Abstract
Recovery of tall alpine herbfields from livestock grazing and drought were examined using 30, 0.6 × 0.9 m permanent quadrats photographed in 1959, 1964, 1968, 1978, and 2001 in Kosciuszko National Park, southeastern Australia. Cover attributes were assessed using a 130-point grid superimposed over each slide/photograph. For the 18 permanent quadrats near Mount Gungartan, where grazing ceased in 1958, there was a pattern of increasing vegetation cover. Bare areas were initially colonized by herbs, with native grasses, the lateral expansion of existing shrubs, and other herb species eventually replacing the colonizing herbs. For the 12 quadrats near Mount Kosciuszko, where grazing ceased 15 years before the survey, vegetation cover was high (around 94%) in all surveys except at the end of a long drought. The drought caused an increase in litter at both locations as graminoids died back. Graminoid cover recovered relatively rapidly and had reached pre-drought levels by 1978 and remained high. Herb cover was dynamic after the initial recovery from grazing, with a trend for reduced herb cover after 1964. The pattern of recovery from these two types of landscape level disturbances differed due to the different responses of the major life forms to bare ground, litter, and existing vegetation cover. The study also highlights the benefits of existing long-term monitoring programs.
Introduction
Mountain areas are subject to a range of natural and anthropogenic landscape scale disturbances. Natural disturbances can include unusual (long or short) periods of snow cover, along with frosts, storms, droughts, and fires during the snow-free period (CitationCostin, 1954; CitationBillings, 1974; CitationNagy et al., 2003). Many mountain areas are also extensively used for forestry, livestock grazing, mining, and tourism; activities that can also result in disturbance (CitationKörner, 1999; CitationLoison et al., 2003). Understanding the ecological processes and patterns of alpine ecosystems recovering from natural and anthropogenic disturbances is important for the conservation and management of these important ecosystems (CitationKörner, 1999; CitationLoison et al., 2003). Alpine ecosystems are subject to very severe climatic conditions and have short growing seasons (CitationBillings, 1974; CitationCostin et al., 2000; CitationNagy et al., 2003). Such factors contribute to the slow growth of plants and may cause the rapid deterioration of vegetation and soils in disturbed areas (e.g., by natural events such as needle ice formation and wind erosion) (CitationKörner, 1999; CitationCostin et al., 2000; CitationUrbanska and Chambers, 2002). Monitoring programs (especially those spanning a decade or more) can provide important data on ecosystem processes and patterns including timing and extent of recovery from large-scale disturbance (CitationBakker et al., 1996; CitationGrabherr, 2003).
In Australia, the highest mountains occur in the Australian Alps in the southeast of the mainland. The area has been subject to a range of natural and anthropogenic disturbances including droughts, fires, mining, livestock grazing, and tourism. Currently much of the area is conserved in a series of national parks. Australia's largest contiguous alpine zone, the Kosciuszko alpine zone, is protected in the largest of these parks, Kosciuszko National Park (6980 km2) (CitationGood, 1992). The main alpine zone covers an area of about 100 km2, consisting of an undulating plateau that gradually rises from the east to a series of north-south–aligned peaks along the Main Range, and that rapidly drops off toward steep valleys along the western side (CitationCostin et al., 2000).
The most extensive and biodiverse community in the Kosciuszko alpine area is the tall alpine herbfield. With over 120 taxa, this alliance is dominated by graminoids (including 15 species of Poaceae, and 9 species of Cyperaceae), and a diverse range of herbs including many Asteraceae (32 species). Tall alpine herbfield occurs in areas that experience 4–9 months of snow cover on “average sites” excluding those with persistent snow cover, strong winds, and water-logged or very stony soils (CitationCostin et al., 2000).
There have been three main landscape-level disturbances to the Kosciuszko alpine flora in the last 200 years: fire, livestock grazing, and drought (CitationGood, 1992; CitationCostin et al., 2000). Fires in pre-European times were probably rare, but after the arrival of European graziers fire frequency increased (CitationCostin, 1954; CitationLeigh et al., 1987; CitationBanks, 1989). Low intensity fires became an integral part of management as pastoralists deliberately burned the alpine pastures in spring and/or autumn to remove the mature snowgrasses (predominantly Poa) which were rather unpalatable to livestock and to promote the more palatable herbs and new growth of the grasses (CitationCostin, 1958; Citationvan Rees, 1982). There have been no extensive fires in the Kosciuszko alpine zone since the ban on grazing and burning in 1944, although small sections of the alpine area burned during the extensive wildfires in January/February 2003 (CitationWorboys, 2003).
European settlers introduced sheep and cattle to the area in the 1830s. Previously, the area had experienced little anthropogenic disturbance and none from ungulates (CitationGood, 1992; CitationGreen and Osborne, 1994; CitationCostin et al., 2000). Grazing by cattle and sheep substantially altered the vegetation with livestock numbers often high, ineffectively managed, and beyond the capacity of the vegetation to sustain even seasonal usage (CitationGood, 1995; CitationCostin et al., 2000). Consequently, livestock grazing resulted in decreases in vegetation cover, opening of snow-grass swards, reductions in palatable species, and changes in vegetation patterns and community structure (CitationHelms, 1893; CitationByles, 1932; CitationCostin, 1954; CitationCostin, 1958; CitationCostin et al., 1959; CitationDurham, 1959; CitationCostin et al., 1960; CitationBryant, 1971).
The combination of regular intense grazing and fire also led to severe sheet erosion in the Kosciuszko alpine zone (CitationBryant, 1971). Livestock grazing and associated burning practices around Mount Kosciuszko were banned in 1944 and in the rest of the park in 1958 (CitationGood, 1992; CitationWorboys and Pickering, 2002). However, some livestock grazing occurred in subsequent years (CitationCostin, 1958; CitationWorboys and Pickering, 2002).
Periodic severe droughts are a natural event in many areas in Australia, including in the Australian Alps (CitationCostin, 1954; CitationGood, 1992). Long periods of low rainfall and limited snow cover can result in alpine vegetation dying back during the summer, resulting in a litter layer of dried grasses (CitationCostin, 1954; CitationKirkwood, 2001). In addition to the direct effects of the drought on vegetation, dry periods appear to favor the activity of some herbivores such as case moth larvae which eat the base of the native snowgrasses, resulting in discrete patches of dead grass (CitationCostin, 1954; CitationGreen and Osborne, 1994; CitationKirkwood, 2001). Droughts can result in changes in the cover and composition of the tall alpine herbfield (CitationCostin, 1954; CitationKirkwood, 2001). However, unlike grazing, there appears to be less potential for soil erosion due to the presence of a thick litter of grass persisting during and after the drought.
