Abstract
This study is presented against the background that climate warming is predicted to continue in much of the Arctic through the next century and that small greenhouse chambers have been used widely to warm tundra communities in order to forecast climate-related changes. It reports results from up to 8 years of experimental warming with ∼1 m2 open-top chambers (OTCs) at four tundra communities near Barrow (71°18′N, 156°40′W) and Atqasuk (70°29′N, 157°25′W) in northern Alaska. Between 1994 and 2002 the OTCs increased the mean growing season air temperature by between 0.6 and 2.2°C, depending on the site and year. The change in average July soil temperature recorded over 3 years at 10 cm depth due to the OTCs varied between −0.8 and 0.7°C, depending on the site. Changes in soil temperature did not result in detectable differences in thaw depth at any site. This is interpreted to be the result of the small size of the OTCs and possibly changes in vegetation. The differences in warming profiles between sites are important for the biological interpretation of manipulative warming experiments. These profiles also illustrate that vertical heat exchange varies according to plant community type and that this may be an important consideration as the region undergoes climatic change.
Introduction
Concerns about the regional impacts of climatic change have motivated an increased focus on the relationship between species and ecosystem structure and function and temperature over the past decade (CitationIPCC, 2001a; CitationRoot et al., 2003). Most attention to date has been given to climate warming, due to the significance of increased temperature on many biological processes (CitationLong and Woodward, 1988; CitationMinorsky, 2002) and the increase in temperature being experienced globally (CitationIPCC, 2001b).
There are now many groups of researchers experimentally warming plant and animal communities to forecast change. The most prominent of these include the International Tundra Experiment (ITEX, see CitationHenry and Molau, 1997; CitationArft et al., 1999; CitationHollister et al., 2005a; CitationWalker et al., 2006), and the Global Change and Terrestrial Ecosystem Network of Experimental Warming Studies (GCTE-NEWS, see CitationShaver et al., 2000; CitationRustad et al., 2001). Warming experiments are now prevalent in many community types but are greatest in number (over 30 in ITEX alone, CitationHollister, 2003) and of longest duration (e.g., over 20 yr, CitationChapin et al., 1995; CitationChapin and Shaver, 1996) in tundra environments. This is partly because the Arctic is predicted to warm more than other regions of the world (CitationIPCC, 2001a) and tundra communities are believed to be vulnerable to changes in temperature (CitationBliss et al., 1973; CitationEverett and Fitzharris, 1998). At this writing, at least 70 published papers have reported vegetation change resulting from small (generally ∼1 m2) warming manipulations in tundra systems (CitationHollister, 2003).
Passive warming experiments, which warm small patches of tundra, have been popular to date because of their effectiveness in remote tundra-dominated locations with extreme climates and where line power and other technical services are often limited. An undesirable characteristic of chamber-based warming is the limited ability to control the amount of temperature change, which has been criticized in some reviews (CitationDebevec and MacLean, 1993; CitationKennedy, 1995a; CitationMarion et al., 1997; CitationWookey and Robinson, 1997). In a review of several passive chamber designs, CitationMarion et al. (1997) found that chambers warm the average daily temperature, increase the daily range of temperature, reduce canopy turbulence, lower incident radiation, and reduce relative humidity. Chambers were also found to have a variable effect on soil temperatures and may interfere with herbivory and pollination. CitationMarion et al. (1997) detected no change in CO2 concentration, soil moisture, or thaw depth. CitationMarion et al. (1997) concluded that as the design of chambers became increasingly closed the warming potential of the chamber increased, as did the potential impact from unwanted experimental artifacts. For these reasons preference has been given to open-top chamber (OTCs) in most studies.
Owing to a lack of standardization, the reviews cited above (CitationDebevec and MacLean, 1993; CitationKennedy, 1995a; CitationMarion et al., 1997; CitationWookey and Robinson, 1997) were unable to unambiguously isolate the cause for the differences in response to chamber design or site characteristics. This paper describes and contrasts the long-term performance of OTCs at four study sites differing in location and tundra type, and emphasizes the response of soil temperature and thaw depth to experimental warming. The objective is to identify and describe the differences in performance of the same chamber design across sites.
The response of plant species to warming at these sites is reported elsewhere (CitationHollister and Webber, 2000; CitationHollister, 2003; CitationHollister et al., 2005a, Citation2005b). In warmed plots, canopy height has increased, there has been an increase in the growth of many vascular plant species (CitationHollister et al., 2005b), and an increase in the relative cover of graminoids or shrubs and a decrease in lichens or bryophytes (CitationHollister et al., 2005a). Although there has not been an increase in leaf area or above-ground biomass, there has been an increase in standing dead plant matter (CitationHollister, 2003).
