Abstract
On Axel Heiberg Island, Nunavut, Canada, a banded vegetation pattern occurred on a hillside where patterned ground and unidirectional abiotic fluxes, such as downslope water flow or wind, were not present. The parent material was the obvious source of the plant pattern, as the soils occurred on five distinct types of alluvial deposits. To examine the observed pattern, plants were inventoried and soils were sampled in July 1999. Twelve vascular species of plants, but no non-vascular species, were present at the site. Neither water, often thought to limit plant distribution in the High Arctic, nor any of the other measured soil variables, predicted plant abundance. The best predictor of plant abundance, based on regression tree analysis, was total soil nitrogen; however, higher plant density was associated with lower nitrogen. The five soil types differed in plant density and soil properties. Even though the sand soil always had soil nutrients equal to or lower than the blocky clay soil, the sand and clay soils had the highest plant density and the blocky clay soil the lowest. Although the vegetation pattern is obvious, the underlying mechanism creating the pattern is not.
Introduction
The Queen Elizabeth Islands represent the northernmost land in Canada. This group of High Arctic islands covers more than 400,000 km2, but only approximately 175 vascular plant species are found there (CitationPorsild and Cody, 1980). Although the geographic distribution of these species has been documented, the ecological and physiological tolerances controlling species distributions are less well understood. Many factors combine to affect plant function at these latitudes: a short growing season, cold air and soil temperatures, high wind speeds, permafrost, soil churning, low sun angles, low precipitation, and low nutrient concentrations (CitationSavile, 1972). In fact, this entire region has been described as a marginal environment for plant growth (CitationSvoboda and Henry, 1987). The poorly developed soils are typically little modified by vegetation; thus, surficial geology plays a large role in determining where plants in general and particular species can grow (CitationEdlund, 1994).
In the High Arctic the environmental variables controlling plant distribution and abundance depend on scale. Regional-scale patterns of plant diversity in the High Arctic are correlated with summer temperature, while at the local scale, soil moisture seems to drive the distribution of plants (CitationEdlund and Alt, 1989; CitationBatten and Svoboda, 1994). Soil moisture in the High Arctic is usually related to topographic position and microtopographic features, such as patterned ground. Cryptogamic crust also tends to increase soil moisture. Because of the higher soil moisture, the protected sites for germination, and a reduction in soil cryoturbation, greater cryptogamic cover tends to increase vascular plant diversity and abundance (CitationAnderson and Bliss, 1998; CitationGold and Bliss, 1995; CitationBliss et al., 1994). Although water seems to control much of the structure and function of High Arctic plant communities, plants do not appear to be water stressed even in the driest soils (CitationGold and Bliss, 1995).
Vegetation patterns at many spatial scales are often controlled by a variety of differences in soil properties including parent material and topography (CitationJenny, 1994). In the High Arctic, like most regions, certain plant species tend to be grow on soils derived from calcareous bedrock (CitationPorsild and Cody, 1980). The chemical composition in layers of the Mancos Shale in the western United States creates predictable vegetation patterns (CitationPotter et al., 1985). In arid regions, coarser textured soils support higher plant densities because these soils actually provide more water for plants (CitationNoy-Meir, 1973). Different plant communities and soils develop based on topographic position largely driven by differences in soil moisture (CitationBillings, 1973). Although many soil factors affect plant distribution, in the High Arctic, most research has focused on the effects of patterned ground (CitationAnderson and Bliss, 1998).
