Abstract
Climate-driven northern limits of cold-hardy conifers, Norway spruce (Picea abies (L.) Karst.) and Scots pine (Pinus sylvestris L.), reach latitude 70°N in Fennoscandia. Spruce timberline is located at significantly lower latitudes than pine timberline, but factors contributing to this exceptional pattern have remained obscure. We assessed soil-water and soil-nutrient regimes across the spruce timberline in Finnish Lapland through the use of terrestrial electromagnetic (EM) measurements. These included soil dielectric permittivity (ε), which is positively correlated to soil-water content (θv); gamma radiation (γ), which is negatively correlated to soil-water content (θv); and electrical conductivity (σa), which is a measure of solute content of a soil. The soil EM data were acquired at 15 sites carrying mature (older than 150 yr) stands dominated by either spruce or pine; the sites fall along a 50-km-long transect that crosses the spruce timberline. The correlation and artificial neural network (ANN) analyses revealed that the soil-moisture and solute content were significantly higher in glacial tills of Norway spruce stands compared to those of stands formed by Scots pine. The stands dominated by spruce were associated with mesic-wet and nutrient-rich tills derived from mafic amphibole and hornblende gneisses of the Tanaelv Belt. Dry and nutrient-poor till derived from felsic garnet gneisses of the Lapland Granulite appeared to constitute an edaphic dispersal barrier for Norway spruce.
Introduction
East of Fennoscandia, spruce (Picea abies ssp. obovata) forms the coniferous tree limit, reaching 66°20′–69°25′N in the valleys of the Ob and Yenisei rivers in western Siberia, whereas the tree limit of Scots pine (Pinus sylvestris L.) reaches only latitudes 64°30′–66°N, respectively (CitationVeijola, 1998; CitationMacDonald et al., 2000; CitationPayette et al., 2002). In Fennoscandia, the northern limits of Norway spruce (Picea abies (L.) Karst.) and Scots pine both reach latitude 70°N, but the timberline of spruce is located at significantly lower latitudes than that of Scots pine (CitationKallio et al., 1971; CitationVeijola, 1998; CitationMacDonald et al., 2000; ). Timberline is defined here as the species-specific northern limit at which a minimum of 30% of forest canopy closure ceases (CitationSihvo, 2002; see definitions in CitationVeijola, 1998). It is known that solitary spruce trees or stands occur north to Scots pine tree limit (CitationKallio et al., 1971; CitationOksanen, 1995; CitationSihvo, 2002), but the reasons for the exceptional position of the spruce timberline in northern Fennoscandia have remained obscure. Even though spruce is a climax species in the (aspen)-birch-pine-spruce successional sequence (CitationPastor et al., 1999), it is absent over a large region in the northern boreal forests of Lapland () where Scots pine is a dominant species and birch (Betula pendula Roth. and B. pubescens Ehrh.) is a principal associate.
Holocene migration of tree species, treeline fluctuations, and forest disturbances are to a great extent associated with climate-driven factors (CitationMoe, 1970; CitationEronen and Hyvärinen, 1987; CitationAgee, 1998; CitationRichardson, 1998; CitationKremenetski et al., 1999; CitationBarber et al., 2000; CitationMacDonald et al., 2000; CitationKullman, 2002; CitationPayette et al., 2002; CitationPitkänen et al., 2002; CitationKirdyanov et al., 2003). On the basis of the radiocarbon (14C) dates of subfossil Scots pine logs, pine spread to northern Fennoscandia by 8000–7000 yr BP and reached its maximum distribution by 6000–4000 yr BP, followed by gradual retreat to its modern range limit (CitationEronen and Hyvärinen, 1987; CitationPayette et al., 2002). As evidenced by 14C-dated subfossil trunks, the expansion of spruce to sites north of the modern treeline in Eurasian Russia occurred by 9000–8000 yr BP, and the retreat of spruce to its current treeline position occurred by 4000–3000 yr BP (CitationKremenetski et al., 1998; CitationMacDonald et al., 2000). Also, spruce macrofossils well above the present alpine tree limit in the Caledonian mountain range in central Sweden indicate that spruce had been a vital part in the forest migration during the early Holocene time period (8000–11,000 yr BP; CitationKullman, 2002). According to the conceptual migration model, spruce reached its northernmost range limit as late as 3000–3300 yr BP in northeast Fennoscandia (CitationMoe, 1970; CitationKremenetski et al., 1999; cf. CitationMacDonald et al., 2000; CitationPayette et al., 2002).
Climatic tolerances of the timberline tree species have been intensively studied in the circumpolar subarctic (CitationNikolov and Helmisaari, 1992; CitationStolbovoi and Nilsson, 1999; CitationKirdyanov et al., 2003). Spruce and pine species in high northern latitudes are able to tolerate a wide range of environmental conditions, but limited information is available on the soil-driven constraints applying to these tree species. Topographic barriers, i.e., mountain ranges, may play an important role in the migration patterns (CitationRupp et al., 2001), but also unfavorable soil conditions may constitute species-specific dispersal barriers in the subarctic region (CitationTimoney et al., 1993; CitationSutinen et al., 2002). Soil-moisture content (θv) and nutrients in soil solution are two of the most important causative factors that control tree growth, species composition, and diversity (CitationTimoney et al., 1993; Citationvan Praag et al., 1997; CitationReimann et al., 1998; CitationRichardson, 1998; CitationVejre, 1999; CitationSalmela et al., 2001; CitationAlawi, 2002; CitationSutinen et al., 2002); hence regional variability in the soil θv and nutrient regimes may also be a critical driver for the species-specific timberline and treeline positions.
We targeted soil electromagnetic (EM) surveys across the Norway spruce timberline in Finnish Lapland () to see whether the soil θv and/or nutrient regimes are different within the timberline spruce forest from those north of the timberline. The measurements of the soil ε, γ, and σa provided us with a basis to assess soil-driven factors, soil-moisture availability, and solute content as contributors to the Norway spruce timberline in northeast Fennoscandia.
