ABSTRACT
We analyzed lichen species composition and biomass in 815 plots on 163 sites across wild reindeer regions in Norway, ranging from ranges with a long history of very low grazing pressure to heavily grazed sites. Reindeer density (1974–2000) and lichen biomass were well correlated for sites with comparable snow cover, altitude and terrain (R 2 = 0.81, P = 0.006, n = 12). Absence of grazing for potentially several centuries has virtually resulted in a monoculture consisting of Cladina stellaris, Flavocetraria nivalis, and Alectoria ochroleuca (Syn. Bryocaulon ochroleuca). Light grazing in terms of 20 to 30% removal of initial lichen cover easily eroded Cladina stellaris from exposed ridges by cratering and trampling by reindeer through the snow, while Flavocetraria nivalis persisted longer. This decline in lichen cover observed along a historic grazing gradient further resulted in increasing cover of bare ground, but less than expected from lichen removal due to gradual colonization of other species, such as mosses (incl. Polytrichum piliferum), crustose and fruticose lichens, dwarf shrubs (Arctostaphylos spp., Empetrum nigrum, Loiseleuria procumbens), and graminoids, particularly rushes (Juncus trifidus). Moderate grazing may thus increase plant diversity on ridges compared to ungrazed lands, and hence strongly influence gradients in biomass, composition and abundance of ridge communities across the landscape.
Introduction
Herbivores are well known to influence plant succession patterns, nutrient cycling, and vegetation community structures (CitationOksanen and Oksanen, 1989; CitationOlofsson et al., 2002, Citation2004a, Citation2004b, Citation2005; CitationVirtanen et al., 2006). There are numerous studies on the role of ungulate grazing for vegetation composition (i.e., CitationManseau et al., 1996; CitationAugustine and McNaughton, 1998; CitationBråthen and Oksanen, 2001; Citationvan der Wal, 2006). In arctic and alpine environments, where diversity of ungulates is much lower than in tropical ecosystems, permafrost, geomorphology and the short growing season provide highly different conditions for plant growth and colonization of disturbed or heavily grazed areas than in more temperate and tropical regions (CitationForbes and Jefferies, 1999; Citationvan der Wal et al., 2001a, Citation2001b).
Ground lichens comprise a primary food source for reindeer and caribou (Rangifer tarandus) in alpine regions, along with grasses, sedges, herbs, and dwarf shrubs (CitationBergerud, 1974; CitationGaare and Skogland, 1975; CitationHelle, 1984; CitationDanell et al., 1994; CitationParker et al., 2005). Due to their slow growth, lichens can easily be depleted by intensive grazing at high Rangifer densities (CitationSkogland, 1985; CitationOuellet et al., 1993; CitationJefferies et al., 1994; CitationBoudreau and Payette, 2004a; CitationMysterud, 2006). Depletion of lichens following an increase in reindeer or caribou populations has been recorded throughout the circumpolar North, such as in Norway (CitationTveitnes, 1980; CitationSkogland, 1988; CitationGaare, 1995; CitationJohansen and Karlsen, 2005), West-Greenland (CitationStaaland and Olesen, 1992), eastern Canada (CitationCouturier et al., 1990; CitationManseau et al., 1996), Russia (CitationAhti and Oksanen, 1990; CitationVilchek, 1997; CitationCrittenden, 2000), and in particular in the extreme situations of introduced caribou herds to islands without a previous history of grazing (CitationKlein, 1968, Citation1987; CitationHeard and Ouellet, 1994).
A depletion of the lichen cover has often been observed in conjunction with increased vulnerability to icing conditions, reduced body weights of Rangifer, as well as increased mortality rates and low calf production (CitationMessier et al., 1988; CitationJohansen and Karlsen, 2005). Although reindeer and caribou can adjust to lichen-free diets (CitationLeader-Williams, 1988; CitationMathiesen et al., 1999), lichens are a vital carbohydrate source for most Rangifer in winter.
The abundance of sites available for grazing in winter, and thus the availability of lichens and food plants, depends mainly on snow conditions (CitationPruitt, 1959; CitationSkogland, 1978; CitationHelle, 1984; CitationAdamczewski et al., 1988) and terrain ruggedness (CitationNellemann, 1996). Reindeer will dig craters on exposed ridges with shallow snow cover to access the lichens. In this cratering process, some lichens are kicked loose in the snow, while others are more resistant to trampling and are grazed at the bottom of the craters. The composition of lichens will therefore provide an indication of both grazing pressure and of the sensitivity of lichens to cratering (CitationNellemann et al., 2000). This is especially true in tundra and coastal regions, where strong winds and freeze-thaw processes pack the snow and form ice layers, but is also important in inland and alpine areas. Lichens may in particular constitute a large proportion of the diet in late winter, when windblown ridges support the only readily available forage (CitationBergerud, 1974; CitationGaare and Skogland, 1975; CitationMathiesen et al., 2000).
The decisions of where to eat, when to eat and what to eat are complex functions of factors such as the nutritional demands of the animal, the availability and quality of forage, cost of foraging, predator avoidance, terrain preferences, and environmental changes (CitationSchluter, 1981; CitationMangel and Clark, 1986; CitationJohnson et al., 2001). Reindeer and caribou can adapt to a variety of environmental conditions and landscapes using different foraging strategies (CitationJohnson et al., 2001), and are often classified as intermediate mixed feeders based on characteristics of their digestive system (CitationMathiesen, 1999; CitationHofmann, 2000; CitationClauss et al., 2006; but see also CitationKlein, 1985 and CitationStaaland and White, 1991). CitationJohnson et al. (2001) found that woodland caribou selected sites with shallow snow and a relatively high abundance of specific lichen species. Choices varied somewhat according to the scale studied; at the finest scale measured, reindeer chose between different lichen species present.
Rangifer lichen preferences have also been studied under more controlled conditions through numerous rumen content and fecal analyses, as well as in cafeteria trials (CitationGaare and Skogland, 1975; CitationHolleman and Luick, 1977; CitationDanell et al., 1994; CitationMathiesen et al., 2000). Several studies have assessed impacts of reindeer grazing on ground cover vegetation using remote sensing (CitationVäre et al., 1996; CitationColpaert et al., 2003; CitationJohansen and Karlsen, 2005), comparing vegetation in enclosures with grazed areas (CitationLeader-Williams et al., 1987; CitationVäre et al., 1995; Citationden Herder et al., 2003) or by general vegetation surveys (CitationTømmervik et al., 2004), focusing on the increase of plant communities such as shrubs and grasses, as well as bare soil. Several studies have addressed vegetation changes in boreal forests and low-alpine zones (CitationAustrheim and Eriksson, 2001; Citationden Herder et al., 2003; CitationBoudreau and Payette, 2004b; CitationEskelinen and Oksanen, 2006), but few studies have assessed vegetation transitions with different grazing pressure on arctic and alpine ridges from historic ungrazed to heavily grazed locations (CitationHenry and Gunn, 1991; Citationvan der Wal et al., 2001a, Citation2001b). Such ridges are of particular interest in range ecology, being the only sites available to arctic ungulates in late winter; as well as representing severe growth conditions in relation to drainage and exposure. Several studies have documented changes in vegetation composition in boreal forests or in tundra following disturbance or grazing (i.e., CitationForbes and Sumina, 1999; CitationOlofsson et al., 2005), showing changes in lichen composition as well as in colonization rates of bryophytes and graminoids (CitationVäre et al., 1995; CitationVirtanen et al., 1997).
