ABSTRACT
After more than half a century of paleoenvironmental investigations, disagreements persist as to the nature of vegetation type and climate of the Bering land bridge (BLB) during the late Wisconsin (Sartan) glacial interval. Few data exist from sites on the former land bridge, now submerged under the Bering and Chukchi Seas. Two hypotheses have emerged during the past decade. The first, based on pollen data from Bering Sea islands and adjacent mainlands of western Alaska and Northeast Siberia, represents the likely predominant vegetation on the Bering land bridge during full-glacial conditions: graminoid-herb-willow tundra vegetation associated with cold, dry winters and cool, dry summer climate. The second hypothesis suggests that dwarf birch-shrub-herb tundra formed a broad belt across the BLB, and that mesic vegetation was associated with cold, snowier winters and moist, cool summers. As a step towards resolving this controversy, a sediment core from Norton Sound, northeastern Bering Sea was radiocarbon dated and analyzed for pollen content. Two pollen zones were identified. The older, bracketed by radiocarbon ages of 29,500 and 11,515 14C yr BP, contains pollen assemblages composed of grass, sedge, wormwood, willow, and a variety of herb (forb) taxa. These assemblages are interpreted to represent graminoid-herb-willow tundra vegetation that developed under an arid, cool climate regime. The younger pollen zone sediments were deposited about 11,515 14C yr BP, when rising sea level had begun to flood the BLB. This younger pollen zone contains pollen of birch, willow, heaths, aquatic plants, and spores of sphagnum moss. This is interpreted to represent a Lateglacial dwarf birch-heath-willow-herb tundra vegetation, likely associated with a wetter climate with deeper winter snows, and moist, cool summers. This record supports the first hypothesis, that graminoid-herb-willow tundra vegetation extended into the lowlands of the BLB during full glacial conditions of the late Wisconsin.
Introduction
The present-day Bering and Chukchi Seas between western Alaska and eastern Chukotka in Siberia cover a vast continental shelf that was subaerially exposed to varying degrees by lower sea level during Pleistocene glaciations. During the late Wisconsin glacial maximum, an area of the Bering-Chukchi continental shelf in excess of 1.4 × 106 km2 was subaerially exposed when eustatic sea level dropped to about 125 m below present sea level (; CitationFairbanks, 1989; CitationBard et al., 1990). These exposed lands formed a broad connection, or land bridge that has been biologically important for the exchange of plants and animals between Eurasia and the Americas (CitationKurtén and Anderson, 1980; CitationRepenning, 1980; CitationGuthrie, 1990). The Bering land bridge and its southern coast are also likely routes for human entry into the Americas from northeast Asia more than 12,000 14C yr BP; ∼13,800 cal. yr BP (CitationWest, 1996; CitationDixon, 1999, Citation2001; CitationMandryk et al., 2001).
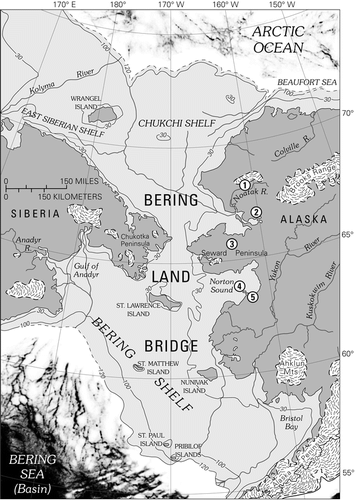
Figure 1 Map of central Beringia, showing extent of Bering land bridge during the late Wisconsin maximum, ∼18,000 14C yr BP. Glacial ice extent is based on CitationKaufman and Hopkins (1986), CitationHamilton (1994), CitationVelichko et al. (1984), and CitationKaufman and Manley (2004). Key sites with pollen evidence for late Wisconsin vegetation discussed in this paper include (1) Kaiyak Lake, northwestern Alaska (CitationAnderson, 1985); (2) Squirrel Lake, northwestern Alaska (CitationAnderson, 1985); (3) Imuruk Lake, Seward Peninsula (CitationColinvaux, 1964; CitationColbaugh, 1968); (4) Core 79-121, Norton Sound (CitationMuhs et al., 2003; this paper); and (5) Zagoskin Lake, St. Michael Island (CitationAger, 1983, Citation2003).
The paleoenvironmental history of the Bering land bridge and the adjacent mainlands has been the subject of scientific investigations for more than a half-century (e.g., CitationHopkins, 1967, Citation1979, Citation1982; CitationHopkins et al., 1960, Citation1981, Citation1982; CitationMcCulloch and Hopkins, 1966; CitationElias and Brigham-Grette, 2001; CitationGuthrie, R. D., 1990, Citation2001; CitationColinvaux, 1964, Citation1981; CitationAger, 1982, Citation2003; CitationBrigham-Grette et al., 2004). However, the limited data relating to the paleoenvironments on the former land bridge itself has led to differing interpretations among researchers about the nature of Bering land bridge vegetation and climate during the late Wisconsin. One group argues that a mesic climate prevailed on the land bridge, even during the maximum of the late Wisconsin glacial interval; and that mesic herb or shrub-herb tundra may have been widespread on the land bridge (CitationGuthrie, 2001; CitationYurtsev, 2001; CitationElias et al., 1996, Citation1997). An alternative reconstruction, based on pollen data from islands in the Bering Sea and the mainland of western Alaska, suggests that the land bridge climate was arid, even near its southern shores, and that the vegetation was predominantly graminoid-herb-willow tundra during full glacial time (CitationAger, 1982, Citation1983, Citation2002, Citation2003; CitationColinvaux, 1981).
In this paper, we present new pollen data and radiocarbon dates from a sediment core from the former Bering land bridge that provides a rare glimpse of environmental conditions that prevailed there during the late Wisconsin and Lateglacial. Pollen data from a single core from the former land bridge is not necessarily representative of environments covering the entire, vast land bridge, but these new data demonstrate the potential for obtaining detailed pollen records from terrestrial sediments recovered from the former land bridge. Previous attempts to extract paleoenvironmental data from cores of terrestrial deposits on the former land bridge focused on peaty horizons, which are rare, except in Lateglacial age sediments, deposited during a time of climatic transition (CitationElias et al., 1992, Citation1996, Citation1997).
This discussion will focus primarily on central Beringia, which is defined for the purposes of this paper as the area of the former Bering land bridge, now submerged under the waters of the Bering and Chukchi Seas, except for scattered islands (). Onshore sites in mainland western Alaska and northeastern Siberia are also discussed. In recent usage, unglaciated regions of northern and interior Alaska and interior Yukon Territory are included in eastern Beringia (CitationElias and Brigham-Grette, 2001). Northeastern Siberia, including Chukotka, is referred to as western Beringia.
Many of the previous paleoenvironmental studies in central Beringia utilized fossil assemblages (e.g., pollen and spores, plant macrofossils, insect fossils) obtained from lake sediment cores and coastal exposures on islands in the Bering Sea and from western mainland Alaska (e.g., CitationColinvaux, 1964, Citation1967a, Citation1967b, Citation1981; CitationColbaugh, 1968; CitationAger, 1982, Citation1983, Citation2002, Citation2003; CitationAnderson, 1985, Citation1988; CitationElias et al., 1996,Citation1997; CitationBrubaker et al., 2001; CitationKurek and Cwynar, 2007).