Materials and Methods
STUDY AREA
The study area in Finnish Lapland (67°56′N–68°30′N; 26°52′E–27°36′E) is characterized by gently undulating glacial terrain reaching 250–380 m (a.s.l.) elevation (), but with a range of treeless granulite fells (i.e., high altitude and strong relief features above timberline) that reaches 450–718 m a.s.l. The effective temperature sum ranges from 692 to 596 degree-days () as computed according to the model presented by CitationOjansuu and Henttonen (1983). A degree-day is a linear temperature sum unit based on daily mean temperature minus the threshold value, > 5°C. The applied model estimates temperature sums for a given location on the basis of the meteorological data by the Finnish Meteorological Institute (FMI), the digital elevation model, and the presence of water bodies (lakes). Climate is cool and humid; the long-term (1961–1990, FMI) annual average temperature ranges from −1.6°C to −2.1°C, the January mean range is −15.1°C to −15.7°C, and the July mean range is +12.8°C to +13.4°C. The mean annual precipitation is 530 mm, and June–September precipitation is 250 mm (period 1961–1990; FMI). Summer precipitation exceeds evaporation by ∼50 mm, but > 40% of precipitation falls as snow, and snowmelt is the major contributor to the runoff (CitationSolantie, 1987).
The study area is located in the Tanaelv Belt–Lapland Granulite (TB-LG) transition () that represents Archean (1.9 Ga) continental crust (CitationMarker, 1985). Rocks of the mafic TB consist of amphibole and hornblende gneisses with a number of concordant bodies and lenses of metamorphic gabbroic, anorthositic, and ultramafic rocks (CitationMarker, 1985). The TB is similar in lithologic composition to the Lapland Greenstone Belt, which typically is covered with forests dominated by Norway spruce and characterized by understory vegetation containing Hylocomium, Polytrichum, and Sphagnum mosses (CitationMarker, 1985; CitationKatila and Tomppo, 2001; CitationSalmela et al., 2001; CitationSutinen et al., 2002; CitationHyvönen et al., 2003). In contrast to the TB, the LG is composed of felsic garnet gneisses. Because of the resulting dry acidic sandy till as parent soil material, forests in the LG are dominated by Scots pine with understory vegetation characterized by Stereocaulon and Cladonia lichens (CitationSalmela et al., 2001). In the transition zone the mafic granulite parts (veins, inclusions), however, are similar in geochemical composition and structural position to those of the rocks of Tanaelv Belt (CitationMarker, 1985). A set of mafic and felsic rocks constitutes a basis for the contrasting lithogeochemical and thereby for the soil nutritional regimes (CitationReimann et al., 1998) across the Norway spruce timberline. The fine fraction content of parent tills may reach 50% in the Lapland Greenstone Belt and TB, but may be <30% in the LG (CitationHänninen, 1997; CitationRuther, 1999; CitationSalmela et al., 2001). Tills are superimposed on the bedrock only as a 1–4-m-thick veneer, but the tops of the fells frequently are frost-shattered bare rock with no or scarce vegetation. The Podzol (Spodosol) was classified as a Typic Haplocryod (see CitationSutinen et al., 2002).
SOIL EM PROPERTIES
Soil-moisture and nutrient regimes were assessed with the aid of terrain EM measurements coupled with airborne measurements by the Geological Survey of Finland. The soil θv can be measured via dielectric permittivity (ε) with time-domain reflectometry (TDR), electrical capacitance probing (ECP), and/or radar surface arrival detection (RSAD) (CitationTopp et al., 1980; CitationHänninen, 1997; CitationRuther, 1999; CitationSutinen et al., 2002). The soil θv is one of the major contributors to the intensity variations of gamma-ray (γ) flux from glacial materials, and a major part (90%) of gamma radiation is emitted from the upper 30 cm of the soil (CitationGrasty, 1997). Hence the differentiation of the root-zone soil-water regimes under pine and spruce stands can be achieved stand-wise with terrestrial methods and at landscape scale with airborne gamma-ray measurements (CitationGrasty, 1997; CitationHyvönen et al., 2003). The soil-solute content, obtained with electromagnetic induction measurements of the soil electrical conductivity (σa), can be used to estimate soil-nutrient regimes and, along with site-specific calibrations, to estimate stand-scale soil-nutrient distributions (CitationMcBride et al., 1990; see CitationRhoades and Oster, 1986; CitationCorwin and Lesch, 2003).
Fifteen sites () were selected randomly. We applied a systematic grid of points with a spacing of 2 km covering the study area. In order to obtain soil data through the TB-LG transition in a short period of time, the point coverage was then clipped by using a 500-m buffered road network. With the aid of a GIS compartment database by the Finnish Forest and Park Service, only the sites with mature stands (older than 150 yr) and with a dominance of either Norway spruce (>65%) or Scots pine (>65%) were included in the study. In the TB, spruce was dominant, and downy birch (Betula pubescens Ehrh.) was a principal associate. In the LG, pine was dominant, and silver birch (B. pendula Roth.) was a common associate of pine (see CitationSutinen et al., 2002); Norway spruce was absent, except on the Kuusipää site (no. 13 in ). At each selected site, the soil EM properties (ε, σa, and γ) were measured along a 100-m-long line by using 1-m point spacing. Out of a total of 1500 measurement points (microsites), 16 were omitted owing to surface stoniness.
The ε values (i.e., soil θv; see CitationTopp et al., 1980) were determined with an electrical capacitance probe (ECP, Adek Ltd, Estonia). The measuring depth of ECP was ∼10 cm in the mineral soil, and the obtained soil volume was 500 cm3 (CitationSutinen et al., 2002). The most commonly used ε/θv calibration is the one presented by CitationTopp et al. (1980). The gamma-ray flux from the glacial tills under stands dominated either by Scots pine or Norway spruce was measured with a portable GS-256 gamma-ray spectrometer (Geophysica Brno, Czech). The spectral windows (total counts, γTot; potassium, γK; uranium, γU; and thorium, γTh) of the GS-256 are equal to those applied by the airborne gamma-ray spectrometer of the Geological Survey of Finland (CitationHyvönen et al., 2003). At each measuring point, a 1-min counting time was applied. The soil σa was obtained inductively with the EM-38 conductivity meter (Geonics Ltd., Mississauga, Ont.). The EM-38 has an intercoil spacing of 1 m, which results in a penetration depth of ∼0–1.5 m in the vertical dipole orientation coil position (CitationCorwin and Lesch, 2003).