Around 1900, wild reindeer were near absent in Norway due to hunting, except for the herd in Snøhetta and northern Rondane. Reindeer in southern Rondane were first reintroduced in 1972 (CitationJordhøy and Skogland, 1985). This means that some of the ranges investigated here have had little or no grazing for around a century (CitationDahl, 1956). Other ranges have been exposed to successive periods of heavy grazing and depletion of the lichen layer since the 1960s (CitationGaare, 1968; CitationSkogland, 1986), hence enabling us to address the entire grazing gradient as it has evolved through the century. Herein, we analyze the composition and colonization of vegetation on ridges with comparable altitude and snow conditions along a grazing gradient.
Methods
Study Area
In south-central Norway, the original wild reindeer regions have been fragmented into some 23 smaller, largely disconnected regions holding separate populations with modest or no interaction. The fragmentation has also resulted in a disproportionate distribution in the availability of summer and winter ranges among the herds (CitationSkogland, 1983). Wild reindeer numbers are monitored by aerial and ground surveys, and regulated through organized hunting in fall. Predation pressure is very low in the study area, predators being limited to a few wolverines (Gulo gulo) and golden eagles (Aquila chrysaetos; CitationMinistry of the Environment, 2003; CitationSæther et al., 2003).
The study was conducted in four wild reindeer regions; Rondane (58 sites, 290 plots), Snøhetta (59 sites, 295 plots), Nord-Ottadalen (21 sites, 105 plots) and Nordfjella (25 sites, 125 plots; , ). There were differences in reindeer densities both within the ranges as well as across them, partly resulting from varying degrees of human activity within the ranges, where areas within 2 to 5 km of resorts and major traffic corridors are used much less by wild reindeer compared to areas away from human activity (CitationNellemann et al., 2000; Citation2001; CitationVistnes et al., 2001; Citation2004). The ranges have also historically been exposed to different grazing pressures as mentioned above. Annual precipitation ranged from 500 to 2500 mm, being highest in the west. We studied reindeer winter grazing grounds between 1000 and 1500 m a.s.l. (in data analyses further restricted to 1100–1400 m a.s.l.) on windblown ridges above the treeline. Vegetation on ridges was dominated by lichen species, but included also occasional grasses, sedges (Carex spp., Kobresia myosuroides, Luzula spp.), rushes (Juncus trifidus), mosses (Racomitrium lanuginosum and Polytrichum piliferum), and dwarf shrubs (Arctostaphylos spp., Loiseleuria procumbens, Betula nana and Empetrum nigrum). Lichens included Cladina stellaris, Flavocetraria nivalis, Alectoria ochroleuca (Syn. Bryocaulon ochroleuca), and to a lesser extent Alectoria nigricans, Bryocaulon divergens, Flavocetraria cucullata, and Thamnolia vermicularis. Lichens on rocks included Hypogymnia (syn. Parmelia) alpicola, Rhizocarpon geographicum, and Umbilicaria spp. Cladina rangiferina, Cladina mitis, Cladonia spp., Stereocaulon spp., and Cetraria spp. were common farther down the ridge, but were in general not included in this study as we focused on exposed ridges which depict grazing patterns most clearly (CitationNellemann et al., 2000).
Figure 1 Location of the study areas Rondane, Nord-Ottadalen, Snøhetta, and Nordfjella wild reindeer regions in Southern Norway.
Table 1 Distribution and time of field work (vegetation and snow registration) for 163 sites in wild reindeer winter grazing habitat.
Lichen and Snow Measurements
A total of 163 sites, each 20 × 50 m, were randomly placed within the four study areas using a 100 × 100-m grid and a random digit table. A 100 × 100-m grid was placed across a 1:50,000 map and x-y coordinates for individual sites were selected using a random digit table, where the ridge nearest the random point was used for the site. A 10 × 10-m grid covering this ridge was used to place the site along the ridge on a similar basis. Sites were restricted to exposed ridges with no or little snow in lichen heaths between 1000 and 1500 m a.s.l. (). In cases when field work was conducted in summer, sites were first marked the previous winter to relocate exposed ridges available in winter. At each site, we measured cover of major lichen species within five 1 × 1-m random plots, using a 1 × 1-m grid system covering the 20 × 50 m site, and, again, the random digit table. If in winter, snow was cautiously removed from the site using a small shovel and a soft brush, ensuring that as few vegetation fragments as possible were removed. Vegetation cover of dominant or common lichen species (Cladina stellaris, Flavocetraria nivalis, Flavocetraria cucullata, Alectoria ochroleuca, Alectoria nigricans, Bryocaulon divergens, Thamnolia vermicularis, Umbilicaria spp., and Hypogymnia alpicola were recorded by counting the number of 10 × 10-cm cells with >50% cover out of 100 cells in a 1 × 1-m vegetation frame (CitationWratten and Fry, 1980; CitationNellemann and Thomsen, 1994). We also recorded potential presence of Cladina rangiferina, Cladina mitis, Cladonia spp., Stereocaulon spp., Cetraria islandica, and Cetraria delisei, although most of these are located further down the snow gradient. Cover of Polytrichum piliferum, Racomitrium lanuginosum, Andreaea rupestris, Arctostaphylos spp., Empetrum nigrum, Loiseleuria procumbens, Betula nana, Juncus trifidus, Luzula spp., Poa spp., and Carex spp. were recorded in the same manner.
Average height of lichen was allocated to class 1 (0–1.5 cm), 2 (1.5–3.0 cm), or 3 (>3.0 cm) by measuring lichen height in five random points within the vegetation frame according to CitationNellemann et al. (2000). We calculated means from the five plots at each site. We used cover-biomass regressions for different lichen heights from Rondane (CitationNellemann et al., 2000) to assess relationships between lichen cover and biomass. Some of the lichens generally had low coverage (0–5%), but may depend upon erosion or grazing of the more dominant lichen species in order to colonize ridges. To develop a general tabular overview of the presence and absence of species at different extents of lichen cover, we divided lichen cover into four classes: >75% lichen cover; 50–74% lichen cover; 25–49% lichen cover, and 0–24% lichen cover. We then recorded the presence and absence of lichens on ridges according to this classification.