In the Australian Alps, including Kosciuszko alpine zone, it was possible to examine the response of vegetation to the impacts of grazing and drought as several long-term monitoring programs have been established (CitationClarke, 1998). One of the oldest was established in 1959 in the Kosciuszko alpine area to examine the recovery of vegetation following the cessation of livestock grazing and to provide insights into plant lifecycles and successional processes (CitationWimbush and Costin, 1979). It includes a series of transects and permanent photographic plots (photoquadrats). Although the transect data from 1959 to 1978 have been analyzed (CitationWimbush and Costin, 1979), the photoquadrats have not previously been analyzed.
The photoquadrats provide a comparison of vegetation patterns at two locations (Kosciuszko and Gungartan) with different land-use histories (CitationWimbush and Costin, 1979). Livestock grazing at Kosciuszko was banned in 1944 and at Gungartan in 1958 (CitationWimbush and Costin, 1979). When the permanent photoquadrats were established in 1959, vegetation at Kosciuszko had largely recovered from grazing and was in near natural condition. At Gungartan, there was reduced vegetation cover and evidence of soil erosion, with livestock grazing having only ceased the year before the photoquadrats were established (CitationWimbush and Costin, 1979). During the 42-year period of the survey, the Kosciuszko alpine zone experienced a severe and prolonged drought from around 1961 to 1967 which dramatically reduced the cover of live grasses and increased litter cover (CitationWimbush and Costin, 1979).
The aim of this study was to document vegetation change and revegetation of bare ground from 1959 to 2001. In this paper, we hypothesize that the two locations, Gungartan and Kosciuszko, will differ in vegetation dynamics reflecting the difference in the time since grazing ceased, with large areas of bare ground present at Gungartan at the start of the study. As well, the patterns of community structure and composition at the life form level, seen in vegetation recovering from grazing and vegetation recovering from drought, are likely to differ. Specifically, this paper documents persistence and replacement patterns at the life form level and revegetation of areas of bare ground and litter. Because of the repeated sampling of the same quadrats, it is possible to examine changes in the percentage cover of different life forms, and patterns of replacement at individual points in each photoquadrat.
Methods
STUDY SITES
To examine the patterns of vegetation recovery after grazing, two valleys in the alpine area of Kosciuszko alpine zone in southeastern Australia were selected that differed in grazing history. In 1959, 12 permanent photoquadrats were established in tall alpine herbfield vegetation in a valley near Australia's highest mountain, Mount Kosciuszko (2228 m), where grazing had ceased in 1944 and where the vegetation was considered to be in a relatively stable state in terms of cover and composition by the authors of the original study (CitationWimbush and Costin, 1979). The 12 quadrats were located adjacent to three permanent transects that traversed the valleys. The transects, and therefore photoquadrats, extended down the exposed north-facing slope of Etheridge Range (1980–2000 m), across the flat Rawson Creek arm of the upper Snowy River (∼1920 m), and onto the sheltered south-facing slope of Mount Clarke (1960–2000 m) (CitationWimbush and Costin, 1979) (). The other 18 quadrats were located approximately 23 km northeast, near Mount Gungartan at a site were grazing had only ceased the year before the study, and where vegetation showed greater evidence of disturbance than at Kosciuszko. The quadrats at Gungartan were also located adjacent to three permanent transects, one of which was on a wind-exposed west-facing slope and the other two on a sheltered east-facing slope (2020–2040 m) (CitationWimbush and Costin, 1979) ().
PERMANENT PHOTOQUADRATS
Each corner of the 0.6 × 0.9 m quadrats was marked by a star picket, with the longest side lying across the slope. The quadrats were photographed by Dane Wimbush and Alec Costin in the years 1959, 1964, 1968, and 1978, and these photographs were generously provided to the authors for analysis. In 2001, the photoquadrats were photographed by one of the authors (Scherrer) using an adaptation of the set-up by CitationWimbush et al., (1967). Only one photoquadrat (#43 at Gungartan Middle) could not be located. In addition, three photographs from the year 1959 (#30, #31, and #36) and two photographs from 1968 (#23 and #24) were missing ().
The right-hand photograph of each of the original stereo pair (1959 to 1979) and the single photograph from 2001 for each photoquadrat were analyzed using a dissecting microscope with a transparent dissection plate that was illuminated from below. An evenly spaced grid of 130 point quadrats (10 rows and 13 columns) was engraved on a transparency that was overlaid onto the photoquadrat slides. Distances between point quadrats on the photograph were equivalent to 75 mm in the field. The cover/vegetation at each point was then assessed using a dissecting microscope and recorded as either graminoid, bare, rock, litter, lichen, moss, or a specific genus of shrub or herb. The information obtained for each photoquadrat was percent cover of cover classes, and succession (replacement of one cover class by another) and persistence of cover classes.
DATA ANALYSIS AND DISPLAY
Differences between cover classes were examined using the software package SPSS version 10.0. The Mann-Whitney U test was used to test for significance of differences in vegetation cover between locations. Repeated measures analyses of variance (ANOVA) and repeated measures contrasts were used to test for statistical significance of change over time. Repeated measures analysis accounts for the fact that the samples were not independent with respect to time. The original survey design was pseudoreplicated with respect to land-use history (time since grazing) (CitationHurlbert, 1984). This was an inherent problem accompanying the dataset that could not be corrected and that limited statistical inferences to the plots analyzed (CitationWahren, 1997). Such restrictions, however, do not necessarily prevent useful ecological interpretations, particularly where data have been collected over several decades and where patterns and processes have been well documented (CitationHurlbert, 1984; CitationBakker et al., 1996; CitationWahren, 1997).
Persistence or succession in cover classes was evaluated by comparing the cover at each point quadrat within a photoquadrat of a particular year to the vegetation at the equivalent point quadrat in the photoquadrat of the subsequent survey. Persistence or change for each cover type from one survey to the next was expressed as percentage. The net change between photoquadrats was also calculated for each period.
Results
VEGETATION COVER
Overall percentage cover of live vegetation was close to 95% at the Kosciuszko photoquadrats in all surveys other than 1968, at the end of the drought, when it dropped to below 63% (). Unlike the Kosciuszko photoquadrats, many of the Gungartan quadrats in 1959 had bare areas showing evidence of erosion. Compared to the Kosciuszko quadrats, percentage cover of vegetation at the Gungartan quadrats was significantly lower in 1959 (Mann-Whitney, p = 0.010, U = 33.5), but in 2001, 43 years after the cessation of livestock grazing at Gungartan, there was no longer a significant difference (p = n.s., U = 77.5). During that period, cover of vegetation at Gungartan increased from about 63% in 1959 to above 85% in 2001 (). Vegetation cover at Gungartan decreased to just 50% at the end of the drought in 1968, but was higher than initial levels in each subsequent survey. Thus, the results indicate that, apart from the effect of drought, vegetation cover at Gungartan is recovering as bare areas are slowly being revegetated.