Methods
EXPERIMENTAL DESIGN AND DATA COLLECTION
Study sites were established in wet and dry vegetation communities near Barrow (71°18′N, 156°40′W) and Atqasuk (70°29′N, 157°25′W) on the North Slope of Alaska (). A brief description of each site, with a key to abbreviations used in subsequent parts of this paper, is presented in . Each site contained 24 experimental plots and 24 control plots in which biotic measurements were made. Each site was established by choosing plots and randomly assigning the treatment factor: warming with the use of hexagonal OTCs constructed of Sun-Lite HPTM fiberglass (Solar Components Corporation, Manchester, New Hampshire). The transmittance of the fiberglass in the visible wavelengths is 86% and less than 5% in the infrared. The design of the 35 cm high chambers incorporates inward-sloping sides separated by 103 cm at the base and 60 cm at the top. The OTCs cover an area of approximately 1 m2. Control plots were 1 m2.
Chambers were installed soon after snowmelt and removed in mid- to late August at the end of each field season. The day of snowmelt was recorded for each plot when all snow within the plot had melted. In years when the site was not observed until after snowmelt due to logistic constraints involved in accessing sites, the date on which plots became snow free was estimated based on nearby soil temperatures. Owing to logistical constraints, 15 August was specified as the last day of a given field season. The growing season in this study, therefore, is defined as extending from the onset of snow-free conditions within the plots, until 15 August. This was reasonable for nearly all vegetative measurements because it occurred after growth had ceased (CitationTieszen, 1972; CitationJohnson and Tieszen, 1976; CitationHollister et al., 2005b). Although it is somewhat early for measurement of active-layer thickness, increases late in the season are normally minor owing to the small increases in heat accumulation in late August and September and the square root relationship between thaw and heat accumulation.
Regional climate measurements, including temperature at screen height (2 m, 107 temperature probe), precipitation (35 cm height, TE525 tipping bucket rain gage), light intensity, and wind speed (35 cm height, 03001 wind sentry), were measured every 15 min, averaged or summed (for rain), and recorded every hour on Automatic Weather Stations (CR10X datalogger) at the dry heath sites at Barrow and Atqasuk. The sensor and logger specifications cited above refer to products from Campbell Scientific Inc., Logan, Utah, U.S.A. Light intensity was recorded every 10 to 20 min and averaged every hour with StowAway™ single-channel miniature dataloggers (Onset Computer Corporation, Pocasset, Massachusetts, U.S.A.). Correlations between the stations and the National Oceanic and Atmospheric Administration's Climate Monitoring and Diagnostics Laboratory (NOAA/CMDL) in Barrow (CitationStone et al., 1996) were used to estimate temperature at screen height when the Campbell weather stations were not operational. A description of this analysis is provided in CitationHollister (2003). The regional climate measurements outlined above were only collected at the dry heath sites. The dry sites were within 500 m of the wet sites. Any reported differences between sites within a geographic region are attributable to differences in snowmelt timing and the subsequent differences in the length of growing season between sites.
Each growing season, plot-based temperature was measured in 5 to 10 of the 24 control/warmed plots. The plots in which temperature sensors were placed were determined randomly each year. At each site an additional two plots per treatment were established in 1998 to provide more detailed information about the effects of OTCs on air and soil microclimate. Data from these plots are referred to as “plot microclimate” hereafter whereas data from the subset of plots in which biotic measurements were made are referred to as “site temperature.” Aerial sensors were housed in radiation shields placed within the plant canopy at approximately 13 cm above the ground and connected to HOBO® or StowAway™ loggers (Onset Computer Corporation, Pocasset, Massachusetts). Soil temperature was measured at depths of 0, 5, 10, 15, 30, and 45 cm using TP101M temperature probes (Measurement Research Corporation, Gig Harbor, Washington, U.S.A.) connected to Campbell Scientific CR10X dataloggers. Temperature was measured every 10 to 80 min.
Depth of thaw was measured at least once at the end of the growing season at each of the 24 plots per treatment where biotic measurements were made. Thaw depths were measured to the nearest cm by inserting a graduated metal rod into the ground until the frozen surface was reached.
DATA ANALYSIS
Reported temperatures were averaged per treatment and point in time. Thawing degree-days based on 0°C were totaled beginning on the day after snowmelt for each plot using hourly data and are henceforth referred to as TDDsm. When no data were recorded within an hour (for recording intervals of greater than 1 h) or if the data were considered erroneous, the mean of measurements immediately before and after was used. When canopy height temperatures were not recorded, screen height temperatures for the region were used to estimate the missing data prior to calculating TDDsm (CitationHollister, 2003). Estimated values for erroneous measurements represented less than 5% of the data set. All plots within a treatment were then averaged to represent mean TDDsm per treatment and site on any given day for each year.
Indices of wind speed, solar intensity, and precipitation were calculated on a relative scale, based on daily conditions. Mean daily wind speed and light intensity were classified as “less than average,” “average,” and “above average.” Wind was classified as “average” if the average daily wind speed near the ground was between 2.4 and 3.9 m sec−1. Light intensity was classified as “average sunny” if mean intensity from 0800 to 1600 h was between 3.40 and 3.55 lum m−1. Precipitation was classified as “no rain,” “trace,” or “rainy.” The latter category was used whenever the daily accumulation totaled more than 1 mm. Rain was classified as trace if the daily sum was between 0 and 1 mm.
