Abstract
Kärkevagge, an alpine ecosystem in the subarctic, has been the subject of scientific study for half a century. We investigated the relationship of its vegetation to soil properties. At the lowest elevations, on stable portions of the landscape dominated by Betula and Empetrum, are found Spodosols developed in glacio-fluvial sediments. Cryosaprists occur in scattered bedrock depressions with bog-type vegetation. Cryofluvents are found in flood-prone areas covered by dwarf-willow thickets. Intermediate elevations have meadow-type vegetation with Cryofluvents on floodplains and on lower colluvial slopes, Cryaquents in wetter areas, and Cryorthents on steeper soliflucted slopes. Above that, on steep, west-facing Dryas-covered colluvial slopes, soils are Ca-rich Eutrocryepts and Haplocryolls. The highest-elevation soils are infertile with poor horizonation, despite their possible antiquity, and vegetation that is largely cryptogams. Dystrocryepts occur on more stable alpine locations; Cryorthents on soliflucted areas; and Haploturbels, with shallow permafrost, above 1400 m. Measured annual soil temperatures range from +2.4°C at a Dryas site to −3.4°C at 1585 m at an alpine cryptogam site. Vegetation distribution in Kärkevagge is related to climatic factors, which are controlled by elevation and landscape position, and edaphic factors, which control soil moisture and nutrient availability.
Introduction
One striking feature of arctic-alpine landscapes is the zonation of plant communities (CitationIsard, 1986; CitationBillings, 1987). This is particularly noteworthy in a low arctic-alpine environment such as Kärkevagge, Swedish Lapland, where elevation differences allow a wide range of plant communities to coexist in a small area. The boundaries between plant communities are often abrupt and generally related to elevation. However, other factors associated with vegetation distribution under arctic-alpine conditions may be less obvious (CitationChapin, 1987). In particular, depth and duration of snow cover, which influence moisture and temperature, have been shown to be important variables in cold regions (CitationShaver et al., 1996; CitationStanton et al., 1994), as have soil moisture and chemistry (CitationIsard, 1986; CitationKnapik et al., 1973; CitationRune, 1953).
Kärkevagge is a glacially sculpted valley in the subarctic of Swedish Lapland (). It has a special scientific interest because it is the study area where Anders CitationRapp (1960) quantitatively determined the factors controlling local landscape denudation. Rapp also noted that soil pH, although commonly about 6.0, was as low as 2.9 in certain areas. He found that these differences had a profound effect on vegetation, which he described as primarily distributed by elevation with secondary controls by moisture and snow cover. More recently, the source of the low pH was found to be sulfuric acid produced by oxidation of pyrite found in some local rock units (CitationDarmody et al., 2001, Citation2002). These findings come from a concentrated research effort into weathering and soil development in the valley (CitationSchlyter et al., 1993; CitationThorn et al., 1999, Citation2001, Citation2002a, Citation2002b; CitationDarmody et al., 2000a, Citation2000b; CitationAllen et al., 2001; CitationCampbell et al., 2001, Citation2002; CitationDixon et al., 2002; ). Although there has been some preliminary work describing the vegetation of the valley (CitationKlintenberg, 1995) and one published report including the total elemental composition of plants from a meadow on a colluvial slope (CitationDarmody et al., 2001), there has been no systematic study of the edaphic and climatic factors associated with vegetation distribution in Kärkevagge. The objective of this research, therefore, was to determine the relationships of vegetation distribution in Kärkevagge to soil climatic, chemical, and physical properties.
Study Area
LOCATION AND CLIMATE OF KÄRKEVAGGE
Kärkevagge is a 5-km-long, U-shaped glacial valley trending NNW-SSE in Swedish Lapland (). Its southern terminus is a steep cirque-like headwall, while at the north end the floor of its mouth hangs at about 600 m a.s.l. above the much larger, west-east-trending Vassijuare valley at approximately 18°18′E, 68°26′N. The steep valley walls of Kärkevagge reveal a complex geology of essentially horizontally bedded metamorphic rocks (CitationKulling, 1964). Kärkevagge is in the western Abisko Mountains, a transition zone between the maritime arctic climate along the Norwegian coast, some 80 km to the west, and a more continental climate to the east (CitationEriksson, 1982). In general, regional precipitation decreases and seasonal temperature differences increase to the east. Annual precipitation is evenly distributed and is about 1000 mm (CitationRapp, 1960), although it varies with elevation. Snow, which represents about 50–75% of the precipitation, covers the region 220 to 240 d yr−1. Snow depth reaches a maximum thickness of 0.75 to 1.5 m in March/April (CitationÅngström, 1974). Mean annual air temperature recorded at a midvalley station at 760 m, was ∼1.7 °C September 1995 through August 1996 (CitationThorn et al., 1999). The Kärkevagge region was deglaciated about 9000 to 8500 14C yr B.P. (CitationKarlén, 1979). Through the Holocene the climate has changed from generally warm and moist, to warm and dry, to cold and moist, and the vegetation responded accordingly (CitationKullman, 1998; CitationBarnekow, 2000; CitationBarnekow and Sandgren, 2001).
Vegetation of Kärkevagge
The general distribution of major vegetation belts in the Scandinavian mountains is governed largely by latitude and elevation, which are the primary controls on temperature and precipitation. At the local scale, the occurrence of a specific vegetation community type depends on, or is mediated by, edaphic conditions, depth and duration of snow cover, aspect, exposure, and human influence (CitationGjaerevoll, 1950, Citation1965; CitationSjörs, 1956). The vegetation in the Kärkevagge catchment is representative of the major vegetation community types in the northern Scandinavian mountains, from the treeline to barren rock on the highest parts of the watershed.
The mouth of the Kärkevagge catchment is occupied by subalpine mountain birch forest (Betula pubescens ssp. tortuosa [Ledeb.] Nyman), and the local forest limit is at an altitude of about 530 m a.s.l. Isolated birch trees occur up to 580 m along Kärkejokka, the main stream draining the valley. Treeline has moved upward during recent years, which may in part be an effect of climate amelioration. However, human activity has clearly affected the forest distribution both through previous forms of reindeer husbandry in the area (CitationEmanuelsson, 1987) and from tree cutting during the building of the Kiruna-Narvik railway at the turn of the previous century (CitationSchlyter et al., 1993). The birch forest is luxurious with an undergrowth of meadow-type vegetation that includes Geranium sylvaticum L., Lactuca alpina L., Ranunculus platanifolius L., Angelica archangelica L., and Trollius europaeus L. along with various Salix species, e.g., S. caprea L., S. glauca L., S. lanata L., and S. lapponum L. At slightly higher elevations, and at more exposed and well-drained sites, the birch forest grades into heath with typical species such as Empetrum hermaphroditum Hagerup, Vaccinium vitis-idaea L., Betula nana L., and scattered Juniperus communis L. Also in this zone Vaccinium myrtillus L. is found in more sheltered positions with better snow protection.
