Abstract
Carbon dioxide fluxes in a dry subarctic heath were examined after 10 and 11 yr of experimental manipulations of temperature, light, and nutrients. The aim was to investigate how growing season carbon (C) balance was affected by the major climatic factors that are expected to change in the future. Carbon flux was measured in closed chambers as uptake through gross ecosystem production (GP), release through ecosystem respiration (ER), and as net ecosystem production (NEP). Diurnal NEP through a day with clear skies at peak growing season was consistently negative through all treatments the first year of measurement, and day-time NEP varied around zero at eight days across the growing season the second year, implying that a net release of C from the ecosystem to the atmosphere may take place during the growing season. Our results suggest that respiration was the main determinant of C balance, and that variations in light levels and temperature could alter the balance between C uptake and C loss. Fertilization strongly enhanced both ER and GP whereas temperature enhancement changed neither ER nor GP. Shading decreased both ER and GP. After harvest of the aboveground plant biomass, the belowground respiration was 72 to 93% of the ER before harvest. The significant treatment effects on belowground respiration after harvest were similar to the effects on ER before harvest. These results suggest that the ER were mainly from belowground respiration, and that the treatments affected the belowground respiration more than the respiration above ground.
Introduction
It is predicted that the current release of anthropogenic greenhouse gases will increase air temperatures considerably and trigger changes in precipitation patterns in the Arctic within the next century (CitationMaxwell, 1997; CitationRowntree, 1997). This will lead to effects on soil temperatures and soil moisture regimes, and indirectly create changes in soil organic matter (SOM) quality as mineralization processes will be affected. Furthermore, light availability may decrease as a result of enhanced cloud formation due to increased evaporation at warmer temperatures (CitationMaxwell, 1992). These changes can potentially affect the carbon (C) balance of arctic ecosystems by altering the photosynthetic uptake relative to respiratory release of C, and lead to changes in the carbon pool sizes. Arctic ecosystem net C balance may be important on a global scale, because arctic and alpine areas of the world contain 14% of the global soil C-pools (CitationPost et al., 1982), most of which is bound in SOM, which potentially can be released to the atmosphere. On the other hand, the C stock in arctic vegetation is low compared to the global average, implying that the vegetation may constitute a significant, potential sink for atmospheric C over the long term (CitationShaver et al., 1992, CitationJonasson et al., 2001).
Most work on C balance in the Arctic has been done in wet and moist ecosystems (CitationBillings et al., 1982; CitationOberbauer et al., 1991; CitationOechel et al., 1993, Citation1995; CitationJohnson et al., 1996, Citation2000; CitationShaver et al., 1998), whereas drier ecosystem types have been less investigated (but see CitationChristensen et al., 1997; CitationJones et al., 1998; CitationWelker et al., 1999, Citation2000; CitationIlleris et al., 2003). Dry ecosystems cover 44% of the low arctic region holding 12% of the region's C stores. In the High Arctic, dry ecosystems cover 58% of the ice-free areas and contain 42% of the terrestrial C stores of the region (data from CitationBliss and Matveyeva, 1992). Due to the extensive area and large C stores of dry arctic ecosystems, it is essential to understand their responses to climate change, and feedback mechanisms affecting the carbon cycle in these areas. The objective of this study was to investigate the long-term effects of altered temperature, nutrient regimes, and light regimes on the diurnal C balance at peak growing season in a dry dwarf shrub–dominated heath in northern Scandinavia. Also, in order to extend the results to an entire growing season, we investigated daytime flux regularly during a growing season.
The effects of experimental manipulations of temperature, radiation, and nutrient levels on carbon cycling in arctic soils and vegetation are variable. Changes in respiratory CO2 release and soil organic matter and litter decomposition rate in warming experiments with temperature enhancement of a few degrees above ambient range from nonexistent in some studies (CitationNadelhoffer et al., 1991; CitationRobinson et al., 1995, Citation1997; CitationJohnson et al., 2000) to increased release in others (CitationHobbie and Chapin, 1998; CitationJones et al., 1998; CitationWelker et al., 1999). Photosynthesis of many arctic plants has optimum ranges between 7 and 10°C higher than the actual summer temperatures (CitationSemikhatova et al., 1992). As a consequence, all other factors being equal, a temperature increase is likely to increase carbon assimilation by the plant biomass. At the ecosystem level, the carbon balance depends on the differences between the rates of photosynthesis and the rates of respiratory C losses from plants, soil biota, and microorganisms. In spite of expected increase of photosynthesis after warming, the gross ecosystem production (GP), i.e. the photosynthetic gain, has been unchanged in some warming studies (CitationWelker et al., 1999; CitationJohnson et al., 2000), and net ecosystem production (NEP), i.e. the difference between GP and ecosystem respiration (ER) has been variable after warming (CitationHobbie and Chapin, 1998; CitationJones et al., 1998; CitationWelker et al., 1999; CitationJohnson et al., 2000).
Few studies have been done on responses to changes in light conditions. If photosynthetically active radiation (PAR) decreases due to increased cloudiness, photosynthesis may decrease. However, this effect on net C balance may be counteracted by the impact of decreased soil temperature on ER. In one experiment with shading of a wet sedge ecosystem, the NEP, which was positive, decreased (CitationJoabsson et al., 1999), indicating that the impact on GP was larger than the impact on ER.
In comparison with the effects of temperature, changes in C balance seem to be more responsive to alterations in soil nutrient status. Fertilizer addition increases both ecosystem C uptake and C release (CitationChristensen et al., 1997; CitationShaver et al., 1998; CitationJohnson et al., 2000). This is because productivity of arctic ecosystems is nutrient-limited due to the fact that most nutrients are bound in SOM and not available for plants (CitationNadelhoffer et al., 1992).
In order to understand responses of the individual components of ecosystem C balance to warming, fertilizer addition, and shading, we investigated both photosynthetic C incorporation and respiratory C losses from plants and decomposers after a decade of experimental manipulation of the heath ecosystem. We expected to find increased photosynthesis and respiration in response to temperature enhancement and nutrient levels. In contrast, we expected shading to decrease photosynthesis and respiration, particularly if it also resulted in cooler soils. We also expected to find a net C uptake in our control plots, because the measurements were done in the growing season.