One vegetation pattern, banded vegetation, is defined as vegetation growing in a linear pattern, often with little plant cover between bands. Three mechanisms have been elucidated to create this pattern in different settings: a unidirectional flow of water downslope, stress from a unidirectional wind, and linear microtopography resulting from freeze-thaw cycles. Banded vegetation, or “tiger stripes,” typically occurs on shallow slopes in arid regions of North America, Australia, and Africa and results from a positive feedback: established plants intercept water and nutrients flowing down the shallow slopes facilitating the establishment of additional plants across the slope (CitationWhite, 1971; CitationAguiar and Sala, 1999). In temperate regions the vegetation may develop a banded pattern when plants are killed by exposure to unidirectional winds. Salt spray, in coastal regions, can kill exposed trees and creates a wave-like regeneration pattern (CitationCampbell, 1998). Similar wave patterns develop in forests in New England and Argentina exposed to high winds from a consistent direction (CitationFoster, 1988; CitationPuigdefabregas et al., 1999). Patterned ground in cold regions can form as stripes oriented down the slope creating non-random vegetation patterns associated with the microtopographic variation (CitationWashburn, 1956; CitationAnderson and Bliss, 1998). However, few banded vegetation patterns have been described in herbaceous communities, and banded vegetation patterns unrelated to patterned ground are undescribed in the arctic literature.
This study documents the contour-parallel banded vegetation pattern in a cold steppe plant community on Axel Heiberg Island in the High Arctic and seeks to determine its potential causes. Microscale vegetation patterns in high arctic plant communities are poorly understood, and banded vegetation patterns unrelated to patterned ground are undescribed. Unlike banded vegetation pattern worldwide, there was neither patterned ground present nor evidence of either unidirectional transport of water or death of plants caused by wind. The obvious origin of the vegetation pattern was the parent material consisting of layered sedimentary deposits. More specifically, this study was designed to answer the following questions:
What is the microscale spatial distribution of plants?
Is this vegetation pattern related to the spatial distribution of soil type/sedimentary deposits? That is, is the plant distribution correlated with soil moisture or other soil variables that vary among the soil types?
Methods
STUDY AREA
This study was conducted at the fossil forest site east of the Geodetic Hills on Axel Heiberg Island, Nunavut, Canada (79°55′N, 89°02′W). Described in detail by CitationChristie and McMillen (1991), the fossil forest site is a hill 1.3 km long and 0.5 km wide, with an elevation of 200–300 m, and is composed of flat-lying, unconsolidated, Eocene sediments with repeated beds of sand, silt, and lignite (). Erosion by wind and mass wasting has been measured at 0.3 cm yr−1 in flat areas and up to 11 cm yr−1 in steeper areas (CitationBigras et al., 1995). Patterned ground was absent in the study area. Rills often develop on the lower slopes, usually beginning in sand beds.
The climate is characterized by low temperatures and scant precipitation. The mean annual temperature at Eureka, 60 km east, is −19°C, and the July–August mean is 4°C. The growing season lasts from approximately 15 June to 25 August (CitationMaxwell, 1981). Precipitation averages 68 mm yr−1, with 22 mm during the growing season, and approximately one-third falling as rain. The weather during 1999, when the data were collected, was particularly warm and dry with a July/August mean temperature of 5.9°C and only 5.8 mm of precipitation (Canadian National Climate Data and Information Archive; http://www.climate.weatheroffice.ec.gc.ca).
VEGETATION AND SOIL SAMPLING
Plants and soils were sampled during the last week of July 1999. Three transects were established down the southeast-facing slope of the site perpendicular to the distinct vegetation bands running across the slope. The transects were 55 m long, approximately 150 m apart, and the slope angle averaged 15°. All plants within one meter of each transect were identified and counted, with each ramet of the clonal species counted as an individual. Plants of non-clonal species were classified as seedlings if neither flowers nor seeds were present or remaining from previous years. Most of these seedlings were small, but larger plants without reproductive structures and even the small plants may in fact be quite old and not truly be seedlings. Each meter of transect (representing an area 2 m2) will be referred to as a plot for a total of 168 plots in the plant data set.
To determine the association between plant cover and soil characteristics, five soil types were described based on the characteristics of the Eocene deposits: blocky clay, clay, sand, white, and organic. The soil types occurred several times along the transects but were not always laterally continuous between transects. All soils were covered in 1–3 cm of homogenous gravel-sized slopewash, transported downhill by contemporary mass-wasting processes. The five soil types were easily distinguishable visually and were separated by abrupt boundaries. The blocky clay soil was composed of large stable aggregates, while the clay soil was sticky and lacked aggregates. Preliminary data indicated some mineralogical differences between these two clay soils, but they both were both gray with a silt loam to silty clay loam texture once the aggregates of the blocky clay soil were crushed (D. Liptzin, unpublished data). The sand soils had a sandy loam texture and occasionally contained a band of lithified and oxidized sandstone. White soils, corresponding to the white layers described by CitationTarnocai et al. (1991), were composed of platy quartz of unknown origin. Organic soils corresponded to the Eocene lignitic beds and contain only small amounts of mineral soil. All soil types ranged in pH from 5 to 6, and were classified as Regosolic Static Cryosols (CitationSoil Classification Working Group, 1998).
