Abstract
Diatoms from Sawtooth Lake (79°20′N, 81°51′W) on the Fosheim Peninsula in Central Ellesmere Island, Canada were analyzed to assess the temporal extent and magnitude of climatic change in the High Arctic during the late Holocene. Diatom results from the sediment cores show an absence of diatoms throughout the last ∼2.5 ka (4.6 m) until the 1920s. However, ca. 1926 (5.3-cm depth), a rapid colonization of diatoms in the lake occurred. Within the uppermost section of the core (∼1920 to ∼1997), the diatom flora shift from a small Fragilaria-dominated assemblage to a more diverse assemblage that is dominated by large planktonic taxa (e.g., Cyclotella bodanica) and large raphid benthic species. The postglacial nature of this assemblage suggests a decrease in ice cover and a concomitant increase in light and nutrient availability for diatom growth over the last ∼75 yr. Of particular significance is this absence of diatoms prior to the ∼1920s, which indicates that environmental conditions of the last ∼75 yr are unlike any of the previous ∼2500 yr.
Introduction
Holocene climate fluctuations and recent warming have been reconstructed from a number of proxy climate data sets in the Northern Hemisphere. These reconstructions show pronounced warming in the last 50 yr, which is unprecedented in relation to the last 1000 yr (CitationMann et al., 1999). While these reconstructions provide much-needed high-resolution data for climate variability before the availability of instrumental data, it is necessary to push this application further back in time to expand our knowledge of natural climate variability throughout the Holocene, thereby placing the magnitude and rate of change in recent years within a longer perspective. This need is especially true in the High Arctic, where global circulation models (GCMs) predict an amplified response to greenhouse gas-induced warming (CitationZwiers, 2002), yet few long-term high-resolution climate data sets exist.
In the Arctic, recent environmental changes have been documented from many paleoclimatic records. These changes are manifested in the increased thickness of lacustrine varves (CitationSmith, 1997; CitationHughen et al., 2000), in recent shifts in fossil algal assemblages preserved in lacustrine sediments (CitationDouglas et al., 1994; CitationDoubleday et al., 1995; CitationGajewski et al., 1997; CitationWolfe, 2000; CitationWolfe and Perren, 2001; CitationSorvari et al., 2002), and from other high-resolution paleoclimatic proxy records (reviewed in CitationOverpeck et al., 1997). These changes are thought to represent recent warming in the Arctic that is unprecedented in the context of recent centuries. The warming is consistent with GCM predictions of climate expected under higher CO2 levels, suggesting that it is, at least in part, the result of anthropogenic influences on global climate.
In this paper, we present a late Holocene paleolimnological record from the sediments of Sawtooth Lake, Ellesmere Island, Nunavut, Canada. The purpose of this study is to determine the nature of recent environmental change from the biological record contained within the sediments by a detailed, high-resolution analysis of the fossil diatoms.
ECOLOGICAL RESPONSE TO CLIMATE CHANGE IN THE HIGH ARCTIC
The development of calibration sets for circumarctic sites (CitationPienitz et al., 1995; CitationLim et al., 2001; CitationWeckström and Korhola, 2001; CitationJoynt and Wolfe, 2001) is rapidly changing the applicability of quantitative models to both high and low arctic sites; however, because of limited autecological information in the High Arctic, hindcasting climatic trends using diatoms has relied historically on qualitative models of their inferred responses to climatic change (CitationSmol, 1983). Because high arctic lacustrine environments are dominated by low temperatures, thick and persistent ice cover, limited light, and a short (often less than 8 wk) growing season, a reduction in ice cover is expected to be the major response of arctic lakes to climatic warming (CitationDouglas and Smol, 1999). A reduction in ice cover has many implications for the aquatic ecology of arctic lakes (CitationRouse et al., 1997; CitationDouglas and Smol, 1999, CitationBattarbee, 2000). It promotes an increased diversity of habitats for autotrophic species, such as diatoms, by increasing light in the euphotic zone for plankton growth, and allows the littoral zone to be colonized by mosses and thus epiphytic species. A longer growing season allows these species to establish themselves and reproduce for longer periods of time, thereby increasing productivity and ecological complexity. In addition, warmer climates accelerate weathering in the watershed. This increased buffering releases more nutrients and dissolved solids into the lake, which augment primary productivity (CitationDouglas and Smol, 1999).
ICE COVER MODEL
The main model used in the Arctic relates diatom species assemblages to changes in ice cover (CitationSmol, 1983). As warming occurs, the ice pan on the lake shrinks, thereby allowing more photosynthetically active radiation to penetrate the lake. The moat (the area between the shore and the ice pan) becomes wider, allowing for the proliferation of deeper water and epiphytic taxa. Convective circulation in the ice-free areas and increased nutrients allow for larger phytoplankton to thrive in the euphotic zone and for larger epiphytic taxa to colonize the littoral zone of the lake. Thus, with warming, a shift from small shallow-water taxa (e.g., Fragilaria pinnata) to larger epiphytic benthic species (e.g., Navicula spp.) and planktonic taxa (e.g., Cyclotella spp., Stephanodiscus spp.) should be apparent (CitationLotter and Bigler, 2000). Indeed, this species transition is also a noted sequence in the glacial/postglacial ontogeny of lakes at lower latitudes (reviewed in CitationRound, 1981).