Methods
SITE DESCRIPTION
The experiments took place at a dwarf shrub–dominated heath close to the treeline at 450 m a.s.l. in northern Sweden near Abisko (68°19′N, 18°51′E). The climate is subarctic with a 30-yr mean annual temperature of −0.8°C, and a yearly precipitation of 304 mm. The growing season lasts from about mid-June to early September. The vegetation is dominated by Cassiope tetragona (L.) D. Don, which makes up about one third of the aboveground biomass (CitationMichelsen et al., 1996). Other dwarf shrubs such as Empetrum hermaphroditum Hagerup, Vaccinium uliginosum L., Rhododendron lapponicum (L.) Wahlenb., and the shrub Betula nana L. are subdominants (CitationHavström et al., 1993), and mosses are abundant beneath the vascular canopy.
The soil consists of a well-developed humus layer of about 15 cm depth with an organic content of approximately 80% (CitationJonasson et al., 1993). Total soil N and P content is 2.3 and 0.12% of SOM, respectively, but only a small proportion is in inorganic, plant-available form (CitationJonasson et al., 1993). For further details of the climate, vegetation, and soil see CitationHavström et al. (1993), CitationJonasson et al. (1993), and CitationMichelsen et al. (1996).
EXPERIMENTAL SET-UP
Temperature, light, and nutrients have been manipulated in a factorial experiment since 1989. Within six replicated blocks covering about 400 m2 there have been two levels of temperature enhancements (T1 and T2), light reduction (S), and control plots (C), all of these with and without fertilizer addition (F), making altogether eight different treatments over a total of 48 plots. Each treated plot had a ground surface area of 1.2 m × 1.2 m. Temperature was enhanced by erecting a 1.2 × 1.2 m, dome-shaped, 50-cm-high, open-top plastic greenhouses over soil and plants. The plastic used was optical neutral 0.05 mm polyethylene film, which reduced PAR by 9% (CitationHavström et al., 1993). The low-temperature enhancement (T1) greenhouse had a 5- to 10-cm open gap above the ground on two sides, which caused an air temperature rise of 2.8°C. The high-temperature enhancement greenhouse (T2) was closed at the ground surface, causing a temperature rise of 3.9°C (CitationMichelsen et al., 1996). The shading treatment was established using similar constructions covered with sack cloth that caused a 64% reduction of incoming light (CitationHavström et al., 1993). This is similar to the reduction in global radiation under the canopy of the nearby birch (Betula pubescens ssp. tortuosa) forest. Fertilizer was added to the plots each year in June as 10 g N m−2, 2.6 g P m−2 and 9 g K m−2 (in 1989 4.9, 1.3, and 6.0 g m−2 of N, P, and K), except in 1993 and in 1998. The experiments are set up in gently sloping terrain, permitting lateral movement of soil water.
MEASUREMENTS OF CO2 EXCHANGE
At peak growing season of 1998, we measured ecosystem CO2 flux at the 48 plots using a portable Li-Cor 6200 infrared gas analyzer (LI-COR, Lincoln, Nebraska, U.S.A.) connected to a transparent acrylic chamber of 35.9 L fitted with a circulation fan. Two days prior to the flux measurements, aluminum bases of 35 × 35 cm were inserted 5 to 13 cm into the soil. They were equipped with a 2-cm-wide water channel in order to ensure an airtight seal to the chamber. One diurnal measurement was performed on 18–19 July starting at 1000 h the first morning and continuing until 1400 h. the following day. The measurements were done in three block-wise cycles lasting for about 8 h per cycle. Hence, each plot was measured three times, with each measurement approximately 8 h apart. On 19 July, when the measurements were completed, aboveground plant biomass was harvested within the chamber bases. In order to investigate the belowground proportion of the fluxes, a second diurnal measurement of the CO2 fluxes within the harvested plots took place over three block-wise cycles from 1000 h on 21 July ending at 1000 h. the following day. The total diurnal CO2 flux in each plot was estimated by interpolation between the three measured time points.
For practical reasons, it was not possible to keep the greenhouse plastic and the shading screens in place during the measurements. Instead, we covered the chambers with greenhouse plastic and sackcloth where appropriate during measurements.
During the measurements, the Li-Cor system logged chamber CO2 concentration, chamber air temperature and photosynthetic active radiation (PAR) every 20 s during a period of 2 min, i.e. during six consecutive periods, from which the mean value was used for further calculations. A full measurement in one plot consisted of a measurement with a transparent chamber, by which net ecosystem production (NEP) was determined, after which the chamber was briefly removed from the base in order to restore the CO2 level to ambient. When replaced on the base, we covered the chamber with a double layer of dark plastic to exclude light, and a new measurement was performed as above. This measurement yielded the ecosystem (plant plus soil) respiration (ER). Gross ecosystem production (GP) was calculated as the difference between NEP and ER. By convention, C fluxes into the ecosystem were set positive, and fluxes out of the ecosystem were set negative.
After completion of each flux measurement at a plot, we recorded the soil temperatures at 4 and 8 cm depth at three positions inside the chambers. Volumetric soil moisture content was also measured at three positions, using a ThetaProbe ML2x (Delta-T Devices Ltd, Cambridge, UK) following the third measurement cycle of each diurnal measurement.
The harvested aboveground biomass was sorted to species level (except for mosses and lichens), dried to constant mass and weighed. Fine root (<1 mm in diameter) biomass was determined by hand-sorting of roots from one-quarter of each of three soil cores per plot, of 4 cm diameter and 10 cm depth, followed by drying and weighing.
In addition, to obtain a seasonal pattern, daytime fluxes were measured eight times during the 1999 growing season using the same methods as in 1998. Measurements were done between 0800 h. and 1800 h on nonrainy days (14 and 22 June, 2 and 13 July, and on 10, 18, 24, and 30 August). No measurements could be done between 13 July and 10 August because of technical problems. Aluminum bases of 25 × 25 cm used for the 1999 measurements were placed in nonharvested plots in late growing season of 1998. The measurements were done in C, T2, F, and T2+F plots with the Li-Cor fitted with a 13.5-L acrylic chamber. Measurements in shaded plots were not done this year.