A hillslope profile was created using slope angles and changes in soil type (). Soils were sampled every 2 m along the transects except in areas with many consecutive plots with blocky clay soils, which were sampled approximately every 5 m. A soil pit was excavated to 50 cm at each sampling location underneath the plant nearest the transect. Pedogenic horizons were never visible, but changes in soil type were noted. The depth of the deepest visible roots was measured. Active layer depth within the soil pit was determined by the presence of ice.
The soil data set was based on seven variables measured on soils from a subset of the plots in the plant data set (n = 49). The top 8 cm of soil was sampled by forcing a 1700 cm3 aluminum box into the soil. Samples were kept in plastic bags until reaching the University of Pennsylvania. The samples were air dried at room temperature to a constant weight. Soils were sieved (2 mm), and the modern litter and roots and Eocene organic matter remaining in the sieve were removed. Organic matter was not removed from the organic soil samples because they were considered organic soils. Gravimetric water content was determined as (wet weight − dry weight)/dry weight. Aggregates > 2 mm were weighed separately to calculate the percentage of the soil in stable aggregates and then remixed with the dry, sieved soil. Organic carbon and total nitrogen concentrations were measured on ground samples using a Carlo Erba elemental analyzer (Fisons Instruments NA1500, Dearborn, MI, U.S.A.) on samples ground in a ball mill. Exchangeable K, Mg, and Ca were extracted from the soil with 1M NH4Cl for 12 h and measured by Inductively Coupled Plasma Atomic Emission Spectrometry (Perkin-Elmer Plasma 400, Shelton, CT, U.S.A.).
DATA ANALYSES
The spatial distribution of the vegetation was described by plotting plant density against distance along each transect. To test if the plants were randomly distributed, the frequencies of plant density were compared against a Poisson distribution. The expected and observed values were combined into 10 bins with the stipulation that the expected value was never less than five.
The plots in the plant data set were ordinated using plant density for plant species with principal components analysis using PC-ORD 4.0. However, since PCA cannot handle plots with no species present, only 129 of the 168 plots could be used in the analysis. In addition, Draba and Melandrium were left out of the analysis since so few individuals were present. A canonical correspondence analysis was done in PC-ORD using the 43 plots in both the plant and the soil data set with a plant density > 0. The rows and the columns were standardized by centering and normalizing for the ordination, and a Monte Carlo test was performed to test for significance of the axes.
The plant and soil data sets were used to test for differences among the five soil types. Differences among soil types based on the variables measured in the soil data set (water, aggregates, total soil N and C, and exchangeable Ca, Mg, and K) were tested with a MANOVA followed by univariate ANOVA models on each of the soil variables using SAS (release 8.02). Two of these variables, exchangeable K and total N, were log transformed to improve normality. Means were calculated for all five soil types, and a post hoc Tukey's test was used to assess differences only among the three common soil types (blocky clay, clay, and sand). The plant data set was analyzed using ANOVA models and post hoc Tukey's comparisons to examine the main effect of soil type on plant density and species density.
Regression models were used to analyze the relationship between soil characteristics and plant density. All seven measured soil variables were tested separately with simple regressions. A series of multiple regression models were built based on the variables predicted to be most important for High Arctic ecosystems. The models were built and tested by adding, in order, water, nitrogen, aggregates, and then the exchangeable cations.
The non-parametric technique, regression tree analysis, was employed to determine which of the soil properties best predicted the plant density using S-Plus 2000 software. This technique partitions the data set (soil data set, n = 49) into two subsets based on the predictor variable that minimizes the variance. A full tree was created with a minimum of three plots in each terminal node. The full tree was pruned to five branches as this was the point where the decrease in variance per parameter added began to decline.