While this model provides the means by which diatom records can be interpreted, it cannot be directly tied to any single meteorological variable, which is necessary for the use of a transfer function. Ice cover does relate to summer warmth (CitationDoran et al., 1996; CitationSchindler et al., 1996), but not linearly. Several factors such as winter temperature, windiness, and internal lacustrine thermal feedbacks complicate the relationship (CitationDoran et al., 1996), as do changes in snow depth, which affect sub-ice circulation, diatom suspension, and photosynthesis (CitationKelley, 1997). In addition, caution must be used in ascribing changes solely to ice cover. A reduction in ice cover is merely one factor among others, such as the increases in nutrient supply, light, and habitat availability that result from climatic warming.
Study Area
Sawtooth Lake (79°20′N, 83°51′W, 280m a.s.l.) is the highest of a chain of three lakes situated within a col in the Sawtooth Mountain Range on the Fosheim Peninsula in central Ellesmere Island (). The surrounding unglacierized landscape is considerably high in relief, with the steep-sided, east-west-trending valley walls reaching 1000 m in elevation. It is a large (260 ha), deep (>90 m) lake that retains considerable ice cover during the summer months and is rarely ice free.
The prevalence of lake ice sharply contrasts with the relatively warm climatic and vegetational characteristics of this intermontane region. Summer temperatures (June, July, and August) average 5.5°C in nearby Eureka (60 km to the north). Mean annual temperature averages −20°C. The area is an extreme polar desert where annual precipitation, which falls mainly as snow, averages 68 mm, making it the driest place in Canada (CitationEnvironment Canada, 2003). The catchment is largely unvegetated: the silt and clay-rich lowlands support an enriched prostrate shrub flora (CitationEdlund and Alt, 1989), whereas the felsenmeer highlands, predominantly composed of the highly erodable carbonate bedrock of the Eureka Sound Group (CitationMiall, 1991), support only a sparse saxifrage-dominated community.
The lake is situated well above Holocene marine limit (<140 m a.s.l.: CitationBell, 1996) and is slightly alkaline (pH 8) and dilute with concentrations of major cations ranging from ∼0–4.5 mg L–1 for Na3+, K+, Mg+, and Si4+, with higher concentrations of Ca2+ (∼20 mg L–1) derived from the surrounding bedrock. Lake water temperatures range between 0 and 2°C, and oxygen levels range between over 15 mg L–1 at the surface to 4 mg L–1 at depth in the distal basin (see ).
The mixing regimen of Sawtooth Lake is complicated. The predominance of ice on the lake prevents classic cold monomictic overturn. However, the inflowing streams from the eastern end of the watershed deliver considerable sediment to the lake in the form of overflows, interflows, and underflows (CitationPatridge, 1999) and effectively mix the lake. A sill (60-m depth) in the middle of the lake acts as a barrier to communication between the two basins and creates an isolated trough in the distal basin. This trough is depleted in oxygen, has relatively higher concentrations of hypolimnetic cations, and allows for the preservation of laminae from which a high-resolution record may be obtained.
Methods
CORING AND DIATOM PREPARATION
A suite of gravity and vibracores was collected from the distal basin of Sawtooth Lake in the summer of 1999. Diatoms were sampled from the 36-cm-long gravity core (10–5) at 1–2-mm intervals to approximate an annual record in the uppermost 6 cm and at 1-cm intervals thereafter to 36 cm. The vibracore (10–6B) was sampled at 10-cm intervals to 4 m; thereafter it was sampled at 5-cm intervals to the base at 4.6 m.
Diatom samples from the short core were freeze-dried and then digested in 30% hydrogen peroxide following standard oxidative techniques (CitationBattarbee, 1986). They were repeatedly rinsed, and then aliquots were evaporated at room temperature on round coverslips in evaporation trays. Coverslips were permanently mounted on plain glass slides using Naphrax® mounting medium (RI = 1.74). At least 300 diatoms were enumerated from the uppermost samples using an Olympus HB microscope equipped with phase optics. Where diatom concentrations declined, at least 5 transects of the slide were counted. For several of these (below 5-cm depth), total diatom counts were less than 5. Diatom species are expressed as relative percentages and as estimated concentrations within the sediment based upon the known weight of the original sample, the volume of the subsequent aliquots taken, and the area of the slide surface counted (CitationBattarbee, 1973). Taxonomic assignations were made using reference literature (CitationKrammer and Lange-Bertalot, 1986–1991).
CHRONOLOGY
The chronology of Sawtooth Lake is complicated. An additional surface lamina recovered in each successive year of coring suggests that the sediments are annually laminated (varved). However, a large erosive turbidite at 2-cm depth and several sections where laminae are disturbed preclude a definitive varve chronology for the uppermost section. Two approaches to this chronological uncertainty were produced.
