Abstract
Our study examined carbon dioxide exchange and nitrogen cycling over two consecutive years (winter and summer) in a grazed alpine grassland and in an embedded long-term grazing exclosure to ascertain whether grazing history had resulted in divergent soil carbon attributes, CO2 exchange rates, and different vegetation C and N and soil N processes. Soil C and N concentrations and masses were significantly higher in the grazed than in the ungrazed area, though grass leaf N was higher in the ungrazed area, as was vegetation biomass. Detectable amounts of CO2 were lost from the grazed and ungrazed areas of this grassland during the winters of 1998, 1999, and 2000, and at 6 of 15 winter flux sample dates, CO2 efflux was greater in the grazed area than in the ungrazed area. The ungrazed area consistently gained more C during the summer months than the grazed area, with net CO2 exchange peaking in mid-July 1998 at nearly 5 μmol m−2 s−1 in the ungrazed area compared to <2 μmol m−2 s−1 in the grazed area. During the 2-yr study period, the grazed area was a carbon source of 170 g C m−2, while the ungrazed area was a carbon sink of 83 g C m−2. Lower N mineralization rates early and late in the summer (1999) in the grazed site at Libby Flats corresponded to reductions in net CO2 exchange and lower plant N content compared to the ungrazed exclosure. Based on these results, we suggest that: (1) long-term grazing in high-altitude rangelands can alter annual CO2 exchange and N dynamics; (2) temporal synchrony in C and N processes occur during the summer; that is, increased C exchange rates accompany increased N mineralization rates; and (3) integrative (total soil C and N) and instantaneous (CO2 exchange and vegetation N) measures of C and N dynamics may not necessarily lead to the same interpretation regarding C sequestration and N cycling in alpine grasslands.
Introduction
Alpine ecosystems are an important forage resource for livestock and wildlife, especially in the Rocky Mountain region of North America and in the Alps of Western Europe, where they are used for summer pasture (CitationBillings, 1979; CitationThilenius, 1979; CitationBliss, 1985; CitationDiemer, 1992; CitationSchappi and Körner, 1997; CitationPovirk et al., 2000). Globally, grazing systems in the alpine life zone represent 3% of the global land area (5.7 × 106 km2) and at least 1% of the global terrestrial C pool (CitationKörner, 1995). Although a multitude of studies have examined biogeochemical cycling in low-altitude rangeland ecosystems (CitationMilchunas and Lauenroth, 1993; CitationKelly et al., 2002; CitationSmith et al., 2002), very few studies have compared carbon and nitrogen processes between high-altitude alpine grasslands that have been grazed as opposed to adjacent areas that have been either ungrazed or released from grazing. Understanding the role of grazing in alpine grasslands C and N biogeochemistry is important today as land managers are considering means by which they may alter carbon sequestration characteristics with grazing management practices (CitationFollett et al., 2000; CitationPovirk et al., 2000, CitationConant and Paustian, 2002). Recently it has been suggested that mountain regions in the central United States are important carbon sinks in global carbon budgets (CitationSchimel et al., 2002). While coniferous forests may be accounting for a large portion of these carbon budgets, alpine grasslands and tundra are spatially an important part of high-elevation systems in the Rocky Mountain region, and their carbon budgets may be vulnerable to changing climates (CitationWalker et al., 1994, 1999; CitationWelker et al., 2001).
Grazing by large herbivores can alter ecosystem processes directly and indirectly through biomass and nutrient redistribution (CitationFrank et al., 2002). Biomass removal by grazers reduces the amount of standing litter affecting soil temperatures. Grazing may alter the magnitude of carbon cycling by shifting plant biomass allocation from roots to shoots or vice versa (CitationHolland and Detling, 1990; CitationHolland et al., 1992; CitationFrank et al. 2002) and by delaying senescence. Urine and fecal deposition by herbivores can also alter soil nitrogen processes, thus affecting net soil N mineralization, plant N content, and photosynthetic processes (CitationHolland and Detling, 1990; CitationFrank and Groffman, 1998; CitationLal et al., 1998; CitationFrank et al., 2000; CitationLeCain et al., 2000). When herbivory results in greater N mineralization and higher leaf N contents (CitationHolland and Detling, 1990; CitationSeagle et al., 1992), then the short-term carbon exchange rates and carbon sequestering ability of grazed areas may become greater than ungrazed or lightly grazed areas due to stimulation of photosynthesis (Painter and Detling, 1981). However, generalizations such as these are not always certain or possible because of different herbivory intensities, intra- and interseasonal variations in temperature and precipitation, differing experimental designs, and statistical interpretations (CitationWilson and Jefferies, 1996; CitationTracy and Frank, 1998), along with intrinsic differences in rangeland ecosystems.
In this study, we quantified the CO2 exchange (net ecosystem flux, ecosystem respiration, gross photosynthesis) and the soil N mineralization patterns as well as total biomass and C and N of soils and plants in an alpine grassland where grazing has been maintained and where it has been eliminated 45 yr ago with an exclosure (i.e., ungrazed area). Our measurements were conducted over two consecutive years, including winter, because non-growing-season C and N dynamics were shown to be important in alpine and winter-dominated ecosystems and C and N biogeochemistry (CitationSommerfeld et al., 1993, Citation1996; CitationBrooks et al., 1996; Brooks et al., 1997; CitationFahnestock et al., 1998, Citation1999; CitationBilbrough et al., 2000).
Materials and Methods
SITE DESCRIPTION AND HISTORY
Our study was conducted at Libby Flats (41°20′N, 106°17′W), located on the east side of the Medicine Bow Range in southeastern Wyoming (U.S.A.) approximately 64 km west of Laramie. The research site is an open alpine grassland located at 3230 m with a 3° slope to the north. The area became subject to intense unregulated grazing (e.g., herds numbering 200,000 in the Medicine Bow National Forest, with 60,000 in the Libby Flats–Medicine Bow Peak vicinity) with the introduction of sheep and cattle in the late 1890s, supporting the local mining and timber industries. Sheep grazing since the mid-1900s has been much more moderate, with 1 to 3 herded bands of ∼2000 sheep rotating within the area each grazing season (July to September). The sheep allotment was not leased after 1995; thus, no livestock were present during the study period. A grazing exclosure (60 m × 25 m) was constructed in the center of the grassland by the U.S. Forest Service 45 yr prior to our study to serve as a visual reference site for the surrounding land that continued to be grazed. We used the exclosure and the grazed area surrounding it for this study. The exclosure is located in the center of a large grassland area (>3 ha), and no apparent microtopographic variance is evident between the exclosure and the grazed area within a 50-m circumference of the exclosure. Because this study was a comparison of 1 ungrazed, or released from grazing, area compared to 1 grazed area (pseudoreplication), we attempted to overcome this deficiency with repeated sampling of replicate plots within each area over a 2-yr period and conducted soil measurements at a second, subsidiary high-elevation grassland study site: Hay Creek. The Hay Creek study site (3000 m a.s.l.) is located 1.6 km north of the Big Horn National Forest Experimental Pasture, near Sheridan, WY. This exclosure was officially established in 1924 but had been in place and used as an observational plot since 1913 (CitationMay, 1954). The area adjacent to the Hay Creek exclosure was, and still is, grazed from June to September by cattle. This grassland is dominated by Festuca idahoensis, Bromus pumpellianus, Geum triflorum, Myosotis alpestris, Cerastium arvense, Lupinus sericeus, Phlox multiflora, Agoseris glauca, Poa canbii, Achillea millefolium, and Carex spp. (CitationMay, 1954). Our findings are primarily limited in scope to the study site at Libby Flats, but because the flora, parent material, and land-use history are representative of other alpine grasslands in the Rocky Mountain region (CitationBillings, 1979; CitationWalker et al., 1994), our findings may have implications for surrounding alpine grasslands.