Until recently, the late Pleistocene paleovegetation and paleoclimate history of northeastern Siberia was little known. Now several well-documented late Pleistocene sites with radiocarbon-dated pollen records have been reported from Chukotka and adjacent areas of western Beringia (e.g., CitationAnderson and Lozhkin, 2001, Citation2002; CitationAnderson et al., 2002; CitationLozhkin et al., 2007). The available evidence from western Alaska and northeastern Siberia indicate that graminoid-herb-willow tundra was the predominant vegetation type in those regions during full glacial time of the late Wisconsin (Sartan) glacial interval. However, little paleoenvironmental evidence has been documented from terrestrial sediments of the now-submerged Bering land bridge. Few sediment cores from the Bering and Chukchi platforms penetrate beyond the marine sediments that have accumulated since the marine transgression that began following the late glacial maximum. A few researchers have studied fossil evidence preserved in sediment cores obtained from the continental shelves of the Bering and Chukchi Seas (CitationColinvaux, 1967a; CitationElias et al., 1992, Citation1996, Citation1997). However, nearly all the samples previously analyzed from sediment cores from the Bering-Chukchi continental shelf date to the latest Wisconsin (Lateglacial, ∼14,000–10,000 14C yr BP; ∼16,300–11,440 cal. yr BP) or early Holocene (10,000–7500 14C yr BP; ∼11,440–8330 cal. yr BP), a time of rapid sea level rise as well as climate and ecological change. One such record is from marine or estuarine sediments that accumulated during the marine transgression of the land bridge (e.g., CitationColinvaux, 1967a). Very few reported samples of land bridge terrestrial sediments are associated with apparent full glacial, late Wisconsin radiocarbon ages more than 14,000 14C yr BP; ∼16,300 cal. yr BP (, west of Port Clarence area; CitationElias et al., 1996, Citation1997).
Figure 2 Map of Norton Sound, northeastern Bering Sea, showing bathymetry (CitationSharma, 1979), and distribution of major (modern) vegetation types (CitationJFSLUPCFA, 1973). Also shown are: (1) site of USGS core 76-121; (2) St. Michael Island, site of Puyuk and Zagoskin Lake pollen records (CitationAger, 1982, Citation1983, Citation2003); (3) Village of St. Michael, site of climate data station (see ); (4) Village of Unalakleet, site of climate data station (see ); (5) City of Nome, site of climate data station (see ); (6) Village of Teller, site of climate data station ().
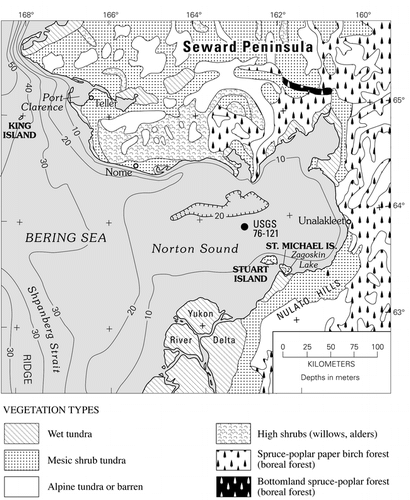
Table 1 Climate data (temperature and precipitation) from weather stations around Norton Sound.
This paper presents evidence from 26 pollen samples and 6 radiocarbon dates from a sediment core (USGS 76-121) obtained in 1976 by the U.S. Geological Survey in central Norton Sound, northeastern Bering Sea ( and ). This record provides the first detailed pollen evidence that can be used to reconstruct vegetation of mid-Wisconsin interstadial and late Wisconsin glacial age from a lowland site on the former Bering land bridge.
Study Site
Norton Sound is a shallow embayment of northeastern Bering Sea, bordered to the north by Seward Peninsula, to the south by the northern part of the Yukon River delta, and to the southeast and east by the Nulato Hills (). Most of present-day Norton Sound is less than 20 m deep, and postglacial sea level rise transgressed across most of the sound during the early to middle Holocene (CitationNelson, 1982; CitationFairbanks, 1989; CitationBard et al., 1990; CitationBauch et al., 2001; CitationMason and Jordan, 2002). The coring site discussed here (USGS 76-121) is located in central Norton Sound at 63°53′N, 162°42′W (). Present-day water depth at the coring site is 17 m.
The Holocene marine transgression of Norton Sound, along with regional ocean currents, tidal currents, sea-ice pressure ridge gouging, and storm waves have all resulted in the erosion and extensive reworking of Holocene marine sediments as well as former land bridge terrestrial sediments younger than about 11,000 14C yr BP; ∼12,875 cal. yr BP in Norton Sound (CitationSharma, 1979; CitationNelson, 1982; CitationOlsen et al., 1982). The terrestrial pollen record recovered from Norton Sound USGS core 76-121 spans only the time between ∼29,500 and 11,500 14C yr BP; ∼34,900 and 12,875 cal. yr BP.
Modern Climate
Modern climate of the Norton Sound area can be inferred from regional climate zones of Alaska and the limited climate data available from the region. Past or presently active weather stations are located at St. Michael, Unalakleet, Nome, and Teller (; ). The Norton Sound region has a mean annual air temperature of about −2 to −4°C. However, summer temperatures in recent decades have been warm enough to support tree growth, even in areas that are not presently forested (; ). Either trees have not yet had time to spread into the lands adjacent to western Norton Sound, or other environmental factors inhibit tree survival in these areas. The lands surrounding Norton Sound fall within the zone of discontinuous permafrost (CitationPéwé, 1975). The Norton Sound area climate is characterized as transitional between Arctic (to the north), continental (to the east and northeast), and maritime (to the southwest) zones (CitationPéwé, 1975). In winter, sea ice forms in the northern Bering Sea and expands southward, sometimes as far south as the Pribilof Islands, with some scattered ice drifting farther south into the deeper waters of Bering Sea basin. Sea ice dominates northern Bering Sea for nearly nine months of the year, from October to June, and is particularly abundant between January and May (CitationRay and Dupré, 1981).
Modern Vegetation
Modern vegetation on the lands surrounding Norton Sound includes mesic shrub tundra, wet tundra in low coastal areas, bog vegetation, alpine tundra in mountainous areas, shrub thickets in lowlands and gullies on uplands, and some areas of lowland boreal spruce forest on southeastern Seward Peninsula and near Unalakleet (; CitationJoint Federal-State Land Use Planning Commission for Alaska (JFSLUPCA), 1973; CitationViereck and Little, 1972, Citation1975; CitationAger, 1982, Citation1983, Citation2003). In areas with boreal forest (), common trees include Picea glauca (white spruce), P. mariana (black spruce), Betula papyrifera (paper birch), and Populus balsamifera (balsam poplar). Common large shrub taxa in the Norton Sound region include Alnus crispa (American green alder), and several Salix (willow) species, e.g., Salix alaxensis, S. glauca, S. lanata, and S. planifolia ssp. pulchra. Common low shrubs, both within boreal forest and moist tundra, include Betula nana (dwarf birch), Salix reticulata (netleaf willow), Spiraea beauverdiana (Alaska spiraea), Potentilla fruticosa (bush cinquefoil), Vaccinium uliginosum (bog blueberry), V. vitis-idaea (mountain cranberry), Ledum decumbens (Labrador tea), Andromeda polifolia (bog rosemary), and Empetrum nigrum (crowberry), following the taxonomic nomenclature of CitationViereck and Little (1972, Citation1975).
Common herbaceous taxa include Cyperaceae (sedges) such as Eriophorum spp. and Carex spp., Polygonum spp. (bistort) and Rumex spp., (sourdock), Epilobium spp. (fireweed), several genera of Ranunculaceae, e.g., Delphinium, Aconitum, several genera of the Asteraceae (e.g., Artemisia, Achillea, Aster, Petasites, Arnica, Senecio), Valeriana, and Pedicularis spp. Nomenclature for herbs generally follows CitationHultén (1968), although a number of plant family names have been updated.
In many areas of the lowlands and gently sloping uplands surrounding Norton Sound today, peat of mostly Holocene age covers extensive areas, and is often more than a meter in thickness (CitationAger, 2003). The main contributors to peat formation in this region are mosses, especially Sphagnum spp., Cyperaceae, Poaceae (grasses), Ericales (heaths) such as Vaccinium spp. and Empetrum nigrum, Salix, and Betula nana. Permafrost with ice wedge polygons exists today in these widespread peat deposits.