BARE GROUND
The extent of bare areas and exposed rock was clearly higher at the Gungartan photoquadrats at all surveys (, ). Bare areas at Kosciuszko averaged 3% with little variation between surveys, and even by the end of the drought in 1968 only reaching 5%. Bare areas at Gungartan, however, were far greater, averaging 23% over the 42 years. In 1959, bare areas at Gungartan averaged 31% but had declined to 12% in 2001. There was a strong increase in 1968, which was likely to have been the effect of the severe drought in the 1960s. The rate of decline of bare areas appears to be slowing. There was an almost linear decline in bare areas from 1959 to 1978 (excluding the drought affected 1968 result) with a rate of about −0.8% per year (). Between 1978 and 2001 bare areas has declined by 4.2% per quadrat or at a yearly rate of about −0.2%.
Initially bare ground was colonized principally by herbs (27% overall, ). Graminoids and shrubs also replaced bare ground between 1959 and 1964 (11% and 8.1%, respectively). As the drought intensified (1964–1968), the amount of bare ground increased from 15.6% to 22.1% (). As vegetation recovered from the drought, bare areas declined, with replacement by graminoids (27%), herbs (17%), shrubs (7.4%), and a few mosses (4.1%) (). Between 1978 and 2001, there were further declines in bare areas (8.1% overall in 2001), as bare ground was colonized by shrubs (26%), graminoids (27%), and herbs (5.9%), with only 40% of the bare ground in 1978 still not covered by vegetation in 2001. There were differences among quadrats in which life forms invaded the bare areas. For the quadrats with large areas of bare ground in 1959, it was principally graminoids (32%), while for those that had medium levels of bare ground it was shrubs (72%).
LITTER
Litter cover in 1959 and in 2001 was below 4% at both locations, but at the end of the drought in 1968 had increased dramatically to 26% at Kosciuszko and 19% at Gungartan before declining to below 6% by 1978 ( and ). Litter had the highest turnover of any of the ground cover categories, with most litter being replaced by vegetation between survey periods (). In the initial survey period (1959–1964), the small amount of litter present was colonized by graminoids (31%), herbs (31%), shrubs (10%), or became bare ground (18%). With the start of the drought, there was an increase in litter mostly due to the death of graminoids. Some of this litter was colonized by graminoids (14%), herbs (14%), and shrubs (9.4%) even during the latter part of the drought (). By 1968, when the drought had caused extensive areas of graminoids to die back and form litter (36% of graminoid cover in 1964 was litter in 1968), litter cover reached its peak (21.5% of total cover, ). Most of this litter was rapidly recolonized by graminoids in the next 10 years (74%), with small areas being colonized by herbs (8.9%) and shrubs (3.8%) and only 3.2% becoming bare ground. During the final survey period, the small remaining amount of litter (4.7% in 1978, 3.4% in 2001) was again colonized principally by graminoids (71%).
LIFE FORM COVER
Overall, graminoids were the most common vegetation form, averaging above 50% in all surveys apart from 1964 and 1968. Early in the drought in 1964, graminoid cover declined at Gungartan to 30%, dropping even further by the end of the drought in 1968 to just 23%. At Kosciuszko graminoid cover had not declined in 1964, but did so in 1968, with only 24% cover compared to 55% four years previously (, ).
In addition to examining changes in the total proportions of each life form it was possible to document succession among the life forms (). Graminoids were the most stable of the life forms. From 57% to 82% points that had graminoids in one survey were graminoids in the following survey for all intervals except the latter part of the drought (from 1964 to 1968). During the drought graminoids died back, with over a third of graminoid cover replaced by litter (36%). However, 40% of the points still remained as graminoids.
When graminoids were replaced by other life forms, it tended to be herbs. This was part of a dynamic between these two life forms, with herbs replacing graminoids and graminoids replacing herbs over the 43 years examined. The relative balance between the two life forms changed during the study. Initially herb cover increased between 1959 and 1964 due to herbs replacing graminoids faster than the reverse along with bare ground being colonized by herbs. Even during the latter part of the drought (1964–1968) there continued to be a net loss of graminoids to herbs. After the drought (1968–1978), the balance again changed with 10 times the number of points changing from herbs to graminoids than graminoids to herbs. This indicated the post-drought recovery of graminoids. During this period, only 28% of herb points remained herbs, showing just how dynamic the cover of this life form can be. Herb cover continued to decline at Gungartan after a peak during the early stages of the drought (30% herb cover in 1964) until 2001, when it accounted for only 6% of cover. In contrast at Kosciuszko herb cover was 29% in 2001.
Overall mean shrub cover increased from below 2% to 20% in 42 years (). Shrubs were absent in 1959 at the Gungartan photoquadrats, but recorded 4% cover at the Kosciuszko photoquadrats. By 2001, cover had reached 22% and 17% at Gungartan and Kosciuszko, respectively. Shrubs increased between the first two survey periods post-grazing due to their replacement of other life forms/cover. Between 1959 and 1964, they replaced bare ground, graminoids, and to a lesser extent herbs and litter (). The small amount of shrub cover in 1959 (1.5%) was dynamic, with most of it being replaced by bare ground, graminoids, herbs, and litter (). This situation changed from 1964 on as areas that had shrubs increasingly tended to remain shrubs, and there was also a net gain in shrub cover from the other life forms/cover. Most of this increase appears to have occurred due to growth and lateral expansion of existing shrubs rather than establishment of new shrubs from seedlings. Also, although shrubs replaced areas of bare ground, they also replaced graminoids, herbs, and litter.
Mosses (1.3%) and lichen (0.04%) remained uncommon in all surveys ().
Discussion
Results from this study provided quantitative information about the composition of tall alpine herbfield, and in particular recovery patterns from livestock grazing and drought. They support and extend other studies of tall alpine herbfield (or equivalent) both in Kosciuszko (CitationCostin, 1954; CitationCostin et al., 1959; CitationWimbush and Costin, 1979; CitationMallen-Cooper, 1990; CitationJohnston, 1998; CitationCostin et al., 2000; CitationKirkwood, 2001; CitationJohnston, 2003; CitationHill and Pickering, in press) and other alpine areas (e.g., CitationMcDougall, 1982; CitationWalsh et al., 1984; CitationBridle, 2000).