STATISTICAL METHODS
A classification of the tills on sites dominated either by Scots pine or Norway spruce was performed with correlation and artificial neural network (ANN) analyses for the terrestrial soil EM data (γK, γTh, ε, and σa). In principle, an ANN is a massively and parallel distributed processor that is able to store the knowledge contained in the experiential data and makes it available for use (CitationHaykin, 1994). Thereby, knowledge is acquired by the network through a training process. When training an ANN, a set of examples consisting of the input parameters (e.g., γK, γTh, ε, and σa) and the corresponding output parameters (e.g., desired site classification) is presented to the ANN. The rules of how to calculate the ANN output, based on given input parameters, are saved in the internal parameters of the ANN (weights between the neurons and the gains of the activation functions).
On the basis of the terrain geophysical data on 15 sites, two ANNs (ANN1 and ANN2) were trained to differentiate glacial tills typified by stands dominated either by Norway spruce or Scots pine. The classification with ANN1 was based on the measured values of γK, γTh, ε, and σa, but because pine is associated with low soil dielectric permittivities (ε < 15; CitationSutinen et al., 2002) the classification with ANN2 was based on the measured values of γK, γTh, and σa. Of a total of 1500 microsites (i.e., measurement points), 644 for pine and 840 for spruce (owing to stoniness, 16 microsites omitted) were interpreted with the ANNs. For the training of the ANNs, only microsites in the vicinity of pine and spruce trees (i.e., no broad-leaved trees present), 877 in number, were preselected. To train ANN1 we selected 117 microsites (41 for pine and 76 for spruce) from the preselected 877 microsites by using a cluster analysis of the input values. The desired output of the ANN1 was 0.9 for pine and −0.9 for spruce microsites, respectively. In order to train the ANN2 we selected 25 microsites (10 for pine and 15 for spruce from the preselected 877 locations) by using a cluster analysis of the input values. Similar to ANN1, the desired output of ANN2 was 0.9 for pine and −0.9 for spruce microsites.
Within training, the internal parameters of the ANNs were changed by a back-propagation algorithm until the ANNs calculated the appropriate desired output for a specific input pattern. To test the generalization behavior of the trained ANNs, the whole set of the EM data (i.e., for 1484 measured microsites) was classified by the trained ANNs. In this test the mean value of the two output values, one from ANN1 and one from ANN2, was used for the final classification.
Results
The range of soil dielectric values (i.e., soil moisture) in tills of Norway spruce stands (8 < ε < 41) was significantly (t ε = 20.77; P < 0.001) wider compared to that of Scots pine stands (3.5 < ε < 15) (; ). The mean soil ε in the spruce sites ranged from ε = 11 ± 1.84 to ε = 21.26 ± 7.71, but northeast of the spruce timberline in the LG, the mean soil ε in the pine sites was in the range 6.44 ± 1.19 < ε < 10.15 ± 2.57. The till on the isolated spruce site at Kuusipää, presumably on a diabase vein and inside the felsic LG, showed much higher soil moisture (ε = 16.73 ± 7.39) than the surrounding till derived from the LG (; see ).
A significant negative correlation (Spearman's r S = 0.58, P < 0.01) was observed between the soil ε and γK as well as between the soil ε and γTh (r S = 0.39, P < 0.01), respectively. The gamma-ray flux from the tills below Scots pine was observed to be much higher (0.7 ≤ γK ≤ 2.2% K) than the flux below Norway spruce (0.2 ≤ γK ≤ 1.5% K; ; see ). A mean γK = 1.58% K ± 0.28 was found for the soil under Scots pine stands, and a mean γK = 1.1% K ± 0.33 was found for the soil under the Norway spruce stands. A major part (95%) of the observations showed soil γK > 0.9% K under Scots pine stands.
The soil-solute content, as expressed with the soil σa and obtained with the EM-38, was significantly higher (t σ = 38.39; P < 0.05) in tills of Norway spruce stands (0.6 < σa < 3.6 mS/m) compared to those of Scots pine stands (0.1 < σa < 2.2 mS/m; ; ). The mean σa values of the till in pine stands in the felsic LG ranged from σa = 0.42 ± 0.16 to σa = 0.86 ± 0.2 mS/m, whereas the mean σa values for tills in spruce stands in the TB transition ranged from σa = 1.39 ± 0.54 mS/m to σa = 2.04 ± 0.79 mS/m. The isolated spruce site at Kuusipää () yielded a mean σa value of 1.53 ± 0.33 mS/m.
The ANN results () indicated soils of spruce and pine trees to be significantly different on the basis of the terrestrial soil EM data. The ANN outputs that are >0.9 correspond to a reliability of 9.5:1 for pine. In contrast, the ANN outputs smaller than −0.9 correspond to a reliability of >80:1 for spruce. The ANN output of 0 demonstrates that some of the soils in the TB-LG transition could be suitable for both conifer species.