Snow depth and hardness were measured at each plot in 105 (64%) of the sites, using a ramsonde penetrometer (CitationSkogland, 1978; CitationNellemann et al., 2000). Hardness (Hr ) was calculated using the following equation: Hr = (Ph × hhd × nhd) d −1 + Q, where Ph is the mass of the load (g), hhd is the height of the hammer drop, nhd is the number of hammer drops, d is the penetration into the snow cover (cm), and Q is the total mass of the ramsonde. We calculated an integrated ram hardness index (IRH) to derive a better measure of forage availability (CitationTucker et al., 1991; CitationVistnes et al., 2001), where IRH = Hr × d. Elevation was recorded using 1:50,000-scaled maps with a contour interval of 20 m, often in combination with GPS.
To evaluate the effect on lichen erosion resulting from potential differences in the sensitivity of lichens to grazing and cratering, we investigated seven ridges in southern Rondane grazed <24 h earlier by herds dominated by bulls. This is an area with an extensive lichen cover and a long history of very low grazing intensity (CitationDahl, 1956; CitationJordhøy and Skogland, 1985). At each of the seven ridges, five craters were selected randomly located >5 m apart. Lichen cover on ground in the crater was measured using the vegetation frame as described above, while lichen fragments in snow were collected within a 2-m radius around each crater. The fragments were then sorted by species, and the volume of each species was measured in a 200-mL cup in field. For comparison purposes, volume of lichen fragments in snow and cover of lichen on ground was converted to percent of total lichen, as lichen cover is closely correlated to lichen volume and biomass (CitationNellemann, 1996; CitationNellemann et al., 2000). By using sites grazed mainly by bulls, we avoided the effect of calves and juveniles grazing extensively on lichens kicked up by older animals around craters. During late winter, the bulls, having lost their antlers and thus unable to defend craters against parturient females, tend to graze in other areas than female-calf groups (CitationEspmark, 1964; CitationThing, 1977; CitationSkogland, 1989; CitationHoland et al., 2004).
Reindeer Observations
Data on reindeer density and corresponding lichen biomass for 12 different subregions were gathered from the studies of CitationNellemann (1996), CitationNellemann et al. (2000; Citation2001), CitationJordhøy et al. 2003, and CitationVistnes et al. (2004). Here, reindeer density was estimated using systematic aerial fixed-wing or ground surveys in winter, following contiguous 4 km wide transects. Fixed-wing surveys were used in Snøhetta, Nord-Ottadalen, northern Rondane and Nordfjella, while reindeer in central Rondane were studied using ground surveys following approximately the same method as for fixed-winged surveys. All reindeer observed were photographed and located using GPS or topographic maps, and the number of reindeer determined. Surveys were conducted one day each winter for 8 yr in Nordfjella (1986–1998) and Nord-Ottadalen (1974–1985), 11 yr in northern Rondane (1987–1997), 5 yr in central Rondane (1991–1996), and 14 yr in central Snøhetta (1975–2000).
Statistical Analyses
Statistical analyses were performed in Sigmastat (CitationSPSS, 1997). Polynomial regression analysis was used to determine the correlation between snow depth, elevation, and reindeer density, respectively, versus lichen biomass. Lichen composition on recently grazed ridges was compared with lichen fragments in the snow using t-tests, except for Cetraria islandica and Alectoria ochroleuca, where a Mann-Whitney rank sum test was used since normality tests failed for these species. In all cases, P-values < 0.05 were considered statistically significant.
Results
Effects on Snow Conditions on Availability of Exposed Ridges
In 85% of sites where snow characteristics were measured, snow depths were below 5 cm, and correlation between snow depth and lichen biomass was therefore very low (R 2 = 0.020). This was mainly a result of using only windblown ridges for intensive sampling. No grazing was observed in any other terrain, where snow depths typically varied between 60 and 200 cm. Ram hardness was on average 8.2 ± 0.7 kg, and integrated ram hardness (IRH) on average 33.2 ± 4.0 kg cm on exposed ridges. All hardness values were below 28 kg (83% below 10 kg) and thus below values found on ridges preferred by reindeer (CitationSkogland, 1978), except for one site with a hardness value of 49.3 kg. This site was excluded from further analyses. Snow conditions were apparently quite similar in all sites visited, reflected also in the low variance among exposed sites.
Effects of Elevation on Lichen Biomass
Biomass was correlated to elevation when including all sites (R 2 = 0.39, P < 0.001, n = 163). To control for this, we included only sites 1100 to 1400 m a.s.l. in further analyses, where elevation only explained 18% of the variance (R 2 = 0.18, P < 0.001, n = 127). The proportion of the three major lichen species was only weakly related to elevation within sites 1100 to 1400 m a.s.l. (Flavocetraria nivalis: R 2 = 0.18, P = 0.003; Cladina stellaris: R 2 = 0.11, P = 0.027, and Alectoria ochroleuca: R 2 = 0.014, P = 0.450).
Effects of Reindeer Density on Lichen Biomass
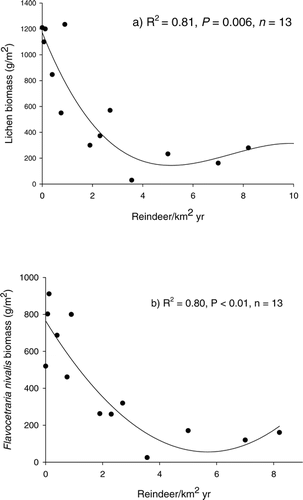
Reindeer density and lichen biomass was well correlated across the regions investigated (R 2 = 0.81; ). Biomass of Flavocetraria nivalis was correlated to reindeer density (R 2 = 0.80; P = 0.008; ). Biomass of the two other species were less closely related to reindeer density (Alectoria ochroleuca; R 2 = 0.36, P = 0.227; Cladina stellaris; R 2 = 0.54, P = 0.094).
Figure 2A–B Correlation between annual reindeer densities (reindeer km−2 yr−1): (a) total lichen biomass (g m−2) and (b) Flavocetraria nivalis. Rondane, Nord-Ottadalen, Snøhetta, and Nordfjella wild reindeer regions, Norway.
Effects of Reindeer Cratering on Lichen Composition
Lichen composition on recently grazed ridges (<24 h) differed significantly from lichen fragments left in the snow surrounding the craters for Flavocetraria nivalis and Cladina stellaris (). The volume of Flavocetraria nivalis was significantly higher on ridges compared to remaining lichen fragments in snow (P < 0.001). The volume of Cladina stellaris was significantly lower on ridges compared to the fragments found in snow (P < 0.001). There was no significant difference between the amount of Alectoria ochroleuca found on ridges and in snow (P = 0.142). This field work was conducted in an area rich on lichen (average 595 g m−2 ± 81, n = 7), corresponding to a lichen cover of >75%.
Table 2 Proportion (%) of total lichen volume in snow or lichen cover on ridge ±SE for three grazed lichen species on seven sites with a long history of low grazing pressure in Rondane, April 2006.
Test results also showed that there was a significant difference for Cetraria islandica but not for Alectoria nigricans between the proportion of lichen remnants observed in the snow versus the proportion observed on the ground. However, volumes were small for these two species which probably explains why they were not recorded on the ridge using the vegetation frame method which requires >50% coverage of at least one 10 × 10 cm cell, while they were registered in small amounts through the collection of lichen fragments in snow.