Methods
Core USGS 76-121 was collected from central Norton Sound by the U.S. Geological Survey in 1976 (CitationNelson, 1982; CitationOlsen et al., 1982). The 10 × 10 cm square sediment core was obtained with a vibracorer. The core has been preserved in a refrigerated archival storage facility maintained by the U.S. Geological Survey in Redwood City, California. The core was sampled for pollen analysis, sediment analysis, and Accelerator Mass Spectrometry (AMS) radiocarbon dating by the authors in 1999. Several previous publications mention or describe this core (CitationNelson, 1982; CitationOlsen et al., 1982; CitationElias et al., 1996, Citation1997, CitationMuhs et al., 2003). The original core was 180 cm long. At the time we sampled the core, the lowermost segment (172.5–180 cm) was missing as a result of previous sampling for a bulk-sediment, conventional radiocarbon date (one of two conventional bulk sediment radiocarbon dates reported for this core in CitationElias et al., 1997) and other analyses. We therefore have no pollen data for the lowermost dated core interval 172.5–180 cm.
The samples obtained from the core in 1999 were processed for pollen analysis using standard maceration methods (CitationDoher, 1980) at the U.S. Geological Survey Cenozoic Palynology Laboratory in Denver, Colorado. Several sediment samples from Core 76-121 were wet-sieved to concentrate fine plant detritus for radiocarbon dating. Pollen analysis was conducted by the senior author, and relied on published pollen floras (e.g., CitationMoriya, 1978) and a collection of modern pollen and spore reference slides housed at the U.S. Geological Survey, Denver, to identify pollen and spore types.
Counts of more than 300 pollen grains (plus spores) were identified and tabulated for all but one sparse sample from 127 cm core depth, which had a pollen sum of 176 grains. Pollen percentages were calculated from a sum that included all pollen types. Spore percentages were calculated from a sum that included all pollen grains + spores counted.
John McGeehin of the U.S. Geological Survey Radiocarbon Laboratory prepared our samples for AMS radiocarbon dating. Calibration of radiocarbon ages was done using the Columbia University calibration website (http://www.radiocarbon.ldgo.columbia.edu/research/radiocarbon.htm).
Results
Core Description
The upper 19 cm of sediments in Core 76-121 (left column, ) consists of bioturbated marine sandy silt containing a few fragments of marine mollusk shells. Between 19 and 91 cm core depths, the sediments are non-marine, and consist of partly laminated olive gray to black silt, within which layers of peaty silt occur sporadically.
Figure 3 Pollen and spore percentage diagram for USGS Core 76-121, central Norton Sound. Lithology column for the core is shown at the left of the diagram, and radiocarbon dates from the core are listed along the right margin, and in . See for location of the coring site within Norton Sound.
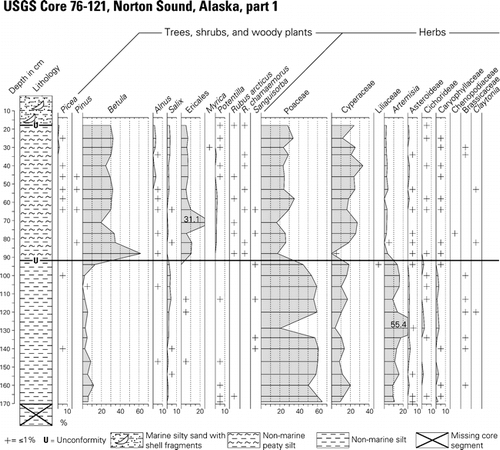
Figure 3 Continued.

Table 2 Radiocarbon ages and calibrated ages for USGS core 76-121. Calibrations follow the Fairbanks 0107 calibration curve (CitationFairbanks et al., 2005). Data from http://radiocarbon.ldeo.columbia.edu/research/radiocarbon.htm on 21 September 2007. The marine reservoir correction for sample NOR-11 is based on data from CitationDumond and Griffin (2002). The USGS conventional bulk-sediment radiocarbon dates shown here were previously reported in CitationElias et al. (1996, Citation1997).
Between core depths of 91 and 172.5 cm, the sediments are massive fine silt in which fine plant detritus is very sparse. No peat layers were observed in that part of the core. The original basal core segment between 172.5 and 180 cm was unavailable to us, but a previously published description of the original core suggests that the missing core segment was similar in lithology to the overlying silty sediments (CitationOlsen et al., 1982). More recently, samples from the massive silt unit within the core below core depth of 91 cm were subjected to geochemical and sedimentological analyses. The data were compared with analytical data from silt deposits from other sites in Alaska and elsewhere (CitationMuhs et al., 2003). The latter study suggests that the silt unit in the lower part of Core 79-121 (NOR-2) represents a probable loess deposit that accumulated on the former Bering land bridge in what is now central Norton Sound during the late Wisconsin glacial interval.
Data obtained from pollen and spore analysis of 26 samples from the non-marine sediments in USGS Core 79-121 were used to construct a pollen and spore percentage diagram (). The pollen diagram and radiocarbon and calibrated ages () from the core provide a record of terrestrial environments on the former Bering land bridge beginning perhaps with the latest part of the middle Wisconsin interstadial (Marine Isotope Stage 3 [MIS 3]), and extending through the late Wisconsin glacial interval (MIS 2), to the latest Wisconsin transition from full glacial to Lateglacial conditions.
No pollen analyses were attempted for the sediments in the upper 19 cm of the core, because the marine deposits have been extensively reworked. Bioturbation, currents, tides, storm waves, and perhaps sea ice gouging of the shallow sea floor have all been identified as agents of sediment reworking in this area (CitationSharma, 1979; CitationOlsen et al., 1982; CitationThor and Nelson, 1981; CitationLarsen et al., 1981). A marine mollusk fragment recovered from near the base of the marine sediments in Core 79-121 yielded an AMS radiocarbon age of 1120 ± 40 14C yr BP (; ). The marine reservoir effect on marine shell material and bone in this region varies between 450 and 750 radiocarbon years (CitationDumond and Griffin, 2002). Marine reservoir effects of similar magnitude have been reported from dated marine fossils from cores on the continental shelf north of Siberia (CitationBauch et al., 2001). Therefore, the thin veneer of marine sediments at the top of the core has probably been reworked repeatedly during the past few centuries, based on the marine reservoir-adjusted radiocarbon age range of the shell fragment (). Erosion of land bridge sediments during the early to middle Holocene marine transgression, plus reworking of uppermost sediments in these shallow waters during major storms have apparently been responsible for the removal of the earliest Holocene and some of the Lateglacial age terrestrial sediment record at this site.
The abrupt transition between massive silts below core depth 91 cm and the partly laminated peaty silt between 19 and 91 cm core depth may mark the position of an unconformity, caused perhaps by wind deflation, thermokarst melting, or shoreline erosion within a shallow pond during Lateglacial time. Three radiocarbon ages from within the peaty silt layer (pollen zone NOR-1) are all very similar (), but they decrease slightly in age with depth. The counting errors at one standard deviation for the reported dates for adjacent samples in pollen zone NOR-1 samples overlap, permitting an interpretation of no significant age differences within this core interval. One of the samples is from a previously-studied core segment that was conventionally dated using a large-volume bulk-sediment sample ().
The slight differences in ages of samples from within zone NOR-1 suggests rapid sedimentation rates in zone NOR-1. The discontinuous sediment laminations and layers of silty peat suggest that the upper deposits coinciding with pollen zone NOR-1 probably accumulated in a shallow, perhaps thermokarst pond or lake. Pollen of aquatic and semi-aquatic plants in this part of the core supports the interpretation of the peaty sediments as lacustrine. The dominant silt-sized mineral fraction of the NOR-1 zone sediments may be partly derived from redeposition of older loess deposits, but some loess deposition is likely to have continued during the late glacial interval when NOR-1 accumulated (CitationMuhs et al., 2003).
