Abstract
In alpine lakes from several regions of the world, sedimentary diatom profiles indicate that rapid shifts in diatom community structure have occurred over the past century. A number of these recent shifts have been attributed to anthropogenic disturbances such as enhanced atmospheric nitrogen (N) deposition or climate change. When these disturbances are coupled, the response of alpine lakes becomes more complex and varies from region to region. The Beartooth Mountain Range, situated on the border between Montana and Wyoming, is part of the central Rocky Mountains; it is considered an area of relatively low N deposition but has experienced an increase in bulk precipitation rates, primarily in the form of snowfall, over the past century. We have examined a 400-yr sediment record from Beartooth Lake and have observed a rapid change in the diatom community structure over the past decade. A typical alpine lake diatom flora, consisting mainly of small Fragilaria sensu lato species, dominated this lake until approximately 1995, at which time Fragilaria crotonensis and Cyclotella bodanica var. lemanica rapidly increased to approximately 30% each of the total assemblage. The diatom assemblages from the tops and bottoms of short cores from three additional lakes in the area also reveal taxonomic shifts. These shifts appear indicative of both increased N loading to these systems as well as changes in thermal stratification patterns.
Introduction
Diatoms are a class of microscopic algae that are sensitive to environmental changes and that produce siliceous fossils that are frequently well preserved in lacustrine sediments. Shifts in diatom assemblage composition are frequently the first indication of ecological perturbations. Sedimentary diatom profiles have been used in the reconstruction of limnological parameters such as lake-water pH (e.g., CitationCharles and Whitehead, 1986; CitationRenberg, 1990; CitationAnderson and Renberg, 1992; CitationBattarbee et al., 1999) and phosphorus concentrations (e.g., CitationAnderson et al., 1990; CitationAnderson and Rippey, 1994; CitationHall and Smol, 1996), as well as climate-related variables, such as salinity in closed-basin lakes (e.g., CitationFritz et al., 1991; CitationMetcalfe, 1995; CitationLaird et al., 1996), dissolved organic carbon concentrations in lakes near treeline (CitationMacDonald et al., 1993), and epilimnetic temperature (e.g., CitationWeckström et al., 1997; CitationJoynt and Wolfe, 2001).
In alpine lakes from several regions of the world, sedimentary diatom profiles indicate that rapid shifts in diatom community structure have occurred over the past century (Arzet, 1987; CitationPsenner and Schmidt, 1992; CitationDrake and Naiman, 2000; CitationWolfe et al., 2001). While these rapid changes are widespread, the mechanisms driving them appear to vary considerably from region to region. These recent shifts in diatom community structure have been attributed to perturbations such as fish stocking, enhanced atmospheric nitrogen (N) deposition (via nutrient enrichment and/or pH changes), climate change, or a combination of these disturbances (CitationPsenner and Schmidt, 1992; CitationSchmidt and Psenner, 1992; CitationSommaruga-Wögrath et al., 1997; CitationDrake and Naiman, 2000; CitationWolfe et al., 2001). Although the nature of the disturbance varies, these alpine lake communities are clearly undergoing reorganization.
Of the major disturbances affecting alpine lakes, enhanced atmospheric N deposition is one of the primary concerns, as human alteration of the global N cycle has doubled the amount of fixed N (predominantly in the forms NH4 + and NO3 −) transferred from the atmosphere to land-based ecosystems (CitationVitousek et al., 1997). Because of atmospheric transport, anthropogenic N can affect systems far removed from the source of emission, as indicated by increased NO3 − concentrations in the Greenland ice sheet (CitationMayewski et al., 1990). Although the increased quantity of fixed N in the atmosphere is now well documented, the extent to which atmospheric N deposition alters nutrient concentrations and ratios in aquatic systems is presently unclear, in part because most aquatic ecosystems also receive nutrient inputs via agricultural and/or urban runoff.