Discussion
DISTRIBUTION OF SPRUCE AND CLIMATE
Spruce timberline parallels the arc-shaped TB-LG transition for ∼100 km in northeast Finnish Lapland (); hence the orientation of the present timberline appears inconsistent with the conceptual model of spruce migration from the southeast during the Holocene (CitationMoe, 1970; CitationKremenetski et al., 1999). However, reconstruction of the postglacial history of spruce is based on pollen records and radiocarbon ages from organic deposits. No macrofossil evidence has been found to indicate distribution of spruce beyond its present timberline. The late expansion of spruce to its northern and northwestern limit by 3000–3300 14C yr BP (CitationMoe, 1970; see CitationKremenetski et al., 1999; CitationPayette et al., 2002) appears to be incoherent with simultaneous retreat of Scots pine as evidenced by pine macrofossils (CitationEronen and Hyvärinen, 1987). As a part of global climatic deterioration—i.e., lower solar insolation and summer temperatures—spruce retreated to its current treeline position at ca. 4000–3000 yr BP in northwest Siberia (CitationMacDonald et al., 2000). Spruce immigrated to the Kola Peninsula between 7000 and 5000 14C yr BP, but radiocarbon dates indicate spruce to have arrived in the northwest Siberian Subarctic at ca. 9000 yr BP, i.e., much earlier than pine (which arrived by ca. 7000 yr BP; CitationKremenetski et al., 1998; CitationMacDonald et al., 2000). The age of subfossil spruce stems (8000–11,000 yr BP) 400–500 m above the present alpine tree limit in the Caledonian mountain range in central Sweden suggests that spruce was not a late successor during early Holocene time (CitationKullman, 2002). Because of the lack of spruce macrofossils in Lapland, however, the conceptual migration model of spruce appears vague with respect to the present spruce timberline.
The Scots pine timberline roughly coincides with the temperature sum of 550 degree-days, but a major part of the spruce timberline is located at significantly lower latitudes (CitationVeijola, 1998; CitationJuntunen et al., 2002; ). Climate-driven factors, crucial determinants of the treeline positions (CitationKallio et al., 1971; CitationNikolov and Helmisaari, 1992; CitationStolbovoi and Nilsson, 1999; CitationMacDonald et al., 2000), seem to play a role in the northwest part of the arc-shaped TB near the Norwegian border, where the 10° to 12°C July isotherm may be associated with the northwestern spruce limit, as shown in Eurasian Subarctic (CitationMacDonald et al., 2000). The annual average temperature range of the study area (from −1.6°C to −2.1°C) is well above the species-specific temperature (annual mean temperature) niches of −2.4°C for pine and −2.3°C for spruce observed in the northern Siberian taiga (CitationStolbovoi and Nilsson, 1999). It seems that spruce, rather similarly to pine, is able to thrive at high altitudes and low-temperature conditions (CitationKallio et al., 1971; CitationNikolov and Helmisaari, 1992; CitationStolbovoi and Nilsson, 1999; CitationMacDonald et al., 2000). The range of the effective temperature sum (692–596 degree-days) at the study sites here () is well above the reported temperature sum minima of 470 degree-days for Norway spruce and 450 degree-days for Scots pine (CitationNikolov and Helmisaari, 1992). Furthermore, the proportion of spruce has a tendency to increase along with degreasing temperature on fertile tills in Lapland (CitationSutinen et al., 2002), hence topoclimatic factors give no explanation for the present spruce timberline in the TB-LG transition.
Climate change in northeast Fennoscandia is evidenced by significant increase in the mean MAM (March-April-May) temperatures over the past 150 yr, particularly since the 1970s (CitationTuomenvirta, 2004). This change appears to be concomitant with the circumpolar reduction of snow-cover area in spring and with earlier “spring greening” (CitationMyneni et al., 1997). On the fertile soil derived from alkaline nepheline syenite rocks of the Khibiny Mountains in the Kola Peninsula, the twentieth-century advance of the spruce treeline suggests an increase in summer temperature by ∼2°C (CitationKremenetski et al., 1999). Also, new regeneration and tree establishment have been observed in the “alpine” spruce treelines in Finland (CitationJuntunen et al., 2002), but no evidence has been found to show new recruitment of spruce on the dry acidic soil of the felsic LG proper. The growth response of Norway spruce to recent climate warming has yet to be studied in the TB-LG transition, but observations elsewhere in the circumpolar region suggest that climate warming does not necessarily result in increased growth of spruce. As an example, reduced growth of Alaskan white spruce (Picea glauca) in the twentieth century has been linked to temperature-induced drought stress (CitationBarber et al., 2000; CitationLloyd and Fastie, 2002). Inverse growth responses to temperature appear to be widespread even in the treeline conditions, but growth declines of white spruce are most common in dry soils (CitationLloyd and Fastie, 2002). In northeast Fennoscandia, climate warming may not result in enhanced migration of spruce to the dry, acidic, and nutrient-poor till of the felsic LG, but the Greenstone Belt–Tanaelv Belt association (), extending north to Norway, provides a potential “polar” pathway for spruce expansion in the future.
SPRUCE TIMBERLINE AND SOIL MOISTURE
The soil ε (i.e., soil θv) was significantly greater in tills below Norway spruce stands in the mafic TB transition compared to those of Scots pine stands in the felsic LG proper (), indicating soil-moisture availability to be one of the most important drivers for the timberline position of Norway spruce. As in several other studies (CitationSalmela et al., 2001; CitationSutinen et al., 2002; CitationHyvönen et al., 2003) the present results demonstrate Norway spruce to be adapted to a much wider soil-moisture regime (8 < ε < 41) compared to the soil-moisture regime to which Scots pine is adapted (3.5 ◃ ε < 15). Spruce seems to be able to grow even on saturated sites (ε > 30; θv > 0.44 cm3 cm−3), according to the dielectric mixing model presented by CitationHänninen (1997), but is absent on sites at the dry end of the soil-moisture gradient (ε < 8; θv < 0.15 cm3 cm−3). In accordance with our results, the sporadic spruce trees and spruce stands along the northwest tree limit have been found to be associated with edaphically moist sites (CitationOksanen, 1995).