First, we used 210Pb and 137Cs as a guide to the chronology. However, due to dilution, the 210Pb curve for Sawtooth Lake does not decay exponentially with depth. Rather, the 210Pb profile tends to oscillate with lamina thickness, suggesting that old Pb has been washed in from the watershed. However, a strong 137Cs peak stratigraphically correlated with ∼4.0-cm depth in core 10–5 anchors the chronology and is believed to be valid due to the prevalence of clays, which retard the vertical migration of Cs in the sediments (CitationCrusius and Anderson, 1995). In addition, the constant rate of supply (CRS) model (CitationAppleby and Oldfield, 1978) produced a date of 1964, which closely matches that of the 137Cs-inferred date of 1963. These factors suggest that the Pb-based chronology provides a reliable estimate of age within the uppermost 6 cm.
Second, we counted and measured varve thickness as outlined in CitationFrancus et al. (2002). Given that samples for radiometric analysis spanned 0.5 cm of sediment, and the time interval represented by each sample must range over a decade in places, one would not expect exact correspondence between the varve count and the radiometric peaks. Nevertheless, the correspondence that we found is quite consistent with the interpretation of the varve chronology (compare CitationFrancus et al., 2002). These varve counts place the base of the core at least 2550 varve yr BP. In spite of these uncertainties, the timing of the major changes in diatom assemblages, as we will discuss further, can be constrained within a decade. Diatom samples were taken from individual laminae in an effort to approximate an annual record and were assigned ages based on linear interpolation between 210Pb-generated dates.
STATISTICAL METHODS
Diatom species >1% abundance in any one sample were employed in statistical analyses. Gradient lengths were assessed using detrended correspondence analysis (DCA) on CANOCO software (Citationter Braak, 1990). Principle Components Analysis (PCA) was chosen for further analysis due to the linear response of the data, indicated by the short gradient length (1.896) of the DCA 1 axis. The PCA was run using a centered covariance data matrix and then plotted as both a species scatter () and as downcore first-axis scores ().
BIOVOLUME
Diatom biovolume (μm3 g–1 dry sediment) was estimated for diatom species using the BIOVOL program (CitationKirschtel, 1996). Size ranges were estimated from reference photographs from Sawtooth Lake and other girdle views from other high arctic lakes (usually smaller than temperate-cell size ranges). In cases where girdle views were unavailable, volumes were estimated from similarly sized species. Due to the inherent uncertainty in estimating cell volumes, the values are used to provide a rough estimate of changes in diatom production over time rather than absolute numbers. Despite these uncertainties, volumes calculated here were comparable to other, published diatom biovolumes (compare CitationJoynt and Wolfe, 2001).
Results
GENERAL DIATOM STRATIGRAPHY
Diatoms are absent from the entire retrieved sedimentary record of Sawtooth Lake below 5.3-cm depth (∼1926). However, in the uppermost 5.3 cm, diatom concentrations increase rapidly and oscillate between 0 and 3.5 × 107 valves g–1 dry sediment. Of a total of 66 species seen in the lake sediments, only 21 diatom species were recorded in >2% relative abundance (). The siliceous remains of chrysophytes are equally variable and appear periodically throughout the core, but are most abundant at the top, where they range from 0 to 1 × 107 cysts g–1 dry sediment () and below 4-m depth, where they were inconsistently observed.
A trend in the diatom stratigraphy is apparent over the last 80 yr (). In general, diatoms increase in both size and diversity toward the surface: from the early dominance of small Fragilaria species to larger planktonic Cyclotella species and larger benthic taxa (Navicula, Cymbella, etc.) in recent decades. This general trend is superimposed upon the higher-frequency oscillations in species dominance, which punctuate the record.
When diatoms first appear in the lake ca. 1926, it is as a low concentration and low biovolume population of Fragilaria pinnata. Fragilaria species diversify quickly and are replaced by larger taxa, both benthic and planktonic, which reach a maximum circa 1980. In the early 1990s, concomitant with a large turbidite, diatom species diversity and concentration within the sediments plummet. The mid- to late 1990s are marked by a return to high species diversity and concentration.
ORDINATION
The PCA of diatom species highlights trends within the data (). Negative values on the first axis (λ = 64%) represent “warmer”-condition taxa: large, high-biomass, centric diatoms (Cyclotella bodanica and Stephanodiscus alpinus) and larger benthic, epiphytic species belonging to the genera Achnanthes, Cocconeis, Cymbella, Denticula Diploneis, Navicula, Pinnularia, and Nitzschia. The positive, “colder” side of the axis is dominated by the presence of small Fragilaria pinnata (and the occasional Campylodiscus noricus), which is typically associated with late glacial lakes at more temperate latitudes (CitationHaworth, 1976; CitationStabell, 1985; CitationMarciniak, 1986). The second axis (λ = 17%) shows intermediate species: other Fragilaria and small benthic taxa (e.g., Amphora pediculus, Caloneis spp.). These groupings can therefore be seen as a gradient from “cold” to “warm. “The downcore diatoms appear to follow this “ice-cover” gradient with the transition from cold, ice-covered conditions to warmer conditions in the last few decades (see ). Indeed, this trend echoes the climate trends of the region where 6 of the warmest years on record were in the last decade (CitationEnvironment Canada, 2003).