The low-alpine vegetation belt is composed of heath, meadow, and willow communities. In Kärkevagge, it ranges from 980 m a.s.l. in shaded areas of the southern parts of the valley to 1100 m on the west-facing slope. The development of the heaths is governed by wind exposure, snow cover, and water availability. Exposed, windy sites with poor snow cover have an incomplete vegetation cover that includes stunted E. hermaphroditum, Diapensia lapponica L., and Saxifraga oppositifolia L. The more continuous vegetation cover on less exposed, dry sites is dominated by E. hermaphroditum, V. vitis-idaea, and V. myrtillus. On more moist sites, B. nana and Salix sp. become more abundant. In wetter areas in the lower parts of the low-alpine belt, Salix dominates, where low- and medium-height species S. lanata, S. lapponum, and S. glauca form thickets.
Alpine meadows are common in the low-alpine belt in areas with extensive snow cover. Large belts of low-herb meadows occur in Kärkevagge above the heaths and in sheltered positions with sufficient snow cover in the heath areas, i.e., along smaller streams and other topographically favored places. Common species are Viola biflora L., Alchemilla alpina L., Polygonum viviparum (L.) Gray, Oxyria digyna (L.) Hill, Ranunculus acris L., Festuca ovina L., Phleum alpinum L., Deschampsia flexuosa (L.) Trin., and Carex bigelowii Torr. Ex Schwein. At well-drained and climatically favorable sites, tall-herb meadow vegetation may develop with typical species such as Angelica archangelica, Geranium sylvaticum, Melandrium rubrum (Weigel) Garcke, Trollius europaeus, and Myosotis decumbens Host. The influence of more base-rich lithologies is evident in the meadow flora on the eastern slopes of the valley as evidenced by the presence of Dryas octopetala L., Salix reticulata L., Saussurea alpina (L.) DC., Parnassia palustris L., and Silene acaulis (L.) Jacq. At better-drained sites this vegetation grades into a Dryas heath community typical of calcareous soils, which shares a large stock of species with base-rich meadow communities.
The meadows at the western side of Kärkevagge are more moist, solifluction is more pronounced, and snow patches more numerous and extensive, as compared with the eastern side, owing to a greater deposition of snow controlled by dominating winter westerly winds. Snow cover exerts a strong influence on vegetation patterns. Where snow cover duration is longer, snow-patch vegetation is present but includes meadow species such as Viola biflora and Oxyria digyna. Typical snow-patch vegetation is very species poor and dominated by Salix herbacea L. and S. polaris Wahlenb., together with species such as Cassiope hypnoides (L.) D. Don, Gnaphalium supinum L., and Veronica alpina L. that can cope with the very short growing period. Extensive areas of snow-patch vegetation occur at lower elevations on the western side of the valley than on the east side. Where snow cover is more extended, snow-patch vegetation becomes very sparse and species poor. Salix polaris and a few moss and lichen species dominate the vegetation of the most long-lived snow patches.
At the highest elevations on thin soils, regolith, or coarser rock rubble substrate, plant cover is minimal with only a limited number of higher plant species represented, e.g., Ranunculus glacialis L. and Luzula arcuata (Wahlenb.) Sw. Cryptogams, i.e., mosses and lichens, are found at the highest and most exposed sites.
Methods and Materials
FIELD METHODOLOGY
Sites were selected to represent most of the common vegetation community types in the valley (CitationKlintenberg, 1995) on typical geomorphic surfaces (). Soil pits were excavated by hand to 1.5 m or until the water table or immovable rock was encountered. Soils were examined and described in the field using standard techniques (CitationSoil Survey Staff, 1993), and the soils were sampled by horizon. In total, 42 soil pits were sampled from 10 vegetation community types, and 227 soil samples were retrieved for later analyses (CitationDarmody et al., 2000a). Two of the vegetation types, referred to as “wind heath” (WH), a stunted Empetrum community, and “midalpine” (MA), an alpine-meadow transitional community, were not replicated. Therefore, they were excluded from the statistical analyses and will only be discussed in passing.
Soil temperature was monitored at 11 selected sites by means of miniature loggers buried at 10 cm that recorded temperatures 4 times daily for 1 yr in 1995–1996 (CitationThorn et al., 1999). Air temperature was recorded at the research hut in the center of the valley at 760 m a.s.l. (CitationThorn et al., 1999). Growing degree days were calculated as the annual sum of the degrees exceeding a reference temperature of 0, 0.5, 5, or 10°C, but no upper limit was applied (CitationMaxwell, 1992). Isothermal days were computed as the sum of days when the soil temperature was 0 ± 0.5°C.
Lab Methodology
The soils were oven-dried in the lab and passed through a 10-mesh sieve to isolate the <2-mm fraction, which was saved for further analysis. Particle-size analysis was by sieving for sands and by hydrometer for silt and clay (CitationGee and Bauder, 1986). A 1:1 soil to distilled water suspension was used to determine pH (CitationMcLean, 1982). Extractable elements were determined by inductively coupled plasma spectroscopy on 1:10 soil : Mehlich 3 extracts (0.2N CH3COOH + 0.25N NH4NO3 + 0.013N HNO3 + 0.015N NH4F + 0.001M EDTA) (CitationMehlich, 1984). Cation exchange capacity (CEC) was estimated by summation of Mehlich 3 extracted cations. Base saturation (BS) was determined using exchangeable H estimated from SMP buffer pH at pH 7.8 (CitationMcLean, 1982). Organic carbon content was determined with a Leco C analyzer; as there were no free carbonates in the soil, total C was attributed to organic matter. Total N was determined with a Kjeldahl digestion followed by detection of ammonia using a Lachat Flow Injection Analyzer. Statistical evaluations were done at the α = 0.05 level using Sigma Stat statistical software (SPSS Inc., Chicago, IL). Statistical tests included ANOVA on means or ranks followed by pairwise multiple comparison procedures by SNK or by Dunn's method, as appropriate.
Results
PHYSICAL PROPERTIES
Our findings are consistent with generalized descriptions of vegetation zonation in the watershed (CitationRapp, 1960; CitationKlintenberg, 1995), where elevation is the primary controlling factor (). Slope gradients ranged up to 78% and were steepest on the colluvial slopes, which were covered with Dryas and meadow vegetation types. Bog-type vegetation was confined to semiclosed bedrock depressions, which had the lowest slope gradients, 1 to 4%. Willow-type vegetation was primarily confined to wetter areas, such as along the floodplains, and consequently had low slope gradients, 1 to 14%. Soil texture was generally coarse, with clay contents nowhere exceeding 7%. Coarse fragment contents were great, typically ranging from 10 to 66% by volume (CitationDarmody et al., 2000a). Consequently, particle size families were coarse-loamy to sandy-skeletal (CitationSoil Survey Staff, 1999). The coarsest-textured soils were found at the wind heath and Empetrum heath sites and at one willow site. Low clay content and high coarse-fragment contents make for poor soil moisture storage and, together with landscape position, dictate soil drainage and plant available water. Bog vegetation was confined to ponded sites. At the other moisture storage extreme, Empetrum heath was largely confined to the excessively drained soils formed from coarse outwash deposits on kame terraces and eskers, or on the tops of the large boulders that litter the valley floor and gives it its name. Vegetation community types in order of decreasing wetness were bog, willow, solifluction meadow, meadow, midalpine meadow, alpine, birch, Dryas, Empetrum, and wind heath.