Forb and graminoid species dominate the vegetation inside and outside the exclosure at Libby Flats and include Deschampsia cespitosa, Poa alpina, Polygonum bistortoides, Trifolium spp., Artemesia scopulorum, Geum rossii, Achillea millefolium, Cerastium arvense, Phleum alpinum, Solidago multiradiata, and several Carex species. Soils at Libby Flats are fine, loamy, mixed Entic Humic Dystrocryepts (CitationUSDA, 1999).
Precipitation during the summer in southeast Wyoming is generally in the form of intermittent thunderstorm activity or, more rarely, snowfall. Annual precipitation, recorded at the nearby Glacier Lakes Ecosystem Experiments Site (GLEES), is approximately 120 cm, with summer precipitation accounting for less than 20% (CitationNADP, 1999). Precipitation during our study period is summarized in , with winter snow being almost 50% greater in 1999 than in 1998; summer precipitation was 25% lower in 1999 than in 1998. If we compare years on a “water-year” basis (October–September), precipitation was greater in 1999 than in 1998. Over our 2-yr study period, this area received 302 cm of precipitation, 33% more then the average for the 6 previous 2-yr periods (CitationNADP, 1999).
CARBON DIOXIDE EXCHANGE MEASUREMENTS
Carbon dioxide exchange patterns at Libby Flats were quantified by taking measurements during the growing (snow-free) and nongrowing (i.e., snow-covered) seasons from January 1998 to January 2000. During the nongrowing season when snow was present, we used a CO2 concentration gradient approach with a simple diffusion model to estimate wintertime losses of C from grazed and ungrazed areas through snowpack. This method has been documented elsewhere (CitationSommerfeld et al., 1993, Citation1996; CitationBrooks et al., 1996; CitationFahnestock et al., 1998, Citation1999), but briefly, it entails measuring CO2 concentrations at the snow-atmosphere and snow-soil interfaces with an infrared gas analyzer (PP Systems EGM-1) and calculating diffusional CO2 loss from the soil to the atmosphere. Values are subsequently corrected for temperature, pressure, and nonlinear diffusional flow (i.e., tortuosity) resulting from snow density differences throughout the snowpack. Measurements were taken during the winters of 1998 (1 January–12 May), 1999 (12 December–3 June), and early 2000 (6 January). Weather permitting, 50 randomly located measurements were taken both inside and outside the exclosure during each sample date. Snow depths were recorded at each measurement location. The snow depth increased throughout the winter of 1998, was generally consistent in the winter of 1999, and was not different between the grazed and ungrazed areas (). Profiles of snow density and temperature were made from snow pits dug at each site during each sample date, where soil temperatures were recorded (). In addition to our CO2 flux estimates, these measurements of snow depth and soil temperature allowed us to evaluate their relationship (). Soil temperature may be one of the controlling factors of soil respiration and CO2 efflux in winter (CitationBrooks et al., 1996; CitationSchimel and Clein, 1996; CitationFahnestock et al., 1998).
Growing-season CO2 exchange measurements were taken with an infrared gas analyzer (Licor, LI-6200) connected to a transparent chamber (CitationVourlitis et al., 1993) that was placed over preselected plots at the time of each measurement. Experimental plots (0.75 m × 0.75 m) were selected in July 1997 within the Libby Flats exclosure (8 plots) and in the adjacent grazed area (7 plots). Square polyethylene bases (75 cm × 75 cm × 20 cm high with a 5-cm horizontal lip) were set 15 cm into the soil to provide a seal for CO2 flux measurements (CitationVourlitis et al., 1993). All grazed plots were located between 10 m and 30 m from the exclosure so that the differences between the grazed and ungrazed areas could not be attributed to differences in slope or aspect.
Carbon dioxide exchange was measured during the late spring, summer, and fall of 1998 (12 June–12 September) and in 1999 (20 June–12 November). Diurnal CO2 measurements were taken ca. every 4 hr on each sampling date, and sampling occurred approximately every 1 to 4 wk during the snow-free season. A 75 cm × 75 cm × 40 cm transparent chamber was sealed to each chamber base at the time of each measurement. Four small fans continuously mixed air in the chamber during measurements. Once CO2 concentrations within the chamber stabilized (typically 30–60 s), net CO2 exchange determinations were made for each plot. After these net CO2 exchange measurement (net ecosystem exchange [NEE]), the chamber was covered with an opaque cloth to prevent photosynthesis, and measurements of ecosystem respiration rates were determined. Whole ecosystem C assimilation (i.e., gross photosynthesis) was subsequently calculated by subtracting the respiration measurements from the net CO2 exchange measurements, similar to techniques we have used for carbon exchange characteristics of other alpine (CitationWelker et al., 1999) and arctic (CitationJones et al., 1998; CitationWelker et al., 2000) systems.
SOIL AND VEGETATION MEASUREMENTS
In July 1997, 10 soil samples were collected in the ungrazed exclosure and grazed area at Libby Flats, and in August 1998 at the Hay Creek site (CitationMay, 1954), from 3 depths (0–7.5 cm, 7.5–15 cm, 15–30 cm) for analyses of total C and N content using a 2-m King Tube. Soils were air dried, processed through 2 sieves (0.168 cm and 0.084 cm), and analyzed for total C and N concentration on a Carlo-Erba CHN analyzer. Soil bulk density was determined to a depth of 5 cm, as herbivore effects are often confined to the upper soil profiles (CitationElliott et al., 1999; CitationSchuman et al., 1999). At the Hay Creek site only soil C and N measurements were conducted.
Net N mineralization rates were measured using an in situ soil core incubation method (CitationDiStefano and Gholz, 1986). Eight sets of mineralization measurements were made from 24 June through November 1999 in the grazed and ungrazed areas. For each measurement period, paired soil cores were installed in close proximity to each other. One soil core was harvested immediately for an initial measurement of inorganic N and gravimetric water content. An ion-exchange resin was placed at the bottom of the second core as a leachate trap, and the core was returned to the soil for an incubation period of 2 to 4 wk. Entire soil cores were returned to the lab and processed within 4 hr of sample collection. Coarse roots and gravel were removed, and gravimetric soil water content was determined on a 10-g subsample. Nitrogen was extracted from a second 10-g subsample with 2M KCl for 1 hr. Filter extracts of both soils and resins were analyzed for NH4 + and NO3 − using a continuous-flow autoanalyzer. Leachate N values from resin bags were added to the N values from the incubated soil cores. The difference in N concentration between the initial (T0) and final (T1) measurement of each period was divided by the number of incubation days to yield daily net N mineralization rates. At each sample date, 3 soil temperature measurements were recorded adjacent to the soil cores using a thermocouple placed 5 cm below the surface.