Effects of Reindeer Density on Ridge Vegetation Composition
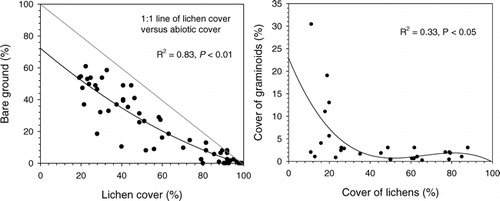
Areas with little or no record of grazing were completely dominated by 5- to 10-cm-thick mats of Cladina stellaris, Flavocetraria nivalis, and Alectoria ochroleuca, only penetrated by Betula nana (). Flavocetraria nivalis became increasingly dominant among the lichens with increasing grazing pressure, while Cladina stellaris rapidly declined even at mild grazing levels (50–74% lichen cover) becoming almost absent at a 60% lichen cover (). Further grazing reduced overall lichen cover, resulting in a gradual increase in bare ground (abiotic cover) on grazed sites (). The increase in bare ground was however 5 to 20% lower than the extent of the removed lichen cover as a result of the spreading and colonization of other species (). When investigating ridges with moderate to heavy grazing levels, other lichen species like Cladonia spp., Alectoria nigricans, Bryocaulon divergens, and Thamnolia vermicularis became present, as well as Umbilicaria spp. and Hypogymnia alpicola on small rocks formerly covered by heavy lichen mats.
Table 3 Presence and abundance of common plant species on ridges along a grazing gradient from near absence of grazing by wild reindeer (75–100% lichen cover) to heavily grazed sites (0–25% lichen cover; CitationNellemann et al., 2000, Citation2001; CitationVistnes et al., 2001, Citation2004). Wild reindeer winter habitat, Norway.
Figure 3 The relationship between total cover of lichens and decline in cover of Cladina stellaris with increasing grazing pressure. Rondane, Nord-Ottadalen, Snøhetta, and Nordfjella wild reindeer regions, Norway.
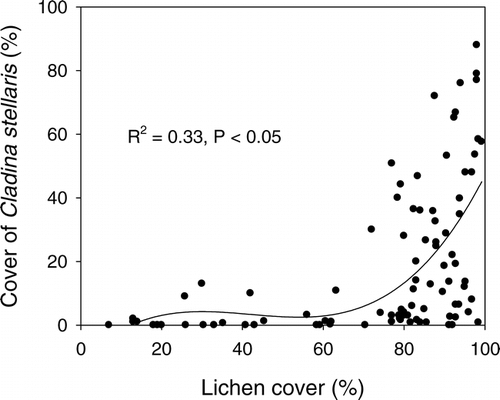
Figure 4A–B Relationship between total lichen cover: (a) cover of bare ground and (b) cover of graminoids (mainly Juncus trifidus). Rondane, Nord-Ottadalen, Snøhetta, and Nordfjella wild reindeer regions, Norway.
Discussion
Effects of Snow, Terrain, and Elevation on Lichen Distribution
Snow can pose a major barrier to reindeer food sources in winter, determining which parts of the range are available for grazing (CitationPruitt, 1959; CitationFancy and White, 1985; CitationAdamczewski et al., 1988; CitationCollins and Smith, 1991). In addition, lichen biomass and composition may vary with elevation (CitationDahl, 1956; CitationArseneau et al., 1997; CitationGrytnes et al., 1999; CitationBruun et al., 2006) and microtopography (CitationNellemann et al., 2000; CitationBruun et al., 2006). Indeed, Cladina stellaris tends to grow in areas with a slightly greater snow cover than Flavocetraria nivalis, but this is true mainly farther down the ridge where this species can be observed with other Cladina species (CitationDahl, 1956). To avoid any bias from this, we deliberately confined our sampling exclusively to lichen heath ridges with very shallow snow cover and no larger coverage of Cladina rangiferina, Cladina arbuscula, or Cladina mitis.
Given this limited range of variation in snow cover, elevation, and terrain, the second most important factor influencing lichen composition and biomass is probably grazing intensity. Optimal-foraging theory predicts that animals foraging in a patchy landscape will choose forage-rich patches, often estimated by protein or energy content, over poor patches (CitationMacArthur and Pianka, 1966; CitationBerteaux et al., 1998). Free-ranging animals are, however, also influenced by stochastically varying environments when making their foraging decisions (CitationSchluter, 1981; CitationMangel and Clark, 1986). In late winter, ridge crests may be the only vegetation type that is not completely covered with snow (CitationAdamczewski et al., 1988). Although reindeer are excellent diggers in snow, they will forage on wind-blown ridges and bluffs with softer and more shallow snow than what is average at the general site (CitationLaPerriere and Lent, 1977; CitationCollins and Smith, 1991; CitationJohnson et al., 2001) to reduce energy expenditure. Lichens may thus be almost completely removed from the top of wind-blown ridges, and still be abundant at the snow-covered sides of the ridges only a few meters away (CitationNellemann et al., 2000).
Lichens as Forage in Winter
Reducing unnecessary digging in deep or hard snow is vital during winter when forage is limited. This is particularly important as lichens are rich on readily digestible carbohydrates, but low on proteins (CitationHelle, 1984; CitationSvihus and Holand, 2000). Their content of some 2–5% crude protein (except for 7–8% in Stereocaulon spp) is below what is needed for maintenance (CitationStaaland et al., 1983; CitationGarmo, 1986; CitationNieminen and Heiskari, 1988; CitationParker et al., 2005). Even with a recycling of nitrogen in urea (CitationHove and Jacobsen, 1975) and reduced appetite in winter (CitationPösö, 2005), Rangifer will most often suffer from a protein deficiency in winter, the nitrogen level will be below the requirements of rumen bacteria, and digestibility will be reduced (CitationHuot, 1989; CitationGerhart et al., 1996). An additional negative effect is that intake of digestible carbohydrates, found in lichens, will increase the microbial nitrogen requirement and thus the loss of fecal nitrogen (CitationVan Soest, 1994). Probably to increase nitrogen intake, reindeer commonly eat a varied diet also consisting of more protein-rich grasses like Deschampsia flexuosa (CitationHelle, 1984; CitationDanell et al., 1994; CitationAagnes et al., 1995), even when there is no shortage of lichens.
Effects of Grazing and Cratering on Lichen Composition
The reduction of Cladina stellaris with increased grazing (, ) indicated that this is a species heavily used by reindeer when available, and that it is easily eroded (CitationArseneault et al., 1997; Citationden Herder et al., 2003; CitationBoudreau and Payette, 2004a). Several studies have also documented that reindeer prefer Cladina stellaris to lichen species such as Stereocaulon paschale, and prefer lichens above moss and grass (CitationHolleman and Luick, 1977; CitationDanell et al., 1994). These studies did however not include Flavocetraria nivalis. CitationStorheier et al. (2002) found a higher digestibility of Flavocetraria nivalis (70%) than of Cladina stellaris (50%), and CitationSvihus and Holand (2000) found less soluble fiber and glucose in Cladina stellaris than in Flavocetraria nivalis. Cladina stellaris however had higher total monosaccharide content than Flavocetraria nivalis (CitationSvihus and Holand, 2000).