The response of alpine lakes to enhanced N deposition becomes more complex when coupled with changes in climate, and thus varies from region to region. Alpine lakes are sensitive to climate change. For example, an apparent coupling of pH and air temperatures has been observed in lakes of the eastern Alps (CitationPsenner and Schmidt, 1992; CitationSommaruga-Wögrath et al., 1997). Higher mean air temperatures are correlated with higher pH in these lakes, likely relating to enhanced weathering rates and increased base cation supply, and/or stimulated within-lake (microbial) alkalinity production (CitationSchindler et al., 1986). This may explain why recent increases in acidic, inorganic N deposition from industrial sources have not resulted in higher dissolved inorganic N (DIN) concentrations and attendant decreased lake-water pH; the limnological response to current climate warming has thus far offset the impacts of acid deposition. Similarly, in the southern Rocky Mountains, despite the prediction of acidification from atmospheric deposition (CitationKling and Grant, 1984; CitationCaine, 1995), recent paleolimnological analyses indicate no diatom evidence whatsoever of decreased lake pH (CitationWolfe et al., 2001). However, CitationWilliams et al. (1996) have recorded episodic decreases in pH and alkalinity in lakes of the Niwot Ridge in the Colorado Front Range. In contrast, modern limnological surveys in the central Rocky Mountains show no evidence of acidification occurring in that region (CitationMiller and Bellini, 1996; CitationTheriot et al., 1997).
Due to the geographic variability of the interplay between climate change and enhanced atmospheric N deposition, alpine lakes in different regions will not likely respond uniformly to these changes. In lakes of the southern Rocky Mountains (e.g., Colorado Front Range), shifts in fossil diatom assemblages intensified during the 1950s (according to 210Pb chronology), at a sediment depth of 5 cm below the mud-water interface (CitationWolfe et al., 2001). The timing of these shifts coincides with the widespread implementation of the Haber-Bosch process for the production of commercial N fertilizers. Rates and magnitudes of recent ecological shifts in this area far exceed those of any natural, preindustrial changes (CitationBaron et al., 2000; CitationWolfe et al., 2001). Here we report the recent shifts in diatom community structure observed in the Beartooth Range, which is part of the central Rocky Mountains . As indicated in , the Beartooth Range is an area with much lower N deposition rates than the Colorado Front Range. Diatom shifts in the fossil records of Beartooth lakes are discussed in reference to recent changes in climate and N deposition patterns.
Site Description
The Beartooth Mountains lie to the northeast of Yellowstone National Park, along the border of Montana and Wyoming, and comprise a large section of the Absaroka-Beartooth Wilderness Area . The geology of the region is dominated by Precambrian granitic rock, which was uplifted 50 million years ago, and is some of the oldest exposed crust (2.8–3.2 billion years) on the continent. The area is characterized by sweeping alpine plateaus and large U-shaped glacial valleys.
Over 600 permanent lakes have been identified in the 900-km2 Beartooth primitive area. As a result of the slow-weathering bedrock, lakes in the region generally have low silica and phosphorus levels. Nitrate concentrations are also low in these lakes . Many of the lakes in the Beartooth area lie above 2500 m, with treeline ranging between 2750 and 3000 m. As a result, many of the lakes in forested areas sit in basins that may have been barren of forest vegetation in the recent past. Many of the lakes of the region were historically fishless, but fish have been introduced over the past century to virtually all of the drainages on the Beartooth Plateau. Records from the Wyoming Game and Fish Department indicate that stocking in most of these lakes began in the 1920s.
Methods
In September of 1999, a 40-cm core of organic sediments was collected from Beartooth Lake (Park County, Wyoming) with a modified K-B gravity corer (CitationGlew, 1989) and extruded in consecutive 0.5-cm increments in the field. The diatom species composition of 14 samples was determined (as described below), resulting in a diatom profile for this lake for the past 400 yr. The chronology of recent sediments was based on 210Pb distillation and alpha spectrometry methods (modified from CitationEakins and Morrison, 1978), and dates and sedimentation rates were determined according to the constant flux:constant sedimentation rate (CF:CSR) model (CitationOldfield and Appleby, 1984). The results are essentially the same as those from the Constant Rate of Supply model, but the CF:CSR model provides a more conservative sedimentation rate. The stratigraphy of 210Pb activity also verifies the stratigraphic integrity of the core.