Even though the soil θv varies seasonally owing to climatic events, the spatial pattern of the soil θv is primarily dependent on the soil physical properties, such that soil θv positively correlates, e.g., with fine fraction content in tills under forest stands (CitationHänninen, 1997; CitationSutinen et al., 2002). The clay fraction content in tills in Lapland is rather low (<10%); the fine fraction content of parent tills may reach 50% in the Lapland Greenstone Belt and TB, but may be <30% in the LG (CitationHänninen, 1997; CitationRuther, 1999; CitationSalmela et al., 2001). Several studies have shown that the seasonal soil-moisture content is higher in silty tills below spruce stands compared to sandy tills below Scots pine stands (CitationHänninen, 1997; CitationSutinen et al., 2002; CitationHyvönen et al., 2003); hence the variation in the soil θv values (3.5 < ε < 41; 0.042 < θv < 0.52 cm3 cm−3) through the TB-LG transition evidently was due to spatial heterogeneity of the soil physical properties. In the humid climate of Lapland, precipitation exceeds evaporation by ∼50 mm (CitationSolantie, 1987), suggesting that soil texture, structure, and water-retention characteristics, rather than evaporation, are pronounced in determining the spatial distribution of soil-water regimes (CitationHänninen, 1997; CitationSalmela et al., 2001). Relationships between spruce growth and soil moisture are not well known in the Fennoscandian Subarctic (cf. CitationKirdyanov et al., 2003), but based on the southern boreal experiments, reduced growth of Norway spruce is coincidental with soil drought (CitationAlawi, 2002; see CitationBarber et al., 2000; CitationLloyd and Fastie, 2002). In the present spruce timberline soil, drought evidently plays only a minor role in the TB (), but because of low retention characteristics of tills in the LG, the drought effect cannot be ruled out as one of the potential constraints for spruce.
The terrestrial gamma-ray measurements indicated a significant negative correlation between the soil ε (i.e., θv) and the soil γK and soil γTh, hence supporting the previous studies showing the attenuation of gamma-ray flux to be primarily due to the soil θv (CitationGrasty, 1997; CitationHyvönen et al., 2003). The soil γK was significantly greater under Scots pine stands (mean γK = 1.58%) compared Norway spruce stands (mean γK = 1.06%; and ). In accordance with several other reports (CitationTimoney et al., 1993; CitationRichardson, 1998; CitationSutinen et al., 2002; CitationHyvönen et al., 2003), the regional distribution of soil-moisture regimes has been shown here to be soil driven. The soil θv was high (indicated by low soil γK) within the TB transition, but significantly low (high soil γK) in the LG proper as shown by the by the airborne gamma-ray data (). Hence, the 60–80-km-wide LG (see ) may constitute a regional drought barrier for spruce in a similar manner as dry and acidic soils present a barrier for white spruce in the Canadian Subarctic (CitationTimoney et al., 1993).
Scots pine dominates drier habitats with sandy acidic soils, such as till in the LG (; ). Low soil moisture seems to be a prerequisite factor for Scots pine (), such that the edaphic constraint appeared to be ε > 15 (i.e., θv > 0.27 cm3 cm−3). This finding coincides with that of several other reports from Fennoscandia and northwest Russia indicating that Scots pine stands occur only on dry sandy soils (CitationHänninen, 1997; CitationAgee, 1998; CitationKremenetski et al., 1999; CitationMacDonald et al., 2000; CitationSalmela et al., 2001; CitationJasinski and Angelstam, 2002; CitationPitkänen et al., 2002; CitationSutinen et al., 2002; CitationHyvönen et al., 2003). In contrast to the situation in Fennoscandia and the Kola Peninsula, the range limit of Scots pine is at a significantly lower position than that of spruce or Siberian larch (Larix sibirica (Ledeb.)) in the northwest Russian Siberia (CitationMacDonald et al., 2000). The reverse behavior of northern limits of Eurasian spruce and pine species is not well understood, but it may be associated with differences between the soils derived from the rocks of the Baltic Shield and those derived from Cambrian sedimentary rocks in subarctic Siberia.
SPRUCE TIMBERLINE AND SOIL-SOLUTE CONTENT
The soil-solute content, as expressed by the soil σa (CitationRhoades and Oster, 1986), was significantly higher in tills below Norway spruce stands in the TB than those below stands occupied by Scots pine in the felsic LG proper (). The σa values of the till in the LG were low, such that a major part (95%) of the observations were σa < 0.6 mS/m. Even though a strong correlation has been shown between the soil σa and soil attributes associated with forest productivity, such as exchangeable Ca and Mg as well as cation-exchange capacity (CEC) (CitationMcBride et al., 1990), low soil σa values in the till of the LG may only indirectly be associated with the low concentrations of Ca and Mg. However, the concentrations of Ca and Mg are high in tills of the Greenstone Belt–Tanaelv Belt, but low within the felsic Lapland Granulite (CitationReimann et al., 1998). In contrast to these bases, concentration of Al in till of the TB is low compared to that in the LG (CitationReimann et al., 1998). Because the growth and yield of Norway spruce have been observed to be dependent on soil Ca and Mg (CitationVejre, 1999), an inhibitory effect of Al on the uptake of Mg (Citationvan Praag et al., 1997) may disfavor spruce on till of the LG. In a similar way, CitationTimoney et al. (1993) found concentrations of N, Ca, and Mg to be insufficient for white spruce on Canadian Shield tills derived from acidic crystalline rocks.
Previous studies have demonstrated that lithology has pronounced impact on the initial quantity of base cations and nutrient status of the sites (CitationTimoney et al., 1993; CitationReimann et al., 1998; CitationRichardson, 1998); hence the treeline spruce occurrence at the Kuusipää site (; ) inside the LG is puzzling. The mean σa = 1.53 ± 0.33 mS/m of the till indicates a source rock type other than the felsic garnet gneisses of the LG (), possibly the diabase dikes that perpendicularly cross the LG (CitationPihlaja, 1987). On the Kuusipää site, spruce forms a mixed treeline with mountain birch (Betula pubescens ssp. czerepanovii) at 596 degree-days in a similar way as on fertile soil of the alkaline Khibiny Mountains in the Kola Peninsula (CitationKremenetski et al., 1999). The climate-driven (“alpine”) treelines of spruce in Finnish Lapland are attributed to fertile tills derived from mafic and intermediate-composition rock types, such as the ultramafic rocks and amphibolites in Pallastunturi in western Lapland (treeline at 500 m a.s.l. and 594 degree-days) and the biotite-rich diorites in Sarmitunturi in eastern Lapland (410 m a.s.l and 607 degree-days) (see locations in ; CitationJuntunen et al., 2002).