Soil classification was not universally well correlated with vegetation community types as used in this study (CitationSoil Survey Staff, 1999). This reflects the edaphic effects of small-scale topography on microclimate, snow cover, and hydrology as well as landscape history and dynamics. However, some general associations were identified. Bogs were associated exclusively with Cryosaprists, highest-elevation alpine sites with Haploturbels, Dryas sites with Eutrocryepts and Haplocryolls, and birch and Empetrum sites at the lower elevations with Haplocryods (). As CitationRapp (1960) noted, the best-developed soils in the valley, those showing the most contrasting horizons, were found at the lowest elevations under birch and Empetrum. These vegetation types are the most vigorous in the area, with abundant above-ground biomass, and they produce organic acids that promote soil profile development. The well-developed soils also suggest that these landscape positions are stable, as opposed to most other locations in the valley that show evidence of instability. The unstable landscape positions include floodplains and colluvial and soliflucted slopes. At these locations the soils are Cryorthents or Cryofluvents with buried soils that indicate mass movement, solifluction, erosion, or deposition.
Chemical Properties
Despite Kärkevagge's relatively small geographical area, its soils exhibit a wide range of chemical properties (). Measured pH ranged from a high of 8.2 to a low of 3.5. High-pH soils were associated with high extractable Ca content, which was the most variable parameter measured, ranging from 30–5010 ppm. The Ca is derived from weathering of localized calcareous rocks (CitationCampbell et al., 2001). Likewise, the macronutrient K is generally abundant throughout the area due to the prevalence of micaceous schists; its content ranges from 4–303 ppm. Another potentially important nutrient derived from local rocks is S, which ranged from 15–180 ppm and may originate, in part, through pyrite weathering (CitationDarmody et al., 2001). One floodplain site, W4, produced enough H2S to smell of rotten eggs (CitationDarmody and Fanning, 1977). Phosphorus and N are generally considered the most limiting plant nutrients in the subarctic (CitationJonasson and Michelsen, 1996). In Kärkevagge, extractable P content ranged from 6 to 107 ppm. The source is presumably apatite in the bedrock. The other macronutrient, N, ranged from 0.04 to 2.1% in the surface soil samples. It was closely related (R2 = 0.89, n = 40) to the soil organic carbon content, which ranged from 0.1 to 48% in all the samples, with the highest organic matter contents in the surface A and O horizons (). Plotting the distribution with depth of measured soil parameters shows the importance of bioaccumulation of nutrients in the soil organic matter at the soil surface. The surface O and A horizons were higher in CEC, organic C, S, P, Ca, Mg, K, Mn, Cu, and Zn than the subsurface horizons ().
When surface soil fertility parameters are evaluated over the vegetation groups, some general patterns emerge (). The bog and Empetrum heath soils had statistically the highest amounts of both C and N and the alpine soils the lowest. However, the C/N ratio did not follow the same pattern. The highest C/N ratio, 33, was in the Empetrum heath soils, but a statistically indistinguishable C/N ratio, 31, was found in the soil with the lowest organic C content, the alpine soil. This high ratio indicates that the organic matter is resistant to decomposition in both situations, but production of organic matter is enhanced in the Empetrum soils and inhibited in the alpine soils.
Extractable P and K contents in the surface soils, along with S, did not vary by vegetative group. In contrast, the important element Ca showed strong associations with vegetative groups; it was greatest in the Dryas, meadow, solifluction meadow, and willow sites. The Dryas sites are on the upper slopes below the cliffs where calcareous rocks are exposed. Other high-Ca sites are on stream courses where Ca-rich waters are common (CitationCampbell et al., 2001), or below the Dryas sites on the colluvial slopes. In contrast, the alpine sites had the lowest measured Ca as well as the lowest Mg content. These sites are on the ridge crests, stratigraphically above the calcareous rock and, in addition, may be the most leached due to their antiquity (CitationAllen et al., 2001). Both pH and base saturation tend to follow the Ca content and are statistically related to it (R2 = 0.84 and 0.81, respectively; n = 46 for both). The highest surface horizon pH was with the Dryas soils and the lowest were with the alpine, birch, and Empetrum heath soils. The low pH in the birch and heath soils was reflected in the strong podzolization that these soils showed. As would be expected, the base saturation trends followed soil pH, with the highest base saturation in the Dryas soils and the lowest in the alpine soils. Cation exchange capacity showed similar trends and was strongly correlated (R2 = 0.77, n = 46) with Ca content. Extractable Al followed trends generally opposite to pH and Ca: it was lowest in the Dryas soils and highest in the alpine soils. Most micronutrients did not show an association with vegetation groups, although the meadow sites were highest in Fe and Cu, and B was highest in the Dryas soils.
When grouped by drainage class, the poorly drained soils tended to have higher levels of the parameters measured (). These include pH, base saturation, Ca, Mg, and B. This is probably due to the conveyance of Ca and other nutrients to the poorly drained sites in ground and surface water. Many ephemeral streams that feed the wet locations in the landscape are charged with nutrients, particularly Ca (CitationDarmody et al., 2000b; CitationCampbell et al., 2001). Organic C was not different between the two drainage groups, due to the high amount of organic C accumulated both in the poorly drained bog soils and in the well-drained heath soils. However, the C/N ratio was higher, 28, in the well-drained soils than in the poorly drained soils, 17. This indicates that the organic matter in the well-drained soils is more resistant to decomposition.
Thermal Properties
At the drainage basin scale, topography, which largely controls soil moisture and most importantly soil temperature, is the dominant control over plant distribution in Kärkevagge. The alpine sites, at the highest elevation, had lowest mean annual (MAT) and winter (MWT) soil temperatures (at 10 cm) as well as the lowest minimum temperatures recorded (). The other sites did not vary much in MAT, ranging from 1.2 to 2.4°C, or in MWT, ranging from a low of −1.6°C in one Dryas site to a high of 0.2°C in the solifluction meadow site. Mean summer temperatures (MST) overlapped somewhat, with the alpine sites coolest, ranging from 0.8 to 6.2°C, and the Dryas sites warmest, ranging from 7.7 to 8.0°C. Although permafrost is theoretically present in soils with MAT <0°C, which includes all the alpine sites, that special climatic situation was encountered only at site A5 at an elevation of 1590 m. Permafrost may be found at lower elevations in Swedish Lapland, particularly in organic soils where palsas are found, generally east of Abisko. However, it can be expected within 2 m of the soil surface only at elevations above 1400–1500 m in the area of Kärkevagge, according to CitationÅkerman and Malmström (1986). Where there is evidence of permafrost within 2 m and cryoturbation, as at site A5, the soil classification is Haploturbel (CitationDarmody et al., 2000a).