Pollen and Spore Analysis
The pollen and spore percentage diagram () can be divided into two pollen and spore assemblage zones. The older (zone NOR-2) spans core depths 92–180 cm. This zone is bracketed by a previously published basal bulk radiocarbon age of 29,500 ± 340 14C yr BP; ∼34,918 ± 362 cal. yr BP (CitationElias et al., 1996, Citation1997), and a limiting age of 11,515 ± 40 14C yr BP; ∼13,419 ± 108 cal. yr BP () from near the base of the overlying Zone NOR-1. A third radiocarbon sample from this zone was obtained from core depth 140–142 cm. Fine plant detritus from this sample yielded an age of 22,780 ± 100 14C yr BP; ∼27,283 ± 134 cal. yr BP ().
Pollen and spore assemblage zone NOR-2 is characterized by abundant pollen of Poaceae (50–60%), Cyperaceae (10–15%), Salix (<5%), Artemisia (10–15%), and other herbs that are generally present in amounts of <2%, e.g., Asteraceae subf. Asteroideae, Asteraceae subf. Cichorideae, Caryophyllaceae, Brassicaceae, Saxifragaceae, Ranunculus, Caltha, Thalictrum, Coptis, Polemonium, and Valeriana. Spore-producing plants such as Polypodiaceae (ferns), Lycopodium spp. (clubmoss), Sphagnum, and other mosses are very rarely represented in zone NOR-2 sediments.
The graminoid-herb pollen assemblage in zone NOR-2 is quite similar to full-glacial pollen assemblages described from St. Michael Island (CitationAger, 1982, Citation1983, Citation2003), Imuruk Lake on north-central Seward Peninsula (CitationColinvaux, 1964; CitationColbaugh, 1968), northwestern Alaska near Kotzebue Sound (; CitationAnderson, 1985, Citation1988), and other sites in northwest Alaska (CitationMann et al., 2002; CitationOswald et al., 1999). An AMS radiocarbon age of 22,780 ± 100 14C yr BP () on fine plant detritus collected from near the mid-point within pollen assemblage zone NOR-2 indicates that most of the silt unit was deposited during the late Wisconsin, rather than the late mid-Wisconsin interstadial.
The pollen and spore assemblages found in zone NOR-2 suggest that the dominant vegetation growing on former land bridge lowlands (now the floor of Norton Sound) during the late Wisconsin glacial interval (MIS-2) was a graminoid-herb tundra with some scattered, probably low-growing willows. The composition of the herb flora, similar to but somewhat less diverse than seen in the Zagoskin Lake full-glacial record, suggests vegetation of grasses, sedges, and forbs. Some of the forb taxa (e.g., Artemisia, Asteroideae, Cichorideae, Brassicaceae, Caryophyllaceae) are often associated with disturbed habitats (e.g., frost boils, loess deposition, heavy grazing pressure). Loess deposition was probably an important factor in maintaining the productivity of the graminoid-herb vegetation. The dominance of pollen of Poaceae and abundance of Artemisia closely resembles full glacial pollen assemblages from Zagoskin Lake, 65 km southeast of the Norton Sound core site ( and ; CitationAger, 2003). However, in the Zagoskin Lake core, the transition from full-glacial graminoid herb tundra to a more mesic Betula-Ericales shrub tundra apparently began ca. 15,000–14,000 14C yr BP (∼18,100–16,300 cal. yr BP). The radiocarbon age from near the base of the silty peat unit (pollen zone NOR-1, ; ) could therefore be more than 3000 14C years younger than the transitional event marking this zone boundary in the Zagoskin Lake core and in most other pollen, plant macrofossil, and insect fossil records from Beringia (e.g., CitationElias et al., 1996, Citation1997; CitationAger, 1982; Citation2003; CitationClague et al., 2004; CitationHopkins, 1982). However, the age for the transition from graminoid-herb zone to shrub birch-herb zone was, until recent years, based on bulk-sediment conventional radiocarbon ages, which tend to be older than dated macrofossils from the same samples (e.g., CitationAbbott and Stafford, 1996). Recent pollen studies using AMS-dated macrofossils in western and interior Alaska suggest that the transition from graminoid-herb zone to shrub birch zone was later, probably about 13,000–11,800 14C yr BP; ∼15,140–13,650 cal. yr BP (e.g., CitationBrubaker et al., 2001; CitationMann et al., 2002; CitationOswald et al., 1999; CitationBigelow and Edwards, 2001). If the macrofossil ages for the regional vegetation change to shrub birch-herb tundra are correct, then there may be only a minor unconformity at the NOR-1/NOR-2 zone boundary. If an unconformity is present, it may have been the result of thermokarst melting during the Lateglacial. If the sedimentation rate calculated for the lower half of zone NOR-2 is projected upwards in the core to the NOR-1/NOR-2 boundary, the extrapolated age for the zone boundary is about 13,000 14C yr BP; ∼15,140 cal. yr B.P.
Pollen Assemblage Zone NOR-1 (core depth 21 to 91 cm; age range ∼11,500–11,700 14C yr BP; ∼13,360–13,560 cal. yr BP) occurs in a slightly peaty silt to fine sand that accumulated rapidly, judging from the similarity of radiocarbon dates on peaty plant detritus from within the zone (; ). Pollen assemblage zone NOR-1 is characterized by a greater abundance of shrub and low woody plant taxa, such as pollen of Betula (probably dwarf birch, Betula nana, 20–62%), Salix (0–3.2%), Ericales: Ericaceae + Empetrum (5–11%), and Rubus chamaemorus (<1%). Small amounts of Picea (0–2%) and Pinus (pine, <1%) are present in the core above core depth 65 cm, and may represent reworked fossils or long distance wind-transported grains. Herbaceous taxa in zone NOR-1 are dominated by pollen of Poaceae (20–40%) and Cyperaceae, 18–30%. Other herbs, such as Apiaceae, Brassicaceae, Saxifragaceae, Thalictrum, and Polemonium are usually represented in amounts of less than 1% of the pollen sum. Aquatic and semi-aquatic/wet habitat plants are relatively well represented in zone NOR-1, compared with zone NOR-2 samples. Such wetland taxa include Hippuris, Myriophyllum, Nuphar, Potamogeton, Sparganium, Caltha, and Menyanthes. Spore-producing plants are far better represented in NOR-1 than in NOR-2. Spore types represented include Lycopodium spp., and Polypodiaceae fern spores. Spores of mosses, especially Sphagnum mosses, are relatively abundant in some NOR-1 samples, whereas they are rare in NOR-2 samples.
The paucity of plant detritus within the sediments of zone NOR-2 suggests that local environmental conditions were not conducive to the accumulation and preservation of peat during full-glacial times of the late Wisconsin. Pollen grains of woody plants, other than Salix, are poorly represented, and pollen of aquatic and semi-aquatic plants are rare in Zone NOR-2, as are spores of ferns and Sphagnum mosses. This suggests that the landscape was dominated by vegetation with few peat-forming plants present.