Although our results seemed to indicate that Cladina stellaris was preferred above Flavocetraria nivalis, as the first disappeared early with grazing, this may also be explained by alternative or complimentary hypotheses. CitationGaare and Skogland (1975) found that reindeer had a very low preference of Cladina stellaris, even though it made up the largest volume of biomass removed from the ridges. They estimated a spillage factor of 10, and concluded that the considerable waste of Cladina stellaris largely must have been caused by cratering activity. Indeed, several studies have shown that Cladina stellaris is highly sensitive to grazing and is easily eroded (CitationArseneault et al., 1997; Citationden Herder et al., 2003; CitationBoudreau and Payette, 2004a). These results correspond with our comparison of lichen on ridges with lichen fragments in the snow removed through cratering, as well as the rapid decline in this species along the grazing gradient. The reason why Cladina stellaris disappears rapidly with grazing is likely caused by the different morphology of the two species. The Cladina stellaris thallus is easily removed in its entirety in big lumps, whereas the Flavocetraria nivalis thallus is more attached to the ground and is gradually cut down when grazed. It may therefore be more energy efficient for reindeer to graze lumps of Cladina stellaris than Flavocetraria nivalis. As we only included sites grazed by bulls mainly, we also avoided the effect of animals with a lower social status (such as calves) influencing the observations through secondary grazing on lichens fragments in the snow or in craters dug by older animals (CitationSkogland, 1989).
Alectoria ochroleuca is a less studied species although it often is dominant in volume. CitationArseneault et al. (1997) suggested that this species was avoided by caribou. CitationGaare (1968) observed that Alectoria ochroleuca had almost entirely disappeared from grazed sites although it was not found in rumen samples, and concluded that it must have been removed through trampling alone. Our results imply however that it also might have been grazed, as few fragments were found at newly grazed sites. It has also been documented that Alectoria ochroleuca is rich on soluble fiber and glucose (CitationSvihus and Holand, 2000). CitationOksanen and Virtanen (1995) concluded that reindeer had low preference for Alectoria ochroleuca as it holds high levels of lichenic acids, but that it still may be grazed when other species are unavailable.
Colonization and Vegetation Change on Grazed Ridges
Grazing has profound effects on the composition of lichens, but also on the composition and colonization of other species on the ridges through the removal of dominant species and the exposure of bare soil (CitationOlofsson, 2001; Citation2006). Within our study areas, some ranges have been nearly ungrazed for at least a century (CitationDahl, 1956; CitationJordhøy and Skogland, 1985), while others have experienced varying grazing pressure for millennia, also including heavy grazing for the past decades (CitationBarth 1996; CitationSkogland, 1986; CitationVistnes et al., 2004). This has resulted in a gradient from a near monoculture of Cladina stellaris, Alectoria ochroleuca, and Flavocetraria nivalis (CitationDahl, 1956) on ridges with little or no grazing, to moderately grazed ridges with higher species diversity and heavily grazed ridges exposed to erosion. The high proportion of Cladina stellaris in the snow around craters suggests high vulnerability to erosion and grazing, and was also well reflected in coverage of this species along the gradient of lichen cover. Moderate grazing by the Rondane herd is reflected in the steep decline in Cladina stellaris (CitationNellemann et al., 2000; CitationBoudreau and Payette, 2004a). In other parts of our study areas where ranges were heavily grazed in the 1960 to 1980s, Cladina stellaris is still absent from exposed ridges (CitationHenry and Gunn, 1991; CitationNellemann, 1996; CitationVistnes et al., 2004).
Most interesting however, is the transition in vegetation observed along these grazing gradients (). When lichen mats completely covered the ground, only penetrated by Betula nana (CitationOksanen and Virtanen, 1995), removal of the lichens by cratering, grazing, and trampling exposed bare ground. This bare ground, especially when reaching levels of 30 to 40% cover, was gradually colonized by mosses and crustose lichens (CitationBoudreau and Payette, 2004b). The observed 10 to 25% increase in new plant cover following removal of lichens by grazing was likely a result of lowered competition among remaining species, as well as colonization of bryophytes like Polytrichum piliferum, Racomitrium lanuginosum, and Andreaea rupestris, dwarf shrubs like Arctostaphylos spp., Empetrum nigrum, and Loiseleuria procumbens and rushes like Juncus trifidus (, ; ). The colonization of graminoids was however highly variable, and most prominent for the most heavily grazed sites, possibly being an effect of fertilization from faeces and urine given that these small narrow ridges are used repeatedly by many animals (CitationOlofsson et al., 2001, Citation2004b; CitationOlofsson, 2006). There appeared to be a slight increase in dwarf shrubs, in particular of Arctostaphylos spp. and Loiseleuria procumbens, while Empetrum nigrum did not increase very much (CitationOlofsson et al., 2005). At the most heavily grazed sites, coverage remained low also of dwarf shrubs, likely because they are frequently pulled out of the ground by reindeer when grazing or snow conditions are marginal. Here, only a few graminoids established, particularly the rush Juncus trifidus. Even mosses had low coverage when abiotic ground covered up to 90%, possibly a result of grazing and trampling (Citationvan der Wal and Brooker, 2004). Along with mosses, typically Polytrichum piliferum () and Andreaea rupestris, we also observed a colonization of moderately grazed ridges by other lichens, particularly of crustose lichens, Cladonia spp., A. nigricans, Bryocaulon divergens, and Thamnolia vermicularis, thus increasing diversity of both lichens and mosses compared to sites with low or no grazing pressure (CitationAustrheim and Eriksson, 2001; CitationOlofsson et al., 2001; Citationden Herder et al., 2003; CitationBoudreau and Payette, 2004b; CitationEskelinen and Oksanen, 2006).
Figure 5 Impacts of grazing: (a–c) near absence of grazing has led to a 5- to 15-cm-thick monoculture layer of only Cladina stellaris, Alectoria ochroleucha, and Flavocetraria nivalis; (d) moderate grazing where small “white” rocks not yet overgrown with Rhizocarpon spp. or Umbilicaria spp. suggest removal of fruticose lichens by grazing in the past couple of decades; and (e–f) high grazing pressure with a gradual colonization by crustose lichens, Polytrichum piliferum, Andreaea rupestris, Loiseleura procumbens, and a few persistent Flavocetraria nivalis.

In conclusion, moderate grazing on wind-blown ridges in winter reduced overall biomass, but increased diversity of both mosses, lichens, graminoids and dwarf shrubs, possibly by exposing bare ground, reducing competition, and increasing nitrogen turnover through more exposure to the sun and addition of feces and urine. Very heavy grazing and trampling may in a short term be beneficial to a range of colonizing species, but in the long term destroy vegetation cover and result in erosion on exposed ridges.