The ANN classification for the terrestrial soil EM data indicated significant differences between the tills of spruce stands and those of pine (), suggesting good generalization behavior of the trained ANN. An advantage of this technique is that with an ANN of proper size (number of neurons), complex relationships between input and output parameters can be represented. A disadvantage of a classification using ANNs is the difficulty of suggesting causal connections to get the desired output from the internal parameters of the ANN. However, the ANN differentiation of spruce and pine coincides with contrasting lithology, lithogeochemistry, soil chemistry, and soil physical properties between the mafic TB and felsic LG (CitationMarker, 1985; CitationReimann et al., 1998; CitationSutinen et al., 2002; ; ). The poor nutrient availability (low soil σa) and low water content (low soil ε and high γK) in the till of the LG evidently are insufficient for spruce (CitationReimann et al., 1998; CitationVejre, 1999; CitationAlawi, 2002; CitationSutinen et al., 2002; see also Citationvan Praag et al., 1997); hence the acidic sandy till of the LG provides low edaphic potential for spruce to migrate north-northeast.
NATURAL DISTURBANCES
In accordance with forest inventories (CitationKallio et al., 1971; CitationKatila and Tomppo, 2001; CitationSihvo, 2002), this study indicated the absence of Norway spruce, but dominance of Scots pine, in the felsic LG (). Here, the percentage of pine may exceed 93%, and silver birch is a common associate of pine (CitationSutinen et al., 2002). Hence, we argue the conceptual model of a static (aspen)-birch-pine-spruce successional sequence, i.e., boreal forests repeat the same regeneration and succession stages after natural disturbances, particularly after forest fires in the boreal environment (see CitationPastor et al., 1999; CitationFrelich and Reich, 1998). According to this static concept, Norway spruce, as a late-successional or climax species, should replace Scots pine, but it appears to be unable to invade nutritionally constraining glacial materials, i.e., granulitic tills with σa < 0.6 mS/m. In contrast, Scots pine, as an early-successional or pioneer species, is absent on fine-grained tills with constraining soil-water content (ε > 15; θv > 0.27 cm3 cm−3; ), i.e., tills derived from fine-grained mafic rocks of the TB and Lapland Greenstone Belt (CitationSutinen et al., 2002; CitationHyvönen et al., 2003; see also CitationRichardson, 1998). Shade-tolerant Norway spruce is known to be adapted to the development of long-term, old-growth continuity; hence the structure of Norway spruce–dominant forests with downy birch as the principal associate of spruce in the TB may in part result from gap dynamics and self-replacement without succession of Scots pine (CitationDrobyshev, 1999).
Successional stages after natural disturbances, such as forest fires, are associated with rotation period and severity of disturbances (CitationAgee, 1998; CitationFrelich and Reich, 1998). Fire-return intervals and fire severity, as attributed to soil-moisture regimes, significantly vary within boreal pine and spruce forests (CitationAgee, 1998; CitationJasinski and Angelstam, 2002). Scots pine has a moderate-severity fire regime (CitationAgee, 1998), but fire susceptibility is much higher for dry pine heaths compared to that for mesic-wet sites of Norway spruce (CitationJasinski and Angelstam, 2002). Through the Holocene (i.e., since 10,000 yr BP), a fire-return interval of 220–260 yr in pine forests has been recorded in eastern Finland (63°N); however, historical fire cycles are only tens of years or less (CitationPitkänen et al., 2002). In contrast to pine heaths, the fire-rotation period of mesic-wet and fertile spruce sites may be as long as 2600 yr (CitationJasinski and Angelstam, 2002). The fire regime of the study area has yet to be described, but the airborne gamma-ray data over the TB-LG transition evidently show discrete soil-moisture regimes (). Classification of the airborne gamma-ray data (CitationHyvönen et al., 2003), associated with studies on charcoal layers in peat deposits (CitationJasinski and Angelstam, 2002; CitationPitkänen et al., 2002), may bring new insight to the rotation and severity of fire disturbances within the soil-driven timberline forests of Norway spruce in Lapland.