Results
VEGETATION DISTRIBUTION
The distribution of plants along the transects clearly shows the banded pattern (). The mean plant density on all the plots was 28 plants m−2, and a Chi-square test revealed that the plants were non-randomly distributed (df = 9, χ2 = 1496, P < 0.001). The vegetation bands were largely a consequence of the high abundance of two clonal species: Alopecurus alpinus and Stellaria crassipes. However, other species also occurred in the bands, and these two species occurred outside of the bands. The proportion of non-clonal plants classified as seedlings was significantly higher outside of the bands (F 1,101 = 5.91, P < 0.05). Every band, defined as plant density > 15 plants · m−2, was associated, at least in part, with some soil type other than the blocky clay soil (). Often there were no plants in the blocky clay soil, for up to 8 consecutive meters along the transect. The sand soil always had some plant cover.
The vegetation bands corresponded to the sedimentary deposits, with fewer plants occurring in the blocky clay soils. Even though the blocky clay soils supported lower plant densities, they had similar or higher concentrations of moisture and nutrients compared to the other soils, suggesting a physical, not a chemical cause. There were some instances where the blocky clay soil supported high densities of plants, but there was almost always another soil type less than 0.5 m below the soil surface (). Even though neither water nor nutrients were higher in the blocky clay soil less than 0.5 m above non-blocky clay soils, there were significantly more plants in these soils compared to other blocky clay soils (F 1,25 = 17.90, P < 0.0005).
Twelve species of herbaceous perennial vascular plants occurred in the study area (). Notably, non-vascular species were absent from the transects. Some species were abundant and found on all soil types (Alopecurus alpinus, Cerastium alpinum, Erysimum pallasii, Papaver radicatum, Puccinellia angustata, and Stellaria crassipes). Of these ubiquitous species only E. pallasii occurred more frequently on a relative basis on blocky clay soil than on sand soil (). Others were rare regardless of soil type (Draba sp., Luzula confusa, and Melandrium sp.). Three species were limited almost exclusively to the sand soil, but were often abundant there (Arnica alpina, Oxyria dignya, and Taraxacum pumilum). The first PCA axis of the plant species composition splits the sand soil, and the three species that grow only there, from the other soil types and species (). The second PCA axis separates the four other soil types. The Monte Carlo test indicated that the axes of the CCA were not significant, and thus the results will not be discussed.
EFFECTS OF SOIL TYPES
Neither soil moisture nor nitrogen, often thought to be limiting to High Arctic plants, could predict plant density. Neither could any other soil variable individually or any of the tested multiple regression models. However, species density was negatively related to the percentage of aggregates (F 1,41 = 10.91, P < 0.005). The regression tree analysis provided a way to determine which soil variable best explained the difference in plant density. The pruned tree revealed that nitrogen was best predictor such that plots with a total soil nitrogen less than 0.0985% tended to have more plants (). The low nitrogen node was further subdivided such that the highest plant density occurred in plots with fewer aggregates and higher calcium and the lowest plant density occurred in plots with more aggregates and higher carbon.
The MANOVA analysis indicated significant differences among the soil types (Pillai's trace = 1.71, df = 28,164, F = 4.39, P < 0.001). The univariate ANOVA models revealed that the three common soil types differed significantly in moisture, calcium, magnesium, nitrogen, plant density, species density, and aggregates but not in potassium or carbon. In general, organic soil had the highest values for soil nutrients, sand soil and white soil had the lowest values, and clay soil and blocky clay soil were intermediate (). Clay soil had significantly more moisture than sand and blocky clay soils. Blocky clay soil had significantly more aggregates than both sand and clay soils. Sand soil had lower concentrations, though not always significantly lower, of all measured nutrients and water and the fewest aggregates. However, sand soil had significantly higher species density than blocky clay and clay soils and higher plant density than blocky clay soil. Clay soil also had higher species density and plant density than blocky clay soil.