References Cited
- Aagnes, T. H. , W. Sørmo , and S. D. Mathiesen . 1995. Ruminal microbial digestion in free-living, in captive lichen-fed, and in starved reindeer ( Rangifer tarandus tarandus ) in winter. Applied and Environmental Microbiology 61:583–591.
- Adamczewski, J. Z. , C. C. Gates , B. M. Soutar , and R. J. Hudson . 1988. Limiting effects of snow on seasonal habitat use and diets of caribou ( Rangifer tarandus groenlandicus ) on Coats Island, Northwest Territories, Canada. Canadian Journal of Zoology 66:1986–1996.
- Ahti, T. and J. Oksanen . 1990. Epigeic lichen communities of taiga and tundra regions. Vegetatio 86:39–70.
- Arseneau, M. J. , L. Sirois , and J. P. Ouellet . 1997. Effects of altitude and tree height on the distribution and biomass of fruticose arboreal lichens in an old growth balsam fir forest. Ecoscience 4:206–213.
- Arseneault, D. , N. Villeneuve , C. Boismenu , Y. LeBlanc , and J. Deshaye . 1997. Estimating lichen biomass and caribou grazing on the wintering grounds of northern Québec: an application of fire history and Landsat data. Journal of Applied Ecology 34:65–78.
- Augustine, D. J. and S. J. McNaughton . 1998. Ungulate effects on the functional species composition of plant communities: Herbivore selectivity and plant tolerance. Journal of Wildlife Management 62:1165–1183.
- Austrheim, G. and O. Eriksson . 2001. Plant species diversity and grazing in the Scandinavian mountains—patterns and processes at different spatial scales. Ecography 24:683–695.
- Barth, E. K. 1996. Trapping systems for reindeer, ancient activities and traditions in Rondane. Technical Report 1996, Norwegian Institute for Cultural Heritage Research, Trondheim.
- Bergerud, A. T. 1974. Relative abundance of food in winter for Newfoundland caribou. Oikos 25:379–397.
- Berteaux, D. , M. Crete , J. Huot , J. Maltais , and J. P. Ouellet . 1998. Food choice by white-tailed deer n relation to protein and energy content of the diet: a field experiment. Oecologia 115:84–92.
- Boudreau, S. and S. Payette . 2004a. Growth performance of Cladina stellaris following caribou disturbance in subarctic Quebec. Écoscience 11:347–355.
- Boudreau, S. and S. Payette . 2004b. Caribou-induced changes in species dominance of lichen woodlands: An analysis of plant remains. American Journal of Botany 91:422–429.
- Bruun, H. H. , J. Moen , R. Virtanen , J. A. Grytnes , L. Oksanen , and A. Angerbjörn . 2006. Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. Journal of Vegetation Science 17:37–46.
- Bråthen, K. A. and J. Oksanen . 2001. Reindeer reduce biomass of preferred plant species. Journal of Vegetation Science 12:473–480.
- Clauss, M. , R. R. Hofmann , J. Hummel , J. Adamczewski , K. Nygren , C. Pitra , W. J. Streich , and S. Reese . 2006. Macroscopic anatomy of the omasum of free-ranging moose ( Alces alces ) and muskoxen ( Ovibos moschatus ) and a comparison of the omasal laminal surface area in 34 ruminant species. Journal of Zoology 270:346–358.
- Collins, W. B. and T. S. Smith . 1991. Effects of wind-hardened snow on foraging by reindeer ( Rangifer tarandus ). Arctic 44:217–222.
- Colpaert, A. , J. Kumpula , and M. Nieminen . 2003. Reindeer pasture biomass assessment using satellite remote sensing. Arctic 56:147–158.
- Couturier, S. , J. Brunelle , D. Vandal , and G. St-Martin . 1990. Changes in the population dynamics of the George River Caribou Herd, 1976–87. Arctic 43:9–20.
- Crittenden, P. D. 2000. Aspects of the ecology of mat-forming lichens. Rangifer 20:127–139.
- Dahl, E. 1956. Mountain vegetation in South Norway and its relation to the environment. Ph.D. dissertation, University of Oslo, Norway. Skrifter Norsk Vitenskapsakademi 3:174.
- Danell, K. , P. M. Utsi , R. T. Palo , and O. Eriksson . 1994. Food plant selection by reindeer during winter in relation to plant quality. Ecography 17:153–158.
- den Herder, M. , M. M. Kytöviita , and P. Niemelä . 2003. Growth of reindeer lichens and effects of reindeer grazing on ground cover vegetation in a Scots pine forest and a subarctic heathland in Finnish Lapland. Ecography 26:3–12.
- Eskelinen, A. and J. Oksanen . 2006. Changes in the abundance, composition and species richness of mountain vegetation in relation to summer grazing by reindeer. Journal of Vegetation Science 17:245–254.
- Espmark, Y. 1964. Studies in dominance-subordination relationship in group of semi-domestic reindeer ( Rangifer tarandus L.). Animal Behaviour 12:420–426.
- Fancy, S. G. and R. G. White . 1985. Energy expenditures by caribou while cratering in snow. Journal of Wildlife Management 49:987–993.
- Forbes, B. C. and R. L. Jefferies . 1999. Revegetation of disturbed arctic sites: constraints and applications. Biological Conservation 88:15–24.
- Forbes, B. C. and O. I. Sumina . 1999. Comparative ordination of low arctic vegetation recovering from disturbance: Reconciling two contrasting approaches for field data collection. Arctic, Antarctic, and Alpine Research 31:389–399.
- Gaare, E. 1968. A preliminary report on winter nutrition of wild reindeer in the southern Scandes, Norway. Symposium, Zoological Society London 21:109–115.
- Gaare, E. 1995. A hypothesis to explain lichen-Rangifer dynamic relationship. Rangifer 17:3–7.
- Gaare, E. and T. Skogland . 1975. Wild reindeer food habits and range use at Hardangervidda. In Wielgolaski, F. E. , editor. Fennoscandian Tundra Ecosystems. Part 2. Analysis and Synthesis. Ecological studies vol. 17 Berlin Springer. 195–205.
- Garmo, T. H. 1986. Chemical composition and in vitro dry matter digestibility of indigenous mountain pasture plants in different plant groups (Preliminary report). Rangifer 6:14–22.
- Gerhart, K. L. , R. G. White , R. D. Cameron , and D. E. Russell . 1996. Body composition and nutrient reserves of arctic caribou. Canadian Journal of Zoology 74:136–146.
- Grytnes, J. A. , H. J. B. Birks , and S. M. Peglar . 1999. Plant species richness in Fennoscandia: evaluating the relative importance of climate and history. Nordic Journal of Botany 19:489–503.
- Heard, D. C. and J. P. Ouellet . 1994. Dynamics of an introduced caribou population. Arctic 47:88–95.
- Helle, T. 1984. Foraging behavior of the semidomestic reindeer ( Rangifer tarandus L) in relation to snow in Finnish Lapland. Report from Kevo Subarctic Research Station Rovaniemi University of Oulu. 19: 35–47.