Vegetation samples for C and N concentration determination and total biomass were collected in mid-July 1999 from 10 randomly placed quadrats (0.25 m2) in the grazed and ungrazed areas. All vegetation in each quadrat was harvested to the soil surface and separated in grass and forb categories. Green leaves were separated from dead leaves and stems, all vegetation was oven-dried at 60°C for 48 hr, and total biomass was measured to the nearest 0.1 g. Green leaf biomass was ground to a fine powder using a Wiley mill and a mortar and pestle, and subsamples were analyzed for total C and N concentration on a Carlo-Erba CHN analyzer.
STATISTICAL ANALYSIS
A two-way, repeated-measures analysis of variance (CitationSAS Institute, 1994) was used to detect significant differences (P < 0.05) in summer CO2 exchange between sample dates and between the ungrazed and grazed treatments, with plots treated as replicates. Separate analyses were performed on the net CO2 exchange rate and the rates of photosynthesis and ecosystem respiration. General Linear Model procedures (CitationSAS Institute, 1994) were used to detect differences (P < 0.05) in rates of wintertime CO2 efflux and total soil and plant C and N concentration between grazed and ungrazed treatments, with the soil data separated by soil depth. When significant interactions occurred between sample date and grazing treatment, individual t-tests were used to test for significant (P < 0.05) grazing treatment effects for CO2 exchange characteristics at individual sample dates. Soil inorganic N pools and net N mineralization rates were each analyzed with a two-factor ANOVA using General Linear Models with grazing treatment and sampling date as the main effects. Box plots and normal probability plots of residuals were used to assess normality and outliers. Where appropriate (i.e., P < 0.05 in the main analysis), means were compared using the Tukey-Kramer test.
Results
WINTER CO2 EXCHANGE
Detectable levels of CO2 were lost from these grasslands to the atmosphere in all three winters on Libby Flats. Carbon dioxide efflux was measurable even when air and soil temperatures were fairly cold (e.g., on 1 January 1998, air and soil temperatures were −7.0 and −2.9°C, respectively) () and snow depth was relatively shallow (ca. 20 cm; ). Snow depth and temperature were found to be linearly related () (r2 = 0.4). Overall, there was a significant (P < 0.01) interaction between sample date and grazing treatment effect on winter CO2 efflux. At several sample dates during the winter measurement periods (1 January, 22 April, and 12 May 1998; 3 March and 3 June 1999; and 6 January 2000), the grazed area lost significantly more carbon (i.e., was a greater C source) than the ungrazed area (). The range of carbon lost during winter varied between −0.2 and −1.5 mol CO2 m−2 s−1 (note that negative numbers signify loss of C to the atmosphere, while positive values indicate net C gain by the ecosystem).
GROWING SEASON CO2 EXCHANGE
The grazed area at Libby Flats generally had lower rates of net ecosystem CO2 exchange (i.e., lower C-sink activity) during the snow-free period (∼early June to mid-October) than the ungrazed area, especially during the first summer of study (). A significant (P < 0.01) interaction between sample date and grazing treatment was found for net CO2 exchange in both the 1998 and 1999 growing seasons. In 1998, net CO2 exchange was significantly higher in the ungrazed than in the grazed area on half of all sample dates, while in 1999 this difference was evident at 25% of the sample dates. Lower growing-season carbon gain in the grazed relative to the ungrazed area was due to a combination of lower photosynthetic rates and higher respiration rates (). These higher respiration rates correspond to higher soil temperatures in the grazed as opposed to the ungrazed area ().
Diurnal patterns of NEE depict the short-term temporal nature of carbon exchange at this site in addition to the effects of grazing history (). Early in the summer, both of these systems are net C sources to the atmosphere, but by mid-July, during all hours of the day, both grazed and ungrazed areas net C sources, being greater in the ungrazed. At sunset, while the grazed area is becoming a C source, the ungrazed area remains a C sink. Maximum rates of daily NEE occur in late July in both the grazed and ungrazed systems, being almost two-fold higher in the ungrazed.
We estimated the carbon budget of these systems by taking the mean daily CO2 flux for each sample date (average of the diurnal or average of efflux measured during the day in winter) and multiplying it by the number of days assigned to represent a period between sampling dates (CitationWelker et al., 1999, Citation2000; CitationBubier et al., 2002). We then converted the mass of CO2 to the mass of CO2-C to estimate the CO2-C exchange in the grazed and ungrazed areas over this 2-yr period (CitationWelker et al., 1999, Citation2000). The grazed area was a carbon source of 170 g C m−2, while the ungrazed area was a carbon sink of 83 g C m−2 during the 2-yr period.
SOIL AND VEGETATION PROPERTIES
Total soil C and N concentrations in the upper soil layer (0–7.5 cm) were 67% and 60% greater in the grazed than in the ungrazed area, respectively (). However, there were no significant differences in soil C and N between grazed and ungrazed areas in the lower soil layers, and no differences between bulk density or soil C:N ratios at any depth (). This pattern of higher C and N in the upper profile was also observed for Hay Creek, though the values were slightly higher than those from Libby Flats (). Total N of live green leaves of aboveground vegetation, on the other hand, was significantly (P < 0.05) higher in the ungrazed than grazed area. Total biomass was significantly (P < 0.04) higher in the ungrazed area (225 g m−2) compared to total biomass in the grazed (177 g−2) ().
Rates of nitrification varied significantly over the 1999 growing season and were highest in spring and lowest in late fall. Grazing history had no detectable effect on nitrification rates (). However, there was a significant (P < 0.05) effect of grazing history on net N mineralization rates at certain dates in 1999 (). Rates of N mineralization were highest early in the growing season for both grazing treatments, with significantly higher rates in the ungrazed than grazed area in late June to early July (). Rates declined rapidly with the onset of summer, and no significant differences between grazing treatments were observed during this period. Net N mineralization rates increased in early fall and were once again higher in the ungrazed than grazed area between August 30 and September 23 (). This pattern switched, however, between late September and early October, with the ungrazed area having lower net N mineralization rates than the grazed area, though the absolute rates were exceedingly low.
There was a significant (P = 0.01) date by grazing treatment interaction for extractable soil NH4 +, but extractable soil NO3 − levels were only affected by sample date (P = 0.02) (). Soil NH4 + levels were significantly (P < 0.05) higher in the ungrazed grassland in early and mid-July when concentrations were at their greatest. Soil nitrate levels did not differ between grazing treatments and were also at their greatest concentrations in late July. There was a significant date by grazing treatment interaction (P = 0.01) on gravimetric soil moisture levels (), with higher soil moisture content in the ungrazed than the grazed area in early summer. Soil temperatures, measured at 5 cm depth, were similar in grazed and ungrazed areas ().