Only 5 out of 55 soil pits had an active layer less than 50 cm deep, but these soils always had at least some plants in them. The maximum rooting depth tended to increase with the number of plants. However, it is not known whether this relationship is due to soil limitations on root growth or whether it is simply an artifact because with more plants there is a greater chance that one will root more deeply.
Discussion
The obvious banded vegetation pattern at the fossil forest site is intriguing because the cause of the pattern is not obvious. Although the plant banding mirrored the banding of the underlying geology, this plant pattern did not exist around the entire fossil forest hill, nor on other hills nearby with similar sedimentary deposits. Unfortunately, none of the sites without banding were sampled to examine what might be different about those sites. In general, there was low plant density on the entire fossil forest hill except for the vegetation bands within the study area. There is no literature on micro-scale vegetation patterns in the High Arctic where the variation in plant species and abundance was not due to cryogenic processes, micro-topographic relief, or cryptogram abundance. None of the measured soil variables appears to explain the plant pattern. The regression tree analysis revealed that total soil nitrogen could best separate plots into low and high plant densities. However, a simple regression did not find a significant effect of nitrogen on plant density, perhaps because of the small number of samples collected from such different soil types.
Water is often thought to control plant distribution in the High Arctic, but the effects of soil moisture on plant function in the High Arctic are poorly understood. CitationGold and Bliss (1995) found that plant abundance was higher in wetter soils even though plants were not water stressed in the drier soils. In the present study there was a non-significant but negative relationship between soil moisture and plant density both within and among soil types. The data in the present study were collected near the end of the growing season of a warm and dry year; thus, these soils should never become much drier nor the active layer much deeper. Since plants grew even where the active layer was only 30 cm deep, depth to active layer should not determine plant distribution in colder years. It is possible that differences in water would be more evident early in the growing season, or that it is the timing of the permafrost melting that is important. Slope and aspect affect the pattern of snow distribution in winter as well as summer moisture, but the entire study area had a similar slope and aspect (CitationWoo and Young, 1997).
Soil texture may complicate the interpretation of the results. Although both blocky clay and clay soils have a higher percentage of water than the sand soil, this does not necessarily mean more plant-available water. In fact, in some arid regions, coarser textured soils support higher plant densities because these soils actually provide more water for plants (CitationNoy-Meir, 1973). However, this does not seem to be the case here, as the clay and the sand soils support similar numbers of plants. Although the clay and the blocky clay soils had similar textures when the aggregates were disintegrated, the soil aggregates may not provide enough surface area for roots or they may affect how the soil freezes. The ecological tolerance of most High Arctic species are thought to be wide, but it is not clear why some species grew on all soil types while others grew only on the sand soil (CitationMuc et al., 1989).
There is likely variation in soil temperature at this site because of the large differences in soil moisture. However, if moisture is the major control of soil temperature, then the blocky clay soil with intermediate moisture should also have intermediate temperatures. The slope angle may also affect the microsite temperature as it varies from 6–22°C along the transects. However, the bands of plants occur on both the shallow slopes and the steep slopes. Thus, soil temperature is unlikely to be the cause of the overall vegetation pattern. Grasses, sedges/rushes, and non-graminoids in the High Arctic seem to differ in their soil temperature responses (CitationBrooker and van der Wal, 2003). However, with so few species present, it is difficult to compare functional group response to temperature. In addition, only non-graminoid species at this site showed any preference for soil type, and most species occurred regardless of soil moisture.
The vegetation pattern is difficult to explain from patterns of either asexual reproduction or seed germination. Since the main band-forming species were both clonal, they should spread in any direction equally from where they first establish if there were no variation in soil properties. Seedling establishment may be limited in the High Arctic in general, but all five soil types were covered by the same slopewash material. Thus, the soil type should not affect the earliest stages of seedling establishment. Similar to the clonal plants, plants from seeds should develop in bands if protection by established plants is essential. In a polar desert community on Ellesmere Island, high seedling densities were associated with other plants or desiccation cracks (CitationElberling, 2000). However, at this site there were no desiccation cracks and the seedlings were actually more numerous between the bands.
