vaccines: increased immunogenicity with an optimal safety profile. Expert Rev Vaccines 2003;2(2):197-204
- Radošević K, Rodriguez A, Mintardjo R, et al. Antibody and T-cell responses to a virosomal adjuvanted H9N2 avian influenza vaccine: impact of distinct additional adjuvants. Vaccine 2008;26(29):3640-6
- Baldwin SL, Bertholet S, Reese VA, et al. The importance of adjuvant formulation in the development of a tuberculosis vaccine. J Immunol 2012;188(5):2189-97
- Filipazzi P, Pilla L, Mariani L, et al. Limited induction of tumor cross-reactive T cells without a measurable clinical benefit in early melanoma patients vaccinated with human leukocyte antigen class I–modified peptides. Clin Cancer Res 2012;18(23):6485-96
- Mavroudis D, Bolonakis I, Cornet S, et al. A phase I study of the optimized cryptic peptide TERT572Y in patients with advanced malignancies. Oncology 2006;70(4):306-14
- Du L, Zhao G, Sun S, et al. A critical HA1 neutralizing domain of H5N1 influenza in an optimal conformation induces strong cross-protection. PLoS One 2013;8(1):e53568
- Budimir N, Haan A, Meijerhof T, et al. Heterosubtypic cross‐protection induced by whole inactivated influenza virus vaccine in mice: influence of the route of vaccine administration. Influenza Other Respir Viruses 2013;7(6):1202-9
- Frenck RW Jr, Belshe R, Brady RC, et al. Comparison of the immunogenicity and safety of a split-virion, inactivated, trivalent influenza vaccine (Fluzone®) administered by intradermal and intramuscular route in healthy adults. Vaccine 2011;29(34):5666-74
- Rahman F, Dahmen A, Herzog‐Hauff S, et al. Cellular and humoral immune responses induced by intradermal or intramuscular vaccination with the major hepatitis B surface antigen. Hepatology 2000;31(2):521-7
- Mooney AJ, Li Z, Gabbard JD, et al. Recombinant parainfluenza virus 5 vaccine encoding the influenza virus hemagglutinin protects against H5N1 highly pathogenic avian influenza virus infection following intranasal or intramuscular vaccination of BALB/c mice. J Virol 2013;87(1):363-71
- Siu Y, Teoh K, Lo J, et al. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J Virol 2008;82(22):11318-30
- Kitabatake M, Inoue S, Yasui F, et al. SARS-CoV spike protein-expressing recombinant vaccinia virus efficiently induces neutralizing antibodies in rabbits pre-immunized with vaccinia virus. Vaccine 2007;25(4):630-7
- Takasuka N, Fujii H, Takahashi Y, et al. A subcutaneously injected UV-inactivated SARS coronavirus vaccine elicits systemic humoral immunity in mice. Int Immunol 2004;16(10):1423-30
- Du L, Zhao G, Li L, et al. Antigenicity and immunogenicity of SARS-CoV S protein receptor-binding domain stably expressed in CHO cells. Biochem Biophys Res Commun 2009;384(4):486-90
- Khosrow Tayebati S, Ejike Nwankwo I, Amenta F. Intranasal drug delivery to the central nervous system: present status and future outlook. Curr Pharm Des 2013;19(3):510-26
- CDC. Seasonal Influenza (Flu). Available from: www.cdc.gov/flu/protect/whoshouldvax.htm [Last accessed on 24 March 2014]
- Liew FY, Russell SM, Appleyard G, et al. Cross‐protection in mice infected with influenza A virus by the respiratory route is correlated with local IgA antibody rather than serum antibody or cytotoxic T cell reactivity. Eur J Immunol 1984;14(4):350-6
- Kuiken T, Fouchier RA, Schutten M, et al. Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet 2003;362(9380):263-70
- Martina BE, Haagmans BL, Kuiken T, et al. Virology: SARS virus infection of cats and ferrets. Nature 2003;425(6961):915-15
- Roberts A, Vogel L, Guarner J, et al. Severe acute respiratory syndrome coronavirus infection of golden Syrian hamsters. J Virol 2005;79(1):503-11
- de Wit E, Prescott J, Baseler L, et al. The Middle East respiratory syndrome coronavirus (MERS-CoV) does not replicate in Syrian hamsters. PLoS One 2013;8(7):e69127
- Yao Y, Bao L, Deng W, et al. An animal model of MERS produced by infection of Rhesus macaques with MERS coronavirus. J Infect Dis 2014;209(2):236-42
- Coleman CM, Matthews KL, Goicochea L, Frieman MB. Wild-type and innate immune-deficient mice are not susceptible to the Middle East respiratory syndrome coronavirus. J Gen Virol 2014;95(Pt 2):408-12
- Falzarano D, de Wit E, Rasmussen AL, et al. Treatment with interferon-[alpha] 2b and ribavirin improves outcome in MERS-CoV-infected rhesus macaques. Nat Med 2013;19(10):1313-17
- Munster VJ, de Wit E, Feldmann H. Pneumonia from human coronavirus in a macaque model. N Engl J Med 2013;368(16):1560-2
- de Wit E, Rasmussen AL, Falzarano D, et al. Middle East respiratory syndrome coronavirus (MERS-CoV) causes transient lower respiratory tract infection in rhesus macaques. Proc Natl Acad Sci USA 2013;110(41):16598-603
- Devitt E. Lack of small animal model hinders MERS coronavirus research. Nat Med 2013;19(8):952-2
- Zhao J, Li K, Wohlford-Lenane C, et al. Rapid generation of a mouse model for Middle East respiratory syndrome. Proc Natl Acad Sci USA 2014;111(13):4970-5
- Reardon S. Nature news. Available from: www.nature.com/news/biologists-make-first-mouse-model-for-mers-1.14634 [Last accessed on 22 March 2014]
- Narayanan K, Maeda A, Maeda J, Makino S. Characterization of the coronavirus M protein and nucleocapsid interaction in infected cells. J Virol 2000;74(17):8127-34