Short cores were also extracted in July 2000 from three additional lakes in the Beartooth Mountains: Beauty, Emerald, and Long (all in Park County, Wyoming). Sediment samples from only the top and bottom of each core were collected, to assess whether any change in diatom stratigraphy had occurred. The bottom depth of each core was Beauty, 12 cm; Emerald, 21 cm; Long, 26 cm. Because complete cores were not collected, the 210Pb dates of these sediment samples were not determined. While the dates for each bottom sample will differ, this “top and bottom” sediment core approach (CitationDixit and Smol, 1994) is simply used to document whether any shift in community structure has occurred in the past 100–200 yr.
Diatom slides were prepared from digested (30% H2O2) sediment samples. The processed samples were settled onto coverslips and mounted onto slides with Naphrax®. A minimum of 300 valves per slide was counted under oil immersion on a Nikon E400 Eclipse microscope with phase-contrast under oil-immersion at 1000× magnification. Diatom taxonomy was based primarily on CitationKrammer and Lange-Bertalot (1986–1991) and CitationCamburn and Charles (2000).
Carbon (C) and N content of lake sediments were determined on freeze-dried samples by combustion and gas chromatography on an elemental analyzer (Carlo Erba 1106).
Results
The 210Pb dates reveal that the 40-cm core from Beartooth Lake captures the last 400 yr of sediment deposition (, ). The top 15 cm provide a record for the past 100 yr in this lake. An average sedimentation rate of 1 cm per 10 yr is common in alpine lakes situated at or above treeline, where sedimentation rates are typically low.
A total of 27 diatom taxa were found in the sediment core from Beartooth Lake, with a profile of the major taxa displayed in . The profile indicates that prior to the 1990s, a typical alpine diatom flora dominated in this system. The assemblage consisted mainly of small Fragilaria sensu lato species (i.e., Staurosira and Staurosirella sensu; CitationRound et al., 1990), including F. construens (Ehr.) Grun., F. construens v. venter (Ehr.) Grun., and F. pinnata Ehr., all of which are common in systems situated at high elevations or high latitudes (CitationLotter et al., 1999). However, from 1995 to the present, a shift in fossil assemblages is apparent, with Fragilaria crotonensis Kitton and Cyclotella bodanica var. lemanica (O. Müller ex. Schröter) Bachmann rapidly increasing to comprise approximately 30% each of the total assemblage. Both of these taxa were found sporadically further down the core, but never comprised more than 1% of the total assemblage until the 1990s. Small Fragilaria species were still abundant in these samples, but comprised a much smaller fraction of these assemblages.
Given the relatively constant rate of total sediment accumulation , C and N profiles in Beartooth Lake sediments indicate a recent increase in the rate of organic sedimentation corresponding with the recent changes in diatom flora. Virtually all the C in the sediments of these lakes was organic C, as measured by incremental loss on ignition at 550 °C and 1100 °C (CitationDean, 1974). Sediments below 10 cm in the Beartooth Lake core were typically 3–4% C and 0.3–0.4% N by weight, while surface sediments had more than double these fractions of C and N. Sediment C:N atomic ratios ranged from 10 to 12 in the upper 20 cm of the core, indicating primarily autochthonous sources of organic material and suggesting increased in-lake productivity through the past century. This interpretation is further supported by the fact that hypolimnetic waters in Beartooth Lake remain cold and oxygenated throughout the year, minimizing long-term variability in diagenetic processes that might otherwise complicate relationships between productivity and rates of organic matter accumulation in lake sediments (CitationMeyers, 1997).