Discussion
The full-glacial silt deposit and the pollen it contains (Pollen Zone NOR-2) appear to have accumulated under conditions where evaporation exceeded precipitation. With silty soils and ground cover of graminoid-herb-willow tundra vegetation, and a general lack of peat accumulation, the permafrost table was probably substantially lower than is the case in the region today. Under current conditions of vegetation and climate (, ), much of the landscapes surrounding Norton Sound are mantled with a thick peat layer that insulates the soil and allows the permafrost table to persist at shallow depths. During glacial times it is likely that winter snow cover was thinner than it is today (CitationGuthrie, 1990, Citation2001; CitationHopkins, 1982), which would likely result in a longer growing season in spite of full-glacial climatic conditions prevailing during much of the late Wisconsin. CitationHopkins (1982) summarized evidence for arid conditions in Beringia such as widespread eolian deposits, sand wedges, low organic content of sediments, and evidence for lower lake levels or dry lake basins. CitationBarber and Finney (2000) and CitationAbbott et al. (2000) studied late Quaternary lake level fluctuations in interior Alaska and estimated that precipitation was 35–75% lower than at present prior to 12,500 14C yr BP; ∼14,480 cal. yr BP. Lakes were shallow or dried out. A consensus view of full glacial climates of Beringia remains elusive, however. CitationElias (2001) suggests that maximum mean summer temperatures may have been about 6.4°C cooler than modern conditions in eastern Beringia, based on present-day climatic tolerance limits of modern insect species found in dated fossil assemblages from the region. However, a recent study of chironomid assemblages from the Zagoskin Lake core on St. Michael Island in Norton Sound suggests that mean summer temperatures during the late glacial maximum were only about 1–4°C cooler than today (CitationKurek and Cwynar, 2007). If correct, this temperature reconstruction suggests that the region's graminoid-herb-willow tundra vegetation may have been controlled more by aridity than cold growing season temperatures. This is consistent with the occurrence of trace amounts of pollen of Populus pollen in full-glacial sediments in the Zagoskin Lake core from St. Michael Island (CitationAger, 2003). Trees rarely grow where mean July temperatures fall much below 10°C. Recent pollen-based biome reconstructions and vegetation modeling for Beringia also suggest that aridity was a major factor favoring development of graminoid-herb tundra during full glacial conditions (CitationBigelow et al., 2003; CitationKaplan et al., 2003).
Deeper permafrost and water tables, silty soils, aridity, and high evaporation rates would also be unfavorable for widespread development of mesic and wetland habitats. However, some mesic habitats apparently existed on the Bering land bridge, as indicated by macrofossils of mesic to wetland habitat plants and insects found preserved in sediments in central Beringia (CitationElias et al., 1996, Citation1997; CitationElias, 2001). The pollen evidence presented here, however, shows very low percentages of mesic habitat plants on the land bridge. It is likely, therefore, that mesic to wetland habitats probably occupied only scattered and restricted areas of the Bering land bridge during full-glacial conditions.
The graminoid-rich vegetation on the land bridge and on what are now adjacent mainlands and islands of western Alaska apparently provided suitable forage to support the late Pleistocene large grazing mammals (Bison priscus; Mammuthus primigenius; Rangifer tarandus; Equus; Saiga) that wandered across Beringia. According to CitationGuthrie (1990, Citation2001), most of the grazing mammal species of Beringia required nutrient-rich and/or abundant graminoid-forb vegetation of a more steppe-like character than the predominantly mesic to wet, shrubby, mossy tundra communities blanketing much of Alaska today. Most present-day tundra communities in Alaska would not provide suitable forage and habitats for many of the large grazing mammals that roamed Beringia during the Wisconsin (MIS 2, 3, and 4). The available plant macrofossil data from late Wisconsin age full glacial deposits in Beringia (e.g., CitationGoetcheus and Birks, 2001; CitationZazula et al., 2006, Citation2007; CitationMatthews, 1982) suggest that grasses, sedges (e.g., Kobresia) and herbs predominated over much of the landscapes of Beringia, but most of the species involved are generally found today in dry habitats within lowland and alpine tundra. Fossils of plant species now found in prairie grasslands or true steppe environments are rarely found in Alaskan Beringian full-glacial deposits (e.g., CitationGoetcheus and Birks, 2001). In Yukon Territory, plant macrofossils of some steppe plant species have been identified from late Wisconsin deposits, and this appears to reflect a more extreme aridity in easternmost Beringia (CitationZazula et al., 2006; CitationZazula et al., 2007).
The new pollen evidence from core 76-121 does not support the hypothesis (CitationElias et al., 1996, Citation1997; CitationYurtsev, 2001; CitationGuthrie, 2001) that the Bering land bridge may have been largely covered by mesic herb tundra or mesic shrub tundra during full glacial times. However the new evidence does strongly support the interpretations presented by CitationElias et al. (1996, Citation1997) that mesic birch-shrub-herb tundra probably did cover much of the land bridge as well as adjacent mainlands during the Lateglacial (∼14,000–10,000 14C yr BP; ∼16,300–11,440 cal. yr BP), a time when rapid sea level rise was underway. The marine transgression would have allowed a more maritime climate to migrate inland adjacent to the rapidly changing coastline.
Climate models for full-glacial Beringia suggest moist air masses from the North Pacific flowing north would have resulted in moister conditions in central Beringia, especially along the southern margins of the land bridge (CitationBartlein et al., 1991; CitationBartlein et al., 1998). However, the distribution of eolian deposits and paleowind directions derived from late Wisconsin dune orientations suggest that dominant winds in central Beringia were from the northeast, rather than from the south (CitationMuhs et al., 2003). Pollen and sediment evidence from the Pribilof Islands (CitationColinvaux, 1981) indicate arid conditions and graminoid-herb vegetation on the southern margin of the land bridge during full glacial late Wisconsin. Whatever air masses from the North Pacific penetrated central Beringia at that time appear to have carried little moisture, and the moisture might have been seasonally restricted (CitationHoefle et al., 2000). Another line of evidence that suggests arid conditions in central Beringia is the relatively small extent of glaciers that formed in mountains adjacent to the Bering land bridge during the late Wisconsin glacial interval compared to older glacial deposits in the region (; CitationKaufman and Hopkins, 1986; CitationManley et al., 2001; CitationKaufman and Manley, 2004; CitationVelichko et al., 1984).
The Bering land bridge covered a vast area of the continental shelf between western Alaska and Chukotka during the maximum stage of the late Wisconsin glacial interval. We do not suggest that the data from a single marine core that penetrates to terrestrial sediments of land bridge age necessarily characterizes the vegetation and climates of the entire land bridge during full glacial time. However, this new record from Norton Sound is consistent with most of the existing paleoecological and paleoclimatic evidence for full glacial conditions in western Alaska, the islands within the area of the former land bridge (CitationAger, 1982, Citation1983, Citation2002, Citation2003; CitationColinvaux, 1964, Citation1967a, Citation1967b, Citation1981; CitationAnderson, 1985, Citation1988; CitationBrubaker et al., 2001; CitationClague et al., 2004; CitationGoetcheus and Birks, 2001) and northeast Siberia (CitationAnderson and Lozhkin, 2001, Citation2002; CitationAnderson et al., 2002; CitationLozhkin et al., 2007; CitationBrigham-Grette et al., 2004). The findings presented in this paper represent an initial step towards reconstructing the full-glacial vegetation and climate of the Bering land bridge.
The shift from graminoid-herb-willow tundra to shrub-herb-moss tundra vegetation with aquatics, semi-aquatics, and Sphagnum suggests that both a climatic change and vegetation change had occurred following full glacial time in central Beringia. The implied change in climate is towards moister conditions, perhaps with a thicker snow accumulation in winter, and perhaps a shorter growing season because of the likely longer persistence of snow (CitationGuthrie, 2001). The shift towards moister climates probably coincided with greater cloud cover during the growing season. The increase in moisture likely resulted from warmer sea surface temperatures in the North Pacific, changing atmospheric circulation patterns as ice sheets waned, and the progressive flooding of the Bering land bridge, which would result in a greater influence of maritime air masses in central Beringia and adjacent mainlands.