The results highlight some general patterns for the recovery of herbfields/grasslands in mountain regions. For example this and other studies have found that (1) the recovery of alpine climax communities from perturbations is often slow, and can be in the order of decades (CitationUrbanska and Chambers, 2002; CitationErschbamer et al., 2003; CitationGrabherr, 2003); (2) the colonization of bare ground in alpine regions can be very slow and is often slower than the colonization of litter or establishment of seedlings adjacent to existing plants (“nurse effect”) (CitationUrbanska and Chambers, 2002); (3) species/life form composition changes during recovery from disturbance, sometimes with early colonizers such as some herbs replaced by other species such as graminoids (CitationWebber and Ives, 1978; CitationEbersole, 2002; CitationFischer and Wipf, 2002); and (4) recovery patterns can be affected by the intensity of damage, with sites still differing in composition of vegetation decades after the original disturbance (CitationUrbanska and Chambers, 2002; CitationErschbamer et al., 2003).
This study found that in absence of landscape-level disturbances such as livestock grazing and drought, tall alpine herbfield was characterized by nearly entire vegetation cover (∼94% at Kosciuszko and 84% at Gungartan in 2001) with little bare ground or litter. Such an extensive vegetation cover in alpine areas has been found to provide effective control of surface runoff and soil loss, and provides protection for seedling establishment and revegetation of the small patches of bare ground (CitationCostin, 1958; CitationCostin et al., 1959; CitationMallen-Cooper, 1990; CitationJohnston, 1998; CitationUrbanska and Chambers, 2002; CitationJohnston, 2003).
The tall alpine herbfield community was dominated by graminoids (above 50% cover outside the drought period), followed by herbs (22%, average of all years) and shrubs (9%). Similar or slightly higher cover values for graminoids have been reported from other studies of this community in the Kosciuszko alpine area (CitationKirkwood, 2001; CitationScherrer, 2004; CitationHill and Pickering, in press), and from other alpine areas in Australia (Victorian Alps, CitationMcDougall, 1982; Tasmania, CitationBridle, 2000).
PATTERNS OF RECOVERY FROM LIVESTOCK GRAZING
Grazing history and site characteristics influence the effects of livestock grazing on an area's vegetation (CitationLeigh et al., 1991; CitationMilchunas and Lauenroth, 1993). The introduction of livestock grazing to areas with little or no history of grazing can cause physical damage to vegetation and soils and increase pressures on more palatable plant species through selective grazing (CitationCostin, 1958; CitationWimbush and Costin, 1979; Citationvan Rees, 1982; CitationNoy-Meir et al., 1989; CitationMilchunas and Lauenroth, 1993). Trampling can cause physical damage to plants, disturb the soil structure, and change the local hydrology by creating drainage channels (CitationWilliams, 1990; CitationMilchunas et al., 1989).
The flora of the Kosciuszko alpine zone is not adapted to livestock grazing and trampling, as native megafauna are largely absent and there was no livestock grazing until the 1830s (CitationCostin, 1954; CitationGreen and Osborne, 1994; CitationCostin et al., 2000). The deep soils developed in the absence of hoofed mammals and thus are highly susceptible to erosion from such disturbance (CitationCostin, 1954; CitationGood, 1992, Citation1995). The introduction of grazing by cattle and sheep substantially altered alpine vegetation in Australia (CitationCostin, 1954, Citation1958; CitationCarr and Turner, 1959; CitationBryant, 1971; CitationWimbush and Costin, 1979; CitationMcDougall, 1982; CitationWalsh et al., 1984; Citationvan Rees et al., 1985; CitationWilliams and Ashton, 1987; CitationWahren, 1997; CitationBridle and Kirkpatrick, 1999; CitationBridle, 2000).
In areas with native megafauna, such as mountain goats or sheep, the introduction of low numbers of livestock may have less impact, as grazing and trampling are already a natural part of the system. Prolonged selective grazing is likely to have affected the species composition and vegetation stature by reducing the competitive pressures for less palatable and low statured species (CitationNoy-Meir et al., 1989; CitationSal et al., 1986; Citationvan Rees et al., 1985; CitationBook et al., 1995; CitationKiehl et al., 1996; CitationBridle, 2000). Consequently, pastures which are adapted to grazing pressure are often dominated by horizontal growth forms, especially among the palatable species (CitationSal et al., 1986; CitationBridle, 2000). The flora of the European Alps, for example, which has evolved with a native megafauna and centuries of livestock grazing, may have changed with time to a different but stable ecosystem adapted to grazing. In contrast, the dominance of tall forbs observed in alpine areas of the Australian mainland may be a reflection of the absence of vertebrate herbivores (CitationWimbush and Costin, 1979; CitationKirkpatrick, 1989; CitationBridle, 2000).
The impacts of grazing on tall alpine herbfields in Australia were apparent in the 1959 photoquadrats which showed clear differences between the two locations in extent of bare ground, amount of exposed rock, and vegetation cover. For example, at Gungartan in 1959, vegetation cover was only 63% compared to 93% at Kosciuszko, or conversely 31% of the Gungartan photoquadrats were bare ground compared with only 3% at Kosciuszko.
Initially these bare areas at Gungartan were colonized by herbs (1958–1964). A parallel study analyzing data from long-term transects adjacent to the Gungartan and Kosciuszko photoquadrats found that as bare patches became smaller, they started to be colonized by graminoids and shrubs as well as herbs (CitationScherrer, 2004). The parallel transect study showed that herbs such as the naturalized herb Acetosella vulgaris and native herb Euchiton spp. were able to colonize bare areas through germination, while graminoids replaced bare ground largely by colonization from the edges of bare patches (CitationScherrer, 2004).
In this study, bare areas generally consisted of one or more large patches per quadrat, rather than sparse vegetation with intertussock spaces. Colonization of bare ground appears to be dependent on the size of the gap. Smaller gaps in this and other studies are colonized faster than larger gaps, with colonization in the small gaps benefiting from the expansion of adjacent plants and/or from protection provided by these plants to seedlings (nursery effect) (CitationWilliams, 1992; CitationUrbanska and Chambers, 2002).
Shrubs were only a minor component of the tall alpine herbfield community in 1959, but have increased over the past four decades to around 20% of cover in 2001. This increase appeared to be largely due to the growth of established shrubs and less the result of shrub seedling establishment in bare areas, as has been observed in other studies (CitationWilliams and Ashton, 1987; CitationWilliams and Ashton, 1988; CitationWilliams, 1992). There were a few examples, however, of shrubs establishing within maturing herbs as part of secondary succession (CitationScherrer, 2004).