Banded vegetation can usually be ascribed to one of several mechanisms. In cold regions banding has always been associated with patterned ground, which did not occur at this site, nor was there any evidence of cryoturbation. Vegetation bands in most other regions are caused by the directional flow of water or wind. No data are available about the wind directions at this site, but wind-caused death of plants seems unlikely. Rainfall is so limited and the slopewash so coarse that overland flow is extremely unlikely. Overland flow during snowmelt is a possibility, but none was observed from late June until snowmelt was completed in 1999. Water may flow in the soils along the boundary between soil types because of large texture differences, but this cannot be the explanation for the higher number of plants when blocky clay is near another soil type because there were no measurable differences in soil properties (). Distinct vegetation patterns have been described on the Mancos Shale in the western United States because of variations in the bedrock composition (CitationPotter et al., 1985). However, at the fossil forest site the soils are derived from fluvial sediments, which should mix thoroughly material from a variety of sources.
Plant community composition and structure in the arctic can usually be predicted with environmental variables such as temperature, moisture, or pH, at various spatial scales (CitationGould and Walker, 1999; CitationEdlund and Alt, 1989). However, at the fossil forest site this was not possible. Banded vegetation can be caused by several mechanisms, but none seem likely causes of the pattern, either. Further research could help elucidate whether the parent material, topographic position, aspect, hydrology or some unique combination of these factors explains the banded vegetation.
FIGURE 1. A view of the fossil forest hill. The flat-lying sedimentary deposits are clearly visible, but the bands of plants running parallel to these deposits are more difficult to see because of the low plant density. The black box illustrates the orientation of the transects

FIGURE 2. Hillslope profile for transect 1. The elevations of the soil types other than blocky clay are listed at right with sand soil at 227.4–228.8 m, clay soil at 231.8–232.6 m, organic soil at 235.4–235.9 m, white soil at 238.6–239.2 m, organic soil at 239.8–240.5 m, and clay soil at 240.5–240.8 m. Blocky clay soil occurs everywhere else and is depicted with the hatched shading. The slopewash is shown with the dotted shading; its depth is exaggerated vertically approximately 30 times. The vertical line demonstrates how the depth to non-blocky clay soils was calculated
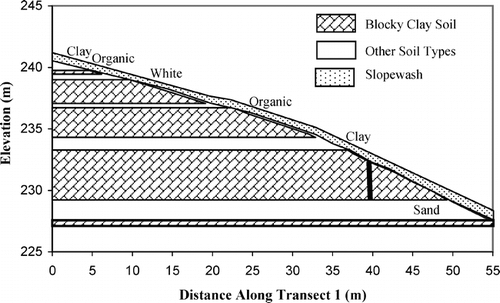
FIGURE 3. Plant density along the three transects. Zero is the uphill end of the transect. Large open symbols represent the four non-blocky clay soil types
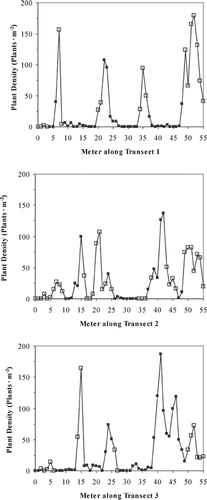
FIGURE 4. Relationship between plant density for all blocky clay soils and the calculated depth to a non-blocky clay soil
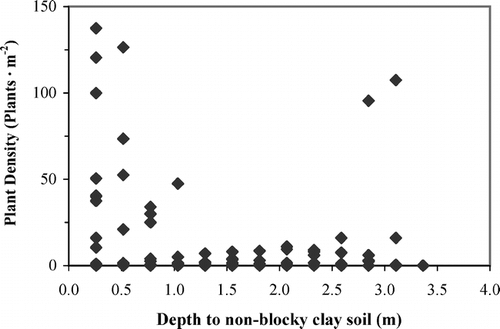
FIGURE 5. PCA ordination of the plots containing at least one plant. PCA axis 1 explained 24% and PCA axis 2 explained 17% of the variance in species composition. Means for ten species of plants (open squares) and all five soil types (crosses) are plotted. Species codes are the first two letters of the genus
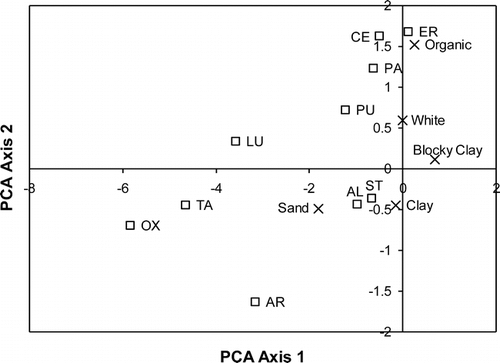