The relative differences between the fossil diatom assemblages of the tops and bottoms of the cores from Beauty, Emerald, and Long Lakes reveal that a number of changes have occurred relatively recently in these lakes. Small Fragilaria species have declined in abundance in all three lakes, although they still remain common in Long Lake (comprising approximately 30% of the modern assemblage). Asterionella formosa Hassall comprised a larger fraction of the modern assemblages in both Long and Emerald Lakes, while Cyclotella rossi (Grun.) Håkansson and Tetracyclus glans (Ehr.) Mills increased in abundance in Beauty Lake. Aulacoseira lirata (Ehr.) Ross and Cyclotella stelligera Cleve & Grun. showed less consistent trends, as each increased in abundance in some cases but declined in others.
Discussion
The diatom fossil records from these four lakes in the Beartooth Mountains indicate that these communities have recently undergone changes. In the case of Beartooth Lake, with a complete record available for analysis, this community shift appears rapidly and very recently (within the last decade). In all cases, small Fragilaria species, which are typical alpine lake taxa, have declined. In contrast, taxa such as Fragilaria crotonensis, Asterionella formosa, and a number of Cyclotella species have increased.
It is currently unclear which factors are driving these shifts in diatom communities. With the shift in diatom taxa occurring within the last decade in Beartooth Lake, this is not likely the result of fish stocking, which was initiated in the 1920s in this lake. However, while the Beartooth Mountains are an area of relatively low N deposition , bulk precipitation rates have increased over the past century, primarily in the form of snowfall . This increase in precipitation has contributed to an increase in the rate of N deposition in this area. Higher precipitation rates can also enhance weathering of nutrients, such as silica and phosphorus, and base cations from the watershed (CitationWhite and Blum, 1995). While bulk precipitation rates began to increase in the middle of the last century, the observed shifts in the diatom community did not occur until the mid-1990s. This lag may be due to lower rates of N deposition in the Beartooth Mountains relative to areas further south , as well as to the nearly continuous vegetation cover of the Beartooth Lake watershed, which provides a transient sink for N derived from precipitation. In contrast, CitationWolfe et al. (2001) observed rapid responses in the diatom communities of two lakes of the Colorado Front Range to have occurred several decades earlier. Not only is this is an area of higher N deposition, the investigated lakes occupy poorly vegetated catchments dominated by talus, implying limited terrestrial sinks for excess N. This confirms the importance of edaphic factors in regulating the timing and extent of limnological shifts associated with atmospheric deposition.
Based on observations of the pH tolerance ranges for the taxa in these sediment records, the shifts in diatom community structure in this area do not indicate pH changes, as most of the taxa found throughout the cores are circumneutral (CitationDixit et al., 1999; CitationCamburn and Charles, 2000). However, these community shifts, as well as the C and N data, are consistent with altered nutrient dynamics in these systems. This is supported by the decline in small taxa, which are typically good competitiors for nutrients due to their high surface area to volume ratio (CitationReynolds, 1984), and the increase in larger taxa, particularly those such as Fragilaria crotonensis and Asterionella formosa, which are typically found in mesotrophic systems. Both F. crotonensis and A. formosa have also increased in abundance in lakes of the Colorado Front Range; CitationWolfe et al. (2001) have inferred that these shifts are the result of enhanced atmospheric N deposition in this area, as the distribution of F. crotonensis is associated with high N:P ratios (CitationInterlandi et al., 1999) and the growth of A. formosa from alpine lakes has been stimulated by N additions (CitationMcKnight et al., 1990; CitationYang et al., 1996).