DROUGHT IMPACTS AND RECOVERY
Mountain ecosystems are often subject to distinct weather patterns that are related to local topography and which may contribute to high local variation of precipitation patterns (CitationTheurillat et al., 1998). In alpine areas, such precipitation often occurs in the form of snow and may cover the vegetation for a considerable proportion of the year (CitationKörner, 1999). A drought thus may affect vegetation directly due to lack of water or indirectly due to the absence of a protective snow cover exposing vegetation and soils to extreme temperatures, wind, and radiation. Drought may cause plant stress and even mortality, generally resulting in increases in litter and decreases in vegetation cover (CitationWimbush and Costin, 1979).
Drought conditions at the Kosciuszko and Gungarten study sites during the 1960s caused clear changes in the total vegetation cover and in the relative cover of herbs and graminoids. However, the patterns of change differed between the two locations. It appears that the nearly intact vegetation cover at Kosciuszko was more “drought proof” than at the more recently grazed Gungartan. For example, at Kosciuszko the main effect of the drought was graminoids dying back and being replaced by litter, which only occurred during the latter part of the drought. There was little apparent effect of the drought on total herb cover at Kosciuszko other than a slight increase (5%) between 1959 and 1964 and almost no change (<2% increase) between 1964 and 1968.
In contrast, at Gungartan, herbs and graminoids appeared to be more susceptible to the drought. In addition to herbs colonizing bare ground between 1959 and 1964, they also replaced graminoids resulting in a substantial reduction in graminoid cover by 1964. This could have just been part of the pattern of recovery from grazing alone, or it could be the result of the drought affecting graminoid cover at Gungartan earlier than at Kosciuszko because Gungartan vegetation was still in the early stages of recovery from grazing. After this initial increase in herb cover at Gungartan, herbs in turn were affected by drought. Between 1964 and 1968 herb cover declined as herbs died back and were replaced by bare ground or litter.
Recovery of graminoids at both locations was relatively fast, with graminoid cover increasing to pre-drought levels by 1978 at both locations. Herb cover, however, declined at both locations post-drought with cover in 2001 well below levels recorded in 1959. Similar effects of the drought were recorded in the adjacent transects (CitationWimbush and Costin, 1979; CitationScherrer, 2004).
The replacement of graminoids by herbs between 1958 and 1964 could have been due to a decrease in resource competition as the grasses died in the early stages of the drought and a resulting pulse of nutrients from the dead plant material. Such nutrient boosts have also been observed in post-fire studies (CitationGimeno-Garcia et al., 2000; CitationRomanyà et al., 2001) and following herbivory by native moth larvae which left behind a layer of dead leaves similar to the graminoid litter resulting from drought (CitationKirkwood, 2001). Increased nutrient availability have been shown to enhance the germination and growth of forb species in alpine environments, generally favoring early successional, fast growing species such as colonizing herbs (CitationMallen-Cooper, 1990; CitationKörner, 1999).
The relatively rapid recovery from drought, with the replacement of litter by graminoids, and to a lesser extent herbs found in this study, is consistent with other studies that have found that litter benefits recovery of alpine vegetation by stabilizing the soil surface, decreasing water loss, and moderating soil temperatures, reducing the risk of frost heave (CitationUrbanska and Chambers, 2002).
LIMITATIONS AND BENEFITS OF THE STUDY
There are important advantages of long-term research studies such as this. Shorter term (<5 years) studies and surveys can capture the present state of an environment and are often used to assess seasonal changes (e.g., CitationGreen, 1997; CitationBridle and Kirkpatrick, 2001; CitationBuckland et al., 2001) or for pre- and post-impact assessment (e.g., CitationStrehlow et al., 2002). Such field studies may, however, be affected by shorter term climatic conditions (e.g., persistent freezing conditions without a protective cover by snow), which could distort the results. Longer term (10 to 50+ years) studies may be able to examine longer term trends, cyclical patterns, and provide information on longer term stability of systems, the recovery process from disturbance, and the timeframe involved (e.g., CitationWimbush and Costin, 1979; CitationLundt, 1998; CitationMcDougall, 2001; CitationRoques et al., 2001). Longer term vegetation studies, for example, can provide information on (1) the temporal patterns of vegetation change (what is happening?); (2) successional patterns in response to particular events (why is it happening?); and (3) interactions contributing to successional change (how is it happening?) (CitationBakker et al., 1996). Vegetation studies over 10 to 50 years may facilitate the description of the effects of external causes of change (e.g., low rainfall) and the generation of hypotheses on internal causes and mechanisms of change (e.g., species competition, allelopathy) (CitationBakker et al., 1996). For example, permanent plots monitored for several years or preferably for decades are considered to be central to the assessment of mechanisms and causes of vegetation dynamics and successional change (CitationAustin, 1981; CitationBakker et al., 1996; CitationHerben, 1996).
Ecological studies examining vegetation change in alpine regions over periods of more than 50 years are rare. In recent years, with increased interest in factors such as climate change many old vegetation study sites have been resurveyed and rephotographed (e.g., CitationGrabherr et al., 1994; CitationGrabherr, 2003; CitationMoiseeve and Shiyatov, 2003; CitationPauli et al., 2003; CitationVirtanen et al., 2003). Studies spanning 10–50 years are few, but include several important examples from the Australian Alps. These include the grazing exclusion plots in the Bogong High Plains established by CitationCarr and Turner (1959) and reassessed by CitationWahren (1997) and grazing transects and photoquadrats established by CitationWimbush and Costin (1979) at Kosciuszko and analyzed in this study.
In addition to benefits of utilizing existing long-term studies there are also limitations. One problem common to permanent plot studies is their limited spatial distribution (CitationCarr and Turner, 1959; CitationAustin, 1981; CitationBridle and Kirkpatrick, 1999). Site replication is important, as the extrapolation of results from single-location studies may be limited by chance local events (CitationSwaine and Greig-Smith, 1980). As the permanent photoquadrats in the Kosciuszko alpine zone were not randomly placed, the generalization of the results to the wider alpine area needs to be approached with caution. Nevertheless, the sites were selected based on the considerable experience of the researchers and were considered by them to be representative of the vegetation of the area at the time of establishment (CitationWimbush and Costin, 1979). Furthermore, the photoquadrats were spread over a distance of about 2 km at both locations, providing a considerable spatial spread and independence among quadrats. The observed past and present-day variation between the two locations supports this.
The detection of long-term population trends may be obscured by short-term variation, such as the strong response of graminoids and herbs to drought as observed in this study (CitationLesica and Steele, 1996). Thus a single spatio-temporal pattern observed in long-term plots may result from different processes (CitationHerben, 1996). A frequent and regular sampling regime increases the chances of detecting and separating such patterns (CitationAustin, 1981). Hence, it is recommended that sampling of the photoquadrats continue at shorter intervals, to increase the potential to detect patterns of change and relate them to environmental conditions.