Discussion
The C and N characteristics of this grazed and ungrazed area were different in the patterns and magnitudes of carbon exchange, carbon sequestration budgets, and in the carbon and nitrogen concentrations of soils and vegetation. Based on our initial discovery that the soil carbon and nitrogen contents were significantly higher in the grazed area (1997), our studies began with the expectation that the grazed area would have higher rates of net CO2 exchange that would be accompanied by higher vegetation N contents and higher soil N availability (N mineralization). These postulates were, however, not manifested in our winter and summer carbon exchange measurements, our estimates of CO2-C budgets, or our measurements of soil and plant N contents. For instance, the grazed area was a C source over this sampling period while the ungrazed area was a C sink, with the grazed area having lower biomass. These differences correspond to the summer temperature measurements whereby soils were warmer in the grazed area, which would account for the higher rates of ecosystem respiration, lower C-sink strength, and overall CO2-C source activity of the grazed area during our study period.
Our measurements of carbon exchange depicted three important findings: (1) Carbon loss in winter was detectable at all measurement periods (winter soil temperatures were always above −5°C, a theoretical threshold of microbial activity (CitationSchimel and Clein, 1996; CitationBrooks et al., 1996) and there were measurement periods during the snow-covered period, when the grazed area had significantly greater rates of carbon loss than the ungrazed exclosure, contributing to their annual C-source attributes (). This is the first documentation that shows that the processes that occur in summer (i.e., livestock and wildlife grazing) carry over to affect winter C cycling. Because the snow cover in the two systems was uniform, the greater rates of winter carbon loss (carbon source) are likely the result of the higher soil C contents in the grazed area (), as opposed to being the result of warmer winter soil temperatures (CitationSchimel et al., 2004). (2) There were also periods during the snow-free period (summer) when the rates of net CO2 flux were lower in the grazed area compared to the ungrazed system, which is the result of generally lower rates of ecosystem photosynthesis in the grazed (primarily 1998) as opposed to the ungrazed area, in conjunction with higher rates of ecosystem respiration in the grazed area (1998 and 1999). The lower rates of gross photosynthesis correspond to the lower plant N content, which typically equates to lower rates of leaf photosynthesis (Field and Mooney, 1986). Collectively, these fluxes equate to a biannual carbon budget in the grazed area of −170 g C m−2 (CO2-C source) (an average of ∼−85 g C m−2 yr−1), while the ungrazed area was a CO2-C sink of 83 g C m−2 over the biannual period (an average of ∼41 g C m−2 yr−1). These estimates of annual CO2-C flux are very similar in magnitude to the mass of C acquired by alpine systems in GLEES (40.9 g C m−2 in 1991, 47.9 g C m−2 in 1992) (CitationSommerfeld et al., 1993), though higher than the values reported for a suite of alpine ecosystems on Niwot Ridge, Colorado, in 1993 and 1994 (0.3–16.9 g C m−2 yr−1) (CitationBrooks et al., 1996). The higher elevation of Niwot Ridge (>3000 m a.s.l.) and its associated shorter growing season could account for the lower magnitudes of annual carbon exchange. In addition, the magnitudes of these fluxes fit within the rates assigned to mountainous regions of the Rocky Mountains (e.g., ∼80 g C m−2 yr−1, CitationSchimel et al., 2002). (3) Our carbon exchange and sequestration findings indicate that integrative measures of ecosystem carbon budgets (soil C contents) may not necessarily correspond to shorter-term, instantaneous measures of CO2 exchange. Clearly the accumulation of soil carbon is indicative of prolonged periods where carbon gains exceed carbon losses, especially in grasslands where secondary stem growth is absent (CitationFollett et al., 2000). However, the time periods by which soil carbon properties are manifested often exceed the measurement periods of short-term studies, such as this and others (Frank et al., 1995; CitationLeCain et al., 2000). We propose that the nature of ecosystem carbon sequestration is dynamic, and while the trajectory for this grazed grassland is positive (accumulating soil carbon), periods of net carbon source activity are intermixed with periods when these grazed grasslands are net carbon sinks (). CitationOechel et al. (1993, Citation1995) report that the North Slope of Alaska (a cold Carex-dominated grassland) has oscillated between being a C sink for extended periods, intermixed with periods of being a net carbon source of CO2 to the atmosphere and being in carbon balance (Oechel et al., 2000).
The interpretation of the carbon dynamics of our ungrazed area (grazing removal exclosure) requires qualification. We have relied on the assumption that the exclosure was placed in a representative area that was not distinct from the surrounding grassland. This assumes, then, that at the time of construction, the soil carbon and nitrogen contents were uniform across the entire study area. If this assumption is true (likely), then we are left with deciphering how the elimination of grazing by livestock and wildlife (no observational or fecal evidence of wildlife grazing) in the exclosure could lead to lower soil carbon and nitrogen pools in the upper soil profile. Soil carbon losses could be the result of greater rates of decomposition and associated greater rates of soil and ecosystem carbon loss; CO2, or methane (Hobbie et al., 2000). If soil temperatures were warmer in the exclosure, then it is possible that greater carbon losses could account for these differences. However, our measurements of soil temperatures indicate that, on the contrary, soils are warmer in the grazed area where standing dead leaf litter is eliminated by herbivores ().
Another possible explanation for the higher soil C and N in the grazed as opposed to the ungrazed area may be the interactions that occur between herbivory (grazing) intensity and carbon allocation to above- and belowground tissue by grasses (CitationWelker et al., 1985; CitationHolland et al., 1992; CitationWelker and Briske, 1992; CitationHolland et al., 1996; CitationFrank et al., 2002). In some instances, grazed grasslands allocate greater proportions of carbon to roots, as compared to ungrazed grasslands, and have higher root turnover, which in turn contributes to greater amounts of soil carbon and nitrogen (Sims and Singh, 1978; CitationFrank et al., 2002). It is likely that reducing the grazing intensity on this alpine grassland by erecting the exclosure may have lead to a reduction in carbon allocation belowground, which would lead to lower rates of root turnover and lower soil carbon accumulation and sequestration over time ().
However, there appears to be a dissconnent between our short-term measures of ecosystem carbon cycling (i.e., CO2 flux measurements) and the longer-term soil carbon dynamics. This disconnect between short- and longer-term processes is not uncommon, and other studies in low-altitude grasslands with divergent grazing histories also indicate that instantaneous rates of CO2 exchange and soil C contents may not be consistent, i.e., grasslands with the highest soil C contents do not necessarily exhibit the highest rates of instantaneous CO2 exchange (CitationLeCain et al. 2000). The processes controlling the linkages between instantaneous rates of net CO2 exchange and soil carbon contents and soil carbon sequestration require further investigation and represent one of the greatest uncertainties in grassland carbon cycling, grazing management, and its interaction with climate.