- Henry, G. H. R. and A. Gunn . 1991. Recovery of tundra vegetation after overgrazing by caribou in Arctic Canada. Arctic 44:38–42.
- Hofmann, R. R. 2000. Functional and comparative digestive system anatomy of Arctic Ungulates. In Haugerud, R. E. , editor. Key note presentations from the 10th Arctic Ungulate Conference, Tromsø 1999. Rangifer 2–3:71–82.
- Holand, O. , H. Gjostein , A. Losvar , J. Kumpula , M. E. Smith , K. H. Roed , M. Nieminen , and R. B. Weladji . 2004. Social rank in female reindeer ( Rangifer tarandus ): effects of body mass, antler size and age. Journal of Zoology 263:365–372.
- Holleman, D. F. and J. R. Luick . 1977. Lichen species preference by reindeer. Canadian Journal of Zoology 55:1368–1369.
- Hove, K. and E. Jacobsen . 1975. Renal excretion of urea in reindeer. Effect of nutrition. Acta Veterinaria Scandinavica 16:513–519.
- Huot, J. 1989. Body composition of the George River caribou ( Rangifer tarandus caribou ) in fall and late winter. Canadian Journal of Zoology 67:103–107.
- Jefferies, R. L. , D. R. Klein , and G. R. Shaver . 1994. Vertebrate herbivores and northern plant communities: reciprocal influences and responses. Oikos 71:193–206.
- Johansen, B. and S. R. Karlsen . 2005. Monitoring vegetation changes on Finnmarksvidda, Northern Norway, using Landsat MSS and Landsat TM/ETM+ satellite images. Phytocoenologia 35:969–984.
- Johnson, C. J. , K. L. Parker , and D. C. Heard . 2001. Foraging across a variable landscape: behavioral decisions made by woodland caribou at multiple spatial scales. Oecologia DOI 10.1007/s004420000573.
- Jordhøy, P. and T. Skogland . 1985. Villreinen i Rondane Sør-området. Jakt og Fiske 6:72–74.
- Jordhøy, P. , O. Strand , C. Nellemann , and I. Vistnes . 2003. Tilbakeføring av Hjerkinn til sivile formål – temautredning Økosystem: Villrein og moskus. Report to The Norwegian Defense Estates Agency 55.
- Klein, D. R. 1968. The introduction, increase, and crash of reindeer on St. Matthew Island. Journal of Wildlife Management 32:350–367.
- Klein, D. R. 1985. Population Ecology: The interaction between deer and their food supply. In Fennessy, P. and K. Drew , editors. Biology of Deer Production 13–22. Royal Society New Zealand Bulletin, 22:.
- Klein, D. R. 1987. Vegetation recovery patterns following overgrazing by reindeer on St. Matthew Island. Journal of Range Management 40:336–338.
- LaPerriere, A. and P. Lent . 1977. Caribou feeding sites in relation to snow characteristics in north-eastern Alaska. Arctic 30:101–108.
- Leader-Williams, N. 1988. Reindeer on South Georgia. The Ecology of an Introduced Population Cambridge Cambridge University Press. 319.
- Leader-Williams, N. , R. I. Lewis Smith , and P. Rothery . 1987. Influence of introduced reindeer upon the vegetation of South Georgia: Results from long term exclusion experiments. Journal of Applied Ecology 24:801–822.
- MacArthur, R. H. and E. R. Pianka . 1966. An optimal use of a patchy environment. American Naturalist 100:603–609.
- Mangel, M. and C. W. Clark . 1986. Towards a unified foraging theory. Ecology 67:1127–1138.
- Manseau, M. , J. Huot , and M. Crête . 1996. Effects of summer grazing by caribou on composition and productivity of vegetation: community and landscape level. Journal of Ecology 84:503–513.
- Mathiesen, S. D. 1999. Comparative aspects of digestion in reindeer. Dr. philos. dissertation, University of Tromsø.
- Mathiesen, S. D. , T. H. Aagnes Utsi , and W. Sørmo . 1999. Forage chemistry and the digestive system in reindeer ( Rangifer tarandus tarandus ) in northern Norway and on South Georgia. Rangifer 19:91–101.
- Mathiesen, S. D. , ØE. Haga , T. Kaino , and N. J. C. Tyler . 2000. Diet composition, rumen papillation and maintenance of carcass mass in female Norwegian reindeer ( Rangifer tarandus tarandus ) in winter. Journal of Zoology, London 251:129–138.
- Messier, F. , J. Huot , D. Le Henaff , and S. Luttich . 1988. Demography of the George River Caribou Herd: Evidence of population regulation by forage exploitation and range expansion. Arctic 41:279–287.
- Ministry of the Environment 2003. Rovvilt i norsk natur. Stortingsmelding nr. 15 (2003–2004). Oslo Ministry of the Environment. 134.
- Mysterud, A. 2006. The concept of overgrazing and its role in management of large herbivores. Wildlife Biology 12:129–141.
- Nellemann, C. 1996. Terrain selection by reindeer in late winter. Arctic 49:339–347.
- Nellemann, C. and M. G. Thomsen . 1994. Terrain ruggedness and caribou forage availability during snowmelt on the Arctic Coastal Plain, Alaska. Arctic 47:361–367.
- Nellemann, C. , P. Jordhøy , O. G. Støen , and O. Strand . 2000. Cumulative impacts of tourist resorts on wild reindeer ( Rangifer tarandus tarandus ) during winter. Arctic 53:9–16.
- Nellemann, C. , I. Vistnes , P. Jordhøy , and O. Strand . 2001. Winter distribution of wild reindeer in relation to power lines, roads and resorts. Biological Conservation 101:351–360.
- Nieminen, M. and U. Heiskari . 1988. Diets of freely grazing and captive reindeer during summer and winter. Rangifer 9:17–34.
- Oksanen, L. and T. Oksanen . 1989. Natural grazing as a factor shaping out barren landscapes. Journal of Arid Environments 17:219–233.
- Oksanen, L. and R. Virtanen . 1995. Topographic, altitudinal and regional patterns in continental and suboceanic heath vegetation of northern Fennoscandia. Acta Botanica Fennica 153:1–80.
- Olofsson, J. 2001. Influence of herbivory and abiotic factors on the distribution of tall forbs along a productivity gradient: a transplantation experiment. Oikos 94:351–357.
- Olofsson, J. 2006. Short- and long-term effects of changes in reindeer grazing pressure on tundra heath vegetation. Journal of Ecology 94:431–440.
- Olofsson, J. , H. Kitti , P. Rautiainen , S. Stark , and L. Oksanen . 2001. Effects of summer grazing by reindeer on composition of vegetation, productivity and nitrogen cycling. Ecography 24:13–24.
- Olofsson, J. , J. Moen , and L. Oksanen . 2002. Effects of herbivory on competition intensity in two arctic-alpine tundra communities with different productivity. Oikos 96:265–272.