BIOMASS
Two main features are apparent in the diatom stratigraphy of Sawtooth Lake. Years with low diatom concentrations are dominated by the small diatom Fragilaria pinnata. Those years with high diatom concentrations have higher diatom diversity and a larger relative abundance of large diatom taxa. These features are evident in the stratigraphic changes in diatom biovolume (see the right column on ). Cyclotella bodanica is roughly 10–15 times the volume of small Fragilaria pinnata and thus requires considerably more resources for proliferation. This change in biomass, from a few small diatoms to a lot of larger diatoms, illustrates the degree to which the diatom production in Sawtooth Lake has exponentially increased within the last 100 yr.
Discussion
Due to the lack of pitting or etching on the valve surfaces, repetitions in laboratory preparations of diatoms, and the oscillations in diatom concentration at the top of the core, the sedimentary sequence is interpreted as recording an environmental rather than taphonomic signal. First-order changes in both the relative abundance and concentration of diatoms suggest that major changes have occurred within the lake environment over the last 100 yr.
Furthermore, the record of annual median grain size at Sawtooth Lake, interpreted as the early-summer snowmelt intensity (CitationFrancus et al., 2002) shows a clear coarsening in the 20th century, from ∼1900 until the 1960s. Unlike the record for diatoms, the coarsening recorded is not unprecedented in the last 400 yr. Nevertheless, both of these independent proxies, i.e., grain size and diatoms, from the same site clearly demonstrate a response to warming during the latter half of the 20th century.
LAKE ONTOGENY
The change in diatoms reflects the ontogeny of Sawtooth Lake and the evolution of diatom production within it. The small Fragilaria pinnata is considered a pioneering species, commonly found in late glacial sediments (CitationHaworth, 1976; CitationStabell, 1985; CitationMarciniak, 1986), whose adaptation to marginal environments allows it to outcompete larger species in marginal environments (CitationLotter et al., 1999; CitationLotter and Bigler, 2000). The autecology of other Fragilaria species is less well known. However, the diatom record from nearby Solstice Lake (CitationWolfe, 2000) suggests that F. pseudoconstruens may be viewed as an intermediate species on a gradient between “glacial” conditions represented by F. pinnata and interglacial conditions marked by larger diatom taxa in high arctic environments.
Indeed, the changes seen over the last 80 yr in Sawtooth Lake are similar in character to changes seen at lower latitudes in the transition from glacial to interglacial conditions and may provide an analogue for the lacustrine response to rapid warming at the beginning of the Holocene.
REGIONAL DIATOM/LACUSTRINE CHANGE
The paleoclimatic records from fossil diatoms across the Canadian High Arctic reveal strikingly similar changes in recent decades (). In general, all of these records depict a distinct transition from small Fragilaria species, which are indicative of low-productivity environments, to larger benthic, periphytic species. While the nature of these transitions is the same, the timing of the shift varies. Several records show the changes occurring in the late 1800s (CitationDouglas et al., 1994; CitationWolfe, 2000), whereas others show it happening sometime within the last 50 yr (CitationGajewski et al., 1997; CitationDoubleday et al., 1995).
Three diatom records from Cape Herschel on Ellesmere Island (CitationDouglas et al., 1994) show a distinct transition from small Fragilaria species to larger epiphytic species (Pinnularia, Cymbella, Navicula) in the uppermost centimeters of the sediment cores. These records indicate that changes that have occurred in the last ∼150 yr (dated by 210Pb) are unprecedented within the context of the last 6000–8000 yr (14C dating). Similar records are found elsewhere in the Arctic. The record from Solstice Lake (CitationWolfe, 2000), a small lake on the Fosheim Peninsula, Ellesmere Island, shows the same timing of the change from small Fragilaria species to larger benthic taxa as the Cape Herschel records. The recent changes in diatoms in these two records show that the magnitude of recent warming since the Industrial Age is unprecedented in the context of the middle to late Holocene.
The record from DV09, a small lake with laminated sediments on northern Devon Island (CitationGajewski et al., 1997), shows that these changes happened in the last 60 yr. Staurosirella (= Fragilaria pinnata group) and other small taxa gave way to the dominance of larger raphid taxa in recent years. In addition, the total diatom concentration has increased dramatically since 1948. These changes are paralleled by an increase in summer melting on the Devon Ice Cap, as revealed by ice-core studies (CitationFisher and Koerner, 1994).
The record from Lower Dumbell Lake (CitationDoubleday et al., 1995), near Alert military base on the north coast of Ellesmere Island, shows the same type of record and timing of change. Diatoms are absent below 1.5 cm in the core. In the uppermost 1 cm, the fossil assemblage shifts from small Fragilaria pinnata to larger epiphytic and planktonic taxa (Cyclotella spp.). Due to the thick ice cover in this northern locale, the change is interpreted to represent a reduction in ice cover over the last ∼50 yr, with the chronology based on the coincident presence of black carbon spherules (combustion products from nearby Alert, which was established in 1950).