Our observation that curtailing grazing in a mountain alpine grassland may change in the short term the sign of CO2 exchange from a CO2-C source to a CO2-C sink has implications for the study of carbon sequestration in the western U.S. mountains (CitationConant and Paustian, 2002; CitationSchimel et al., 2002). In this presentation, CitationSchimel et al. (2002) indicates that changes in land use practices, the recovery of forests from intensive harvesting, and active fire suppression are contributing to the strong C sinks found in the higher elevations of the Rocky Mountain region, approaching 80 g C m−2 yr−1. Our findings suggest that changing grazing practices may also contribute to these “hot spots” of carbon sequestration at high altitudes, as there has been a progressive decline of sheep grazing in high-altitude grasslands, due primarily to reduced consumer demand for red meat, and especially lamb, over the past 40 yr (CitationVavra et al., 1994; CitationLaycock et al., 1996). Considering that the rates of C sequestration of these high-altitude grasslands may approach 40 g C m−2 yr−1, the sign of these systems within the Rocky Mountains may have a large influence on regional carbon processes. While these alpine grassland systems may continue to be used as summer forage by Cervus (elk), the reduction of heavy grazing by sheep may contribute to the strong C sinks observed in the western U.S. mountains. As indicated by others, land use history and carbon cycling are closely linked (CitationFalkowski et al., 2000) and may or may not have just as large an effect on carbon sequestration as changes in climate at high altitudes (CitationSchuman et al., 1999; CitationWelker et al., 1999, CitationConant and Paustian, 2002).
Livestock grazing history at our site appears to have had a long-term positive effect on soil N and C content, especially in the upper soil horizons, similar to that reported by CitationSchuman et al. (1999). This is in contrast to the findings of CitationFrank and Groffman (1998), who found that long-term grazing by native ungulates in Yellowstone National Park, another high-altitude Wyoming grassland, had no effect on any grassland soil variable except gravimetric soil moisture. However, a later study reports that root biomass is higher in grazed than in ungrazed grasslands (CitationFrank et al., 2002), which should in turn equate to higher soil C and N. The higher soil N and C contents we observed at Libby Flats were not, however, mirrored in plant N content, nor were mineralization and nitrification rates consistently higher in grazed sites, as might have been expected. In fact, several times during the growing season, plants in grazed sites had reduced N availability relative to plants in ungrazed sites. This also is in contrast to the findings of CitationFrank and Groffman (1998), who found that net N mineralization in grazed areas was double that of ungrazed areas.
The difference between our observations (grazing having a positive effect on soil C and N contents) and those of CitationFrank and Groffman (1998) may be attributed to the dispersed nature of bison in Yellowstone and thus the patchy nature of microsites where urine and defecation may have had a positive effect on soil N, net primary productivity, and soil C. In contrast, Libby Flats had historical grazing intensities that exceeded thousands of animals in a single open grassland surrounded by subalpine forest, so more uniform grazing, defecation, and urination occurred (CitationDuthie, 1916; CitationPovirk et al., 2001). Bands of sheep are typically herded as very large units to minimize predatory attack; thus, uniform impacts on plants and soils are more likely.
While many studies suggest that large herbivores generally have a positive effect on N cycling (e.g., CitationMcNaughton, 1985; Frank and Evans, 1997; CitationFrank et al., 2000), our study, and those of others, provides contrasting examples. In both our study and those in northwest Wyoming (CitationFrank and Groffman, 1998), there were periods during the growing season when rates of N mineralization were indeed lower in grazed as opposed to ungrazed areas. In our study, leaf N was also lower in the grazed area, supporting our observations of reduced N mineralization rates. CitationTracy and Frank (1998) found that net N mineralization was not significantly (P < 0.05) higher in grazed than in ungrazed areas in northwest Wyoming, nor was shoot N significantly greater in grazed areas. Therefore, it appears that grazing effects on higher-altitude grassland nitrogen mineralization rates are not spatially uniform and merit further investigation before thematic generalizations can be adopted.
Lower N mineralization rates early and late in the summer (1999) in the grazed site at Libby Flats corresponded to lower rates of net CO2 exchange and lower plant N content compared to the ungrazed exclosure. However, this apparent relationship between C and N cycling was not observed in the middle of the growing season; while CO2 gain was much lower in grazed than ungrazed sites at this time, there were no differences in N mineralization or nitrification rates. Because mineralization and nitrification rates are so close to zero, linkages between C and N processes are difficult to detect. It does, however, merit reinforcing that there are indeed phases of the growing season (early summer) when lower rates of nitrification and mineralization (N availability indices) are in line with lower magnitudes of net CO2 flux, suggesting that coupling can occur between C and N processes in this alpine grassland.
Taken together, important insights regarding soil and plant N dynamics and the impact of grazing can be drawn from this and related studies (CitationDormaar et al., 1977; CitationDormaar and Willms, 1990; CitationBauer et al., 1987; CitationWilson and Jefferies, 1996; CitationBardgett et al., 1997, Citation1998; CitationFrank and Groffman, 1998; CitationSchuman et al., 1999; CitationStohlgren et al., 1999; CitationLeCain et al., 2000):
Grazing history can affect the magnitudes, patterns, and amounts of C exchanged with the atmosphere in both predicable and unpredictable ways.
Reducing, and in this case eliminating. livestock grazing may result in time periods when the annual or biannual estimated carbon budgets are different from those where grazing is maintained and may cause the carbon budget sign to switch from being a carbon source to being a carbon sink (CitationConant and Paustian, 2002). However, one needs to keep in mind that these short-term estimates of carbon budgets are not necessarily a reflection of the longer-term carbon sequestration trajectories and may not completely account for changes in belowground carbon processes.
Positive effects of grazing on N mineralization are not necessarily manifested consistently throughout the growing season but may be episodic in nature. There are times when the effect of grazing is positive, periods in which grazing may have a negative effect, and times when there is no grazing effect on N mineralization. These are likely controlled, in part, by the allocation of carbon belowground by plants, the use of root exudates by soil microbes, and their effect on soil N mineralization and precipitation and soil water conditions (CitationHolland and Detling, 1990; CitationHolland et al., 1992, Citation1996).
The effects of grazing on N concentrations and fluxes may not be the same for all graminoid-dominated systems; thus, converting ungrazed systems to grazed systems will not necessarily have a beneficial effect on soil N mineralization or plant-available N and may not lead to increases in carbon sequestration or forage production in all systems (CitationMcNaughton, 1985; CitationCoughenour, 1991; CitationMilchunas and Lauenroth, 1993), nor is it certain that converting heavily (over)grazed systems to ungrazed or lightly grazed systems will alter short- and longer-term carbon processes in predictable ways.
There appear to be periods during the summer when the magnitudes of net carbon flux correspond to the indices of plant nitrogen availability and there are periods when there is very low correspondence between C and N processes, indicative of the complexity in biogeochemical cycling in terrestrial ecosystems.