The resource physiologies of the Cyclotella taxa in these sediment records are not well documented. In large lakes of the Greater Yellowstone Ecosystem, Cyclotella bodanica Grun. is considered a low N specialist (CitationInterlandi et al., 1999), and in general, Cyclotella species are considered good competitors for N. Hence, the increase in these taxa appears inconsistent with increased N loading to these systems. However, it may be indicative of changes in stratification patterns or shifts in light:nutrient ratios. Cyclotella bodanica has high light requirements (CitationInterlandi et al., 1999) and can regulate its buoyancy, which allows it to remain in the epilimnion during stratification. CitationBradbury (1988) also found that this taxon was favored when summer stratification was strong in Elk Lake (Clearwater County, Minnesota). The recent increase in generally larger taxa is also consistent with longer ice-free periods, as small diatoms typically have faster growth rates than larger taxa and thus have an advantage during short growing seasons. A short growing season would also favor benthic and tychoplanktonic forms over planktonic taxa, in that these forms can grow in the shallow margins of lakes, which are the first areas to thaw in spring (CitationSmol, 1988).
While it is currently unclear which factors are driving these shifts in diatom communities, these communities are clearly undergoing changes in lakes of the central Rocky Mountains. These alpine lakes appear highly sensitive to anthropogenic disturbances; our results suggest that the shifts in diatom community structure may be a result of the synergistic effects of nutrient deposition and climate change. We are currently exploring the resource physiologies of the common diatom taxa in this area in an effort to understand the mechanisms behind these changes.
FIGURE 1. Location of the Beartooth Mountain Range, situated in the central Rocky Mountains of North America. Monitoring stations for the National Atmospheric Deposition Program (NADP) are indicated with black circles; data collected at these locations (CitationNADP/NTN 1984–1996) were used to draw the total nitrogen deposition contours. Nitrogen deposition patterns reveal that the central Rocky Mountains are an area of lower total N deposition than the southern Rockies.
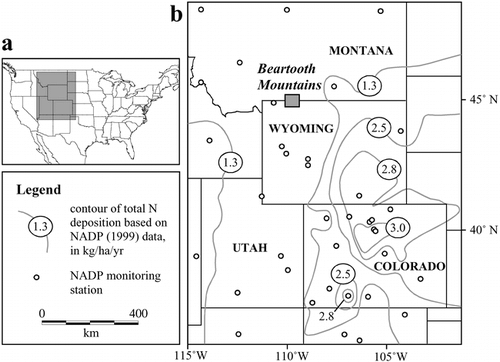
FIGURE 2. Stratigraphic plot of 210Pb activity in the sediment core from Beartooth Lake, and the age-depth model based on the CF:CSR model.
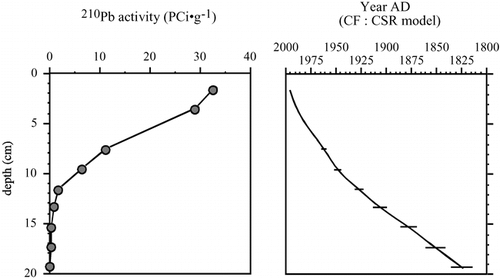
FIGURE 3. Diatom profile, %C, and %N of the sediments from Beartooth Lake, Wyoming. Small Fragilaria species (F. construens, F. construens v. venter, and F. pinnata) are grouped together, and consistently comprise >65% of the assemblages until the top 1.5 cm of the core, at which point they drop to ∼30% of the assemblage. Cyclotella bodanica v. lemanica and Fragilaria crotonensis, both present earlier in the core, each comprise 30% of the assemblages in the top 1.5 cm of the core. 210Pb dating indicates that the top 2 cm of sediment have been deposited since 1995.
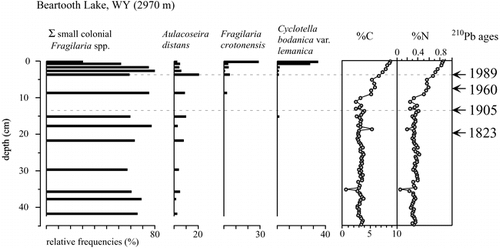
FIGURE 4. Change in the relative frequencies of diatom taxa in the tops and bottoms of cores from three lakes in the Beartooth Mountains. A positive value indicates that this taxon comprises a larger fraction of the modern assemblage when compared to the fossil assemblage at the bottom of the core.