The bird's-eye views provided by photoquadrats such as those used here provide information about only the top layer of vegetation. In tall alpine herbfields, the large graminoids (e.g., snowgrasses), some major herbs, and occasionally shrubs form the top stratum (CitationCostin, 1954). Minor herbs therefore may be underrepresented in scores, unless the top stratum has been removed or damaged by events such as drought, moth herbivory, or trampling. Thus, part of the initial increase by herb cover during the reduction in graminoid cover may be attributed to the increased exposure of minor herbs rather than actual increase in cover.
Conclusions
The results of this study indicate that the recovery of alpine vegetation from the impacts of grazing can take a long time, with differences in the vegetation apparent between the more recently grazed Gungartan quadrats compared to those at Kosciuszko at the start and end of the study, despite a marked decrease in bare ground at Gungartan. The results also highlight that the rate and pattern of vegetation recovery differs depending on whether the disturbance results in a litter layer or bare ground, with more rapid recovery and predominant establishment of graminoids when litter is present (such as after a drought), than when there is bare ground and predominant establishment of herbs (such as after intensive grazing). These findings may have applications for revegetation techniques, such as the application of mulch to disturbed areas to assist a more rapid recovery of vegetation cover. The study again highlights the benefits of longer-term vegetation studies to document and provide understanding of change, forces for change, and their effects.
FIGURE 1. Location of the 30 permanent photoquadrats in the alpine zone of Kosciuszko National Park (shaded areas ≥ 1830 m; contours in meters). (a) Overview of photoquadrat locations; (b) position of the Gungartan photoquadrats; and (c) position of the Kosciuszko photoquadrats
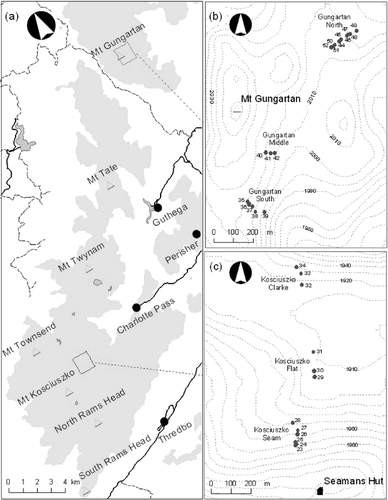
FIGURE 2. Percentage cover of (a) bare ground, (b) litter, and (c) rock in permanent photoquadrats at Kosciuszko (filled squares) and at Gungartan (empty squares) in the alpine zone of Kosciuszko National Park. Values are means ± standard error
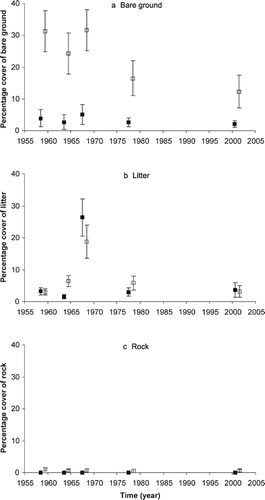
FIGURE 3. Percentage cover of the life forms (a) graminoids, (b) herbs, and (c) shrubs in permanent photoquadrats at Kosciuszko (filled squares) and at Gungartan (empty squares) in the alpine zone of Kosciuszko National Park. Values are means ± standard error
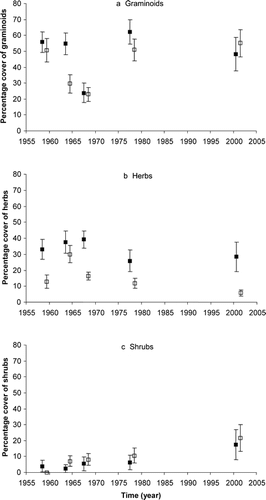
TABLE 1 Summary of location and availability of photographs from 1959 to 2001 at permanent photoquadrats in the alpine zone of Kosciuszko National Park
TABLE 2 Average percent vegetation cover measured at permanent photoquadrats in the alpine zone of Kosciuszko National Park. Values are means ± standard errors (nTotal 1 = 26; nKosciuszko = 10; nGungartan = 16)
TABLE 3 Summary of cover by life forms, litter, rock and bare areas in 30 permanent photoquadrats in the alpine zone of Kosciuszko National Park during five surveys between 1959 and 2001. Values are means ± standard error
TABLE 4 What changes to what: percentage cover by bare ground, litter, graminoids, herbs, and shrubs persisting or changing to a different cover type between the survey periods. Data collected from 30 permanent photoquadrats in the alpine zone of Kosciuszko National Park during five surveys between 1959 and 2001
Acknowledgments
This research was supported by the New South Wales National Parks and Wildlife Service and the Cooperative Research Centre for Sustainable Tourism. The authors thank Dane Wimbush and Alec Costin for their generosity in providing their photographs and answering questions about the details of the original aims and methods. We would also like to thank two anonymous reviewers and Andrea Mayer for their constructive comments on our manuscript.
Notes
Revised ms submitted May 2005
Related Research Data
References Cited
- Austin, M. P. 1981. Permanent quadrats: an interface for theory and practice. Vegetatio 46:1–10.
- Bakker, J. P. , H. Olff , J. H. Willems , and M. Zobel . 1996. Why do we need permanent plots in the study of long-term vegetation dynamics?. Journal of Vegetation Science 7:147–156.
- Banks, J. C G. 1989. A history of forest fire in the Australian Alps. In Good, R. (ed.), The Scientific Significance of the Australian Alps. The Proceedings of the First Fenner Conference 1988. Canberra: Australian Alps National Parks Liaison Committee, 265–280.
- Billings, D. W. 1974. Arctic and alpine vegetation: plant adaptations to cold summer climates. In Ives, J. D., and Barry, R. G. (eds.), Arctic and Alpine Environments. London: Methuen.
- Bock, J. H. , C. L. Jolls , and A. C. Lewis . 1995. The effects of grazing on alpine vegetation: a comparison of the central Caucasus, Republic of Georgia, with the Colorado Rocky Mountains, U.S.A. Arctic and Alpine Research 27:130–136.
- Bridle, K. L. and J. B. Kirkpatrick . 1999. Comparative effects of stock and wild vertebrate herbivore grazing on treeless subalpine vegetation, eastern central plateau, Tasmania. Australian Journal of Botany 47:817–834.
- Bridle, K. L. and J. B. Kirkpatrick . 2001. Impacts of grazing by vertebrate herbivores on the flowering stem production of tall alpine herbs, eastern Central Plateau, Tasmania. Australian Journal of Botany 49:459–470.