FIGURE 1. (a) Winter snow depth, (b) winter soil temperatures, and (c) the regression of snow depth and soil temperatures (r2 = 0.3) on Libby Flats during the winters of 1998, 1999, and 2000
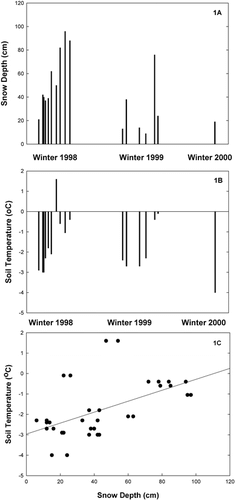
FIGURE 2. Annual net ecosystem CO2 exchange (mean ± 1 SE) in ungrazed and grazed areas at Libby Flats, Wyoming. Asterisks signify significant (P < 0.05) differences between ungrazed and grazed treatments at a sampling date
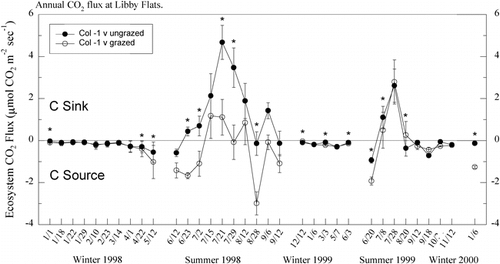
FIGURE 3. Rates of gross photosynthesis and ecosystem respiration in ungrazed and grazed areas at Libby Flats, Wyoming, during the snow-free periods of 1998 and 1999. Asterisks signify significant (P < 0.05) differences between treatments at a sampling date
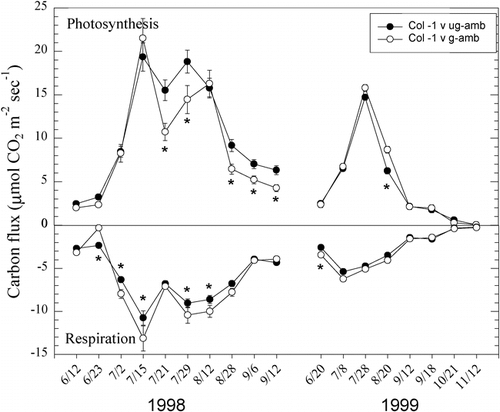
FIGURE 4. Diurnal patterns of net ecosystem exchange (NEE) for 4 sample dates in 1998 at Libby Flats, Wyoming, for areas that were grazed and for areas that were ungrazed
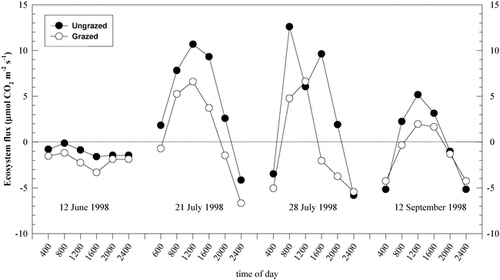
FIGURE 5. Growing-season (1999) patterns of (a) net nitrification and (b) net N mineralization in an ungrazed and a grazed area of Libby Flats, Wyoming. Shown are means ± 1 SE. Asterisks indicate significant (P < 0.05) differences between treatments at a sampling date
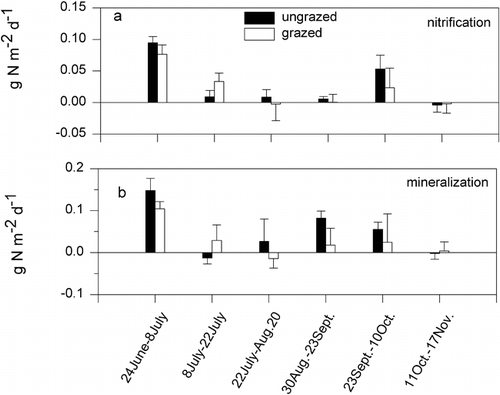
FIGURE 6. Seasonal (1999) patterns of extractable inorganic soil N in grazed and ungrazed areas of Libby Flats, Wyoming. Shown are means ± 1 SE of (a) NH4 +, (b) NO3 −, and (c) total N. Asterisks indicate significant (P < 0.05) differences between treatments at a sampling date
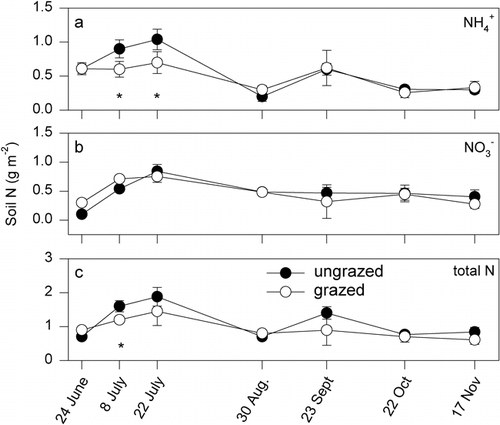
FIGURE 7. Seasonal pattern of summer (a) gravimetric soil water content, measured from 0–10-cm depth, and (b) soil temperature, measured at 5-cm depth in 1999. Shown are means ± 1 SE. Asterisks indicate significant (P < 0.05) differences between treatments at a sampling date
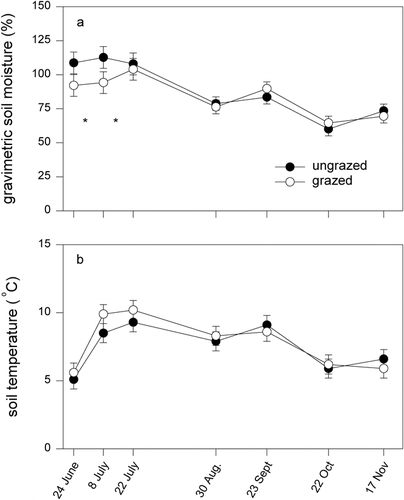
FIGURE 8. Conceptual model depicting the interannual variation in net C exchange as it relates to the long-term carbon-sequestration characteristics of grazed and ungrazed alpine grasslands in southeast Wyoming. While both grazed and ungrazed areas have periods of carbon gain and carbon loss, on average the grazed area accumulates more soil C than the ungrazed area, as indicated by the long-term trajectory
Table 1 Total precipitation for the National Atmospheric Deposition Program site located at Glacier Lakes Ecosystem Experimental site 5 km from the Libby Flats study site (amounts in cm)
Table 2 Soil and vegetation properties of ungrazed and grazed alpine grassland at Libby Flats and at Hay Creek, Wyoming
Acknowledgments
Funding for this research was provided in part by the USDA National Research Initiative Soil and Soil Biology Program (2001789) and the NSF Joint Program for Terrestrial Ecology and Global Change (IBN-97227796).
Related Research Data
References Cited
- Bardgett, R. D. , D. K. Leemans , R. Cook , and P. J. Hobbs . 1997. Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil Biology and Biochemistry 29:1285–1294.
- Bardgett, R. D. , D. A. Wardle , and G. W. Yeates . 1998. Linking aboveground and belowground interactions: how plant responses to foliar herbivory influence soil organisms. Soil Biology and Biochemistry 30:1867–1878.
- Bauer, A. , C. V. Cole , and A. L. Black . 1987. Soil property comparisons in virgin grasslands between grazed and nongrazed management systems. Soil Science Society of America Journal 51:176–182.
- Bilbrough, C. , J. M. Welker , and W. D. Bowman . 2000. Early-spring N uptake by snow covered plants: a comparison of arctic and alpine plant function under snowpack. Arctic, Antarctic and Alpine Research 32:404–411.
- Billings, W. D. 1979. Alpine ecosystems of western North America. In Johnson, D. A. (ed.), Special Management Needs of Alpine Ecosystems. Range Science Series No. 5. Denver, CO: Society for Range Management, 6–21.