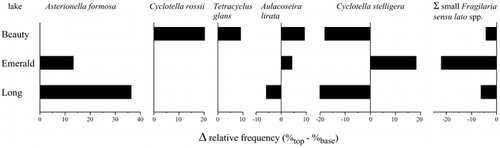
FIGURE 5. Precipitation measurements recorded at Red Lodge, Montana, over the past century, displayed as: (A) total annual; (B) winter; (C) summer. Spline curves are fit to the data with a lambda value of 1000. Total annual precipitation increased over this period, primarily due to increases in winter precipitation. Data are from the Western Regional Climate Center (http://www.wrcc.sage.dri.edu/summary/climsmmt.html).
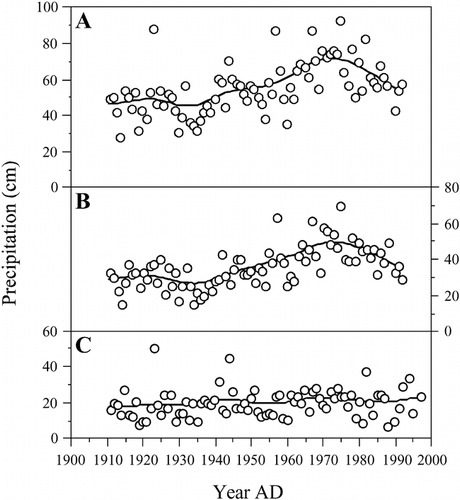
TABLE 1 Dissolved nutrient concentrations measured in a set of lakes from the Beartooth Range in July 2000a
Acknowledgments
We are grateful for the field assistance provided by Misa Saros, Barbara Interlandi, and David Dean. This research is supported by a grant from the National Science Foundation (DEB 0089600). Additional support was provided by a Faculty Research Grant from the University of Wisconsin–La Crosse to JES.
Related Research Data
References Cited
- Anderson, N. J. and I. Renberg . 1992. A paleolimnological assessment of diatom production responses to lake acidification. Environmental Pollution 78:113–119.
- Anderson, N. J. and B. Rippey . 1994. Monitoring lake recovery from point-source eutrophication: the use of diatom-inferred epilimnetic total phosphorus and sediment chemistry. Freshwater Biology 32:625–639.
- Anderson, N. J. , B. Rippey , and A. C. Stevenson . 1990. Change to a diatom assemblage in a eutrophic lake following point source nutrient re-direction: a paleolimnological approach. Freshwater Biology 23:205–217.
- Arzet, K. 1997. Diatomeen als pH-Indikatoren in subrezenten Sedimenten von Weichwasserseen. Dissertation Universität Innsbruck. 266. pp.
- Battarbee, R. W. , D. F. Charles , S. S. Dixit , and I. Renberg . 1999. Diatoms as indicators of surface water acidity. In Stoermer, E.F., and Smol, J.P. (eds.), The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge: Cambridge University Press. 85–127.
- Baron, J. S. , H. M. Rueth , A. P. Wolfe , K. R. Nydick , E. J. Allstott , J. T. Minear , and B. Moraska . 2000. Ecosystem responses to nitrogen deposition in the Colorado Front Range. Ecosystems 3:352–368.
- Bradbury, J. P. 1988. A climatic-limnologic model of diatom succession for paleolimnological interpretation of varved sediments at Elk Lake, Minnesota. Journal of Paleolimnology 1:115–131.
- Caine, N. 1995. Temporal trends in the quality of streamwater in an alpine environment: Green Lakes Valley, Colorado Front Range, U.S.A. Geografiska Annaler 77A:207–220.
- Camburn, K. E. and D. F. Charles . 2000. Diatoms of low-alkalinity lakes in the Northeastern United States. Academy of Natural Sciences of Philadelphia Special Publication 18. 152. pp.