DATA TREATMENT
The measured ecosystem fluxes were corrected at each individual plot for the exact chamber volume, and calculated in units per square meter.
To meet assumptions of normality and homogeneous variances, data were tested by Shapiro-Wilk test and Bartlett's test, respectively. Where appropriate, data were transformed using ln, log, or exp. transformations. Treatment effects in the 1998 data were examined by four-way ANOVA (α level of 0.05) with temperature enhancement (three levels: unheated, open greenhouses, and closed greenhouses), fertilizer addition (two levels: unfertilized and fertilized), shading (two levels: unshaded and shaded), and block (six blocks) as factors. The model used for the ANOVA included the interactions between fertilizer addition and shading and between fertilizer addition and temperature enhancement. The block factor was included to separate the effect of possible differences among the blocks from treatment effects. In order to compare mean CO2 fluxes between the individual treatments and the controls, data were also tested by one-way ANOVA followed by Dunnett's test.
Treatment effects of all data across the 1999 season were tested by three-way repeated measures ANOVA with temperature, fertilizer addition, and block as factors, and the interaction between temperature and fertilizer addition was included in the model.
The treatment effects on the fluxes could be caused by different quantities of plant biomass in the treatments. In order to test the influence of the plant biomass on the 1998 fluxes, Pearson product moment correlations were calculated between the total plant biomass and integrated diurnal GP, ER, and NEP across all treatment plots. In order to test for the treatment effects without the confounding effect of differences in biomass, GP, ER, and NEP from 1998 and 1999 were thereafter analyzed by ANOVAs with biomass as a covariate.
Students t-test was used to compare the 1998 ER in nonharvested and harvested plots. All statistical analyses were performed with SAS (CitationStatistical Analysis System Institute, 1997).
Results
ENVIRONMENTAL PARAMETERS
The mean diurnal air temperatures at the first and second measuring dates in 1998 were 12.3 and 13.3°C, respectively (). This is slightly higher than the 30-yr means for July (11.0°C), and close to the average for July the year of measurement (12.7°C). In 1999, mean air temperatures in June (9.8°C), July (10.8°C), and August (7.8°C) were similar to the 30-yr means, with a slightly warmer June (30-yr mean 8.4°C) and a slightly colder August (30-yr mean 9.7°C) than the average ().
The measurements of ecosystem fluxes in the nonharvested plots in 1998 took place on sunny days with high incoming radiation and stable weather conditions. Mean hourly PAR from 18 July, 0800 h to 19 July, 2000 h was 385 μmol photons m−2 s−1 with daytime hourly maxima of 1264 and 1018 μmol photons m−2 s−1 before midday the two days (). This was appreciably higher than the growing season mean of 1998 (data from 25 June to 29 August) with an hourly mean of 269 μmol photons m−2 s−1 and an average hourly diurnal maximum value of 792 μmol photons m−2 s−1. The longer-term mid-July average diurnal value of incoming PAR in the years 1984 to 1996 was 380 μmol photons m−2 s−1, and the average diurnal hourly maximum value was 970 μmol photons m−2 s−1 (from Abisko Scientific Research Station, www.ans.kiruna.se/ans.htm). In 1999, most measurements were done at radiation levels of between 250 and 500 μmol photons m−2 s−1 due to very cloudy weather conditions.
In 1998, only a few millimeters of precipitation fell between the first and last measuring dates 18–19 and 21–22 July. However, the growing season of 1998 was slightly wetter than the long-term average. In 1999, the total precipitation of 169 mm in June, July, and August was 40% higher than the 30-yr mean of 121 mm. In 1998, the soil temperatures at 4 cm depth on 18–19 July ranged from 6 to 18°C with an average of 9.8 ± 0.2°C, and ranged from 5 to 15°C at 8 cm depth with an average of 7.7 ± 0.1°C. A few days later, when the harvested plots were measured, the temperatures were slightly higher, with a mean value at 4 cm depth of 11.3 ± 0.2°C and at 8 cm depth of 9.1 ± 0.1°C. There was no significant difference in soil temperatures among the treatments when compared across all three cycles of measurements. However, during the third cycle (performed between 0400 h. and 1400 h) at the nonharvested plots, soil temperatures were 0.6 and 1.5°C higher in the warmed plots than in the unwarmed plots at 4 and 8 cm depth (ANOVA, P = 0.01 and 0.005, respectively), and the temperature at 8 cm depth was lower in shaded plots during the entire period (P = 0.009).
Mean soil moisture content throughout the organic horizon in 1998 was 0.18 ± 0.01 m3 m−3 on 18–19 July, corresponding to a gravimetric water content of 170% of the soil dry mass. The moisture content was significantly lower in fertilized (0.16 m3 m−3) than nonfertilized (0.20 m3 m−3) plots (P = 0.004), and in the warmed (0.17 m3 m−3) than in nonwarmed (0.19 m3 m−3) plots (P = 0.019). On 21–22 July the soil moisture level in the harvested plots was slightly higher with a mean of 0.20 ± 0.01 m3 m−3 and, as previously, with significantly (P = 0.039) lower values in the warmed plots (not shown).
ECOSYSTEM GAS FLUXES
The overall diurnal pattern of the CO2 fluxes in 1998 in the control plots is shown in . Gross ecosystem production was highest from midday through the afternoon, while ecosystem respiration was most pronounced in late afternoon. Net ecosystem production was negative most of the day; i.e. there was a net flux of CO2 out of the system at a PAR level below about 500 μmol m2 s−1, which is the approximate light compensation level of the ecosystem.
Total plant biomass (vascular aboveground, root, and mosses) was 1059 g m−2 in control plots. The total biomass was unaffected by warming (P = 0.454) and tended to increase, by 16.6%, in fertilized plots compared to controls (P = 0.084), with no significant interactive effects. By contrast, shading significantly (P = 0.001) reduced biomass by 19.2%.