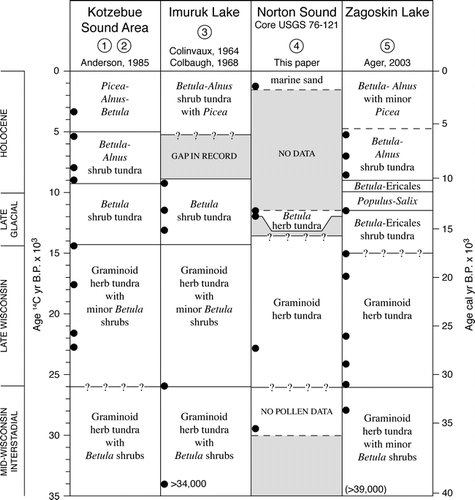
The late glacial vegetation shift to a birch-willow-Ericales-herb-moss tundra suggests that sites on the Bering land bridge as well as upland sites studied on the islands and mainland of western Alaska had broadly similar vegetation over large areas of central Beringia (CitationAger, 2002, Citation2003; CitationClague et al., 2004). This birch zone vegetation has been documented from many sites in Alaska (CitationAger, 1982, Citation1983, Citation2002, Citation2003; CitationAger and Brubaker, 1985; CitationAnderson et al., 2004; CitationBrubaker et al., 2001), as well as in Chukotka and more numerous sites farther west in Siberia (CitationAnderson and Lozhkin, 2002; CitationLozhkin et al., 2007; CitationBrigham-Grette et al., 2004). A chart () showing pollen zonations and chronologies from several key sites in western Alaska shows vegetation histories for sites near Kotzebue Sound, Seward Peninsula, central Norton Sound, and St. Michael Island that can be used to infer the vegetation history (the past ca. 11,500 14C yr; ∼13,360 cal. yr) that is missing from Core 76-121, for the landscapes immediately adjacent to Norton Sound.
Figure 4 Chart comparing central Beringian vegetation histories derived from pollen records from (1) Kaiyak Lake and (2) Squirrel Lake near Kotzebue Sound, northwest Alaska (CitationAnderson, 1985, 1987); (3) Imuruk Lake, Seward Peninsula (CitationColinvaux, 1964; CitationColbaugh, 1968); (4) USGS Core 76-121, Norton Sound (CitationMuhs et al., 2003; this paper); and (5) Zagoskin Lake, St. Michael Island, Norton Sound (CitationAger, 1983, Citation2003).
Acknowledgments
The authors gratefully acknowledge the Marine Geology Team, U.S. Geological Survey, in Menlo Park, California, for permission to sample core USGS 76-121 and other Bering Sea and Chukchi Sea cores during our visit to the marine core archival facilities in Redwood City, California, in 1999. We also thank John P. McGeehin of the U.S. Geological Survey, Reston, Virginia, for preparing our samples for AMS radiocarbon dating. We also gratefully acknowledge Dan Muhs, Paul Carrara, Pat Anderson, and Mary Edwards for their detailed reviews that substantially improved this paper. This research was supported by the U.S. Geological Survey Earth Surface Dynamics Program and is a contribution of the Alaska Quaternary Climates Project.
References Cited
- Abbott, M. B. and T. W. Stafford . 1996. Radiocarbon geochemistry of ancient and modern Arctic lakes, Baffin Island. Quaternary Research 45:300–311.
- Abbott, M. B. , B. P. Finney , M. E. Edwards , and K. R. Kelts . 2000. Lake-level reconstruction and paleohydrology of Birch Lake, central Alaska, based on seismic reflection profiles and core transects. Quaternary Research 53:154–166.
- Ager, T. A. 1982. Vegetational history of western Alaska during the Wisconsin glacial interval and Holocene. In Hopkins, D. M. , J. V. Matthews Jr , C. E. Schweger , and S. B. Young , editors. Paleoecology of Beringia New York Academic Press. 75–93.
- Ager, T. A. 1983. Holocene vegetational history of Alaska. In Wright Jr, H. E. , editor. Late Quaternary Environments of the United States. Vol. 1, The Holocene Minneapolis University of Minnesota Press. 128–141.
- Ager, T. A. 2002. Paleoenvironments of the Bering land bridge and the North Pacific coast during the late Wisconsin and early Holocene. Program and Abstracts of the 17th Biennial Meeting, American Quaternary Association, Anchorage 10–12.
- Ager, T. A. 2003. Late Quaternary vegetation and climate history of the central Bering land bridge from St. Michael Island, western Alaska. Quaternary Research 60:19–32.
- Ager, T. A. and L. B. Brubaker . 1985. Quaternary palynology and vegetational history of Alaska. In Bryant Jr, V. M. and R. G. Holloway , editors. Pollen Records of Late-Quaternary North American Sediments Dallas American Association of Stratigraphic Palynologists Foundation. 353–384.
- Anderson, P. M. 1985. Late Quaternary vegetational change in Kotzebue Sound area, northwestern Alaska. Quaternary Research 24:307–321.
- Anderson, P. M. 1988. Late Quaternary pollen records from the Kobuk and Noatak river drainages, northwestern Alaska. Quaternary Research 29:263–276.
- Anderson, P. M. and A. V. Lozhkin . 2001. The Stage 3 interstadial complex (Karginski/middle Wisconsin interval) of Beringia: variations in paleoenvironments and implications for paleoclimatic interpretations. Quaternary Science Reviews 20:93–125.
- Anderson, P. M. and A. V. Lozhkin . 2002. Late Quaternary vegetation and climate of Siberia and the Russian Far East (palynological and radiocarbon database) U.S. National Oceanic and Atmospheric Administration (NOAA) Paleoclimatology Program, and Russian Academy of Sciences, Far East Branch, North East Science Center, Magadan, Russia. 1–369.
- Anderson, P. M. , A. V. Lozhkin , and L. B. Brubaker . 2002. Implications of a 24,000-yr palynological record for a Younger Dryas cooling and for boreal forest development in northeastern Siberia. Quaternary Research 57:325–333.
- Anderson, P. M. , M. E. Edwards , and L. B. Brubaker . 2004. Results and paleoclimate implications of 35 years of paleoecological research in Alaska. In Gillespie, A. R. and S. C. Porter , editors. The Quaternary Period in the United States Amsterdam Elsevier. 427–440.
- Barber, V. A. and B. P. Finney . 2000. Late Quaternary climate reconstructions for interior Alaska based on paleo–lake level data and hydrological models. Journal of Paleolimnology 24:29–41.
- Bard, E. , B. Hamelin , R. G. Fairbanks , and A. Zindler . 1990. Calibration of the 14C timescale over the past 30,000 years using mass spectrometric U-Th ages from Barbados corals. Nature 345:405–410.
- Bartlein, P. J. , P. M. Anderson , M. E. Edwards , and P. F. McDowell . 1991. A framework for interpreting paleoclimatic variations in eastern Beringia. Quaternary International 10–12:73–83.
- Bartlein, P. J. , K. H. Anderson , P. M. Anderson , M. E. Edwards , C. J. Mock , R. S. Thompson , R. S. Webb , T. Webb III , and C. Whitlock . 1998. Paleoclimate simulations for North America over the past 21,000 yr: features of the simulated climate and comparisons with the paleoenvironmental data. Quaternary Science Reviews 17:549–585.
- Bauch, H. A. , T. Mueller-Lupp , E. Taldenkova , R. F. Spielhagen , H. Kassens , P. M. Grootes , J. Thiede , J. Heinemeier , and V. V. Petryashov . 2001. Chronology of the Holocene transgression at the north Siberian margin. Global and Planetary Change 31:125–139.
- Bigelow, N. H. and M. E. Edwards . 2001. A 14,000 yr paleoenvironmental record from Windmill Lake, central Alaska: Lateglacial and Holocene vegetation in the Alaska Range. Quaternary Science Reviews 20:203–215.