- Bridle, K. L. 2000. The Effects of Vertebrate Herbivore Grazing on the Alpine Vegetation of the Eastern Central Plateau, Tasmania. PhD Thesis, School of Geography and Environmental Studies, University of Tasmania, Hobart, Australia.
- Bryant, W. G. 1971. Deterioration of vegetation and erosion in the Guthega catchment area, Snowy Mountains, New South Wales. Journal of the Soil Conservation Service of New South Wales 27:209–226.
- Buckland, S. M. , K. Thompson , J. G. Hodgson , and J. P. Grime . 2001. Grassland invasions: effects of manipulations of climate and management. Journal of Applied Ecology 38:301–309.
- Byles, B. U. 1932. A reconnaissance of the mountainous part of the River Murray catchment in New South Wales. Forestry Bureau Bulletin 13:1–34.
- Carr, S. G M. and J. S. Turner . 1959. The ecology of the Bogong High Plains II. Fencing in grassland C. Australian Journal of Botany 7:34–63.
- Clarke, J. 1998. Science in High Places: Research and Monitoring in the Australian Alps. 1. Introduction and Sites in New South Wales. Canberra: Australian Alps Liaison Committee.
- Costin, A. B. 1954. A Study of the Ecosystems of the Monaro Region of New South Wales with Special Reference to Soil Erosion. Sydney: Soil Conservation Service of New South Wales.
- Costin, A. B. 1958. The Grazing Factor and the Maintenance of Catchment Values in the Australian Alps. Melbourne: Commonwealth Scientific and Industrial Research Organization.
- Costin, A. B. , D. J. Wimbush , D. Kerr , and L. W. Gay . 1959. Studies in Catchment Hydrology in the Australian Alps I. Trends in Soils and Vegetation. Melbourne: Commonwealth Scientific and Industrial Research Organisation, Division of Plant Industry Technical Paper No. 13.
- Costin, A. B. , D. J. Wimbush , and D. Kerr . 1960. Studies in Catchment Hydrology in the Australian Alps. II. Surface Run-off and Soil Loss. Melbourne: Commonwealth Scientific and Industrial Research Organisation, Division of Plant Industry Technical Paper No. 23.
- Costin, A. B. , M. Gray , C. Totterdell , and D. Wimbush . 2000. Kosciuszko Alpine Flora. Melbourne: Commonwealth Scientific and Industrial Research Organisation.
- Durham, L. J. 1959. Indicators of land deterioration in Snowy Mountain catchments. Journal of the Soil Conservation Service of New South Wales 15:251–276.
- Ebersole, J. J. 2002. Recovery of alpine vegetation on small, denuded plots, Niwot Ridge, Colorado, U.S.A. Arctic, Antarctic, and Alpine Research 34:389–397.
- Erschbamer, B. , R. Virtanen , and L. Nagy . 2003. The impacts of vertebrate grazers on vegetation in European high mountains. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 377–396.
- Fischer, M. and S. Wipf . 2002. Effect of low-intensity grazing on the species-rich vegetation of traditionally mown subalpine meadows. Biological Conservation 104:1–11.
- Gimeno-Garcia, E. , V. Andreu , and J. L. Rubio . 2000. Changes in organic matter, nitrogen, phosphorus and cations in soil as a result of fire and water erosion in a Mediterranean landscape. European Journal of Soil Science 51:201–210.
- Good, R. B. 1992. Kosciuszko Heritage. Sydney: National Parks and Wildlife Service of New South Wales.
- Good, R. B. 1995. Ecologically sustainable development in the Australian Alps. Mountain Research and Development 15:251–258.
- Grabherr, G. 2003. Alpine vegetation dynamics and climate change—a synthesis of long-term studies and observations. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 399–409.
- Grabherr, G. , M. Gottfried , and H. Pauli . 1994. Climate effects on mountain plants. Nature 369:448.
- Green, K. 1997. Inter-annual, season and altitudinal differences in invertebrate activity in the Snowy Mountains. Victorian Naturalist 114:222–229.
- Green, K. and W. Osborne . 1994. Wildlife of the Australian Snow-country. Sydney: Reed Books.
- Helms, R. 1893. Report on the grazing leases of the Mount Kosciuszko Plateau. Agricultural Gazette of New South Wales 1893:530–531.
- Herben, T. 1996. Permanent plots as tools for plant community ecology. Journal of Vegetation Science 7:195–202.
- Hill, W. and C. M. Pickering . in press. Vegetation associated with different walking track types in the Kosciuszko alpine area, Australia. Journal of Environmental Management. .
- Hurlbert, S. H. 1984. Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54:187–211.
- Johnston, S. W. 1998. Managing degraded alpine humus soils in Kosciuszko National Park, New South Wales: 1—soil properties. Proceedings of the ASSI National Soils Conference, April. Brisbane: ASSI, Brisbane, 1999 pp.
- Johnston, S. W. 2003. Function and sustainability of Australian alpine ecosystems: studies in the tall alpine herbfield community, Kosciuszko National Park, New South Wales, Australia. In Janet Mackay and Associates (eds.), Celebrating Mountains. Proceedings of an International Year of Mountains Conference, 25–28 November 2002. Canberra: Australian Alps Liaison Committee, 410.
- Kiehl, K. , I. Eischeid , S. Gettner , and J. Walter . 1996. Impacts of different sheep grazing intensities on salt marsh vegetation in northern Germany. Journal of Vegetation Science 7:99–106.
- Kirkpatrick, J. B. 1989. The comparative ecology of mainland Australian and Tasmanian alpine vegetation. In Good, R. (ed.), The Scientific Significance of the Australian Alps. Canberra: Australian Alps National Parks Liaison Committee, 125–142.
- Kirkwood, A. B. 2001. The effects of Grazing by Moth Larvae on Plants and Soil Properties in Tall Alpine Herbfield. Honours Thesis, School of Environmental and Applied Sciences, Griffith University, Gold Coast, Australia.
- Körner, C. 1999. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin: Springer Verlag.
- Leigh, J. H. , D. J. Wimbush , D. H. Wood , M. D. Holgate , A. Slee , M. C G. Stanger , and R. I. Forrester . 1987. Effects of rabbit grazing on subalpine environment I. Herbaceous and shrubby vegetation. Australian Journal of Botany 35:3433–3464.
- Leigh, J. H. , D. H. Wood , A. V. Slee , and M. D. Holgate . 1991. The effects of burning and simulated grazing on productivity, forage quality, mortality and flowering of eight subalpine herbs in Kosciuszko National Park. Australian Journal of Botany 39:97–118.