A variation of the Stefan solution for thaw depth (sensu CitationJumikis, 1977) was used to describe the relationship between temperature and thaw. Following CitationNelson and Outcalt (1987), a simplified version of the Stefan solution can be written in linear form as
where Z
t represents thaw depth, E is an “edaphic term” representing soil thermal, textural, surface, and moisture properties, and C is a “climatic term” defined as the square root of the thawing degree-day sum (DDT), measured at screen height and scaled to compatible units. The close dependence of end-of-season thaw depth on the square root of the thawing index facilitates treatment of thaw as a linear regression problem. The rate of thaw progression (i.e., the slope of the regression), therefore, is governed by the edaphic factor E, the characteristic values of which are associated with specific land cover classes. This approach has been used successfully in both boreal and tundra locations by CitationHinkel and Nicholas (1995), CitationNelson et al. (1997), CitationBrown et al. (2000), CitationKlene et al. (2001), and CitationShiklomanov and Nelson (2002). The latter two studies demonstrated that characteristic values of E for specific vegetation-soil associations are relatively stable on an interannual basis. Because TDDsm, measured at canopy height, was considered in this study to be a better representation of microclimate than DDT, we used regressions of the form
to relate thaw depth to temperature.
End-of-season thaw depth was analyzed for treatment effect as a single factor repeated ANOVA using CitationSAS (2000). The analyses were run separately for each site, and an overall analysis was run on all sites.
Results
SITE TEMPERATURE
The average date of snowmelt and growing season temperature was variable among sites, years, and treatments (). A three-factor ANOVA of the canopy height temperature data presented in found site treatment and year to be significantly different (P values of <0.001 for each factor). Recorded screen height growing season temperature at Atqasuk was, on average, between 3.8 and 5.2°C warmer than at Barrow depending on the year (1999–2001). Mean screen height growing season temperatures were up to 0.9°C warmer in the wet communities than the dry communities within a region (Barrow or Atqasuk) due to later snowmelt at wet sites, and the measurement period therefore becoming biased to the warmer duration of the snow-free period. Mean growing season temperature at screen height was between 0.4 and 0.9°C lower than canopy height temperatures (except where temperatures were estimated). A two-factor ANOVA of the amount of experimental warming presented in found site and year to be significantly different (P values of <0.001 and 0.041, respectively). The average growing-season temperature at canopy height was between 0.6 and 2.2°C higher in the warmed plots than in control plots, depending on the site and year.
TDDsm varied between 169 and 854, depending on site, year, height, and treatment (); the relative difference between sites, heights, and treatments was also variable between years. TDDsm was higher at Atqasuk than at Barrow, higher in dry heath sites than in wet meadow sites, higher when measured at canopy height than at screen height, and higher in warmed plots than in control plots. Average growing season TDDsm, estimated at screen height at each site, was 569, 555, 243, and 227 for the AD, AW, BD, and BW sites, respectively. The average increase in TDDsm when calculated at canopy height, rather than at screen height, at sites AD, AW, BD, and BW, was 11.5, 10.9, 22.9, and 20.5%, and the average difference between warmed and control plots was 15.5, 9.1, 36.8, and 34.4%, respectively.
PLOT MICROCLIMATE
Mean daily July temperature varied by day (). The overall pattern of above-ground temperatures was similar between sites within years. This was particularly true for control plots within a geographic region (Barrow or Atqasuk). Below-ground temperatures generally followed the seasonal above-ground progression, but the strength of the relationship lessened with increasing depth below ground surface. The relative effect of OTCs varied greatly by day, and the patterns were different between sites. The largest OTC warming at canopy height occurred during days that were sunny, had no rain, and were windy.
The mean July vertical temperature profile for 1999–2001 showed that the presence of the OTCs did not disrupt the normal sequence of the vertical temperature profile: average daily July temperature increased from screen height to the ground surface and decreased gradually with soil depth (). Mean minimum July air temperature was approximately the same regardless of height or treatment, and average July air temperature recorded over the control plots was approximately the same within geographic regions (Barrow and Atqasuk) and year. The largest differences between sites were temperature at the ground surface. The change in temperature with depth was variable between sites, treatments, and years. Most of the warming at canopy height within OTCs was due to increases in daily maximum temperatures. The warming effect of OTCs was generally greatest at canopy height or at the ground surface, and decreased with depth. The average difference between treatments in July soil temperature for years 1999–2001 at 10 cm depth was 0.6, −0.8, 0.3, and 0.7°C for the AD, AW, BD, and BW sites, respectively.