- Charles, D. F. and D. R. Whitehead . 1986. The PIRLA project: Paleoecological investigation of recent lake acidification. Hydrobiologia 143:13–20.
- Dean, W. E. 1974. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparisons with other methods. Journal of Sediment Petrology 44:242–248.
- Dixit, S. S. and J. P. Smol . 1994. Diatoms as indicators in the Environmental Monitoring and Assessment Program–Surface Waters (EMAP-SW). Environmental Monitoring and Assessment 31:275–306.
- Dixit, S. S. , J. P. Smol , D. F. Charles , R. M. Hughes , S. G. Paulsen , and G. B. Collins . 1999. Assessing water quality changes in the lakes of the northeastern United States using sediment diatoms. Canadian Journal of Fisheries and Aquatic Sciences 56:131–152.
- Drake, D. C. and R. J. Naiman . 2000. An evaluation of restoration efforts in fishless lakes stocked with exotic trout. Conservation Biology 14:1807–1820.
- Eakins, J. D. and R. T. Morrison . 1978. A new procedure for the determination of lead-210 in lake and marine sediments. International Journal of Applied Radiation and Isotopes 29:531–536.
- Fritz, S. C. , S. Juggins , R. W. Battarbee , and D. R. Engstrom . 1991. Reconstruction of past changes in salinity and climate using a diatom-based transfer function. Nature 352:706–708.
- Glew, J. R. 1989. A new trigger mechanism for sediment samplers. Journal of Paleolimnology 2:241–243.
- Hall, R. I. and J. P. Smol . 1996. Paleolimnological assessment of long-term water-quality changes in south-central Ontario lakes affected by cottage development and acidification. Canadian Journal of Fisheries and Aquatic Sciences 53:1–17.
- Interlandi, S. J. , S. S. Kilham , and E. C. Theriot . 1999. Responses of phytoplankton to varied resource availability in large lakes of the greater Yellowstone ecosystem. Limnology and Oceanography 44:668–682.
- Joynt III, E. H. and A. P. Wolfe . 2001. Paleoenvironmental inference models from sediment diatom assemblages in Baffin Island lakes (Nunavut, Canada) and reconstruction of summer water temperature. Canadian Journal of Fisheries and Aquatic Sciences 58:1222–1243.
- Kling, G. W. and M. C. Grant . 1984. Acid precipitation in the Colorado Front Range: an overview with time predictions for significant effects. Arctic and Alpine Research 16:321–329.
- Krammer, K. and H. Lange-Bertalot . 1986–1991. Bacillariophyceae. In Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., and Mollenhauer, D. (eds.), Süßwasserflora von Mitteleuropa. Vols. 2(1–4). Stuttgart/Jena: Gustav Fischer Verlag.
- Laird, K. R. , S. C. Fritz , K. A. Maasch , and B. F. Cumming . 1996. Greater drought intensity and frequency before AD 1200 in the Northern Great Plains, USA. Nature 384:552–555.
- Lotter, A. F. , R. Pienitz , and R. Schmidt . 1999. Diatoms as indicators of environmental change near arctic and alpine treeline. In Stoermer, E.F., and Smol, J.P. (eds.), The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge: Cambridge University Press. 205–226.
- MacDonald, G. M. , T. W. D. Edwards , K. A. Moser , R. Pienitz , and J. P. Smol . 1993. Rapid response of treeline vegetation and lakes to past climate warming. Nature 361:243–246.
- Mayewski, P. A. , W. B. Lyons , M. J. Spencer , M. S. Twickler , C. F. Buck , and S. Whitlow . 1990. An ice-core record of atmospheric response to anthropogenic sulfate and nitrate. Science 346:554–556.
- McKnight, D. M. , R. L. Smith , J. P. Bradbury , J. S. Baron , and S. Spaulding . 1990. Phytoplankton dynamics in three Rocky Mountain lakes, Colorado, U.S.A. Arctic and Alpine Research 22:264–274.