Snow cover is an important factor influencing soil temperature (CitationWilliams and Gold, 1976). It insulates the soil, keeping it cool in summer and warm in winter. The minimum air temperature we recorded was −26.6°C, but most of the soils never experienced temperatures below −2°C at the 10-cm depth where they were recorded. Snow falls in Kärkevagge before many of the soils freeze, and isothermal conditions existed for more than 250 d at 10 cm deep in the soils at the midalpine meadow, solifluction meadow, and birch sites and at one Dryas site. The daily mean temperature recorded at birch site 1 clearly showed the effect of a deep snow cover. The soil temperature at 10 cm never dropped below 0°C, whereas at alpine site 4, the snow cover was not sufficient to prevent the temperature dropping to −15°C ().
Growing degree days (GDD) are often used to assess the progress of plant growth and maturation (CitationMaxwell, 1992). They are calculated by subtracting from the daily mean air temperature a reference threshold temperature, then summing over days, excluding degrees over a maximum threshold reference temperature. For conventional midlatitude agricultural crops the low reference temperature is 10°C and the upper is 30°C. However, it is unclear what reference temperatures we should use. Arctic and alpine plants are very cold tolerant; alpine plant roots may grow at temperatures as low as 0°C (CitationBillings, 1974). However, at soil temperatures below 5°C, B. pubescens root growth and N uptake are nil, and raising the temperature above 15°C does not increase the rates (CitationKarlsson and Weih, 1996). Only one site, SM6, had temperatures that slightly exceeded 15°C on a total of only 5 d. Given the uncertainties, we calculated GDD at low reference soil temperatures of 0, 0.5, and 5°C, with no upper reference temperature.
GDDs at 0, 0.5, and 5°C are all closely related with correlation coefficients ranging from 0.95 to 0.99, so they essentially are equivalent and tell a similar story. All GDDs were lowest at the alpine and midalpine meadow sites. This is obviously attributable to the high elevation of the alpine sites (, ). However, at the midalpine site, long snow cover and location at the head of the valley, where there is little direct sunshine, accounts for the cold soil temperatures (). Indeed, maximum temperature recorded at MA1 is less than that of site A4, located 340 m above it. The highest GDD0 was found at the Dryas and the Empetrum heath sites. The Dryas sites were at midelevations, generally well drained, and on west-facing slopes, so they had relatively low heat capacity and received maximum insolation. The Empetrum sites were at low elevation and excessively drained, giving them higher air temperatures and low heat capacity. Despite lush vegetative growth, the birch sites had low GDD due not to low minimum temperatures, but to their long period of snow cover and isothermal conditions.
Cryptogam vegetation found at alpine site A4 testifies to a harsh climate despite the fact that it had a GDD similar to the more lushly vegetated midalpine and birch sites. This discrepancy can be explained when the GDD is plotted against minimum temperature, where all alpine sites stand out with low minimums in contrast to the midalpine and birch sites (). Another way to compare the soil climates is to look at the accumulation of GDD through the summer (). The Dryas sites are the first to accumulate 50 GDD units and continue to accumulate GDD units past Julian day 260, keeping pace with the air temperatures. The growing seasons at the alpine sites and the snow-covered and wet sites (B1, MA1, SM6) start late. However, after the snow melts, the birch and solifluction sites continue to accumulate GDD units well into the fall, unlike the alpine sites and site MA1, which cool rapidly in the fall. In terms of their maximum accumulation of GDD0.5 units, the solifluction site SM6 kept increasing until Julian day 336, while the highest-elevation site, A5, stopped at Julian day 252. Other sites with similar late-season accumulation of GDD0.5 units were H1 and B1. With GDD5 units, there is a different pattern. The Dryas sites remain warm longer into the fall than do the lower-elevation sites B1, H1, SM6, or the alpine sites. Site A5 stopped accumulating GDD5 units on Julian day 239 and GDD0.5 units on Julian day 252. Thus, the growing season, defined as the number of days between when the first GDD0.5 unit is accumulated and the day when no additional units are added, is as short as 49 d at A5 or as long as 157 d at SM6. Similarly, the extremes for the GDD5 growing year are only 19 d for A5 but 108 d at DH2, again showing the harsh climate at the alpine sites and the warmer climate at the midelevation sites with favorable aspect. It should be noted that length of the growing season may not be as important to plant productivity and species composition as soil fertility and protection from wind (CitationShaver et al., 1996). This is exemplified by the wind heath (, site WH1) of sparse and stunted vegetation with very poorly expressed soil found on the exposed summits of coarse-textured eskers in the lower valley.
Discussion
The gross control of vegetation distribution in Kärkevagge is climatic, but at a smaller spatial scale, snow cover and soil drainage, texture, and chemistry are significant. Both the minimum temperatures experienced and the length of the growing season are important. Latitude controls the general climate, but microclimate is largely controlled by topoclimatic factors, snow cover, soil moisture, exposure, aspect, etc. Snow cover has a great role in modifying the microclimate, both preventing low air temperatures from reaching into the soil and delaying warmup in the spring. Soil moisture is controlled by soil texture as influenced by topography. The wettest sites in Kärkevagge (, bog 1 and 2) had organic soils associated with small bedrock depressions isolated from the main stream channels, which otherwise over time would deposit mineral matter to fill in the depressions and dilute the organic matter content below organic soil levels. In contrast, the driest soil, found at the wind heath site (, site WH1) on an esker in the lower valley, was very coarse textured and poorly developed.
EMPETRUM AND BIRCH SOILS
These vegetation types were at the lowest elevation in the valley. This is also the most stable area owing to low slope gradients and isolation from both depositional and erosional forces that are very active in much of Kärkevagge. Here, parent materials were less influenced by Kärkevagge's lithology and reflect materials transported by the larger glacier that occupied Vassijuare valley. The stability of the landscape, as well as the low pH parent materials, allowed processes driven by organic acids produced by Empetrum and Betula to reduce parent material pH from a high of 5.0 in the C horizon to as low as 3.5 in the surface horizons. Lower soil pH was found only where soils were impacted by sulfuric acid generated by pyrite oxidation (CitationDarmody et al., 2001). In addition, soil horizonation is best expressed under Empetrum and birch vegetation because the organic acids promote movement of Fe, Al, and organic C, i.e., the process of podzolization, forming spodic horizons, and thus Spodosols.
Empetrum is efficient at mobilizing nutrients and accumulating them at the surface. For example, the associated soils had the lowest overall levels of Ca, but the mean ratio of Ca in the topsoils to the subsoils (A/C) was highest, 2.7, with this vegetation type (). Despite the fact that Empetrum heath soils are excessively drained, they have the thickest histic epipedons and most organic C at the surface of the soils in Kärkevagge (). The organic matter produced by Empetrum may not be easily decomposed due to its high C/N ratio and the droughty conditions. Overall, Empetrum soils were infertile and lowest in micronutrients Cu, Zn, and Fe but highest in extractable Al. The low metal contents show the effect of the chelating agents produced by the Empetrum plants that complex metals and expedite their movement in the soil profile. The high Al content may be a significant challenge for most plants growing on these soils. An extractable Ca/Al ratio of <1 may be phytotoxic (CitationMcLean, 1982), and in the Empetrum subsoil the Ca/Al ratio is 0.3.