THAW DEPTH
Average thaw depth at the end of the growing season () varied significantly between sites and years (P value <0.001) but not between treatments (P value = 0.567). The same results were obtained when analyses were run by site: interannual differences were significant (P value <0.001) but differences between treatments were not (P value: AD = 0.818, AW = 0.279, BD = 0.635, BW = 0.132). When the first two weeks of thaw depths were analyzed separately, there was no overall effect of treatment, but there was a significant interaction between treatment and site (P value = 0.009). The √(TDDsm) parameter was correlated with the progression of thaw depth at all four sites (). The correlation between thaw depth and √(TDDsm) was strongest when analyzed by treatment. The slope of the line describing the relationship between thaw depth and √(TDDsm), used to estimate the edaphic term E, varied by site. Within individual sites the slope was lower for the OTC plots relative to the control plots.
Discussion
OTC performance varied considerably in the different tundra communities studied. The amount of OTC warming was greatest for maximum daily temperatures and varied by day, leading to substantial differences between years. Differences in the amount of warming between sites were most pronounced at the soil surface and resulted in differences in the amount of soil warming. The large differences in warming at the soil surface should be viewed with caution because minor differences in microtopography and sensor placement have the potential to greatly influence soil surface temperature (CitationKlene et al., 2001). Temperature at 10 cm depth, which is a more spatially integrative and robust measurement, also showed considerable variability in OTC soil warming. While other studies have also found differences in chamber soil warming between sites, the differences were small or the interpretation was confounded by differences in chamber design (CitationCoulson et al., 1993; CitationWookey et al., 1993; CitationMarion et al., 1997).
The vertical and temporal distribution of temperatures was similar to previously reported tundra studies (e.g., CitationCorbet, 1972; CitationWeller and Holmgren, 1974). Nearly all the observed differences in microenvironment between sites in a region occurred at the soil surface or below ground. Differences in the amount of OTC-induced soil warming emphasize the relative importance of factors other than air temperature, such as direct sunlight, which may also change with climate change. It is unclear, therefore, whether regional climate change will result in soil warming (CitationKennedy, 1997). The daily and annual differences are important to consider when accurately characterizing the air or soil temperature profiles (CitationMyers and Pitelka, 1979). The variability in temperatures between days and years detected in this study confirms that long monitoring periods and use of multiple sensors are necessary to accurately access the potential range in near-surface temperatures for a given site (cf. CitationKlene et al., 2001).
Differences in OTC soil warming between sites may be attributable to differences in plant cover and standing water, as well as to unintended influences by the chambers. Where plant cover was nearly complete in the BW site, ground surface temperatures were, on average, higher in the OTCs. Where plant cover was lower and there was exposed ground in the BD site, ground surface temperature in the OTCs was lower at the daily maximum, probably due to attenuation of direct sunlight by the chamber walls. The AD site also had bare ground, but the lighter color and high albedo of the sandy soils may have reduced the direct impact of radiative warming. At the AW site, the presence of the OTCs may have reduced absorption of solar radiation by the shallow standing water.
The dry heath sites thawed faster and more deeply than the wet meadow sites given the same amount of heating, as shown by the value of their edaphic term at these locations. The increased depth of thaw in the dry sites relative to the wet sites is consistent with the overall pattern for the region: owing to their high thermal conductivity and the lack of latent heat effects, dry coarse-grained soils generally experience relatively deep thaw. Thaw in this study's wet sites, which are subject to periodic inundation and have thick insulating moss layers, is somewhat shallower than in its dry sites, but they are large relative to thaw depths recorded in other land cover types in the region (CitationNelson et al., 1998; CitationHinkel and Nelson, 2003). Results from this study are, therefore, consistent with those obtained in earlier work. The most likely reason why OTCs appear to have little to no effect on depth of thaw is the small size of the chambers, which has little influence on lateral heat exchange at depth. It is also possible that the taller plant canopy and increase in standing dead plant matter in the warmed plots may have also increased the thermal resistance of the vegetation layer. Although soil temperatures were changed inside the OTCs, the effect decreased rapidly with depth in a manner similar to other studies (CitationZhang and Welker, 1996; CitationMarion et al., 1997). Under larger closed-top chambers, thaw depths have been shown to increase (CitationChapin et al., 1995; CitationBret-Harte et al., 2001). At all sites the edaphic term (E) for the OTC plots was lower than that of the control plots.
Amplification of the range of daily temperatures due to a disproportionate increase in daily maximum temperature, the reduction of wind, and the potential uncoupling of soil and air temperatures in the OTCs are generally considered inconsistent with climate-change forecasts (CitationKennedy, 1995a, Citation1995b; CitationIPCC, 2001b). Experimental artifacts are not problems unique to OTC warming. Even mechanical heating devices such as soil heating cables (CitationHillier et al., 1994; CitationOberbauer et al., 1998) and infrared heaters (CitationNijs et al., 2000) have drawbacks beyond the obvious financial costs and infrastructure maintenance. Such devices can provide a specified amount of warming, but may also create non-natural vertical temperature profiles and other unwanted experimental artifacts such as soil drying (CitationHarte et al., 1995). The lack of complete experimental control and the accompanying artifacts are tradeoffs inherent to in situ field studies. As with all field studies, it is important to properly document and address any potential experimental artifacts.