FIGURE 6. Pruned regression tree of the soil data set (n = 49). A minimum of three plots occur in each terminal node. The number of branches of the tree was determined by examining when the decrease in variance per node added to the tree began to decline. The variable and the value determining the binary separation are displayed at each node. If the rule is true, the right branch is followed; if false, the left branch. Numbers in boxes at the terminal nodes are the expected value, which is the mean value, of plant density for the group of plots meeting the criteria leading to the node

TABLE 1 Effect of soil type on absolute plant abundance. Values are the mean plant density (plants · m−2) for each species on each soil type. Total is the absolute number of plants of each species, and mean is the average plant density or species density for all soil types combined. Numbers in parentheses after soil type are the number of plots for each soil type. Means and standard errors were calculated for the total plant density and species density for each soil type
TABLE 2 Effect of soil type on relative plant abundance. Values for species are the percentage of plots of each soil type in which a species occurred. Values for plant density are the percentage of plots of each soil type in which plant density was zero, low, medium, or high. % is the percentage of plots of each soil type
TABLE 3 Soil chemical and physical properties of the five soil types. Means followed by different superscript letters are significantly different (Tukey's test p < 0.05). Only blocky clay, clay, and sand soils were compared
Acknowledgments
C. J. Williams, P. Semonche, D. R. Vann, J. B. Smith, J. R. Smith, B. A. Lepage, and A. H. Johnson provided assistance in experimental design, data collection, interpretation of the geologic setting, and moral support. Polar Continental Shelf Project and the Andrew W. Mellon Foundation provided financial and logistical support. M. Dranoff assisted with the soil analyses. W. D. Bowman, T. R. Seastedt, T. A. Forbis, K. N. Suding, K. D. LeJeune, H. E. Reed, W. A. Gould, C. C. Cleveland, and C. Rumbaitis-del Rio provided helpful comments on the manuscript. Two reviewers, R. M. M. Crawford and I. D. Hodkinson, also greatly improved this manuscript, especially the clarity and conciseness.
Notes
Revised ms submitted August 2005
References Cited
- Aguiar, M. and O. E. Sala . 1999. Patch structure, dynamics, and implications for the functioning of arid ecosystems. Trends in Ecology and Evolution 14:273–277.
- Anderson, D. G. and L. C. Bliss . 1998. Association of plant distribution patterns and microenvironments on patterned ground in a polar desert, Devon Island, N.W.T., Canada. Arctic and Alpine Research 30:97–107.
- Batten, D. S. and J. Svoboda . 1994. Plant communities on the uplands in the vicinity of the Alexandra Fiord lowland. In Svoboda, J., and Freedman, B. (eds.), Ecology of a polar oasis Alexandra Fiord, Ellesmere Island, Canada. Toronto: Captus University Publications, 97–110.
- Bigras, C. , M. Bilz , D. W. Grattan , and C. Gruchy . 1995. Erosion of the Geodetic Hills Fossil Forest, Axel Heiberg Island, Northwest Territories. Arctic 48:342–353.
- Billings, W. D. 1973. Arctic and alpine vegetations: similarities, differences, and susceptibility to disturbance. BioScience 23:697–704.
- Bliss, L. C. , G. H R. Henry , J. Svoboda , and D. I. Bliss . 1994. Patterns of plant distribution within two polar desert landscapes. Arctic and Alpine Research 26:46–55.
- Brooker, R. and R. van der Wal . 2003. Can soil temperature direct the composition of High Arctic plant communities?. Journal of Vegetation Science 14:535–542.