FIGURE 1. Airborne magnetic total intensity map of northern Finland. The study area is shown by a black rectangle. Norway spruce timberline (solid lines) (taken from CitationSihvo, 2002) is based on 30% crown coverage. Pine timberline (dashed lines) (adopted from CitationJuntunen et al., 2002) is based on the same crown coverage. LG = Lapland Granulite Belt, TB = Tanaelv Belt (CitationMarker, 1985), LGB = Lapland Greenstone Belt, S = Sarmitunturi, P = Pallastunturi
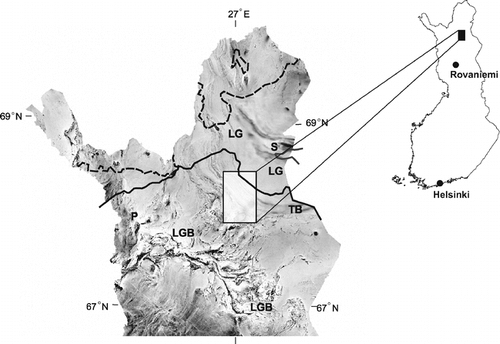
FIGURE 2. Compilation of airborne gamma-ray (potassium window) data and digital elevation model of the study area. Measured sites are at the numbered black dots (see ). Main roads presented as black lines
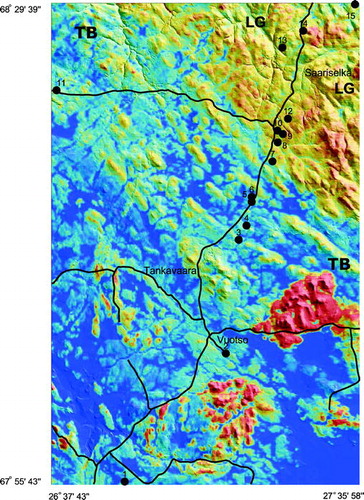
FIGURE 3. Density plot of soil gamma-ray (A), soil dielectric (B), and soil electrical conductivity values (C) for the sites dominated either by Norway spruce or Scots pine in Tanaelv Belt–Lapland Granulite transition in Finnish Lapland
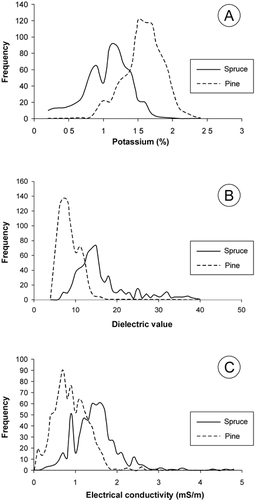
FIGURE 4. Artificial neural network classification for Norway spruce and Scots pine in Tanaelv Belt–Lapland Granulite transition in Finnish Lapland
TABLE 1 Site locations, lithology, tree dominance, elevation, temperature sum, mean soil dielectric values, soil gamma-ray values, and soil electrical conductivity of the study sites
Acknowledgments
This research was a part of the projects Mosaic pattern of forest soils and Timberline, financially supported by the Ministry of Agriculture and Forests and the Academy of Finland. We thank Päivi Hannula, Reijo Raasakka, and Markku Virtanen for field assistance, Viena Arvola for editorial help, and two anonymous referees for providing constructive comments. We acknowledge the staff of the Finnish Forest and Park Service, Yrjö Norokorpi and Juha Sihvo for allowing us to use the biotope inventory data from Lapland, and Olli Sandström and Juha Salmi for allowing us to use the PATI forest compartment database.
Notes
Revised ms submitted February 2005
Related Research Data
References Cited
- Agee, J. K. 1998. Fire and pine ecosystems. In Richardson D. M. (ed.), Ecology and biogeography of Pinus. Cambridge: Cambridge University Press, 193–213.
- Alawi, G. 2002. The impact of soil moisture on stem growth of spruce forest during a 22-year period. Forest Ecology and Management 166:17–33.
- Barber, V. A. , G. P. Juday , and B. P. Finney . 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 405:668–673.
- Corwin, D. L. and S. M. Lesch . 2003. Application of soil electrical conductivity to precision agriculture: Theory, principles and guidelines. Agronomy Journal 94:455–471.
- Drobyshev, I. V. 1999. Regeneration of Norway spruce in canopy gaps in Sphagnum-Myrtillus old-growth forest. Forest Ecology and Management 15:71–83.
- Eronen, M. and P. Hyvärinen . 1987. Radiocarbon-dated subfossil pines from Finnish Lapland. Geografiska Annaler 69A:297–304.
- Frelich, L. E. and P. B. Reich . 1998. Disturbance severity and threshold responses in the boreal forest. Conservation Ecology [online] 2(2): 7. Available from the Internet. URL: http://www.consecol.org/vol2/iss2/art7 (accessed 15 August 2005).
- Grasty, R. L. 1997. Radon emanation and soil moisture effects on airborne gamma-ray measurements. Geophysics 62:1379–1385.
- Hänninen, P. 1997. Dielectric coefficient surveying for overburden classification. Geological Survey of Finland, Bulletin, 396: 72 pp.
- Haykin, S. 1994. Neural networks: A comprehensive foundation. New York: Macmillan College Publishing Company.
- Hyvönen, E. , M. Pänttäjä , M-L. Sutinen , and R. Sutinen . 2003. Assessing site suitability for Scots pine using airborne and terrestrial gamma-ray measurements in Finnish Lapland. Canadian Journal of Forest Research 33:796–806.
- Jasinski, K. and P. Angelstam . 2002. Long-term differences in the dynamics within a natural forest landscape-consequences for management. Forest Ecology and Management 161:1–11.
- Juntunen, V. , S. Neuvonen , Y. Norokorpi , and T. Tasanen . 2002. Potential for timberline advance in northern Finland, as revealed by monitoring during 1983–99. Arctic 55:4 384–361.
- Kallio, P. , U. Laine , and Y. Mäkinen . 1971. Vascular flora of Inari Lapland. 2. Pinaceae and Cupressaeae. Reports from Kevo Subarctic Research Station 8:73–100.
- Katila, M. and E. Tomppo . 2001. Selecting estimation parameters for the Finnish multisource National Forest Inventory. Remote Sensing of Environment 76:16–32.
- Kirdyanov, A. , M. Hughes , E. Vaganov , F. Schweingruber , and P. Silkin . 2003. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 17:61–69.
- Kremenetski, C. V. , L. D. Sulerzhitsky , and R. Hantemirov . 1998. Holocene history of the northern range limits of some trees and shrubs in Russia. Arctic and Alpine Research 30:317–333.
- Kremenetski, C. , T. Vaschalova , and L. Sulerzhitsky . 1999. The Holocene vegetation history of the Khibiny Mountains: Implications for the postglacial expansion of spruce and alder on the Kola Peninsula, northwestern Russia. Journal of Quaternary Science 14:29–43.
- Kullman, L. 2002. Boreal tree taxa in the central Scandes during the Late-Glacial: Implications for Late-Quaternary forest history. Journal of Biogeography 29:1117–1124.
- Lloyd, A. H. and C. L. Fastie . 2002. Spatial and temporal variability in the growth and climate response of treeline trees in Alaska. Climatic Change 52:481–509.
- MacDonald, G. M. , A. A. Velichko , C. V. Kremenetski , O. K. Borisova , A. A. Goleva , A. A. Andreev , L. C. Cwynar , R. T. Riding , S. L. Forman , T. W D. Edvards , R. Aravena , D. Hammarlund , V. N. Szeicz , and V. N. Gattaulin . 2000. Holocene treeline history and climatic change across northern Eurasia. Quaternary Research 53:302–311.