The performance of a warming manipulation is of greatest interest when comparing the biological response of warming experiments. When making comparisons, it is important to recognize that the treatments themselves may have performed differently between sites and years. For example, based on the variable amount of OTC warming, one might predict that the response of plants would be least in the AW site and greatest in the BW site, and that all sites would have had the smallest response in 2000. The mechanisms contributing to observed biological response might also vary between sites. For example, because many researchers believe that below-ground processes leading to increased nutrient availability influence plant response to warming (CitationChapin, 1983; CitationWookey and Robinson, 1997; CitationShaver and Jonasson, 1999) and because the below-ground response to OTCs varied considerably between sites, the influence of below-ground processes on plant performance may vary between sites. Therefore, the interpretation of results from warming experiments is limited if the performance of the warming mechanism is not well documented.
The influence of regional warming on thaw depth is complicated by the influence of vegetation, which is also expected to change (CitationWalker et al., 2003). Quantifying the potential negative feedbacks of the thermal resistance of changing vegetation on thaw depth is an important direction for future research. It is possible that changes in thaw depth and vegetation will act on different time scales and as a result may be out of phase during rapid regional warming events. It is our hope that studies such as this, that concurrently measure thaw depth and vegetation, will be able to address this and other issues to present a holistic approach for furthering understanding how Arctic terrestrial ecosystems will respond to climate warming.
FIGURE 1. Photographs of each study site in relation to average summer temperature and soil moisture. Each site had an array of 24 experimentally warmed p1ots, with the use of open-top chambers (OTCs), and 24 control plots. One of the automated weather stations used to characterize regional climate is shown in the photograph of the Barrow Dry Heath site

FIGURE 2. Thawing degree-day totals from snowmelt until 15 August (TDDsm) measured at screen height (2 m) and canopy height (13 cm) over control and experimentally warmed plots at the four study sites during years 1994–2001. Error bars represent the maximum and minimum
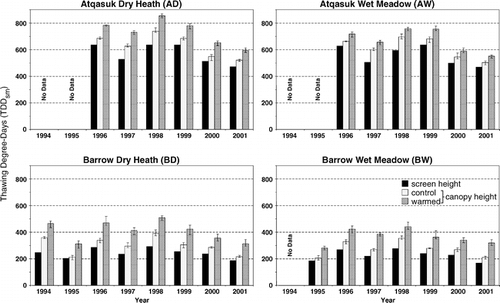
FIGURE 3. Progression of daily average temperatures recorded in July 2000 for sensors placed above ground (S = screen height; 2 m, 13 cm, 0 cm) and below ground (5, 15, 30, 45 cm) in control (C) and experimentally warmed (W) plots at the four study sites. The daily sun (top), rain (middle), and wind (bottom) conditions are displayed above the temperature graph according to the legend displayed in the figure
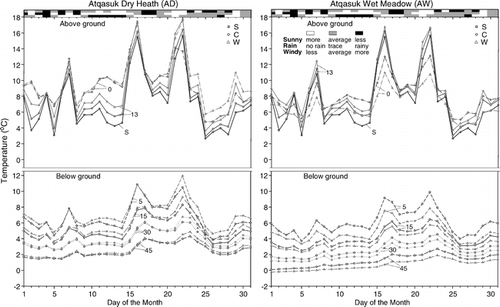
FIGURE 3. (Continued)
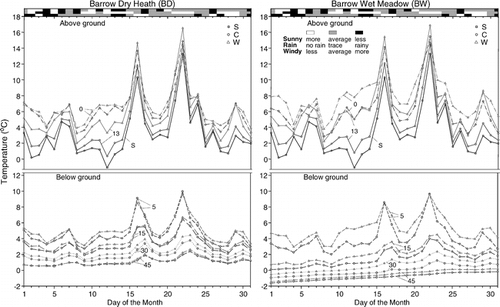
FIGURE 4. Mean daily vertical distribution of minimum (dotted dashed line), average (solid line), and maximum (dashed line) temperature in July for the years 1999–2001 at the four study sites and the change in these temperatures with height in the warmed plots relative to the control plots
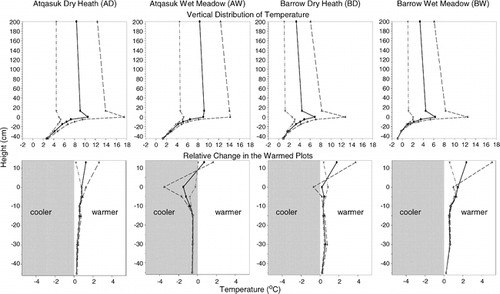
FIGURE 5. Relationship between thaw depth and thawing degree-day totals since snowmelt (TDDsm) recorded at canopy height (13 cm) at the four study sites. Thaw depths and TDDsm are the average for each treatment on a given day. The edaphic term (E) is the slope of the regression with a y-intercept of 0
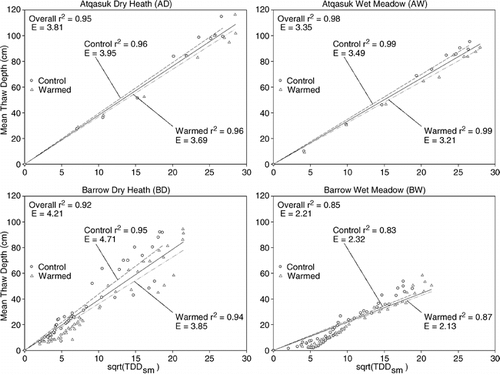