These records all suggest that diatoms in high arctic lakes and ponds, regardless of lake size, latitude, or altitude (), have most likely responded to a decline in ice cover and a concomitant increase in light availability, growing season length, epiphytic habitats, and increased nutrient supply, all of which have likely occurred as a result of recent (last ∼100 yr) warming. Similar records from across the circumpolar region depict the same types of ecological change (CitationSorvari et al., 2002; CitationKorhola et al., 2002). This inferred warming appears to be unique within the context of the late Holocene. Unfortunately, the resolution of these records and the resulting discrepancy in the timing of the shifts preclude any detailed, decadal, regional synthesis. The temporal inconsistency probably relates to differences in sampling and dating resolution, levels of bioturbation, and the size of the lakes, which affect their threshold of sensitivity to climatic perturbations.
OTHER PROXIES
Other high arctic records from different proxies echo the findings of these paleolimnological records (reviewed in CitationOverpeck et al., 1997; CitationSerreze et al., 2000). Varve thickness records from both high (CitationSmith, 1997; CitationLamoureux and Bradley, 1996) and low arctic (CitationHughen et al., 2000) sites depict similar warming starting in the early to mid-1900s. Similarly, the melt record from the Agassiz Ice Cap (CitationKoerner, 1997) and negative glacier mass balances from circumarctic glaciers (CitationDyurgerov and Meier, 2000; CitationDowdeswell et al., 1997) suggest that the last 50 yr have been warm within the context of the last 3000 yr. Tree rings from lower latitudes in the Arctic (CitationBriffa et al., 1995) also show that the last 50 yr were warmer than average. Likewise, global CO2 records from the last 50 yr show that the length of the arctic growing season has increased (CitationOechel et al., 1993). In all, these findings suggest that diatoms are sensitive indicators of recent changes in the arctic environment and that the changes in arctic lakes since the 1920s are regional, not local, in nature.
In terms of Holocene climate, the diatom record from Sawtooth Lake highlights the magnitude of recent changes in the arctic environment. Because the record only extends back to ∼2500 yr BP and does not include a record of the warmer early Holocene (CitationBradley, 1990), it shows the apparent shift from the cold conditions of the late Holocene to greenhouse-gas-induced climatic warming of the last 75 yr. As noted previously, recent changes reflect a rapid transition—within a century—from one of the coldest periods of the late Holocene (the “Little Ice Age”) to one of the warmest (CitationBradley, 2000).
Conclusions
The diatoms from Sawtooth Lake echo findings from across the High Arctic that suggest recent, regional climatic warming has taken place. Shifts in the diatom species suggest that ice cover has decreased over the last 75 yr. This change indicates that environmental conditions of the last ∼75 yr are unlike any of the preceding ∼2550 yr. In addition, it provides an analogue for the types of changes seen in temperate lacustrine environments following deglaciation and should, therefore, provide insight into the larger changes that will take place in high latitudes with increased warming.
FIGURE 1. Location of Sawtooth Lake and other sites referred to in text. Bathymetric map and location of the sediment cores
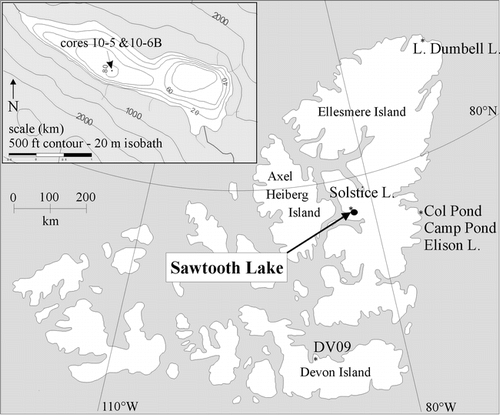
FIGURE 2. Concentration of (a) diatoms and (b) chrysophyte cysts within core 10–5. Diatoms are absent below 6 cm. The gray band marks the presence of a turbidite