- Bigelow, N. H. , L. B. Brubaker , M. E. Edwards , S. P. Harrison , I. C. Prentice , P. M. Anderson , A. A. Andreev , P. J. Bartlein , T. R. Christensen , W. Cramer , J. O. Kaplan , A. V. Lozhkin , N. V. Matveyeva , D. F. Murray , A. D. McGuire , V. Y. Razzhivin , J. C. Ritchie , B. Smith , D. A. Walker , K. Gajewski , V. Wolf , B. H. Holmqvist , Y. Igarashi , K. Kremenetskii , A. Paus , M. F. J. Pisaric , and V. S. Volkova . 2003. Climate change and Arctic ecosystems: 1. Vegetation changes north of 55°N between the Last Glacial Maximum, mid-Holocene, and present. Journal of Geophysical Research—Atmospheres 108:D19 8170. doi:10.1029/2002JD002558.
- Brigham-Grette, J. , A. V. Lozhkin , P. M. Anderson , and O. Y. Glushkova . 2004. Paleoenvironmental conditions before and during the Last Glacial Maximum. In Madsen, D. B. , editor. Entering America: Northeast Asia and Beringia before the Last Glacial Maximum Salt Lake City University of Utah Press. 29–61.
- Brubaker, L. B. , P. M. Anderson , and F. S. Hu . 2001. Vegetation ecotone dynamics in Southwest Alaska during the Late Quaternary. Quaternary Science Reviews 20:175–188.
- Clague, J. J. , R. W. Mathewes , and T. A. Ager . 2004. Environments of northwestern North America before the Last Glacial Maximum. In Madsen, D. B. , editor. Entering America: Northeast Asia and Beringia before the Last Glacial Maximum Salt Lake City University of Utah Press. 63–94.
- Colbaugh, P. R. 1968. The Environment of the Imuruk Lake Area, Seward Peninsula, Alaska during Wisconsin time. Master's thesis. Ohio State University, Columbus. 1–118.
- Colinvaux, P. A. 1964. The environment of the Bering land bridge. Ecological Monographs 34:297–329.
- Colinvaux, P. A. 1967a. A long pollen record from St. Lawrence Island, Bering Sea (Alaska). Palaeogeography, Palaeoclimatology, Palaeoecology 3:29–48.
- Colinvaux, P. A. 1967b. Quaternary vegetational history of Arctic Alaska. In Hopkins, D. M. , editor. The Bering Land Bridge Stanford Stanford University Press. 207–231.
- Colinvaux, P. A. 1981. Historical ecology of Beringia: the south land bridge coast of St. Paul Island. Quaternary Research 16:18–36.
- Dixon, E. J. 1999. Bones, Boats & Bison—Archeology and the First Colonization of Western North America Albuquerque University of New Mexico Press. 1–322.
- Dixon, E. J. 2001. Human colonization of the Americas: timing, technology and process. Quaternary Science Reviews 20:277–299.
- Doher, L. I. 1980. Palynomorph preparation procedures currently used in the Paleontology and Stratigraphy laboratories, U.S. Geological Survey. U.S. Geological Survey Circular 830:1–29.
- Dumond, D. E. and D. G. Griffin . 2002. Measurements of the marine reservoir effect on radiocarbon ages in the eastern Bering Sea. Arctic 55:77–86.
- Elias, S. A. 2001. Mutual climatic range reconstructions of season temperatures based on Late Pleistocene fossil beetle assemblages in eastern Beringia. Quaternary Science Reviews 20:77–91.
- Elias, S. A. and J. Brigham-Grette . 2001. Beringian paleoenvironments—Festschrift in honor of D. M. Hopkins. Quaternary Science Reviews 20:1–574.
- Elias, S. A. , S. K. Short , and R. L. Phillips . 1992. Paleoecology of late-glacial peats from the Bering Land Bridge, Chukchi Sea shelf region, northwestern Alaska. Quaternary Research 38:371–378.
- Elias, S. A. , S. K. Short , C. H. Nelson , and H. H. Birks . 1996. The life and times of the Bering land bridge. Nature 382:60–63.
- Elias, S. A. , S. K. Short , and H. H. Birks . 1997. Late Wisconsin environments of the Bering Land Bridge. Palaeogeography, Palaeoclimatology, Palaeoecology 136:293–308.
- Fairbanks, R. G. 1989. A 17,000-year glacio-eustatic sea level record: influence of glacial melting rates on the Younger Dryas event and deep-ocean circulation. Nature 342:637–642.
- Fairbanks, R. G. , R. A. Mortlock , T-C. Chiu , L. Cao , A. Kaplan , T. P. Guilderson , T. W. Fairbanks , and A. L. Bloom . 2005. Marine radiocarbon calibration curve spanning 0 to 50,000 years B.P. based on paired 230Th/234U/238U and 14C dates on pristine corals. Quaternary Science Reviews 24:1781–1796.
- Goetcheus, V. G. and H. H. Birks . 2001. Full-glacial upland tundra vegetation preserved under tephra in the Beringia National Park, Seward Peninsula, Alaska. Quaternary Science Reviews 20:135–147.
- Guthrie, R. D. 1990. Frozen Fauna of the Mammoth Steppe Chicago University of Chicago Press. 1–323.
- Guthrie, R. D. 2001. Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammoth tooth pits, buckles, and inside-out Beringia. Quaternary Science Reviews 20:549–574.
- Hamilton, T. D. 1994. Late Cenozoic glaciation of Alaska. In Plafker, G. and H. C. Berg , editors. The Geology of Alaska. The Geology of North America, Vol. G-1 Boulder Geological Society of America. 813–844.
- Hoefle, C. , M. E. Edwards , D. M. Hopkins , D. H. Mann , and C-L. Ping . 2000. The full-glacial environment of the northern Seward Peninsula, Alaska, reconstructed from the 18,000-year old Kitluk Paleosol. Quaternary Research 53:143–153.
- Hopkins, D. M. , editor. 1967. The Bering Land Bridge Stanford Stanford University Press. 1–495.
- Hopkins, D. M. 1979. Landscape and climate of Beringia during late Pleistocene and Holocene time. In Laughlin, W. S. and A. B. Harper , editors. The First Americans: Origins, Affinities, and Adaptations New York Gustav Fischer. 15–41.
- Hopkins, D. M. 1982. Aspects of the paleogeography of Beringia during the late Pleistocene. In Hopkins, D. M. , J. V. Matthews Jr , C. E. Schweger , and S. B. Young , editors. Paleoecology of Beringia New York Academic Press. 3–28.
- Hopkins, D. M. , F. S. McNeil , and E. B. Leopold . 1960. The coastal plain at Nome, Alaska: A late Cenozoic type section for the Bering Strait region. Report of the 11th International Geological Congress 4:46–57.
- Hopkins, D. M. , P. A. Smith , and J. V. Matthews . 1981. Dated wood from Alaska and the Yukon: implications for forest refugia in Beringia. Quaternary Research 15:217–249.
- Hopkins, D. M. , J. V. Matthews Jr , C. E. Schweger , and S. B. Young , editors. 1982. Paleoecology of Beringia New York Academic Press. 1–489.
- http://www.radiocarbon.ldgo.columbia.edu/research/radiocarbon.htm. http://www.wrcc.dri.edu/summary/climsmak.html .
- Hultén, E. 1968. Flora of Alaska and Neighboring Territories Stanford Stanford University Press. 1–1008.
- Joint Federal-State Land Use Planning Commission for Alaska (JFSLUPCA) 1973. Major Ecosystems of Alaska Map Washington, D.C U.S. Geological Survey, scale 1: 2,500,000.