- Metcalfe, S. E. 1995. Holocene environmental change in the Zacapu Basin, Mexico: a diatom-based record. The Holocene 5:196–208.
- Meyers, P. A. 1997. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes. Organic Geochemistry 27:213–250.
- Miller, W. and M. Bellini . 1996. Trophic state evaluation of selected lakes in Grand Teton National Park. Unsolicited Proposal Program, University of Wyoming, National Park Service Research Station. 111. pp.
- NADP/NTN [National Atmospheric Deposition Program/National Trans Network] 1984–1996. NADP/NTN annual data summary, precipitation chemistry in the United States. NADP/NTN Coordination Office, Natural Resource Ecology Laboratory. Fort Collins: Colorado State University.
- Oldfield, F. and P. G. Appleby . 1984. Empirical testing of 210Pb-dating models for lake sediments. In Haworth, E.Y., and Lund, J.W.G. (eds.), Lake Sediments and Environmental History. Minneapolis: University of Minnesota Press. 93–124.
- Psenner, R. and R. Schmidt . 1992. Climate-driven pH control of remote alpine lakes and effects of acid deposition. Nature 356:781–783.
- Renberg, I. 1990. A 12000 year perspective of the acidification of Lilla Oresjon, southwest Sweden. Philosophical Transactions of the Royal Society of London, series B 327:357–361.
- Reynolds, C. S. 1984. The Ecology of Freshwater Phytoplankton. Cambridge: Cambridge University Press. 384. pp.
- Round, F. E. , R. M. Crawford , and D. G. Mann . 1990. The Diatoms: Biology and Morphology of the Genera. Cambridge: Cambridge University Press. 747. pp.
- Schindler, D. W. , M. A. Turner , M. P. Stainton , and G. A. Linsey . 1986. Natural sources of acid-neutralizing capacity in low-alkalinity lakes of the Precambrian Shield. Science 232:844–847.
- Schmidt, R. and R. Psenner . 1992. Climate changes and anthropogenic impacts as causes for pH fluctuations in remote high alpine lakes. Documenta Istituto Italiano Idrobiologia 32:31–57.
- Smol, J. P. 1988. Paleoclimate proxy data from freshwater arctic diatoms. Verhandlungen der Internationalen Vereinigung von Limnologen 23:837–844.
- Sommaruga-Wögrath, S. , K. Koinig , R. Schmidt , R. Sommaruga , R. Tessandri , and R. Psenner . 1997. Temperature effects on the acidity of remote alpine lakes. Nature 387:64–67.
- Theriot, E. C. , S. C. Fritz , and R. W. Gresswell . 1997. Long term limnological data from the larger lakes of Yellowstone National Park. Arctic and Alpine Research 29:304–314.
- Vitousek, P. M. , J. D. Aber , R. W. Howarth , G. E. Likens , P. A. Matson , D. W. Schindler , W. H. Schlesinger , and D. G. Tilman . 1997. Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications 7:737–750.
- Weckström, J. , A. Korhola , and T. Blom . 1997. Diatoms as quantitative indicators of pH and water temperature in subarctic Fennoscandian lakes. Hydrobiologia 347:171–184.
- White, A. F. and A. E. Blum . 1995. Effects of climate on chemical weathering in watersheds. Geochimica et Cosmochimica Acta 59:1729–1747.
- Williams, M. W. , M. Losleben , N. Caine , and D. Greenland . 1996. Changes in climate and hydrochemical responses in high-elevation catchment, Rocky Mountains. Limnology and Oceanography 41:939–946.
- Wolfe, A. P. , J. S. Baron , and R. J. Cornett . 2001. Anthropogenic nitrogen deposition induces rapid ecological changes in alpine lakes of the Colorado Front Range (USA). Journal of Paleolimnology 25:1–7.
- Yang, J. R. , F. R. Pick , and P. B. Hamilton . 1996. Changes in the planktonic diatom flora of a large mountain lake in response to fertilization. Journal of Phycology 32:232–243.