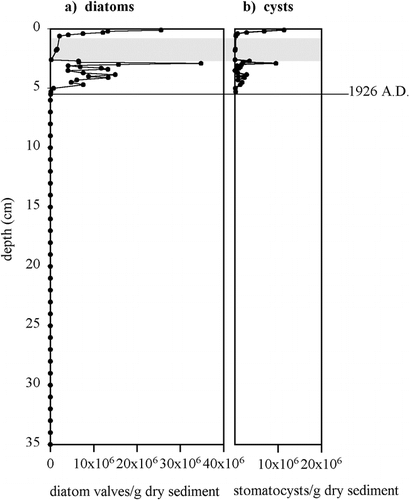
FIGURE 3. Percentage diagram for Sawtooth Lake diatoms and their concentration within the sediments. Sum of other benthic taxa includes Achnanthes, Denticula, Diploneis, Hannaea, Navicula, and Nitzschia as well as other less well-represented genera such as Caloneis, Cocconeis, Cymbella, Diatoma, and Pinnularia. (See for taxa list.) The gray band marks the presence of a turbidite
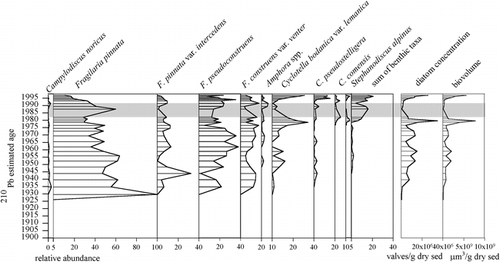
FIGURE 4. PCA biplot of species indicating “cold” and “warm” taxa
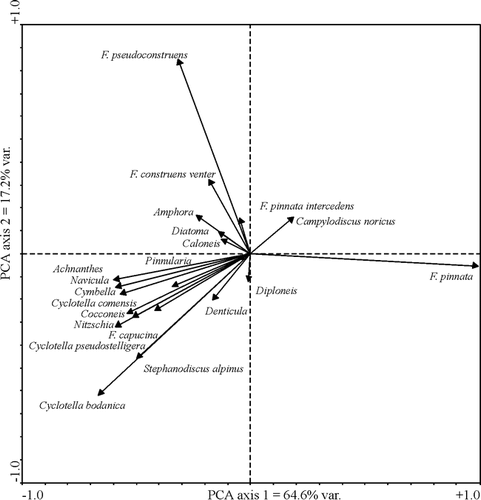
FIGURE 5. PCA axis 1 scores showing inferred cold-warm transition in recent decades
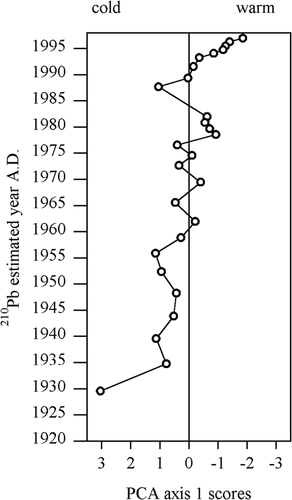
Table 1 Major cation water chemistry
Table 2 Diatom species >2% abundance in any sample
Table 3 Location, altitude, depth, and length of lakes with records referred to in text and on the location map
Acknowledgments
This research was supported by NSF (Earth System History program) grant #9708071 to the University of Massachusetts. Logistical support was provided by Polar Continental Shelf Project. We thank the field crews of 1998, 1999, and 2000 and Frank Keimig for technical assistance. We also thank A. P. Wolfe and E. Saulnier-Talbot, whose comments greatly improved the manuscript. This is PARCS contribution no. 196.
Related Research Data
References Cited
- Appleby, P. G. and G. Oldfield . 1978. The calculation of 210Pb dates assuming constant rate of supply of unsupported 210Pb to the sediment. Catena 5:1–8.
- Battarbee, R. W. 1973. A new method for estimating the absolute microfossil numbers with special reference to diatoms. Limnology and Oceanography 18:647–653.
- Battarbee, R. W. 1986. Diatom analysis. In B. E. Berglund (ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. Chichester: Wiley, 527–570.
- Battarbee, R. W. 2000. Palaeolimnological approaches to climate change, with special regard to the biological record. Quaternary Science Reviews 19:107–124.
- Bell, T. 1996. The last glaciation and sea level history of Fosheim Peninsula, Ellesmere Island, Canadian High Arctic. Canadian Journal of Earth Sciences 33:1075–1086.
- Bradley, R. S. 1990. Holocene paleoclimatology of the Queen Elizabeth Islands, Canadian High Arctic. Quaternary Science Reviews 9:365–384.
- Bradley, R. S. 2000. Past global changes and their significance for the future. Quaternary Science Reviews 19:391–402.
- Briffa, K. R. , P. D. Jones , F. H. Schweingruber , S. G. Shiyatov , and E. R. Cook . 1995. Unusual twentieth-century summer warmth in a 1000-year temperature record from Siberia. Nature 376:156–159.
- Crusius, J. and R. F. Anderson . 1995. Evaluating the mobility of 137Cs, 239+240Pu, and 210Pb from their distributions in laminated lake sediments. Journal of Paleolimnology 13:143–155.
- Doran, P. T. , C. P. McKay , W. P. Adams , M. C. English , R. A. Wharton , and M. A. Meyer . 1996. Climate forcing and thermal feedback of residual lake-ice covers in the High Arctic. Limnology and Oceanography 41:839–848.
- Doubleday, N. C. , M. S V. Douglas , and J. P. Smol . 1995. Paleoenvironmental studies of black carbon deposition in the High Arctic: a case study from northern Ellesmere Island. The Science of the Total Environment 160: /161. 661–668.
- Douglas, M. S V. and J. P. Smol . 1999. Freshwater diatoms as indicators of environmental change in the High Arctic. In Stoermer, E. F. and Smol, J. P. (eds.), The Diatoms: Applications for the Environmental Earth Sciences. Cambridge: Cambridge University Press. 469 pp.
- Douglas, M. S V. , J. P. Smol , and W. Blake Jr. . 1994. Marked post–18th century environmental change in high-arctic ecosystems. Science 266:416–419.