TABLE 1 Summary of the physical attributes of each site including abundant and characteristic plant species. Site abbreviations, used throughout this paper, are AD (Atqasuk Dry Heath), AW (Atqasuk Wet Meadow), BD (Barrow Dry Heath), and BW (Barrow Wet Meadow)
TABLE 2 Average calendar day of OTC installation (set-up) and snowmelt and mean daily temperatures (°C) from installation until 15 August recorded at screen height (S, 2 m) and at canopy height (13 cm) over control (C) and experimentally warmed (W) plots from years 1994–2001 at each study site. Information in italics is estimated
TABLE 3 Average depth of thaw (cm) recorded at the end of the field season at each site (AD—Atqasuk Dry Heath, AW—Atqasuk Wet Meadow, BD—Barrow Dry Heath, BW—Barrow Wet Meadow) for control (C) and experimentally warmed (W) plots during the years 1994–2001. Values in parentheses are standard deviations (n = 24)
Acknowledgments
We thank the U.S. National Science Foundation, Office of Polar Programs, Arctic System Science Program for its support of this work (OPP #9714103), and the many people responsible for collecting and organizing the data, particularly Christian Bay, Lisa Walker, and Steve Rewa. We also thank our many ITEX colleagues and two anonymous reviewers for sharing their insights and constructive criticism. Any opinions, findings, conclusion, or recommendations expressed in this paper are those of the authors and do not necessarily reflect the views of the National Science Foundation. Identification of specific products and manufacturers in the text does not imply endorsement by the National Science Foundation.
Notes
Revised ms submitted May 2005
References Cited
- Arft, A. M. , M. D. Walker , J. Gurevitch , J. M. Alatalo , M. S. Bret-Harte , M. R T. Dale , M. C. Diemer , F. Gugerli , G. H R. Henry , M. H. Jones , R. D. Hollister , I. S. Jónsdóttir , K. Laine , E. Lévesque , G. M. Marion , U. Molau , P. Mølgaard , U. Nordenhall , V. Raszhivin , C. H. Robinson , G. Starr , A. Stenström , M. Stenström , Ø Totland , P. L. Turner , L. J. Walker , P. J. Webber , J. M. Welker , and P. A. Wookey . 1999. Responses of tundra plants to experimental warming: meta-analysis of the international tundra experiment. Ecological Monographs 64:491–511.
- Bliss, L. C. , G. M. Courtin , D. L. Pattie , R. R. Riewe , D. W A. Whitfield , and P. Widden . 1973. Arctic tundra ecosystems. Annual Review of Ecology and Systematics 4:359–399.
- Bret-Harte, M. S. , G. R. Shaver , J. P. Zoerner , J. Johnstone , J. L. Wagner , A. S. Chavez , R. F. Gunkelman IV , S. C. Lippert , and J. A. Laundre . 2001. Developmental plasticity allows Betula nana to dominate tundra subjected to an altered environment. Ecology 82:18–32.
- Brown, J. , K. M. Hinkel , and F. E. Nelson . 2000. The circumpolar active layer monitoring (CALM) program: Research designs and initial results. Polar Geography 24:165–258.
- Chapin III, F. S. 1983. Direct and indirect effects of temperature on arctic plants. Polar Biology 2:47–52.
- Chapin III, F. S. and G. R. Shaver . 1996. Physiological and growth responses of arctic plants to a field experiment simulating climatic change. Ecology 77:822–840.
- Chapin III, F. S. , G. R. Shaver , A. E. Giblin , K. J. Nadelhoffer , and J. A. Laundre . 1995. Response of arctic tundra to experimental and observed changes in climate. Ecology 76:696–711.
- Corbet, P. S. 1972. The microclimate of arctic plants and animals, on land and in fresh water. Acta Arctica 18:7–43.
- Coulson, S. , I. D. Hodkinson , A. Strathdee , J. S. Bale , W. Block , M. R. Worland , and N. R. Webb . 1993. Simulated climate change: the interaction between vegetation type and microhabitat temperatures at Ny Ålesund, Svalbard. Polar Biology 13:67–70.