Soils at the birch sites were chemically and pedologically similar to the Empetrum heath sites but did not have thick fibrous surface O horizons. Like Empetrum soils, birch soils classify as Haplocryods, which show influence of chelating agents on soil chemistry. The difference between the two is soil moisture storage. Birch vegetation in Kärkevagge is predominantly found on finer-textured tills than the heath at the same elevation, which is found on coarse-textured outwash. The finer texture imparts greater soil moisture storage. In addition, birch sites occur in less exposed locations and thus capture more snow. However, the boundary between the two vegetation types has shifted. This change is evidenced by decomposing birch roots found in the Empetrum soils, away from any live birch trees, likely due to deforestation associated with railroad construction (CitationRapp, 1996; CitationSchlyter et al., 1993). This indicates that birch trees could grow on the very coarse-textured soils but have yet to reoccupy those soils. As with the Empetrum soils, birch soils have relatively low CEC, pH, BS, Ca, Mg, Cu, and Zn and show strong horizonation as evidenced by Fe and Al distributions ( and ). Although the Empetrum and birch soils were the only Spodosols in Kärkevagge, there is evidence of weak podzolization occurring in some alpine, Dryas, and meadow soils, where Bw horizons have been recognized ().
DRYAS
Dryas octopetala is well known as a calciphilic species, and it stands out as the dominant species on high-pH, calcium-rich soils in Kärkevagge. For example, extractable Ca content in the Dryas surface soils averaged 25 times greater than that in the alpine surface soils (). However, in contrast to the Empetrum soils, the Dryas soils had less Ca increase in the surface horizon, giving these soils a low A/C horizon Ca ratio of 1.4 (). In addition, the Dryas soils had the highest Ca/Mg ratios of all the soils. Dryas sites were on the upper colluvial slopes, where they received drainage water from the calcareous rocks exposed above them in the cliffs. The high Ca content and productive vegetation allowed these Dryas soils to accumulate a relatively large pool of organic matter. This led to the development of a Mollisol in one Dryas location, assumed to be the most northerly documented in the world (CitationDarmody et al., 2000a). Dryas soils also stood apart in that they had the lowest extractable Al, as expected given the high pH. Their extractable Ca/Al ratio was 54.4 in the surface and 11.6 in the subsoil, the highest of all the sites.
ALPINE
In contrast to the highly fertile Dryas soils, the alpine soils had the lowest levels of soil fertility. Their stratigraphic position places them above local calcareous rocks, and they are developed in residuum from schist, so the bedrock is not a particularly good source of nutrients. The cold, harsh conditions contribute to the poor productivity of these soils, which may be the oldest in Kärkevagge. They are protected from water erosion and deposition, as is commonly found at lower elevations, and are often armored by a pavement of coarse fragments, which, although indicative of frost heaving and past wind erosive forces, affords some protection from current erosion. In addition, there is evidence that regional glaciation involved cold-based ice in some ridge crest locations, which would preclude glacial erosion and deposition (CitationKleman and Hättestrand, 1999; CitationAllen et al., 2001). Thus, the low fertility of these soils can largely be explained by their antiquity, which has caused considerable loss of plant nutrients over the millennia.
Poor plant productivity, together with the cold that both prevents more efficient plants from growing and causes cryoturbation, can be evoked to explain the lack of well-expressed horizonation in the alpine soils. Alpine plants were the worst at accumulating nutrients in their topsoils, having mean A/C horizon ratios of 0.9, 2.9, 1.0, 1.9, and 1.2 for Ca, organic C, Mg, K, and P, respectively (, , and ). Owing to their windswept location, any nutrients that plants have translocated to their aboveground tissues are liable for removal by wind erosion as the tissues die. The CEC, base saturation, organic C, and extractable Ca, Mg, Ca/Mg ratio, and B were lowest in the alpine soils (). Alpine soils also were highest in extractable Al, giving them a mean Ca/Al ratio of 0.25 in the surface and 0.27 in the subsoil, the lowest we measured and within the phytotoxic range (CitationMcLean, 1982).
MEADOW AND SOLIFLUCTION MEADOW
Meadow and solifluction meadow soils were more moist than the Dryas or Empetrum soils. They were chemically and physically similar and both supported luxurious stands of grasses and forbs. Solifluction meadow sites were on the wetter, western side of the valley and contained buried A or O horizons due to the solifluction lobes overriding surfaces downslope. On the eastern slopes, meadow soils also contain buried surfaces due to intermittent deposition of colluvium. Both vegetation types had fertile soils, as evidenced by the relatively high values of CEC, pH, BS, P, S, Ca, and Mg ().
WILLOW AND BOG
Willow sites occur in the wetter portions of the landscape, and their soils show influence of increased organic matter production, reduced decomposition, and groundwater discharge. While their composition varied from place to place, these soils generally were among the highest in CEC, pH, Ca, Mg, and K. The bog soils were in the wettest locations and had the greatest amount of organic C and N throughout the soil profile ().
Summary
Soils in Kärkevagge are quite variable and show a wide range in chemical and physical properties controlled by local soil conditions operating on general elevational trends. Based on our research, the soil-vegetation relationships in Kärkevagge can be simplified into four general groups. First, at the lowest elevations, about 500–700 m a.s.l., are the podzols, Typic Haplocryods associated with birch and Empetrum heath vegetation. These soils are developed on well-drained, coarse-textured, glacio-fluvial deposits of generally low fertility. They have the best soil horizonation of all soils found in the area, and the heath soils have the thickest histic epipedons. They are intermediate in soil temperature, with a growing season, when soil temperatures at 10 cm are above 5°C, of about 75–83 d.
The second group, at about 600–900 m a.s.l., is the more poorly drained meadow, willow, and solifluction meadow soils. These are developed on coarse, loamy-textured alluvium or colluvium of moderately high soil fertility. They commonly have multiple buried thin O or A horizons and exhibit only slight horizonation, if any. The soil classifications include Cryofluvents, Cryorthents, and Cryaquents. Soil temperatures here are cooler, with a growing season of about 57–68 d. Included in this group are small patches of organic soils, Cryosaprists, in bedrock depressions with bog-type vegetation.
The third group is the highly fertile soils that support Dryas vegetation. These are at about 650–900 m a.s.l., and are developed on steeply sloping, well-drained, coarse-loamy colluvium. They have the highest pH, base saturation, and Ca content of any soil in the area and are also the warmest, with a growing season of about 87–108 d. The soils occasionally have buried A or O horizons and, at most, weakly developed profiles. Their classifications include Cryorthents and Eutrocryepts. At one location, the presence of a dark, organic-rich Mollic epipedon supports a soil classification of Haplocryoll.