- Bliss, L. C. 1985. Alpine. In Chabot, B. F., and Mooney, H. A. (eds.), Physiological Ecology of North American Plant Communities. New York: Chapman and Hall, 41–65.
- Brooks, P. D. , M. W. Williams , and S. K. Schmidt . 1996. Microbial activity under alpine snowpacks, Niwot Ridge, Colorado. Biogeochemistry 32:93–113.
- Brooks, P. D. , S. K. Schmidt , and M. W. Williams . 1997. Winter production of CO2 and N2O from alpine tundra: environmental controls and relationship to inter-system C and N fluxes. Oecologia 110:403–413.
- Bubier, J. , P. Crill , and A. Mosedale . 2002. Net ecosystem CO2 exchange measured by autochambers during the snow-covered season at a temperate peatland. Hydrological Processes 16:3667–3682.
- Conant, R. T. and K. Paustain . 2002. Potential soil carbon sequestration in overgrazed grassland ecosystems. Global Biogeochemical Cyles 16:901–909.
- Coughenour, M. B. 1991. Biomass and nitrogen responses to grazing of upland steppe in Yellowstone's northern winter range. Journal of Applied Ecology 28:71–82.
- Diemer, M. W. 1992. Population dynamics and spatial arrangement of Ranunculus glacialis L., an alpine perennial herb, in permanent plots. Vegetatio 103:159–166.
- DiStefano, D. and R. Gholz . 1986. A proposed use of ion exchange resins to measure nitrogen mineralization and nitrification in intact soil cores. Communications in Soil Science and Plant Analysis 17:989–998.
- Dormaar, J. F. and W. D. Willms . 1990. Effect of grazing and cultivation on some chemical properties of soils in the mixed prairie. Journal of Range Management 43:456–460.
- Dormaar, J. F. , A. Johnston , and S. Smoliak . 1977. Seasonal variation in chemical characteristics of soil organic matter of grazed and ungrazed mixed prairie and fescue grassland. Journal of Range Management 30:195–198.
- Duthie, G. A. 1916. History of the Medicine Bow National Forest. American Heritage Center Historical Records (MBNF Box 5), University of Wyoming, Laramie, WY.
- Elliott, E. T. , J. Heil , E. F. Kelly , and H. C. Monger . 1999. Soil structure and other physical properties. In Robertson, G. P., Coleman, D. C., Bledsoe, C. S., and Sollins, P. (eds.), Standard Soil Methods for Ecological Research. New York: Oxford University Press, 74–85.
- Fahnestock, J. T. , M. H. Jones , P. D. Brooks , D. A. Walker , and J. M. Welker . 1998. Winter and early spring CO2 efflux from tundra communities of northern Alaska. Journal of Geophysical Research 103:29023–29027.
- Fahnestock, J. T. , M. H. Jones , and J. M. Welker . 1999. Winter time CO2 efflux from arctic soils: implications for annual budgets. Global Biogeochemical Cycles 13:775–779.
- Falkowski, P. , R. J. Scholes , E. Boyle , J. Canadell , D. Canfield , J. Elser , N. Gruber , K. Hibbard , P. Högberg , S. Linder , F. T. Mackenzie , B. Moore III , T. Pedersen , Y. Rosenthal , S. Seitzinger , V. Smetacek , and W. Steffen . 2000. The global carbon cycle: a test of our knowledge of Earth as a system. Science 290:291–296.
- Follett, R. F. , J. M. Kimble , and R. Lal . 2000. The potential of U.S. grazing lands to sequester carbon. In Follett, R. F., Kimble, J. M., and Lal, R. (eds.), The Potential of U.S. Grazing Lands to Sequester Carbon and Mitigate the Greenhouse Effect. Chelsea, MI: Ann Arbor Press, 401–430.
- Frank, A. B. , D. L. Tanaka , L. Hofmann , and R. F. Follett . 1995. Soil C and N of Northern Great Plains grasslands as influenced by long-term grazing. Journal of Range Management 48:470–474.
- Frank, D. A. and R. D. Evans . 1997. Effects of native grazers on grassland N cycling in Yellowstone National Park. Ecology 78:2238–2248.
- Frank, D. A. and P. M. Groffman . 1998. Ungulate vs. landscape control of soil C and N processes in grasslands of Yellowstone National Park. Ecology 79:2229–2241.
- Frank, D. A. , P. M. Groffman , R. D. Evans , and B. F. Tracy . 2000. Ungulate stimulation of nitrogen cycling and retention in Yellowstone Park grasslands. Oecologia 123:116–121.
- Frank, D. A. , M. M. Kuns , and D. R. Guido . 2002. Consumer control of grassland plant production. Ecology 83:602–606.
- Holland, E. A. and J. K. Detling . 1990. Plant responses to herbivory and belowground nitrogen cycling. Ecology 71:1040–1049.
- Holland, E. A. , W. J. Parton , J. K. Detling , and D. L. Coppock . 1992. Physiological responses of plant populations to herbivory and their consequences for ecosystem nutrient flow. American Naturalist 140:685–706.
- Holland, J. N. , W. Cheng , and D. A. Crosley, Jr . 1996. Herbivore-induced changes in plant carbon allocation: assessment of below-ground C fluxes using carbon-14. Oecologia 107:87–94.
- Jones, M. H. , J. T. Fahnestock , D. A. Walker , M. D. Walker , and J. M. Welker . 1998. Carbon dioxide fluxes in moist and dry arctic tundra during the snow-free season: responses to increases in summer temperature and winter snow accumulation. Arctic and Alpine Research 30:373–380.
- Kelly, R. D. , E. R. Hunt , W. A. Reiners , W. K. Smith , and J. M. Welker . 2002. Relationships between daytime carbon dioxide uptake and absorbed photosynthetic active radiation for three different mountain/plains ecosystems. Journal of Geophysical Research 107:191–198.
- Körner, Ch 1995. Alpine plant diversity: a global survey and functional interpretations. In Chapin, F. S. and Körner, Ch. (eds.), Arctic and Alpine Biodiversity: Patterns, Causes, and Ecosystem Consequences. Berlin: Springer-Verlag, 45–62.
- Lal, R. , J. Kimble , and R. Follett . 1998. Introduction. In Lal, R., Kimble, J. M., Follett, R. F., and Stewart, B. A. (eds.), Land Use and Soil C Pools in Terrestrial Ecosystems: Management of C Sequestration in Soil. New York: CRC Press, 1–10.
- Laycock, W. A. , D. Loper , F. W. Obermiller , L. Smith , S. R. Swanson , P. J. Urness , and M. Vavra . 1996. Grazing on Public Lands. Task Force Report. No. 29. Council for Agricultural Science and Technology. Ames, IA. 70 pp.
- LeCain, D. R. , J. A. Morgan , G. E. Schuman , J. D. Reeder , and R. H. Hart . 2000. Carbon exchange rates in grazed and ungrazed pastures in Wyoming. Journal of Range Management 53:199–206.
- May, M. 1954. Some effects of mulch on soil temperatures in the Big Horn Mountains. M.S. thesis, Department of Range Management, University of Wyoming, Laramie, WY.
- McNaughton, S. J. 1985. Ecology of a grazing ecosystem: the Serengeti. Ecological Monographs 55:259–295.