- Dowdeswell, J. A. , J. O. Hagen , H. Bjornsson , A. F. Glazovsky , W. D. Harrison , P. Holmlund , J. Jania , R. M. Koerner , B. Lefauconnier , C. S L. Ommanney , and R. H. Thomas . 1997. The mass balance of circum-arctic glaciers and recent climate change. Quaternary Research 48:1–14.
- Dyurgerov, M. B. and M. F. Meier . 2000. Twentieth century climate change: evidence from small glaciers. Proceedings of the National Academy of Sciences 97:1406–1411.
- Edlund, S. A. and B. T. Alt . 1989. Regional congruence of vegetation and summer climate patterns in the Queen Elizabeth Islands, Northwest Territories, Canada. Arctic 42:3–23.
- Environment Canada, 2003. Canadian Climate Normals 1971–2000. http://www.msc-smc.ec.gc.ca/climate/climate_normals/index_e.cfm .
- Fisher, D. A. and R. M. Koerner . 1994. Signal to noise in four ice-core records from the Agasssiz Ice Cap, Ellesmere Island, Canada: details of the last millenium for stable isotopes, melt and solid conductivity. The Holocene 4:113–120.
- Francus, P. , R. S. Bradley , M. B. Abbott , W. Patridge , and F. Keimig . 2002. Paleoclimate studies of minerogenic sediments using annually resolved textural parameters. Geophysical Research Letters, 29, 20, 1998, doi:10. 1029/2002GL015082.
- Gajewski, K. , P. B. Hamilton , and R. McNeely . 1997. A high resolution proxy-climate record from an arctic lake with annually-laminated sediments on Devon Island, Nunavut, Canada. Journal of Paleolimnology 17:215–225.
- Haworth, E. Y. 1976. Two late-glacial (Late Devensian) diatom assemblage profiles from northern Scotland. New Phytologist 77:227–256.
- Hughen, K. A. , J. T. Overpeck , and R. F. Anderson . 2000. Recent warming in a 500-year paleotemperature record from varved sediments, Upper Soper Lake, Baffin Island, Canada. The Holocene 10:9–19.
- Joynt III, E. H. and A. P. Wolfe . 2001. Paleoenvironmental inference models from sediment diatom assemblages in Baffin Island lakes (Nunavut, Canada) and reconstruction of summer water temperature. Canadian Journal of Fisheries and Aquatic Sciences 58:1222–1243.
- Kelley, D. E. 1997. Convection in ice-covered lakes: effects on algal suspension,. Journal of Plankton Research 19:1859–1880.
- Kirschtel, D. B. 1996. BIOVOL Ver. 2.1. http://www.msu.edu/~kirschte/biovol.html .
- Koerner, R. M. 1997. Some comments on climatic records from ice cores drilled in areas of high melt. Journal of Glaciology 43:90–97.
- Korhola, A. , S. Sorvari , M. Rautio , P. G. Appleby , J. A. Dearing , Y. Hu , N. Rose , A. Lami , and N. G. Cameron . 2002. A multi-proxy analysis of climate impacts on the recent development of subarctic Lake Saanajarvi in Finnish Lapland. Journal of Paleolimnology 28:1 59–77.
- Krammer, K. and H. Lange-Bertalot . 1986. Bacillariophyceae. 1. Teil: Naviculaceae. In Ettl, H., Gerloff, J., Heynig, H., and Mollenhauer, D. (eds.), Süsswasserflora von Mitteleuropa, Band 2/1. Stuttgart and New York: Gustav Fischer Verlag. 876 pp.
- Krammer, K. and H: Lange-Bertalot . 1988. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Ettl, H., Gerloff, J., Heynig, H., and Mollenhauer, D. (eds.), Süsswasserflora von Mitteleuropa, Band 2/2. Jena: VEB Gustav Fischer Verlag. 596 pp.
- Krammer, K. and H. Lange-Bertalot . 1991a. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Ettl, H., Gerloff, J., Heynig, H., and Mollenhauer, D. (eds.), Süsswasserflora von Mitteleuropa, Band 2/3. Stuttgart and Jena: Gustav Fischer Verlag. 576 pp.
- Krammer, K. and H. Lange-Bertalot . 1991b. Bacillariophyceae. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema, Gesamtliteraturverzeichnis Teil 1–4. In Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., and Mollenhauer, D. (eds.), Süsswasserflora von Mitteleuropa, Band 2/4. Stuttgart and Jena: Gustav Fischer Verlag. 437 pp.
- Lamoureux, S. F. and R. S. Bradley . 1996. A late Holocene varved sediment record of environmental change from northern Ellesmere Island, Canada. Journal of Paleolimnology 16:239–255.
- Lim, D. S S. , M. S V. Douglas , and J. P. Smol . 2001. Diatoms and their relationship to environmental variables from lakes and ponds on Bathurst Island, Nunavut, Canadian High Arctic. Hydrobiologia 450:1–3 215–230.
- Lotter, A. F. and C. Bigler . 2000. Do diatoms in the Swiss Alps reflect the length of ice cover?. Aquatic Sciences 62:125–141.