- Debevec, E. M. and S. F. MacLean Jr. . 1993. Design of greenhouses for the manipulation of temperature in tundra plant communities. Arctic and Alpine Research 25:56–62.
- Everett, J. T. and B. Fitzharris . 1998. The Arctic and the Antarctic. In Watson, R. T., Zinyowera, M. C., and Moss, R. H. (eds.), The Regional Impacts of Climate Change An Assessment of Vulnerability. A Special Report of IPCC Working Group II. Cambridge: Cambridge University Press, 87–103.
- Harte, J. , M. S. Torn , F. R. Chang , B. Feifarek , A. P. Kinzig , R. Shaw , and K. Shen . 1995. Global warming and soil microclimate: Results from a meadow-warming experiment. Ecological Applications 5:132–150.
- Henry, G. H R. and U. Molau . 1997. Tundra plants and climate change: The International Tundra Experiment (ITEX)—Introduction. Global Change Biology 3:1–9.
- Hillier, S. H. , F. Sutton , and J. P. Grime . 1994. A new technique for the experimental manipulation of temperature in plant-communities. Functional Ecology 8:755–762.
- Hinkel, K. M. and F. E. Nelson . 2003. Spatial and temporal patterns of active layer thickness at circumpolar active layer monitoring (CALM) sites in northern Alaska, 1995–2000. Journal of Geophysical Research–Atmospheres, 108: article 8168.
- Hinkel, K. M. and J. R J. Nicholas . 1995. Active layer thaw rate at a boreal forest site in central Alaska, U.S.A. Arctic and Alpine Research 27:72–80.
- Hollister, R. D. 2003. Response of tundra vegetation to temperature: implications for forecasting vegetation change. Ph.D. thesis. Michigan State University, East Lansing, Michigan, U.S.A., 385 + xxiv pp.
- Hollister, R. D. and P. J. Webber . 2000. Biotic validation of small open-top chambers in a tundra ecosystem. Global Change Biology 6:835–842.
- Hollister, R. D. , P. J. Webber , and C. E. Tweedie . 2005a. The response of Alaskan arctic tundra to experimental warming: differences between short- and long-term responses. Global Change Biology 11:525–536.
- Hollister, R. D. , P. J. Webber , and C. Bay . 2005b. Plant response to temperature in northern Alaska: implications for predicting vegetation change. Ecology 86:1562–1570.
- IPCC, 2001a. Climate Change 2001: The Scientific Basis. Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. 881 pp.
- IPCC, 2001b. Climate Change 2001: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. 1032 pp.
- Johnson, D. A. and L. L. Tieszen . 1976. Above-ground biomass allocation, leaf growth and photosynthesis patterns in tundra plant forms in Arctic Alaska. Oecologia 224:159–173.
- Jumikis, A. R. 1977. Thermal Geotechnics. New Brunswick: Rutgers University Press, 375 pp.
- Kennedy, A. D. 1995a. Simulated climate change: are passive greenhouses a valid microcosm for testing the biological effects of environmental perturbations?. Global Change Biology 1:29–42.
- Kennedy, A. D. 1995b. Temperature effects of passive greenhouse apparatus in high-latitude climate-change experiments. Functional Ecology 9:340–350.
- Kennedy, A. D. 1997. Bridging the gap between general circulation model (GCM) output and biological microenvironments. International Journal of Biometeorology 40:119–122.
- Klene, A. E. , F. E. Nelson , N. I. Shiklomanov , and K. M. Hinkel . 2001. The n-factor in natural landscapes: variability of air and soil-surface temperatures, Kuparuk River Basin, Alaska, U.S.A. Arctic, Antarctic, and Alpine Research 33:140–148.
- Long, S. P. and F. I. Woodward . 1988. Plants and Temperature. Cambridge: The Company of Biologists Limited, 415 + x pp.
- Marion, G. M. , G. H R. Henry , D. W. Freckman , J. Johnstone , G. Jones , M. H. Jones , E. Lévesque , U. Molau , P. Mølgaard , A. N. Parsons , J. Svoboda , and R. A. Virginia . 1997. Open-top designs for manipulating field temperature in high-latitude ecosystems. Global Change Biology 3:20–32.
- Minorsky, P. V. 2002. Global warming—Effects on plants. Plant Physiology 129:1421–1422.
- Myers, J. P. and F. A. Pitelka . 1979. Variations in summer temperature patterns near Barrow, Alaska: Analysis and ecological interpretation. Arctic and Alpine Research 11:131–144.
- Nelson, F. E. and S. I. Outcalt . 1987. A computational method for prediction and regionalization of permafrost. Arctic and Alpine Research 19:279–288.
- Nelson, F. E. , N. I. Shiklomanov , G. R. Mueller , K. M. Hinkel , D. A. Walker , and J. G. Bockheim . 1997. Estimating active-layer thickness over a large region: Kuparuk River Basin, Alaska, U.S.A. Arctic and Alpine Research 29:367–378.