- Milchunas, D. G. and W. K. Lauenroth . 1993. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63:327–366.
- NADP. National Atmospheric Deposition Program (NRSP-3) National Trends Network, 1999. NADP Programs Office, Illinois State Water Survey, Champaign, IL.
- Oechel, W. C. , S. J. Hastings , G. L. Vourlitis , M. A. Jenkins , G. Riechers , and N. Grulke . 1993. Recent changes of arctic tundra ecosystems from a carbon sink to a source. Nature 361:520–523.
- Oechel, W. C. , G. L. Vourlitis , S. J. Hastings , and S. A. Bochkarev . 1995. Change in arctic CO2 flux over two decades: effects of climate change at Barrow, Alaska. Ecological Applications 5:846–855.
- Oechel, W. C. , G. L. Vourlitis , S. J. Hastings , R. C. Zuleuta , L. Hinzman , and D. Kane . 2000. Acclimation of ecosystem CO2 exchange in the Alaskan Arctic in response to decadal climate warming. Nature 406:978–981.
- Painter, E. L. and J. K. Detling . 1981. Effects of defoliation on net photosynthesis and regrowth of western wheatgrass. Journal of Range Management 34:68–71.
- Povirk, K. L. , J. M. Welker , and G. F. Vance . 2000. Carbon sequestration in arctic and alpine tundra and mountain meadow ecosystems. In Follett, R. F., Kimble, J. M., and Lal, R. (eds.), The Potential of U.S. Grazing Lands to Sequester Carbon and Mitigate the Greenhouse Effect. Chelsea, MI: Ann Arbor Press, 189–228.
- SAS Institute, 1994. SAS/Stat User's Guide, Version 6, Fourth edition, Vol. 1. Cary, NC: SAS Institute, Inc. 846 pp.
- Schappi, B. and Ch Korner . 1997. In situ effects of elevated CO2 on the carbon and nitrogen cycling status of alpine plants. Functional Ecology 11:290–299.
- Schimel, J. P. and J. S. Clein . 1996. Microbial response to freeze-thaw cycles in tundra and taiga soils. Soil Biology and Biochemistry 36:217–227.
- Schimel, D. , T. Kittel , S. Running , R. Monson , A. Turnipseed , and D. Anderson . 2002. Carbon sequestration studied in western U.S. mountains. EOS, Transactions, American Geophysical Union 83:445. 449.
- Schimel, E. J S. , C. B. Bilbrough , and J. M. Welker . 2004. Increased snow depth effects on microbial activity and N mineralization in two arctic tundra communities. Soil Biology and Biochemistry 36:217–227.
- Schuman, G. E. , J. D. Reeder , J. T. Manley , R. H. Hart , and W. A. Manley . 1999. Impact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecological Applications 9:65–71.
- Seagle, S. W. , S. J. McNaughton , and R. W. Reuss . 1992. Simulated effects of grazing on soil nitrogen and mineralization in contrasting Serengeti grasslands. Ecology 73:1105–1123.
- Sims, P. L. and J. S. Singh . 1978. The structure and function of ten western North American grasslands: net primary productivity, turnover and efficiencies of energy capture and water use. Journal of Ecology 66:573–597.
- Smith, W. K. , R. D. Kelly , J. M. Welker , J. T. Fahnestock , W. A. Reiners , and E. R. Hunt . 2002. Comparison of leaf-to-aircraft measurements of net CO2 exchange in a sagebrush steppe. Journal of Geophysical Research 107:D15. 9:109.
- Sommerfeld, R. A. , A. R. Mosier , and R. C. Musselman . 1993. CO2, CH4 and N2O flux through a Wyoming snowpack and implications for global budgets. Nature 361:140–142.
- Sommerfeld, R. A. , W. J. Massman , and R. C. Musselman . 1996. Diffusional flux of CO2 through snow: spatial and temporal variability among alpine-subalpine sites. Global Biogeochemical Cycles 10:473–482.
- Stohlgren, T. J. , L. D. Schell , and B. V. Vanden Heuvel . 1999. How grazing and soil quality affect native and exotic plant diversity in Rocky Mountain grasslands. Ecological Applications 9:45–64.
- Thilenius, J. F. 1979. Range management in the alpine zone. In Johnson, D. A. (ed.), Special Management Needs of Alpine Ecosystems. Range Science Series, No. 5. Denver, CO: Society for Range Management, 44–64.
- Tracy, B. F. and D. A. Frank . 1998. Herbivore influence on soil microbial biomass and nitrogen mineralization in a northern grassland ecosystem: Yellowstone National Park. Oecologia 114:556–562.
- USDA, 1999. Soil Taxonomy, 2nd ed. U.S. Department of Agriculture. Agriculture Handbook, No. 436. 869 pp.
- Vavra, M. , W. A. Laycock , and R. D. Pieper . 1994. Ecological implications of livestock herbivory in the west. Denver, CO: Society for Range Management. 345 pp.
- Vourlitis, G. L. , W. C. Oechel , S. J. Hastings , and M. A. Jenkins . 1993. A system for measuring in situ CO2 and CH4 flux in unmanaged ecosystems: an arctic example. Functional Ecology 7:369–379.
- Walker, M. D. , P. J. Webber , E. H. Arnold , and D. E. May . 1994. Effects of interannual climate variation on aboveground phytomass in alpine vegetation. Ecology 75:393–408.
- Walker, M. D. , D. A. Walker , J. M. Welker , A. M. Arft , T. Bardsley , P. D. Brooks , J. T. Fahnestock , M. H. Jones , M. Losleben , A. N. Parsons , T. R. Seastedt , and P. L. Turner . 1999. Long-term experimental manipulation of winter snow regime and summer temperature in arctic and alpine tundra. Hydrological Processes 13:2315–2330.
- Welker, J. M. and D. D. Briske . 1992. Clonal biology of the temperate caespitose graminoid Schizachyrium scoparium : a synthesis with reference to climate change. Oikos 56:357–365.
- Welker, J. M. , E. J. Rykiel , D. D. Briske , and J. D. Goeschl . 1985. Carbon import among vegetative tillers within two bunchgrasses: assessment with carbon-11 labeling. Oecologia 67:209–212.
- Welker, J. M. , K. B. Brown , and J. T. Fahnestock . 1999. CO2 flux in arctic and alpine dry tundra: comparative field responses under ambient and experimentally warmed conditions. Arctic, Antarctic, and Alpine Research 31:308–313.
- Welker, J. M. , M. H. Jones , and J. T. Fahnestock . 2000. Annual CO2 flux from dry and moist arctic tundra: field responses to increases in summer temperature and winter snow depth. Climatic Change 44:139–150.
- Welker, J. M. , W. D. Bowman , and T. R. Seastedt . 2001. Environmental change and future directions in alpine research. In Bowman, W. D., and Seastedt, T. R. (eds.), Structure and Function of an Alpine Ecosystem. New York: Oxford University Press, 285–303.
- Wilson, D. J. and R. L. Jefferies . 1996. Nitrogen mineralization, plant growth and goose herbivory in an arctic coastal ecosystem. Journal of Ecology 84:841–851.