- Lotter, A. F. , R. Pienitz , and R. Schmidt . 1999. Diatoms as indicators of environmental change near arctic and alpine treeline. In Stoermer, E. F., and Smol, J. P. (eds.), The Diatoms: Applications for the Environmental Earth Sciences. Cambridge: Cambridge University Press. 469 pp.
- Mann, M. E. , R. S. Bradley , and M. K. Hughes . 1999. Northern Hemisphere temperatures during the past millennium: inferences, uncertainties, and limitations. Geophysical Research Letters 26:759–762.
- Marciniak, B. 1986. Late glacial Fragilaria flora from lake sediments of the Tatra Mtns and the Alps. In Round, F. E. (ed.), Proceedings of the Ninth International Diatom Symposium, Bristol, England, 233–244.
- Miall, A. D. 1991. Late Cretaceous and Tertiary basin development and sedimentation, Arctic Islands. In Trettin, H. P. (ed.), Geology of the Innution Orogen and Arctic Platform of Canada and Greenland. Geology of Canada, No. 3. Ottawa: Geological Survey of Canada, 437–458.
- Oechel, W. C. , S. J. Hastings , G. Vourlitis , M. Jenkins , G. Riechers , and N. Grulke . 1993. Recent change of arctic tundra ecosystems from a net carbon dioxide sink to a source. Nature 361:520–523.
- Overpeck, J. , K. A. Hughen , D. Hardy , R. Bradley , R. Case , M. Douglas , B. Finney , K. Gajewski , G. Jacoby , A. Jennings , S. Lamoureux , A. Lasca , G. MacDonald , J. Moore , M. Retelle , S. Smith , A. Wolfe , and G. Zielinski . 1997. Arctic environmental change of the last four centuries. Science 278:1251–1256.
- Patridge, W. J. 1999. Modern sediment transport and desposition in a high arctic lacustrine environment: relationship between depositional dynamics and climate: South Sawtooth Lake, Ellesmere Island, Nunavut. B.Sc. thesis, Bates College, Lewiston, ME.
- Pienitz, R. , J. P. Smol , and H. J B. Birks . 1995. Assessment of freshwater diatoms as quantitative indicators of past climatic change in the Yukon and Northwest Territories, Canada. Journal of Paleolimnology 13:1 21–49.
- Round, F. E. 1981. The Ecology of the Algae. Cambridge: Cambridge University Press. 653 pp.
- Rouse, W. R. , M. S V. Douglas , R. E. Hecky , A. E. Hershey , G. W. Kling , L. Lesack , P. Marsh , M. McDonald , B. J. Nicholson , N. T. Roulet , and J. P. Smol . 1997. Effects of climate change on the freshwaters of arctic and subarctic North America. Hydrological Processes 11:873–902.
- Schindler, D. W. , S. E. Bayley , B. R. Parker , K. G. Beaty , D. R. Cruikshank , E. J. Fee , E. U. Schindler , and M. P. Stainton . 1996. The effects of climatic warming on the properties of boreal lakes and streams at the Experimental Lakes Area, Northwestern Ontario. Limnology and Oceanography 41:1004–1017.
- Serreze, M. , J. Walsh , F. Chapin III , T. Osterkamp , M. Dyurgerov , V. Romanovsky , W. Oechel , J. Morison , T. Zhang , and R. Barry . 2000. Observational evidence of recent change in the northern high-latitude environment. Climatic Change 46:150–207.
- Smith, S. V. 1997. A record of environmental change derived from varved lakes along the margin of the Agassiz Ice Cap. Master's thesis, Department of Geosciences, University of Massachusetts, Amherst, MA.
- Smol, J. P. 1983. Paleophycology of a high arctic lake near Cape Herschel, Ellesmere Island. Canadian Journal of Botany 61:2195–2204.
- Sorvari, S. , A. Korhola , and R. Thompson . 2002. Lake diatom response to recent arctic warming in Finnish Lapland. Global Change Biology 8:171–181.
- Stabell, B. 1985. The development and succession of taxa within the diatom genus Fragilaria Lyngbye as a response to basin isolation from the sea. Boreas 14:273–286.
- ter Braak, C. J F. 1990. CANOCO, version 3.10. Wageningen: Agricultural Mathematics Group.
- Weckström, J. and A. Korhola . 2001. Patterns in the distribution, composition and diversity of diatom assemblages in relation to ecoclimatic factors in Arctic Lapland. Journal of Biogeography 28:31–45.
- Wolfe, A. P. 2000. A 6500-year diatom record from southwestern Fosheim Peninsula, Ellesmere Island, Nunavut. In Garneau, M., and Alt, B., (eds.), Environmental Response to Climate Change in the Canadian High Arctic. Geological Survey of Canada Bulletin 529:249–256.
- Wolfe, A. P. and B. B. Perren . 2001. Chrysophyte microfossils record marked responses to recent environmental changes in high- and mid-arctic lakes. Canadian Journal of Botany 79:747–752.
- Zwiers, F. W. 2002. Climate change—the 20-year forecast. Nature 416:690–691.