- Nelson, F. E. , K. M. Hinkel , N. I. Shiklomanov , G. R. Mueller , L. L. Miller , and D. A. Walker . 1998. Active-layer thickness in north-central Alaska: systematic sampling, scale, and spatial autocorrelation. Journal of Geophysical Research 103:28963–28973.
- Nijs, I. , F. Kockelbergh , M. Heuer , L. Beyens , K. Trappeniers , and I. Impens . 2000. Climate-warming simulation in tundra: enhanced precision and repeatability with an improved infrared-heating device. Arctic, Antarctic, and Alpine Research 32:346–350.
- Oberbauer, S. F. , G. Starr , and E. W. Pop . 1998. Effects of extended growing season and soil warming on carbon dioxide and methane exchange of tussock tundra in Alaska. Journal of Geophysical Research–Atmospheres 103:29075–29082.
- Root, T. L. , J. T. Price , K. R. Hall , S. H. Schneider , C. Rosenzweig , and J. A. Pounds . 2003. Fingerprints of global warming on wild animals and plants. Nature 421:57–60.
- Rustad, L. E. , J. L. Campbell , G. M. Marion , R. J. Norby , M. J. Mitchell , A. E. Hartley , J. H C. Cornelissen , and J. Gurevitch . 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126:543–562.
- SAS, 2000. SAS Version 8.1. Cary: SAS Institute Incorporated.
- Shaver, G. R. and S. Jonasson . 1999. Response of arctic ecosystems to climate change: Results of long-term field experiments in Sweden and Alaska. Polar Research 18:245–252.
- Shaver, G. R. , J. G. Canadell , F. S. Chapin III , J. Gurevitch , J. Harte , G. Henry , P. Ineson , S. Jonasson , J. M. Melillo , L. F. Pitelka , and L. E. Rustad . 2000. Global warming and terrestrial ecosystems: A conceptual framework for analysis. BioScience 50:871–882.
- Shiklomanov, N. I. and F. E. Nelson . 2002. Active-layer mapping at regional scales: a 13-year spatial time series for the Kuparuk region, north-central Alaska. Permafrost and Periglacial Processes 13:219–230.
- Stone, R. S. , T. Mefford , E. Dutton , D. Longenecker , B. Halter , and D. Endres . 1996. Barrow, Alaska, Surface Radiation and Meteorological Measurements: January 1992 to December 1994. Boulder: U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Environmental Research Laboratories, 79 pp.
- Tieszen, L. L. 1972. The seasonal course of above ground production and chlorophyll distribution in wet arctic tundra at Barrow, Alaska. Arctic and Alpine Research 4:307–324.
- Walker, D. A. , G. J. Jia , H. E. Epstein , M. K. Raynolds , F. S. Chapin III , C. Copass , L. D. Hinzman , J. A. Knudson , H. A. Maier , G. J. Michaelson , F. Nelson , C. L. Ping , V. E. Romanovsky , and N. Shiklomanov . 2003. Vegetation-soil-thaw-depth relationships along a Low-Arctic bioclimate gradient, Alaska: synthesis of information from the ATLAS studies. Permafrost and Periglacial Processes 14:103–123.
- Walker, M. D. , C. H. Wahren , R. D. Hollister , G. H R. Henry , L. E. Ahlquist , J. M. Alatalo , M. S. Bret-Harte , M. P. Calef , T. V. Callaghan , A. B. Carroll , H. E. Epstein , I. S. Jónsdóttir , J. A. Klein , B. Magnsson , U. Molau , S. F. Oberbauer , S. P. Rewa , C. H. Robinson , G. R. Shaver , K. N. Suding , C. C. Thompson , A. Tolvanen , Ø Totland , P. L. Turner , C. E. Tweedie , P. J. Webber , and A. Wookey P . 2006. Plant community responses to experimental warming across the Tundra Biome. Proceedings of the National Academy of Science of the United States of America (PNAS) 103:5 1342–1346.
- Weller, G. and B. Holmgren . 1974. The microclimate of the arctic tundra. Journal of Applied Meteorology 13:854–862.
- Wookey, P. A. and C. H. Robinson . 1997. Interpreting environmental manipulation experiments in arctic ecosystems: Are ‘disturbance’ responses properly accounted for?. In Crawford, R. M. M. (ed.), Disturbance and Recovery in Arctic Lands. Boston: Kluwer Academic Publishers, 115–134.
- Wookey, P. A. , A. N. Parsons , J. M. Welker , J. A. Potter , T. V. Callaghan , J. A. Lee , and M. C. Press . 1993. Comparative responses of phenology and reproductive development to simulated environmental change in sub-arctic and high arctic plants. Oikos 67:490–502.
- Zhang, Y. Q. and J. M. Welker . 1996. Tibetan alpine tundra responses to simulated changes in climate: above ground biomass and community responses. Arctic and Alpine Research 28:203–209.