- Campbell, D. J. 1998. Salt-wind induced wave regeneration in coastal pine forests in New Zealand. Canadian Journal of Forest Research 28:953–960.
- Christie, R. L. and N. J. McMillan . 1991. Introduction. In Christie, R. L., and McMillan, N. J. (eds.), Tertiary fossil forest of the Geodetic Hills, Axel Heiberg Island, Arctic Archipelago. Geological Survey of Canada Bulletin no. 403, xiii–xvi.
- Edlund, S. A. 1994. The distribution of plant communities on Melville Island, Arctic Canada. In Christie, R. L., and McMillan, N. J. (eds.), The geology of Melville Island, Arctic Canada. Geological Survey of Canada Bulletin no. 450, 247–255.
- Edlund, S. A. and B. T. Alt . 1989. Regional congruence of vegetation and summer climate patterns in the Queen Elizabeth Islands, Northwest Territories, Canada. Arctic 42:3–23.
- Elberling, H. 2000. Spatial pattern of Lesquerella arctica : effects of seed bank and desiccation cracks. Ecoscience 7:86–91.
- Foster, D. R. 1988. The potential role of rime ice defoliation in tree mortality of wave-regenerated balsam fir forests. Journal of Ecology 76:172–180.
- Gold, W. G. and L. C. Bliss . 1995. Water limitations and plant community development in a polar desert. Ecology 76:1558–1568.
- Gould, W. A. and M. D. Walker . 1999. Plant communities and landscape diversity along a Canadian arctic river. Journal of Vegetation Science 10:537–548.
- Jenny, H. A. 1994. Factors of soil formation: a system of quantitative pedology. Mineola, NY: Dover Publications, 288 pp.
- Maxwell, J. B. 1981. Climatic regions of the Canadian arctic islands. Arctic 34:225–240.
- Muc, M. , J. Svoboda , and B. Freedman . 1989. Plant communities of a polar oasis at Alexandra Fiord (79°N), Ellesmere Island. Canadian Journal of Botany 67:1126–1136.
- Noy-Meir, I. 1973. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics 4:25–51.
- Porsild, A. E. and W. J. Cody . 1980. Vascular plants of continental Northwest Territories, Canada. Ottawa, Ontario: National Museum of Natural Sciences.
- Potter, L. D. , R. C. Reynolds Jr. , and E. T. Louderbough . 1985. Mancos Shale and plant community relationships: field observations. Journal of Arid Environments 9:137–145.
- Puigdefabregas, J. , F. Gallart , O. Biaciotto , M. Allogia , and G. del Barrio . 1999. Banded vegetation patterning in a subantarctic forest of Tierra del Fuego, as an outcome of the interaction between wind and tree growth. Acta Oecologica 20:135–146.
- Savile, D. B O. 1972. Arctic adaptations in plants. Ottawa: Canada Department of Agriculture, Monograph No. 6.
- Soil Classification Working Group. 1998. The Canadian System of Soil Classification. Ottawa, Ontario: NRC Research Press.
- Svoboda, J. and G. H R. Henry . 1987. Succession in marginal arctic environments. Arctic and Alpine Research 19:373–384.
- Tarnocai, C. , H. Kodama , and C. Fox . 1991. Characteristics and possible origins of the white layers found in the fossil forest deposits, Axel Heiberg Island. In Christie, R. L., and McMillan, N. J. (eds.), Tertiary fossil forest of the Geodetic Hills, Axel Heiberg Island, Arctic Archipelago. Geological Survey of Canada Bulletin no. 403, 189–200.
- Washburn, A. L. 1956. Classification of patterned ground and review of suggested origins. Geological Society of America Bulletin 67:823–865.
- White, L. P. 1971. Vegetation stripes on sheet wash surfaces. Journal of Ecology 59:615–622.
- Woo, M. K. and K. L. Young . 1997. Hydrology of a small drainage basin with polar oasis environment, Fosheim Peninsula, Ellesmere Island, Canada. Permafrost and Periglacial Processes 8:257–277.