Both diurnal GP and ER at all plots were correlated with total plant biomass (r = 0.53, P = 0.0002 and r = 0.44, P = 0.002, respectively). However, there was no difference in the significance of the fluxes tested with and without biomass as covariate in the ANOVAs, except that the effect of temperature and the combined temperature and fertilizer treatments on ER changed from nonsignificant to significant with the covariate included in the model.
NEP did not differ significantly between the treatments (, ) and, hence, it was consistently negative in all treatments, with losses ranging from 0.5 to 2 g C m−2 d−1. However, warming and shading tended to make NEP more negative (P = 0.12 and P = 0.18, respectively). ER increased in fertilized plots (, ) and there was a significant interaction between the fertilizer and temperature treatments. Even though there was a significant main temperature effect on ER (), Dunnett's test revealed that when treatments were tested individually, only the fertilized and fertilized plus temperature treatments differed from the control. Shading caused a significant decline of the respiration rate (). GP also increased significantly after fertilizer addition, and shading reduced GP (, ), which reached only about 1.3 g C m−2 d−1 as compared with 2.7 g C m−2 d−1 in nonshaded plots. GP in the temperature enhanced plots was not affected by the treatments. There were significant block effects on both GP and ER, probably as a consequence of the sloping terrain.
After harvest of the aboveground biomass, ecosystem respiration was reduced only by 7 to 28% across the different treatments (P < 0.0001, ), suggesting that respiration from belowground was a major component of ER. The difference was more pronounced in the fertilized plots than in the nonfertilized plots. However, the fertilized plots showed a significantly higher ER after harvest than did the nonfertilized plots (). Similarly, ER also was significantly higher in the warmed than in nonwarmed plots () and, as in the nonharvested plots, ER declined after shading.
The seasonal flux measurements of 1999 were similar to, or smaller than the daytime fluxes of the previous season (), and responses to treatments were largely similar (, ). GP in control plots was highest in mid-July and lower both in the beginning and the end of the season, whereas ER was more stable during the season with an exception on 24 August when it was very low. NEP in control plots fluctuated around 0 g C m−2 d−1, but was slightly negative during most of the season ().
As in 1998, ER increased in fertilized plots in 1999 (, ), and was not influenced by temperature enhancement. Also GP increased significantly in fertilized plots across the season, while temperature enhancement had no effect (, ).
Discussion
CO2 FLUXES
The ranges of mid-day CO2 fluxes are close to fluxes reported previously for moss covered ground at this site (CitationChristensen et al., 1997), and from a nearby dry heath (CitationIlleris and Jonasson, 1999). The mid-July, CO2 fluxes measured in 1998 were somewhat higher than CO2 fluxes measured during the 1999 growing season. This is probably due to weather conditions, because the 1998 measurement took place on a warm day with high incoming PAR, while 1999 was cold and cloudy, and the measurements took place at slightly colder days with lower PAR. The NEP in control plots in 1998 was positive only for short time periods of high daytime PAR but was negative when integrated over the diurnal cycle (, ). Similar negative carbon balance was found in all treatments in 1998 (). In 1999 even mid-day fluxes were negative on five of eight days in control plots. We interpret that the negative NEP was due mainly to higher temperature sensitivity of the respiration than of the photosynthesis, leading to a main respiratory control of the carbon flux. Indeed, ecosystem respiration increases exponentially with temperature while GP has a long flat optimum (CitationLindroth et al., 1998). This is in accordance with CitationValentini et al. (2000) who found that net ecosystem production was determined by ecosystem respiration and not by gross primary production, with between-year variations depending on climatic variations along the latitudinal transect of European forests. In ecosystems with low photosynthesizing biomass and high SOM content, this is likely to lead to fluxes out of the system at high temperatures (CitationSoegaard and Nordstroem, 1999; CitationSoegaard et al., 2000). Further, in 1999 GP was constrained by low PAR on mostly cloudy days. During the 1999 growing season 58% of all days between 1 June and 30 August had a mean level of incoming PAR below the compensation point of 500 μmol photons m−2 s−1 between 0800 and 1800 h. The negative C balance we measured was therefore probably caused by a combination of the relationship between a low photosynthesizing leaf biomass of 250 g m−2 in the controls, high stem and root biomass of 620 g m−2, a SOM content of about 5600 g m−2 at this site (CitationJonasson et al., 1999), and high temperatures and/or low incoming PAR values during the days of measurements. Hence, it appears that respiration is the main determinant of the carbon balance in this ecosystem, and the balance between C uptake and loss can vary strongly depending on variations in weather conditions (CitationSoegaard et al., 2000). Low PAR values can, however, affect the C balance further.
The measured CO2 fluxes in both the 1998 and 1999 growing seasons suggest that the ecosystem currently is a source of C to the atmosphere during the growing season. Furthermore, CitationGrogan et al. (2001) found a substantial efflux of CO2 from the plots during the winter season of 1999, indicating that this dry subarctic heath may have experienced an overall annual net loss of C in 1998–1999. Hence, it appears that the large soil C pool probably was built up under climate conditions of different balance between C gains and losses than at present.
Negative diurnal C balances have been reported previously in dry Alaskan tundra (CitationJones et al., 1998; CitationWelker et al., 1999), and warming resulted in increased ER, whereas GP remained unchanged so that NEP decreased. In moist tussock tundra, CitationHobbie and Chapin (1998) found a variation between diurnal positive and negative NEP values depending on weather conditions and phenological stage of the plants, and also CitationOechel et al. (1993, Citation1995) and CitationVourlitis and Oechel (1999) have found both negative and positive growing season C balances in Alaskan moist tussock tundra depending on longer term (decadal) climatic trends.