- Marker, M. 1985. Early Proterozoic (c. 2000–1900 Ma) crustal structure of the northeastern Baltic Shield: Tectonic division and tectogenesis. Norges Geologiske Undersøkelse, Bulletin 403:55–74.
- McBride, R. A. , A. M. Gordon , and S. C. Shrive . 1990. Estimating forest soil quality from terrain measurements of apparent electrical conductivity. Soil Science Society of America Journal 54:290–293.
- Moe, D. 1970. The post-glacial immigration of Picea abies into Fennoscandia. Botaniska Notiser 122:61–66.
- Myneni, R. B. , C. D. Keeling , C. J. Tucker , G. Ascar , and R. R. Nemani . 1997. Increased plant growth in northern high latitudes from 1981 to 1991. Nature 386:697–702.
- Nikolov, N. and H. Helmisaari . 1992. Silvics of the circumpolar boreal forest tree species. In Sugart, H., Leemans, R., and Bonan, G. (eds.), A system analysis of the boreal forest. Cambridge: Cambridge University Press, 13–84.
- Ojansuu, R. and H. Henttonen . 1983. Estimation of the local values of monthly mean temperature, effective temperature sum and precipitation sum from the measurements made by the Finnish Meteorological Office. Silva Fennica 17:143–160. (In Finnish with English summary.).
- Oksanen, L. 1995. Isolated occurrences of spruce, Picea abies , in northernmost Fennoscandia in reaction to the enigma of continental mountain birch forests. Acta Botanica Fennica 153:81–92.
- Pastor, J. , Y. Cohen , and R. Moen . 1999. Generation of spatial patterns in boreal forest landscapes. Ecosystems 2:439–450.
- Payette, S. , M. Eronen , and J. J P. Jasisnski . 2002. The circumpolar tundra-taiga interfaces: Late Pleistocene and Holocene changes. Ambio Special Reports 12:15–22.
- Pihlaja, P. 1987. The diabase of Laanila. Geological Survey of Finland—Report of Investigation 76:189–197.
- Pitkänen, A. , P. Huttunen , H. Jungner , and K. Tolonen . 2002. A 10 000 year local forest fire history in a dry heath forest in eastern Finland, reconstructed from charcoal layer records os a small mire. Canadian Journal of Forest Research 32:1875–1880.
- Reimann, C. , M. Äyräs , V. Chekushin , I. Bogatyrev , R. Boyd , P. de Caritat , R. Dutter , T. E. Finne , J. H. Helleraker , Ø Jæger , G. Kashulina , O. Lehto , H. Niskavaara , V. Pavlov , M-L. Räisänen , T. Strand , and T. Volden . 1998. Environmental geochemical atlas of the central Barents region. Trondheim, Norway: Geological Survey of Norway, NGU-GTK-CKE Special Publication, 745 pp.
- Rhoades, J. D. and J. D. Oster . 1986. Solute content. In Klute, A., (ed.), Methods of soil analysis: Part 1. Physical and mineralogical methods (2nd ed.). Agronomy Monograph 9: 985–1006.
- Richardson, D. M. (ed.),. 1998. Ecology and biogeography of Pinus. Cambridge: Cambridge University Press, 527 pp.
- Rupp, T. S. , F. S. Chapin III , and A. M. Starfield . 2001. Modeling the influence of topographic barriers on treeline advance at the forest-tundra ecotone in northwestern Alaska. Climatic Change 48:399–416.
- Ruther, A. 1999. Electromagnetic characteristics of forest till soils on Norway spruce timberline in Finnish Lapland. M.Sc. thesis, University of Vienna, Vienna, Austria, 55 pp.
- Salmela, S. , R. Sutinen , and P. Sepponen . 2001. Understorey vegetation as an indicator of water content in till soils in Finnish Lapland. Scandinavian Journal of Forest Research 16:331–341.
- Sihvo, J. 2002. Ylä-Lapin luonnonhoitoalueen ja Urho Kekkosen kansallispuiston luontokartoitus. Loppuraportti osa 2: Ylä-Lapin luontotyypit. Metsähallituksen luonnossuojelujulkaisuja, Sarja A, no. 137.
- Solantie, R. 1987. Seasonal water balance components in hydrological regionalization. Aqua Fennica 17:133–145.
- Stolbovoi, V. and S. Nilsson . 1999. Forest and temperature associations of Russia relating to global climate warming. Laxenburg, Austria: IIASA, International Institute for Applied Systems Analysis, Interim Report IR-99-006, 26 pp.
- Sutinen, R. , A. Teirilä , M. Pänttäjä , and M-L. Sutinen . 2002. Distribution and diversity of tree species with respect to soil electrical characteristics in Finnish Lapland. Canadian Journal of Forest Research 32:1158–1170.
- Timoney, K. P. , G. H. La Roi , S. C. Zoltai , and A. L. Robinson . 1993. Vegetation communities and plant distributions and their relationships with parent materials in the forest-tundra of north-western Canada. Ecography 16:174–188.
- Topp, G. C. , J. L. Davis , and A. P. Annan . 1980. Electromagnetic determination of soil water content: Measurements in coaxial transmission lines. Water Resources Research 16:574–582.
- Tuomenvirta, H. 2004. Reliable estimation of climatic variations in Finland. Finnish Meteorological Institute Contribution 43: 82 pp.
- van Praag, H. J. , P. Dreze , and M. Cogneau . 1997. Effects of aluminum on calcium and magnesium uptake and translocation by root segments of whole seedlings of Norway spruce ( Picea abies Karst). Plant and Soil 189:267–273.
- Veijola, P. 1998. The northern timberline and timberline forests in Fennoscandia. The Finnish Forest Research Institute Research Papers, 672: 242 pp.
- Vejre, H. 1999. Stability of Norway spruce plantations in western Denmark: Soil nutrient aspects. Forest Ecology and Management 114:45–54.