- Lesica, P. and B. M. Steele . 1996. A method for monitoring long-term population trends: an example using rare arctic-alpine plants. Ecological Applications 6:879–887.
- Loison, A. , C. Toigo , and J. M. Gaillard . 2003. Large herbivores in European alpine ecosystems: current status and challenges for the future. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 351–366.
- Lunt, I. D. 1998. Two hundred years of land use and vegetation change in a remnant coastal woodland in southern Australia. Australian Journal of Botany 46:629–647.
- Mallen-Cooper, J. 1990. Introduced plants in the high altitude environments of Kosciuszko National Park, south eastern Australia. PhD Thesis, Department of Biogeography and Geomorphology, Research School of Pacific Studies, Australian National University, Canberra, Australia.
- McDougall, K. L. 1982. The Alpine Vegetation of the Bogong High Plains. Melbourne: Soil Conservation Authority.
- McDougall, K. L. 2001. Colonization by alpine plants of a stabilized road verge on the Bogong High Plains, Victoria. Ecological Management and Restoration 2:47–52.
- Moiseeve, P. A. and S. G. Shivatov . 2003. Vegetation dynamics at the treeline ecotone in the Ural Highlands, Russia. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 423–435.
- Milchunas, D. G. , W. K. Lauenroth , P. L. Chapman , and M. K. Kazempour . 1989. Effects of grazing, topography and precipitation on the structure of a semiarid grassland. Vegetatio 80:11–23.
- Milchunas, D. G. and W. K. Lauenroth . 1993. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63:327–366.
- Nagy, L. , G. Grabherr , C. Körner , and D. B A. Thompson . (eds.),. 2003. Alpine Biodiversity in Europe. Berlin: Springer, 477 pp.
- Noy-Meir, I. , M. Gutman , and Y. Kaplan . 1989. Responses of Mediterranean grassland plants to grazing and protection. Journal of Ecology 84:439–448.
- Pauli, H. , M. Gottfried , and G. Grabherr . 2003. The Piz Lindard (3411 m), the Grisons, Switzerland—Europe's oldest mountain vegetation study site. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 443–448.
- Romanyà, J. , P. Casals , and V. R. Vallejo . 2001. Short-term effects of fire on soil nitrogen availability in Mediterranean grasslands and shrublands growing in old fields. Forest Ecology and Management 147:39–53.
- Roques, K. G. , T. G. O'Conner , and A. R. Watkinson . 2001. Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall and density dependence. Journal of Applied Ecology 38:268–280.
- Sal, A. G. , J. M. DeMiguel , M. A. Casado , and F. D. Pineda . 1986. Successional changes in the morphology and ecological responses of a grazed pasture ecosystem in central Spain. Vegetatio 67:33–44.
- Scherrer, P. 2004. Monitoring vegetation change in the Kosciuszko alpine zone, Australia. PhD Thesis, School of Environmental and Applied Sciences, Griffith University, Gold Coast, Australia. Accessible at http://adt.caul.edu.au/ .
- Strehlow, K. , J. S. Bradley , J. Davis , and G. R. Friend . 2002. Short term impacts of logging on invertebrate communities in jarrah forests in south-west Western Australia. Forest Ecology and Management 162:165–184.
- Swaine, M. and P. Greig-Smith . 1980. An application of principal components analysis to vegetation change in permanent plots. Journal of Ecology 68:33–41.
- Theurillat, J-P. , F. Felber , P. Geissler , J. M. Bogat , M. Fierz , A. Fischlin , P. Kuepfer , A. Schluessel , C. Velluti , G. F. Zhao , and J. Williams . 1998. Sensitivity of plant and soil ecosystems of the Alps to climate change. In Cebon, P., Dahinden, U., Davies, H. C., Imboden, D. M., and Jaeger, C. C. (eds.), Views from the Alps—Regional Perspectives on Climate Change. London: MIT Press, 225–308.
- Urbanska, K. M. and J. C. Chambers . 2002. High elevation ecosystems. In Perrow, M. R., and Davey, A. (eds.), Handbook of Ecological Restoration, Volume 2: Restoration in Practice, Vol 2. Cambridge: Cambridge University Press, 376–400.
- van Rees, H. 1982. The diet of free-ranging cattle on the Bogong High Plains, Victoria. Australian Rangeland Journal 4:29–33.
- van Rees, H. , W. A. Papst , K. L. McDougall , and R. C. Boston . 1985. Trends in vegetation cover in the grassland community on the Bogong High Plains, Victoria. Australian Rangelands Journal 7:93–98.
- Virtanen, R. , A. Eskelinen , and E. Gaare . 2003. Long-term changes in alpine plant communities in Norway and Finland. In Nagy, L., Grabherr, G., Körner, C., and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer, 411–422.
- Wahren, C. H A. 1997. Vegetation Dynamics on the Bogong High Plains. PhD Thesis, Monash University, Melbourne, Australia.
- Walsh, N. G. , R. H. Barley , and P. K. Gullan . 1984. The Alpine Vegetation of Victoria Excluding the Bogong High Plains Region. Melbourne: Department of Conservation, Forests and Lands.
- Webber, P. J. and J. D. Ives . 1978. Damage and recovery of tundra vegetation. Environmental Conservation 5:171–182.
- Williams, R. J. 1992. Gap dynamics in subalpine heathland and grassland vegetation in south-eastern Australia. Journal of Ecology 80:343–352.
- Williams, R. J. 1990. Cattle grazing within subalpine heathland and grassland communities on the Bogong High Plains: disturbance, regeneration and the shrub-grass balance. Proceedings of the Ecological Society of Australia 16:255–265.
- Williams, R. J. and D. H. Ashton . 1987. Effects of disturbance and grazing by cattle on the dynamics of heathland and grassland communities on the Bogong High Plains, Victoria. Australian Journal of Botany 35:413–431.
- Williams, R. J. and D. H. Ashton . 1988. Cyclical patterns of regeneration in subalpine heathland communities on the Bogong High Plains, Victoria. Australian Journal of Botany 36:605–619.
- Wimbush, D. H. , D. H. Barrow , and A. B. Costin . 1967. Colour stereophotography for the measurement of vegetation. Ecology 48:150–152.
- Wimbush, D. J. and A. B. Costin . 1979. Trends in vegetation at Kosciusko. III Alpine range transects, 1959–1979. Australian Journal of Botany 27:833–871.
- Worboys, G. 2003. A brief report on the 2003 Australian Alps bushfires. Mountain Research and Development 23:294–295.
- Worboys, G. and C. Pickering . 2002. Managing the Kosciuszko Alpine Area: Conservation Milestones and Future Challenges. Cooperative Research Centre for Sustainable Tourism, Griffith University, Gold Coast, Australia.