The fourth group presents the greatest challenge for plant growth, with the most infertile, coldest soils in the area. These are the alpine soils, developed in colluvium and residuum on the mountain summits, at elevations above about 1100 m a.s.l. They are on the oldest land surfaces in the area. They have the lowest levels of pH, base saturation, Ca, N, and most other soil fertility parameters measured, except Al, which is greatest in this group. The Ca/Al ratio in these soils averages 0.3, which may indicate Al phytotoxicity. These soils also have the shortest growing season, about 19–69 d, and the coldest winter temperatures. The soils are classified as Dystrocryepts or Cryorthents, but where permafrost is within 2 m, they are classified as Haploturbels.
At the lower elevations in Kärkevagge, except where plants are sparse due to damage by late-lying snow, rock falls, mass movements, or animals (including people), growth is vigorous and essentially prevents soil erosion. Indeed, CitationRapp (1960) said that slope wash was of minor importance on the grass-covered slopes. However, erosion has occurred in Kärkevagge in the past, as evidenced by buried soils commonly found in Dryas, willow, meadow, and solifluction meadow sites. Recently, radiocarbon dating of buried soils in one meadow site (CitationHolmqvist and Schlyter, 2000) supports Rapp's observation. Five buried soils down to 22 cm radiocarbon date from 130 to 1280 yr B.P. This indicates that despite steep slope (16%) and harsh climate, there has been little or no erosion at this site, only deposition of materials eroded from upslope. The calculated deposition rate was low, 0.08 to 1.40 mm yr−1. The highest deposition rates date from the Little Ice Age, when the climate was presumably more harsh. We know from digging in these soils that the meadow-type vegetation forms a strong sod, but the alpine cryptogam soils do not enjoy that strengthening at the surface. Consequently, there may have been mass movement off the high alpine sites onto the meadow sites below, burying the surface soils (CitationRapp, 1960; CitationAllen et al., 2001). Fortunately, the high fertility of the soils allows the meadow vegetation to quickly reestablish itself and stabilize the slopes after a depositional event. This resilience in resisting erosion will be tested in the future as Kärkevagge responds to increased recreational and grazing pressures.
FIGURE 1. Location of Kärkevagge, Swedish Lapland showing vegetation group soil sample site locations: H, Empetrum heath; B, birch; Bog, bog vegetation; W, willow; DH, Dryas heath; M, Meadow; A, alpine; MA, midalpine; SM, solifluction meadow; WH, wind heath. Numbers indicate replicate sites within a vegetation group
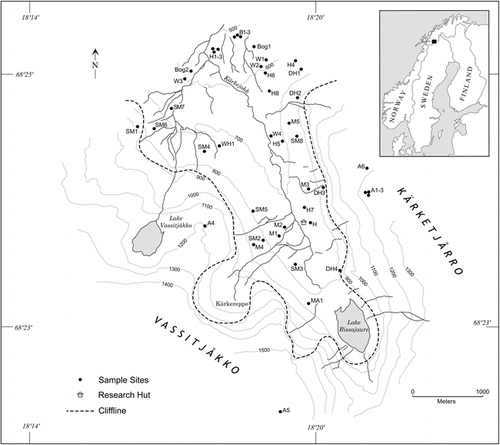
FIGURE 2. Extractable Fe contents by depth in selected solifluction meadow, Empetrum heath, and birch soils. Note the increase in the Bs horizons of the Empetrum and birch soils, indicating podzolization, not seen in the soils from less stable sites such as the solifluction soils
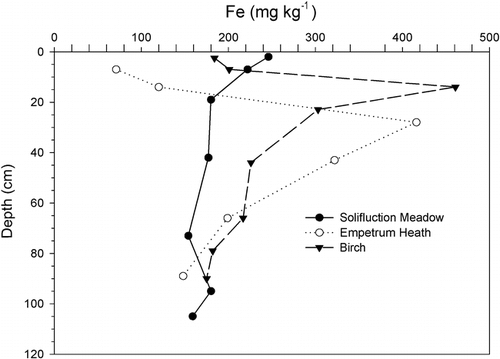
FIGURE 3. Extractable Al contents by depth in selected alpine, birch, Empetrum heath, and Dryas soils. Note the increase in the Bs horizons of the Empetrum and birch soils and in the Bw horizon of the alpine soil, indicating podzolization, not seen in the soils from less stable sites or where Ca dominates the soil chemistry, as in the Dryas soils
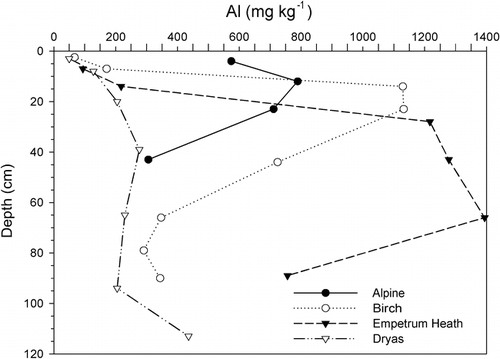
FIGURE 4. Organic carbon (OC) contents by depth in selected alpine, bog, Empetrum heath, and Dryas soils. Note the overall high OC in the bog soils, in the surface of the Empetrum and Dryas soils, and in the Bs horizon of the Empetrum soils and the low OC in the alpine soils
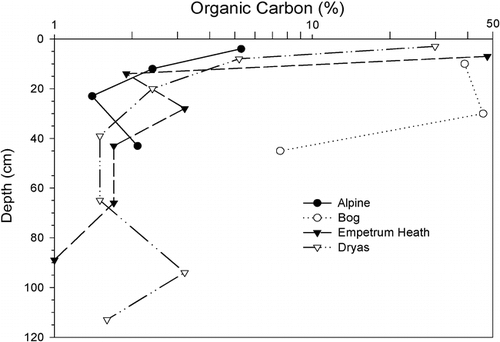
FIGURE 5. Extractable Ca contents by depth in selected alpine, solifluction meadow, Empetrum heath, and Dryas soils. Trends mimic base saturation and pH. Source of Ca in these soils is calcareous rocks exposed in the cliffs of Kärkevagge
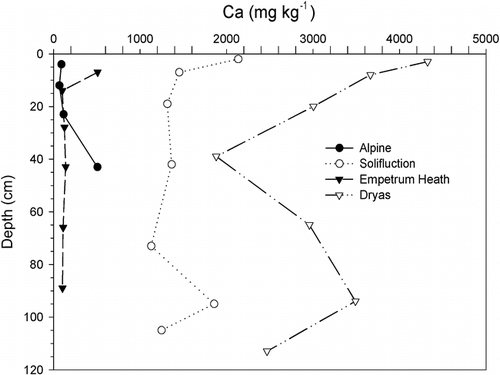