Ecosystem respiration consists partly of plant-associated respiration, and partly of soil faunal and microbial respiration. Since the diurnal ER was reduced by less than 30% after harvest, and because significant effects of fertilizer addition, enhanced temperature and shading were seen on ER both before and after harvest (, ), we assume that the main part of ecosystem respiration came from root and microbial activity, and that the treatment effects were in part due to changes in belowground respiration. However, we cannot exclude that the slightly higher temperature and moisture levels when we measured the fluxes in harvested plots may have increased respiration somewhat in comparison to the level at the measurement with the biomass included. Also, since we did not measure the CO2 flux in a time series after harvest, we are not able to tell whether the flux measured 2 d after harvest was from heterotrophic below ground respiration only, or whether some of the measured flux was a flush effect from plant exudates. Nevertheless, previous studies suggests that the disturbance caused by harvesting of tundra aboveground biomass results in a short initial flush of CO2 that is dissipated within 1 to 2 d (pers. obs.). Hence, the treatment effects on ER both before and after harvest plus the lower flux from harvested than nonharvested plots indicates that a minimum of 7 to 28% of ER came from aboveground plant parts and, hence, a maximum of 72 to 93% came from soil plus plant roots. This is in the same order of magnitude as CitationCraine et al. (1999) found in a similar harvest experiment in temperate grassland.
TREATMENT EFFECTS
Because the use of biomass as covariate did not affect the outcome of the tests of the data, the increased GP in plots with added fertilizer (, ) was not due to a direct effect of higher biomass. The increased GP in fertilized plots in both 1998 and 1999 must be explained by a higher rate of gross photosynthesis, possibly due to changes in species composition with increased abundance of graminoids with high rate of photosynthesis and decline of cryptogams with low photosynthetic rate (CitationGraglia et al, 2001; Citationvan Wijk et al., 2004). It could also be influenced by increased photosynthesis as a function of higher plant tissue concentration of nitrogen in fertilized plots (CitationJonasson et al., 1999). Indeed, the chlorophyll concentration of the most abundant species, Cassiope tetragona, has earlier been observed to increase in the fertilized plots (CitationMichelsen et al., 1996). In accordance with our expectation, diurnal GP decreased in shaded plots, but contrary to our expectation GP in warmed plots in both 1998 and 1999 remained unchanged. This could be because C. tetragona probably is dominating ecosystem GP. This species grows near its southern distributional limit at Abisko, and specimens growing close to the southern distributional limit of the species react less to temperature changes than when they grow further to the north (CitationHavström et al., 1993; CitationWookey et al., 1994; CitationGraglia et al., 1997).
Although the factorial ANOVA showed a significant temperature × fertilizer interaction on diurnal ER in 1998, the effect was small (), and when tested by Dunnett's test against the control plots it was apparent that the fertilizer effect alone was the principal response (see the strong response in the fertilizer treatments, ), whereas warming only tended to increase ER. Furthermore, in the growing season of 1999, only fertilizer addition affected ER (). Since the main part of ER obviously originated from belowground processes (), the increase of ER after fertilizer addition can either be caused by changes in substrate quality or by changes in the microbial communities. The higher plant tissue concentration of nitrogen in the fertilized plots (CitationJonasson et al., 1999) suggests an improved litter and SOM quality probably leading to a higher microbial activity. Also, although earlier analyses of microbial biomass have not shown any changes (CitationJonasson et al., 1999), CitationRuess et al. (1999) found a strong increase of nematodes both in plots with added fertilizer and in plots with temperature enhancement. Because nematodes are predators on both fungi and bacteria, this indicates that the microbial productivity increased, but this was not detectable as increased microbial biomass because the microbes were predated to about the same level as in the control plots. The lower diurnal GP in shaded plots probably caused lower release of root exudate C (CitationJoabsson et al., 1999), thereby reducing the quantity of readily available substrate for ER. Indeed, CitationCraine et al. (1999) found a 35 to 39% lower soil CO2 efflux in shaded plots than unshaded plots, which was explained by a reduced carbohydrate supply to the microorganisms in the rhizosphere.
The unchanged diurnal NEP in the different treatments in both 1998 and 1999 (, , , ) indicates that even though several treatments altered GP, concomitant changes in ER cancelled out any treatment effect on NEP. The result of an increased flux rate both into and out of the system will be a more rapid C turnover, but changes in temperature, nutrient supply or radiation may not necessarily have any strong effects on the whole ecosystem C pool (CitationHobbie and Chapin, 1998). However, the distribution of the ecosystem C may change as soil C stocks are slowly depleted and plant C pools are enhanced (CitationCallaghan and Jonasson, 1995; CitationMcKane et al., 1997). Indeed, this is for instance supported by the fact that graminoids have increased and cryptogams have decreased in the fertilized plots (CitationGraglia et al., 2001; Citationvan Wijk et al., 2004), and grass litter decomposes faster than cryptogam litter and litter of the dominant dwarf shrubs.
The amount of N added annually with the fertilizer in this decade-long field experiment exceeds current low N deposition in the area 50-fold, and strongly exceeds realistic scenarios of enhanced N deposition, but its application provides important information on processes at plant and ecosystem level. By contrast, the experimentally imposed degree of warming and shading are well within the range from scenarios for warming in the Arctic within this century. It is noteworthy that warming and shading tended (P = 0.12 and P = 0.18, respectively) to reduce mid-summer NEP measured in full sunlight, from −0.9 g C m−2 d−1 in controls to −1.5 and −1.4 g C m−2 d−1 in warmed and shaded plots, respectively. Hence, warming and increased cloudiness may tend to accelerate the loss of C from the belowground components of this ecosystem type that apparently already now is experiencing at least periodically negative NEP.