- Kaplan, J. O. , N. H. Bigelow , I. C. Prentice , S. P. Harrison , P. J. Bartlein , T. R. Christensen , W. Cramer , N. V. Matveyeva , A. D. Macguire , D. F. Murray , V. Y. Razzhivin , B. Smith , D. A. Walker , P. M. Anderson , A. A. Andreev , L. B. Brubaker , M. E. Edwards , and A. V. Lozhkin . 2003. Climate change and Arctic ecosystems: 2. Modeling, paleodata-model comparisons, and future projections. Journal of Geophysical Research 108:D-19 8171. doi:10.1029/2002JD002559.
- Kaufman, D. S. and D. M. Hopkins . 1986. Glacial history of the Seward Peninsula. In Hamilton, T. D. , K. M. Reed , and R. M. Thorson , editors. Glaciation in Alaska—The Geologic Record Anchorage Alaska Geological Society. 51–77.
- Kaufman, D. S. and W. F. Manley . 2004. Pleistocene and Late Wisconsin glacial extents across Alaska, U.S.A. In Ehlers, J. and P. L. Gibbard , editors. Quaternary Glaciations—Extent and Chronology, Part II. North America. Developments in Quaternary Science, Vol. 2 Amsterdam Elsevier. 9–27.
- Kurek, J. and L. C. Cwynar . 2007. A chironomid community perspective on paleoclimate: the Last Glacial Maximum and early Holocene in eastern Beringia. Program and Abstracts, Canadian Quaternary Association (CANQUA) Meeting, June 4–8, 2007, Ottawa, Ontario. p. 103.
- Kurtén, B. and E. Anderson . 1980. Pleistocene Animals of North America New York Columbia University Press. 1–442.
- Larsen, M. C. , C. H. Nelson , and D. R. Thor . 1981. Sedimentary processes and potential geologic hazards on the sea floor of the northern Bering Sea. In Hood, D. W. and J. A. Calder , editors. The Eastern Bering Sea Shelf: Oceanography and Resources U.S. Department of Commerce, National Oceanic and Atmospheric Administration. 247–261.
- Leslie, L. D. 1989. Alaska Climate Summaries. Alaska Climate Center Technical Note 5, Arctic Environmental Data Center Anchorage University of Alaska–Anchorage. 1–478.
- Lozhkin, A. V. , P. M. Anderson , T. V. Matrosova , and P. S. Minyuk . 2007. The pollen record from El'gygytgyn Lake: implications for vegetation and climate histories of northern Chukotka since the late middle Pleistocene. Journal of Paleolimnology 37:135–153.
- Mandryk, C. A. S. , H. Josenhans , D. W. Fedje , and R. W. Mathewes . 2001. Late Quaternary paleoenvironments of northwestern North America: implications for inland versus coastal migration routes. Quaternary Science Reviews 20:301–314.
- Manley, W. F. , D. S. Kaufman , and J. P. Briner . 2001. Pleistocene glacial history of the southern Ahklun Mountains, southwestern Alaska: soil development, morphometric, and radiocarbon constraints. Quaternary Science Reviews 21:997–1021.
- Mann, D. H. , D. M. Peteet , R. E. Reanier , and M. L. Kunz . 2002. Responses of an arctic landscape to Lateglacial and early Holocene climatic changes: the importance of moisture. Quaternary Science Reviews 21:997–1021.
- Mason, O. K. and J. W. Jordan . 2002. Minimal late Holocene sea level rise in the Chukchi Sea: arctic insensitivity to global change. Global and Planetary Changes 32:13–23.
- Matthews, J. V. Jr 1982. East Beringia during late Wisconsin time: a review of the biotic evidence. In Hopkins, D. M. , J. V. Matthews Jr , C. E. Schweger , and S. B. Young , editors. Paleoecology of Beringia New York Academic Press. 127–150.
- McCulloch, D. S. and D. M. Hopkins . 1966. Evidence for an early Recent warm interval in northwestern Alaska. Geological Society of America Bulletin 77:1089–1108.
- Moriya, K. 1978. Flora and Palynomorphs of Alaska Tokyo Kodansha Publishing Co. (in Japanese with English plate captions). 1–366.
- Muhs, D. R. , T. A. Ager , J. Been , J. P. Bradbury , and W. E. Dean . 2003. A late Quaternary record of eolian silt deposition in a maar lake, St. Michael Island, western Alaska. Quaternary Research 60:110–122.
- Nelson, C. H. 1982. Late Pleistocene–Holocene transgressive sedimentation in deltaic and non-deltaic areas of the northeastern Bering epicontinental shelf. Geologie en Mijnbouw 61:5–18.
- Olsen, H. W. , E. C. Clukey , and C. H. Nelson . 1982. Geotechnical characteristics of bottom sediment in the northeastern Bering Sea. Geologie en Mijnbouw 61:91–103.
- Oswald, W. W. , L. B. Brubaker , and P. M. Anderson . 1999. Late Quaternary vegetational history of the Howard Pass area, northwestern Alaska. Canadian Journal of Botany 77:570–581.
- Péwé, T. L. 1975. Quaternary geology of Alaska. U.S. Geological Survey Professional Paper 835:1–145.
- Ray, V. M. and W. R. Dupré . 1981. The ice-dominated regimen of Norton Sound and adjacent areas of the Bering Sea. In Hood, D. W. and J. A. Calder , editors. The Eastern Bering Sea Shelf: Oceanography and Resources U.S. Department of Commerce, National Oceanic and Atmospheric Administration. 263–278.
- Repenning, C. A. 1980. Faunal exchanges between Siberia and North America. Canadian Journal of Anthropology 1:37–44.
- Sharma, G. D. 1979. The Alaskan Shelf: Hydrographic, Sedimentary, and Geochemical Environment New York Springer-Verlag. 1–498.
- Thor, D. R. and C. H. Nelson . 1981. Ice gouging on the subarctic Bering Shelf. In Hood, D. W. and J. A. Calder , editors. The Eastern Bering Sea Shelf: Oceanography and Resources, Vol. 1 U.S. Department of Commerce, National Oceanic and Atmospheric Administration. 279–291.
- Velichko, A. A. , L. L. Isayeva , V. M. Makeyev , G. G. Matishov , and M. A. Faustova . 1984. Late Pleistocene glaciation of the arctic shelf, and the reconstruction of Eurasian ice sheets. In Velichko, A. A. , H. E. Wright Jr , and C. W. Barnosky , editors. Late Quaternary Environments of the Soviet Union Minneapolis University of Minnesota Press. 35–41.
- Viereck, L. A. and E. L. Little Jr . 1972. Alaska Trees and Shrubs Washington, D.C U.S. Department of Agriculture Forest Service Handbook 410. 1–265.
- Viereck, L. A. and E. L. Little Jr . 1975. Atlas of United States Trees. Vol. 2, Alaska Trees and Common Shrubs Washington, D.C U.S. Department of Agriculture Forest Service Miscellaneous Publication 1293. 1–19. , 82 plates.
- West, F. H. , editor. 1996. American Beginnings—The Prehistory and Paleoecology of Beringia Chicago University of Chicago Press. 1–576.
- Yurtsev, B. A. 2001. The Pleistocene “tundra-steppe” and the productivity paradox: the landscape approach. Quaternary Science Reviews 20:165–174.
- Zazula, G. D. , C. E. Schweger , A. B. Beaudoin , and G. H. McCourt . 2006. Macrofossil and pollen evidence for full-glacial steppe within an ecological mosaic along the Bluefish River, eastern Beringia. Quaternary International 142–143:2–19.
- Zazula, G. D. , D. G. Froese , S. A. Elias , S. Kuzmina , and R. W. Mathewes . 2007. Arctic ground squirrels in the mammoth-steppe: paleoecology of late Pleistocene middens (∼24 000–29 450 14C yr BP), Yukon Territory, Canada. Quaternary Science Reviews 26:979–1003.