FIGURE 6. Extractable P contents by depth in selected alpine, Empetrum heath, and Dryas soils. Note higher amounts at the surface associate with organic matter except with the alpine soils, where P tends to decrease toward the surface
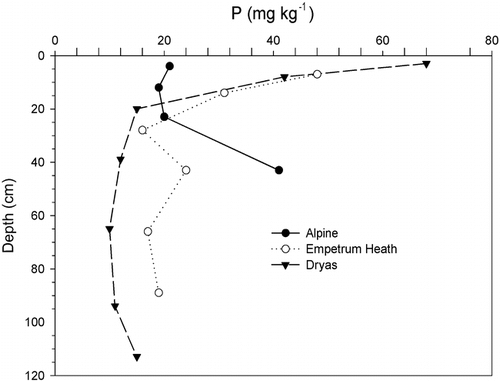
FIGURE 7. Daily mean soil temperatures at 10 cm depth at birch site B1 and alpine site A4. At B1 the logger was in the upper Bs horizon. There was a 2-cm-thick Oh horizon on the mineral soil surface. At A4, the logger was in the upper Bw horizon; there was no organic surface at this site
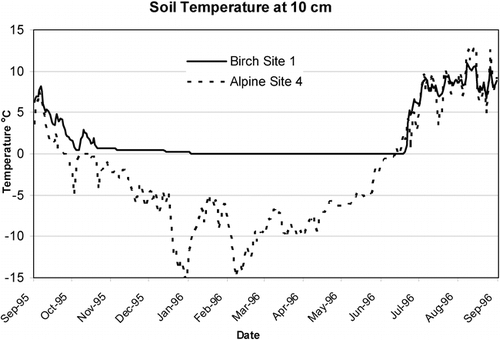
FIGURE 8. Effect of elevation on growing degree days. Legend: A, alpine; B, birch; DH, Dryas; MA, midalpine; H, Empetrum heath; SM, solifluction meadow; air, air temperature at midvalley
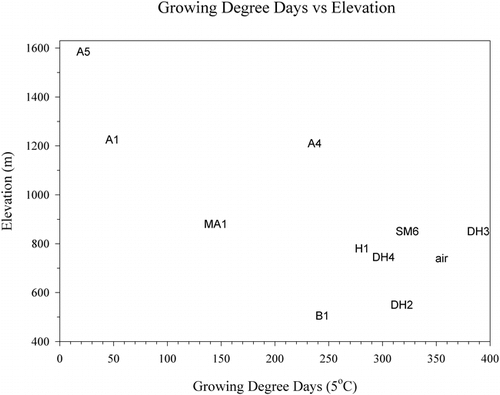
FIGURE 9. Minimum temperature compared to growing degree days. Legend: A, alpine; B, birch; DH, Dryas; MA, midalpine; H, Empetrum heath; SM, solifluction meadow
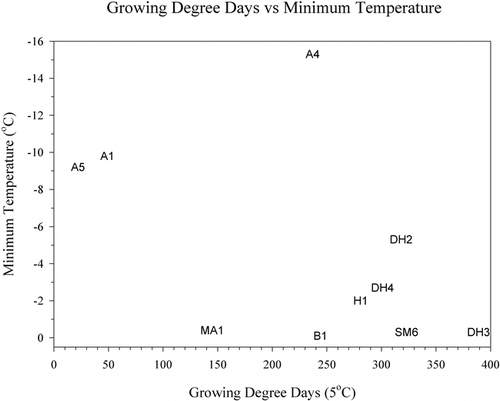
Table 1 Generalized site characteristics of vegetation community/geomorphic sampling sites in Kärkevagge
Table 2 Range of soil fertility parameters found at the vegetation research sites
Table 2 (Cont)
Table 3 Mean values of surface soil parametersa by vegetation community type and drainage
Table 4 Climate data for the vegetation community types
Table 5 Julian date at which cumulative growing degree days (GDD) are achieved at individual sites in Kärkevagge for the year 1996
Acknowledgments
This work was conducted as part of a larger project concerning weathering in Kärkevagge sponsored by a variety of sources. Fieldwork over several years has been funded in part by the Universities of Arkansas, Illinois, and Lund (Sweden) and supported logistically by the Royal Swedish Academy of Sciences Field Station at Abisko. Funding was also provided by grants 5479-95 and 5674-96 from the National Geographic Society and grants BCS-9818917 and BCS-9818667 from the National Science Foundation. We would also like to acknowledge the personal support and encouragement of the late Anders Rapp.
Related Research Data
References Cited
- Allen, C. E. , R. G. Darmody , C. E. Thorn , J. C. Dixon , and P. Schlyter . 2001. Clay mineralogy, chemical weathering, and landscape evolution in arctic-alpine Sweden. Geoderma 99:277–294.
- Åkerman, J. H. and B. Malmström . 1986. Permafrost mounds in the Abisko area, northern Sweden. Geografiska Annaler 68A:155–165.
- Ångström, A. 1974. Sveriges klimat (Swedish climate). Stockholm: Generalstabens Litografiska Ansyalta Forlag. (In Swedish with English summaries.) 156 pp.
- Barnekow, L. 2000. Holocene regional and local vegetation history and lake-level changes in the Torneträsk area, northern Sweden. Journal of Paleolimnology 23:399–420.
- Barnekow, L. and P. Sandgren . 2001. Paleoclimate and tree-line changes during the Holocene based on pollen and plant macrofossil records from six lakes at different altitudes in northern Sweden. Review of Paleobotany and Palynology 117:109–118.
- Billings, W. D. 1974. Adaptations and origins of alpine plants. Arctic and Alpine Research 6:129–142.
- Billings, W. D. 1987. Constraints to plant growth, reproduction, and establishment in arctic environments. Arctic and Alpine Research 19:357–365.
- Campbell, S. W. , J. C. Dixon , R. G. Darmody , and C. E. Thorn . 2001. Spatial variation of early season surface water chemistry in Kärkevagge, Swedish Lapland. Geografiska Annaler 83A:169–178.
- Campbell, S. W. , J. C. Dixon , C. E. Thorn , and R. G. Darmody . 2002. Chemical denudation rates in Kärkevagge, Swedish Lapland. Geografiska Annaler 84A:179–185.
- Chapin III, F. S. 1987. Environmental controls over growth of tundra plants. Ecological Bulletin 38:69–76.
- Darmody, R. G. and D. S. Fanning . 1977. Whiffing sulfur levels of tidal marsh soils. Soil Survey Horizons 18:16–17.
- Darmody, R. G. , C. E. Thorn , J. C. Dixon , and P. Schlyter . 2000a. Soils and landscapes of Kärkevagge, Swedish Lapland. Soil Science Society of America Journal 64:1455–1466.
- Darmody, R. G. , C. E. Thorn , R. L. Harder , J. P L. Schlyter , and J. C. Dixon . 2000b. Weathering implications of water chemistry in an arctic-alpine environment, Northern Sweden. Geomorphology 34:89–100.