FIGURE 1. (a) Photosynthetically active radiation (PAR, solid line) and air temperature (dotted line) during the flux measurements in 1998. (b) Gross ecosystem production (GP), ecosystem respiration (ER), and net ecosystem production (NEP) at control plots in 1998. The points on the curves are calculated as running means of three consecutive measurements during the three cycles of the diurnal measurement. They are only used for illustration of the change in fluxes and not for other calculations. The broken lines show the approximate light compensation point and the zero-line for carbon flux in a and b, respectively
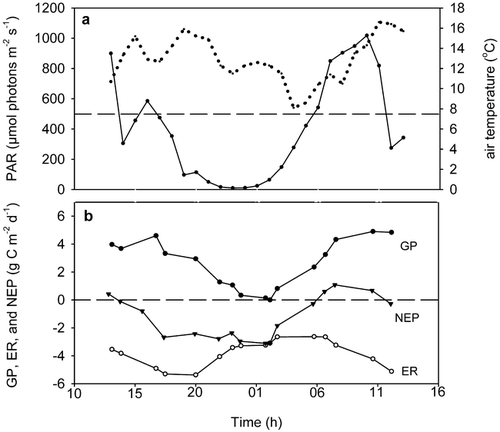
FIGURE 2. (a) Photosynthetically active radiation (PAR, solid line) and air temperature (dotted line) during the flux measurements in 1999. (b) Gross ecosystem production (GP), ecosystem respiration (ER), and net ecosystem production (NEP) at control plots in 1999
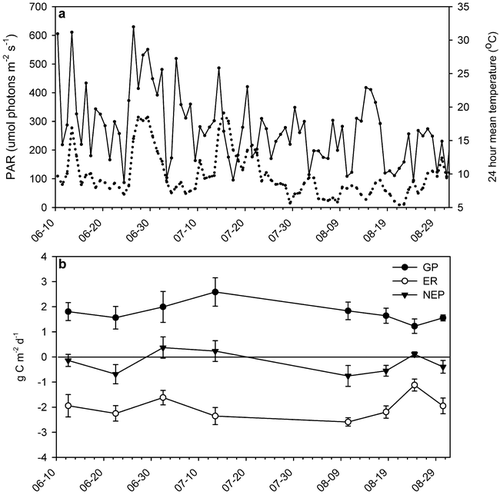
FIGURE 3. Integrated diurnal 1998 mid summer (a) gross ecosystem production (GP), (b) ecosystem respiration (ER), and (c) net ecosystem production (NEP). Data are means ± S.E. of six measurements. Treatments are: C–control, T1–low temperature enhancement, T2–high temperature enhancement, S–shading, F–fertilizer addition
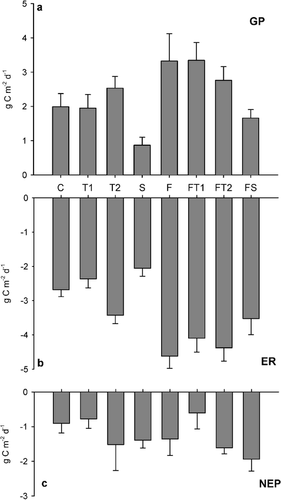
FIGURE 4. Integrated diurnal ecosystem respiration (ER, means ± S.E., n = 6) before (black bars) and after (gray bars) harvest of the aboveground plant biomass. Treatments are: C–control, T1–low temperature enhancement, T2–high temperature enhancement, S–shading, F–fertilizer addition
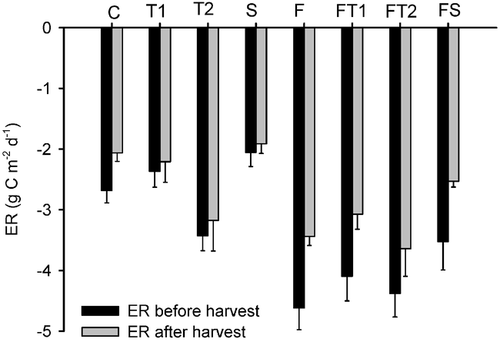
FIGURE 5. Mean seasonal 1999 gross ecosystem production (GP), ecosystem respiration (ER), and net ecosystem production (NEP). Data are seasonal means based on 8 d of measurement ± S.E. (n = 6). Treatments are: C–control, T2–high temperature enhancement, F–fertilizer addition
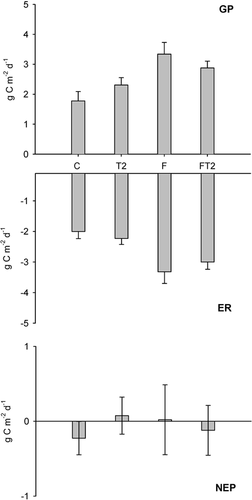
TABLE 1 ANOVAs on diurnal mid-summer CO2 fluxes in 1998 with total plant biomass as covariate. The degrees of freedom (d.f.) of the error-term are 35. Tests without biomass included as covariate gave similar statistical significance of main effects and interactions, except that there were no significant temperature and temperature × fertilizer effect on ER
TABLE 2 Repeated measures ANOVAs on seasonal CO2 fluxes in 1999 with total plant biomass as covariate. The degrees of freedom (d.f.) of the error-term are 35
Acknowledgments
The work was funded by the Danish National Science Research Council, grant 95-01046 and the Swedish Environmental Protection Board, grant 127402. We are much indebted to the Director and the staff at Abisko Scientific Research Station for providing excellent logistic facilities for fieldwork. We are grateful to Peter Jannerup and Lene Østerballe for assistance in the field.
Related Research Data
References Cited
- Abisko Scientific Research Station, Royal Swedish Academy of Sciences. Weather data. www.ans.kiruna.se/ans.htm .
- Billings, W. D. , J. O. Luken , D. A. Mortensen , and K. M. Peterson . 1982. Arctic tundra: A source or sink for atmospheric carbon dioxide in a changing environment?. Oecologia 53:7–11.
- Bliss, L. C. and N. V. Matveyeva . 1992. Circumpolar Arctic vegetation. In: Chapin, F. S., Jefferies, R. L., Reynolds, J. F., Shaver, G. R., and Svoboda, J. (eds.), 1992: Arctic Ecosystems in a Changing Climate. An Ecophysiological Perspective. San Diego: Academic Press, 59–89.
- Callaghan, T. V. and S. Jonasson . 1995. Arctic terrestrial ecosystems and environmental change. Philosophical Transactions of the Royal Society of London A, 352:259–276.
- Christensen, T. R. , A. Michelsen , S. Jonasson , and I. K. Schmidt . 1997. Carbon dioxide and methane exchange of a subarctic heath in response to climate change related environmental manipulations. Oikos 79:34–44.