- Olofsson, J. , P. E. Hulme , L. Oksanen , and O. Suominen . 2004a. Importance of large and small mammalian herbivores for the plant community structure in the forest tundra ecotone. Oikos 106:324–334.
- Olofsson, J. , S. Stark , and L. Oksanen . 2004b. Reindeer influence on ecosystem processes in the tundra. Oikos 105:386–396.
- Olofsson, J. , P. E. Hulme , L. Oksanen , and O. Suominen . 2005. Effects of mammalian herbivores on revegetation of disturbed areas in the forest-tundra ecotone in northern Fennoscandia. Landscape Ecology 20:351–359.
- Ouellet, J. P. , S. Boutin , and D. C. Heard . 1993. Range impacts following the introduction of caribou on Southampton Island, Northwest Territories, Canada. Arctic and Alpine Research 25:136–141.
- Parker, K. L. , P. S. Barboza , and T. R. Stephenson . 2005. Protein conservation in female caribou ( Rangifer tarandus ): Effects of decreasing diet quality during winter. Journal of Mammalogy 86:610–622.
- Pruitt, W. O. 1959. Snow as a factor in the winter ecology of the barren ground caribou ( Rangifer arcticus ). Arctic 12:159–179.
- Pösö, A. R. 2005. Seasonal changes in reindeer physiology. Rangifer 25:31–38.
- Schluter, D. 1981. Does the theory of optimal diets apply in complex environments. American Naturalist 118:139–147.
- Skogland, T. 1978. Characteristics of the snow cover and its relationship to wild mountain reindeer ( Rangifer tarandus tarandus L) feeding strategies. Arctic and Alpine Research 10:569–580.
- Skogland, T. 1983. The effects of density dependent resource limitation on size of wild reindeer. Oecologia 60:156–168.
- Skogland, T. 1985. The effects of density-dependent resource limitations on the demography of wild reindeer. Journal of Animal Ecology 54:359–374.
- Skogland, T. 1986. Movements of tagged and radio-instrumented wild reindeer in relation to habitat alternation in the Snøhetta region, Norway. Rangifer Special Issue 1:267–272.
- Skogland, T. 1988. Tooth wear by food limitation and its life history consequences in wild reindeer. Oikos 51:238–242.
- Skogland, T. 1989. Comparative social organization of wild reinder in relation to food, mates and predator avoidance. Advances in Ethology No. 29.
- SPSS 1997. SigmaStat 2.0 for Windows, User's Manual Chicago SPSS Inc.
- Staaland, H. and C. R. Olesen . 1992. Muskox and caribou adaptation to grazing on the Angujaartorfiup Nunaa range in West Greenland. Rangifer 12:105–113.
- Staaland, H. and R. G. White . 1991. Influence of forage ecology on alimentary tract size and function of Svalbard reindeer. Canadian Journal of Zoology 69:1326–1334.
- Staaland, H. , I. Brattbakk , K. Ekern , and K. Kildemo . 1983. Chemical composition of reindeer forage plants in Svalbard and Norway. Holarctic Ecology 6:109–122.
- Storheier, P. V. , S. D. Mathiesen , N. J. C. Tyler , and M. A. Olsen . 2002. Nutritive value of terricolous lichens for reindeer in winter. Lichenologist 34:247–257.
- Svihus, B. and Ø Holand . 2000. Lichen polysaccharides and their relation to reindeer/caribou nutrition. Journal of Range Management 53:642–648.
- Sæther, B. E. , S. Engen , J. Persson , H. Brøseth , A. Landa , and T. Willebrand . 2003. Analyser av levedyktighet hos Skandinavisk jerv. NINA Fagrapport Norwegian Institute for Nature Research, Trondheim, Norway, 28 pp. 62.
- Thing, H. 1977. Behavior, mechanics and energetics associated with winter cratering by caribou in Northwestern Alaska. Biological Papers of the University of Alaska, 18 Fairbanks University of Alaska. 41.
- Tucker, B. , S. Mahoney , B. Greende , E. Menchenton , and L. Russell . 1991. The influence of snow depth and hardness on winter habitat selection by caribou on the southwest coast of Newfoundland. Rangifer Special Issue 7:160–163.
- Tveitnes, A. 1980. Lavgranskning på Hardangervidda. Research in Norwegian Agriculture 31:287–365.
- Tømmervik, H. , B. Johansen , I. Tombre , D. Thannheiser , K. A. Hogda , E. Gaare , and F. E. Wielgolaski . 2004. Vegetation changes in the Nordic mountain birch forest: The influence of grazing and climate change. Arctic, Antarctic, and Alpine Research 36:323–332.
- van der Wal, R. 2006. Do herbivores cause habitat degradation or vegetation state transition? Evidence from the tundra. Oikos 114:177–186.
- van der Wal, R. , R. Brooker , E. Cooper , and R. Langvatn . 2001a. Differential effects of reindeer on high Arctic lichens. Journal of Vegetation Science 12:705–710.
- van der Wal, R. , S. M. J. van Lieshout , and M. J. J. E. Loonen . 2001b. Herbivore impact on moss depth, soil temperature and arctic plant growth. Polar Biology 24:29–32.
- van der Wal, R. and R. W. Brooker . 2004. Mosses mediate grazer impacts on grass abundance in arctic ecosystems. Functional Ecology 18:77–86.
- Van Soest, P. J. 1994. Nutritional Ecology of the Ruminant Ithaca, N.Y Cornell University. 476.
- Vilchek, G. E. 1997. Arctic ecosystem stability and disturbance: a western Siberian case history. In Crawford, R. M. M. , editor. Disturbance and Recovery in Arctic lands Dordrecht Kluwer Academic Publishers. 179–189.
- Virtanen, R. , H. Henttonen , and K. Laine . 1997. Lemming grazing and structure of a snowbed plant community – A long-term experiment at Kilpisjärvi, Finnish Lapland. Oikos 79:155–166.
- Virtanen, R. , J. Oksanen , L. Oksanen , and V. Y. Razzhivin . 2006. Broad-scale vegetation-environment relationships in Eurasian high-latitude areas. Journal of Vegetation Science 17:519–528.
- Vistnes, I. , C. Nellemann , P. Jordhøy , and O. Strand . 2001. Wild reindeer: impacts of progressive infrastructure development on distribution and range use. Polar Biology 24:531–537.
- Vistnes, I. , C. Nellemann , P. Jordhøy , and O. Strand . 2004. Effects of infrastructure on migration and range use of wild reindeer. Journal of Wildlife Management 68:101–108.
- Väre, H. , R. Ohtonen , and L. Oksanen . 1995. Effects of reindeer grazing on understorey vegetation in dry Pinus sylvestris forests. Journal of Vegetation Science 6:523–530.
- Väre, H. , R. Ohtonen , and K. Mikkola . 1996. The effect and extent of heavy grazing by reindeer in oligotrophic pine heaths in northeastern Fennoscandia. Ecography 19:245–253.
- Wratten, S. D. and G. Fry . 1980. Field and Laboratory Exercises in Field Ecology London Edward Arnold.