- Darmody, R. G. , C. E. Allen , C. E. Thorn , and J. C. Dixon . 2001. The poisonous rocks of Kärkevagge. Geomorphology 41:53–62.
- Darmody, R. G. , S. W. Campbell , J. C. Dixon , and C. E. Thorn . 2002. Enigmatic efflorescence in Kärkevagge, Swedish Lapland: the key to chemical weathering?. Geografiska Annaler 84A:187–192.
- Dixon, J. C. , C. E. Thorn , R. G. Darmody , and S. Campbell . 2002. Weathering rinds and rock coatings from an arctic alpine environment, Northern Scandinavia. Geological Society of America Bulletin 114:226–238.
- Emanuelsson, U. 1987. Human influence on vegetation in the Torneträsk area during the last three centuries. Ecological Bulletins 38:95–111.
- Eriksson, B. 1982. Data rörande Sveriges temperatur-climat (Data concerning the air temperature of Sweden). SMHI Reports. Meteorology and Climatology. RMK 39. (In Swedish with English summaries.).
- Gee, G. W. and J. W. Bauder . 1986. Particle-size analysis. In Klute, A. (ed.), Methods of Soil Analysis, Part 1, 2nd ed. Agronomy Monograph 9. Madison, WI: ASA and SSSA, 383–412.
- Gjaerevoll, O. 1950. The snow-bed vegetation in the surroundings of Lake Torneträsk, Swedish Lapland. Svensk Botanisk Tidskrift 44:387–440.
- Gjaerevoll, O. 1965. Plant cover of the alpine regions. Acta Phytogeographica Suecica 50:257–268.
- Holmqvist, J. and P. Schlyter . 2000. Vesicular-arbuscular mycorrizae for dating multiple paleosols and carbon loss rate determination. Geoderma 97:125–133.
- Isard, S. A. 1986. Factors influencing soil moisture and plant community distribution on Niwot Ridge, Front Range, Colorado, U.S.A. Arctic and Alpine Research 18:83–96.
- Jonasson, S. and A. Michelsen . 1996. Nutrient cycling in subarctic and arctic ecosystems, with special reference to the Abisko and Torneträsk region. Ecological Bulletin 45:45–52.
- Karlén, W. 1979. Deglaciation dates from northern Swedish Lapland. Geografiska Annaler 61A:203–210.
- Karlsson, P. S. and M. Weih . 1996. Relationships between nitrogen economy and performance in the mountain birch Betula pubescens ssp. Tortuosa . Ecological Bulletin 45:71–78.
- Kleman, J. and C. Hättestrand . 1999. Frozen-bed Fennoscandian and Laurentide ice sheets during the last glacial maximum. Nature 402:63–66.
- Klintenberg, P. 1995. The vegetation distribution in the Kärkevagge valley. Lunds Universitets Naturgeografiska Institution. Seminarieuppsatser, No. 32.
- Knapik, L. J. , G. W. Scotter , and W. W. Pettapiece . 1973. Alpine soil and plant community relationships of the Sunshine area, Banff National Park. Arctic and Alpine Research 5:161–170.
- Kullman, L. 1998. Tree-limits and montane forests in the Swedish Scandes: sensitive biomonitors of climate change and variability. Ambio 27:312–321.
- Kulling, O. 1964. Överskit Över Norra Norrbottensfjallens Kaledonberggrund; The geology of the Caledonian rocks of the northern Norbotten Mountains. Sveriges Geologiska Undersökning: Seie Ba Översiktskator Med Beskrivningar, No. 19.
- Maxwell, B. 1992. Arctic climate: potential for change under global warming. In Chapin III, F. S., et al. (eds.), Arctic Ecosystems in a Changing Climate: an Ecophysiological Perspective. San Diego: Academic Press, 11–34.
- McLean, E. O. 1982. Soil pH and lime requirement. In Page, A. L, Miller, R. H., and Keeney, D. R. (eds.), Methods of Soil Analysis, Part 2, 2nd ed. Agronomy Monograph 9. Madison, WI: ASA and SSSA, 199–224.
- Mehlich, A. 1984. Mehlich No. 3 extractant: a modification of Mehlich Mo. 2 extractant. Communications in Soil Science and Plant Analysis 15:1409–1416.
- Rapp, A. 1960. Recent development of mountain slopes in Kärkevagge and surroundings, Northern Scandinavia. Geografiska Annaler 42:65–200.
- Rapp, A. 1996. Photo documentation of landscape change in northern Swedish mountains. Ecological Bulletin 45:170–179.
- Rune, O. 1953. Plant life on serpentines and related rock in the north of Sweden. Acta Phytogeographica Suecica 31:1–139.
- Schlyter, P. , P. Jönsson , R. Nyberg , P. Persson , A. Rapp , C. Jonasson , and J. Rehn . 1993. Geomorphic process studies related to climate change in Kärkevagge, northern Sweden - status of current research. Geografiska Annaler 75A:55–60.
- Shaver, G. R. , J. A. Laundre , A. E. Giblin , and K. J. Nadelhoffer . 1996. Changes in live plant biomass, primary production, and species composition along a riverside toposequence in Arctic Alaska, USA. Arctic and Alpine Research 28:363–379.
- Sjörs, H. 1956. Nordisk växtgeografi. Stockholm: Svenska Bokförlaget Bonniers. 229 pp.
- Soil Survey Staff, 1993. Soil Survey Manual. USDA Handbook 18. Washington, DC: U.S. Government Printing Office. 437 pp.
- Soil Survey Staff, 1999. Soil Taxonomy, 2nd ed. USDA-NRCS Agricultural Handbook 436. Washington, DC: U.S. Government Printing Office. 869 pp.
- Stanton, M. L. , M. Rejmanek , and C. Galen . 1994. Changes in vegetation and soil fertility along a predictable snowmelt gradient in the Mosquito Range, Colorado, U.S.A. Arctic and Alpine Research 26:364–374.
- Thorn, C. E. , J. P L. Schlyter , R. G. Darmody , and J. C. Dixon . 1999. Statistical relationships between daily and monthly air and shallow-ground temperatures in Kärkevagge, Swedish Lapland. Permafrost and Periglacial Processes 10:317–330.
- Thorn, C. E. , R. G. Darmody , J. C. Dixon , and P. Schlyter . 2001. The chemical weathering regime of Kärkevagge, arctic-alpine Sweden. Geomorphology 41:37–52.
- Thorn, C. E. , R. G. Darmody , C. E. Allen , and J. C. Dixon . 2002a. Near-surface ground temperature regime variability in selected microenvironments, Kärkevagge, Swedish Lapland. Geografiska Annaler 84A:289–300.
- Thorn, C. E. , R. G. Darmody , J. C. Dixon , and P. Schlyter . 2002b. Weathering rates of buried machine-polished rock disks, Kärkevagge, Swedish Lapland. Earth Surface Processes and Landforms 27:831–845.
- Williams, G. P. and L. W. Gold . 1976. Ground Temperatures. Canadian Building Digest 180. 4 pp.