- Craine, J. M. , D. A. Wedin , and F. S. Chapin III . 1999. Predominance of ecophysiological controls on soil CO2 flux in a Minnesota grassland. Plant and Soil 207:77–86.
- Graglia, E. , S. Jonasson , A. Michelsen , and I. K. Schmidt . 1997. Effects of shading, nutrient application and warming on leaf growth and shoot densities of dwarf shrubs in two arctic-alpine plant communities. Ecoscience 4:191–198.
- Graglia, E. , S. Jonasson , A. Michelsen , I. K. Schmidt , M. Havström , and L. Gustavsson . 2001. Effects of environmental perturbations on abundance of subarctic plants after three, seven and ten years of treatments. Ecography 24:5–12.
- Grogan, P. , L. Illeris , A. Michelsen , and S. Jonasson . 2001. Respiration of recently-fixed plant carbon dominates mid-winter ecosystem CO2 production in a sub-arctic heath tundra. Climatic Change 50:129–142.
- Havström, M. , T. V. Callaghan , and S. Jonasson . 1993. Differential growth responses of Cassiope tetragona , an arctic dwarf-shrub, to environmental perturbations among three contrasting high- and subarctic sites. Oikos 66:389–402.
- Hobbie, S. E. and F. S. Chapin III . 1998. The response of tundra plant biomass, aboveground production, nitrogen, and CO2 flux to experimental warming. Ecology 79:1526–1544.
- Illeris, L. and S. Jonasson . 1999. Soil and plant CO2 emission in response to variations in soil moisture and temperature and to amendment with nitrogen, phosphorus and carbon in northern Scandinavia. Arctic, Antarctic, and Alpine Research 31:264–271.
- Illeris, L. , A. Michelsen , and S. Jonasson . 2003. Soil plus root respiration and microbial biomass following water, nitrogen, and phosphorus application at a high arctic semi desert. Biogeochemistry 65:15–29.
- Joabsson, A. , T. R. Christensen , and B. Wallen . 1999. Influence of vascular plant photosynthetic rate on CH4 emission from peat monoliths from southern boreal Sweden. Polar Research 18:215–220.
- Johnson, L. C. , G. R. Shaver , D. H. Cades , E. B. Rastetter , K. J. Nadelhoffer , A. E. Giblin , J. A. Laundre , and A. Stanley . 2000. Plant carbon-nutrient interactions control CO2 exchange in Alaskan wet sedge tundra ecosystems. Ecology 81:453–469.
- Johnson, L. C. , G. R. Shaver , A. E. Giblin , K. J. Nadelhoffer , E. R. Rastetter , J. A. Laundre , and G. L. Murray . 1996. Effects of drainage and temperature on carbon balance of tussock tundra microcosms. Oecologia 108:737–748.
- Jonasson, S. , M. Havström , M. Jensen , and T. V. Callaghan . 1993. In situ mineralization of nitrogen and phosphorus of arctic soils after perturbations simulating climate change. Oecologia 95:179–186.
- Jonasson, S. , F. S. Chapin III , and G. R. Shaver . 2001. Biogeochemistry in the Arctic: patterns, processes and controls. In Schulze, E. D., Harrison, S. P., Heimann, M., Holland, E. A., Lloyd, J. J., Prentice, I. C., and Schimel, D. (eds.), Global Biogeochemical Cycles in the Climate System. New York: Academic Press, 139–150.
- Jonasson, S. , A. Michelsen , I. K. Schmidt , and E. V. Nielsen . 1999. Responses in microbes and plants to changed temperature, nutrient and light regimes in the Arctic. Ecology 80:1828–1843.
- Jones, M. H. , J. T. Fahnestock , D. A. Walker , M. D. Walker , and J. M. Welker . 1998. Carbon dioxide fluxes in moist and dry arctic tundra during the snow free-season: responses to increases in summer temperature and winter snow accumulation. Arctic and Alpine Research 30:373–380.
- Lindroth, A. , A. Grelle , and A. Morén . 1998. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity. Global Change Biology 4:443–450.
- Maxwell, B. 1992. Arctic climate: Potential for change under global warming. In Chapin, F. S., III, Jefferies, R. L., Reynolds, J. F., Shaver, G. R., and Svoboda, J. (eds.), Arctic Ecosystems in a Changing Climate. An Ecophysiological Perspective. San Diego: Academic Press, 11–34.
- Maxwell, B. 1997. Recent climate patterns in the Arctic. In Oechel, W. C., Callaghan, T. V., Gilmanov, T., Holten, J. I., Maxwell, B., Molau, U., and Sveinbjörnsson, B. (eds.), Global Change and Arctic Terrestrial Ecosystems. Ecological Studies, 124. New York: Springer-Verlag, 21–46.
- McKane, R. B. , E. B. Rastetter , G. R. Shaver , K. J. Nadelhoffer , A. E. Giblin , J. A. Laundre , and F. S. Chapin III . 1997. Climatic effects on tundra carbon storage inferred from experimental data and a model. Ecology 78:1170–1187.
- Michelsen, A. , S. Jonasson , D. Sleep , M. Havström , and T. V. Callaghan . 1996. Shoot biomass, δ13C, nitrogen and chlorophyll responses of two arctic dwarf shrubs to in situ shading, nutrient application and warming simulating climatic change. Oecologia 105:1–12.
- Nadelhoffer, K. J. , A. E. Giblin , G. R. Shaver , and J. A. Laundre . 1991. Effects of temperature and substrate quality on element mineralization in six arctic soils. Ecology 72:242–253.
- Nadelhoffer, K. J. , A. E. Giblin , G. R. Shaver , and A. E. Linkins . 1992. Microbial processes and plant nutrient availability in arctic soils. In Chapin, F. S., III, Jefferies, R. L., Reynolds, J. F., Shaver, G. R., and Svoboda, J. (eds.), Arctic Ecosystems in a Changing Climate. An Ecophysiological Perspective. San Diego: Academic Press, 281–300.
- Oberbauer, S. F. , J. D. Tenhunen , and J. F. Reynolds . 1991. Environmental effects on CO2 efflux from water track and tussock tundra in Arctic Alaska, U. S. A. Arctic and Alpine Research 23:162–169.
- Oechel, W. C. , S. J. Hastings , G. Vourlitis , M. Jenkins , G. Riechers , and N. Grulke . 1993. Recent change of Arctic tundra ecosystems from a net carbon dioxide sink to a source. Nature 361:520–523.
- Oechel, W. C. , G. L. Vourlitis , S. J. Hastings , and S. A. Bochkarev . 1995. Change in arctic CO2 flux over two decades: Effects of climate change at Barrow, Alaska. Ecological Applications 5:846–855.
- Post, W. M. , W. R. Emanuel , P. J. Zinke , and A. G. Stangenberger . 1982. Soil carbon pools and world life zones. Nature 298:156–159.
- Robinson, C. H. , A. Michelsen , J. A. Lee , S. J. Whitehead , T. V. Callaghan , M. C. Press , and S. Jonasson . 1997. Elevated atmospheric CO2 affects decomposition of Festuca vivipara (L.) Sm. litter and roots in experiments simulating environmental change in two contrasting arctic ecosystems. Global Change Biology 3:37–49.
- Robinson, C. H. , P. A. Wookey , A. N. Parsons , J. A. Potter , T. V. Callaghan , J. A. Lee , M. C. Press , and J. M. Welker . 1995. Responses of plant litter decomposition and nitrogen mineralization to simulated environmental change in a high arctic polar semi-desert and a subarctic dwarf shrub heath. Oikos 74:503–512.
- Rowntree, P. R. 1997. Global and regional patterns of climate change: recent predictions for the Arctic. In Oechel, W. C., Callaghan, T. V., Gilmanov, T., Holten, J. I., Maxwell, B., Molau, U., and Sveinbjörnsson, B. (eds.), Global Change and Arctic Terrestrial Ecosystems. Ecological Studies, 124. New York: Springer-Verlag, 82–109.
- Ruess, L. , A. Michelsen , I. K. Schmidt , and S. Jonasson . 1999. Simulated climate change affecting microorganisms, nematode density and biodiversity in subarctic soils. Plant and Soil 212:63–73.
- Semikhatova, O. A. , T. V. Gerasimenko , and T. I. Ivanova . 1992. Photosynthesis respiration and growth old plants in the Soviet Arctic. In Chapin, F. S., III, Jefferies, R. L., Reynolds, J. F., Shaver, G. R., and Svoboda, J. (eds.), Arctic Ecosystems in a Changing Climate. An Ecophysiological Perspective. San Diego: Academic Press, 169–192.
- Shaver, G. R. , W. D. Billings , F. S. Chapin III , A. E. Giblin , K. J. Nadelhoffer , W. C. Oechel , and E. B. Rastetter . 1992. Global change and the carbon balance of arctic ecosystems. Bioscience 42:433–441.
- Shaver, G. R. , L. C. Johnson , D. H. Cades , G. Murray , J. A. Laundre , E. B. Rastetter , K. J. Nadelhoffer , and A. E. Giblin . 1998. Biomass and CO2 Flux in wet sedge tundras: responses to nutrients, temperature, and light. Ecological Monographs 68:75–97.
- Soegaard, H. and C. Nordstroem . 1999. Carbon dioxide exchange in a high arctic fen estimated by eddy covariance measurements and modelling. Global Change Biology 5:547–562.
- Soegaard, H. , C. Nordstroem , T. Friborg , B. U. Hansen , T. R. Christensen , and C. Bay . 2000. Trace gas exchange in a high-arctic valley. 3. Integrating and scaling CO2 fluxes from canopy to landscape using flux data, footprint modeling, and remote sensing. Global Biochemical Cycles 14:725–744.
- Statistical Analysis Systems Institute, 1997. SAS/STAT Users guide. Release 6.12. SAS Institute, Cary, North Carolina, USA.
- Valentini, R. , G. Matteucci , A. J. Dolman , E. D. Schulze , C. Rebmann , E. J. Moors , A. Granier , P. Gross , N. O. Jensen , K. Pilegaard , A. Lindroth , A. Grelle , C. Bernhofer , T. Grünwald , M. Aubinet , R. Ceulemans , A. S. Kowalski , T. Vesala , Ü Rannik , P. Berbigier , D. Loustau , J. Gudmundsson , H. Thergeisson , A. Ibrom , K. Morgenstern , R. Clement , J. Moncrieff , L. Montagnani , S. Minerbi , and P. G. Jarvis . 2000. Respiration as the main determinant of carbon balance in European forests. Nature 404:861–865.
- van Wijk, M. T. , K. E. Clemmensen , G. R. Shaver , M. Williams , T. V. Callaghan , F. S. Chapin III , J. H C. Cornelissen , L. Gough , S. E. Hobbie , S. Jonasson , J. A. Lee , A. Michelsen , M. C. Press , S. J. Richardson , and H. Rueth . 2004. Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: generalisations and differences in ecosystem and plant type responses to global change. Global Change Biology 10:105–123.
- Vourlitis, G. L. and W. C. Oechel . 1999. Eddy covariance measurements of CO2 and energy fluxes of an Alaskan tussock tundra ecosystem. Ecology 80:686–701.
- Welker, J. M. , K. B. Brown , and J. T. Fahnestock . 1999. CO2 flux in arctic and alpine dry tundra: comparative field responses under ambient an experimentally warmed conditions. Arctic,, Antarctic, and Alpine Research 31:272–277.
- Welker, J. M. , J. T. Fahnestock , and M. H. Jones . 2000. Annual CO2 flux in dry and moist arctic tundra: Fields responses to increases in summer temperatures and winter snow depth. Climatic Change 44:139–150.
- Wookey, P. A. , J. M. Welker , A. N. Parsons , M. C. Press , T. V. Callaghan , and J. A. Lee . 1994. Differential growth, allocation and photosynthetic responses of Polygonum viviparum to simulated environmental change at a high arctic polar semi-desert. Oikos 70:131–139.