ABSTRACT
Species distributions may be limited by substrate boundaries due to constraints at either the germination or seedling establishment life history stages. In a community of alpine tundra plants from the Sierra Nevada, some species are widespread, occurring across all four substrates studied: diorite, granite, hornfels, and marble. Other species are restricted to fewer soil types. We collected seeds from 12 species and performed a reciprocal transplant experiment in growth chambers using native soils. Eleven species germinated and established equally well across all soil types, indicating substrate chemistry did not alone determine species field distributions. However, two patterns emerged that may contribute to our understanding of species composition and dominance across substrates. First, species more dominant on the drier substrates reached at least half their total germination within two weeks of planting. Species more dominant on the wetter substrates required a longer period in wet soil to germinate. Second, there is a strong correlation between species relative abundance and percent germination, indicating that high percent germination may contribute to some species’ dominance. To determine the influence of soil type on plant size, a subset of species were grown for more time. All species were larger on hornfels and marble than on granite and diorite, indicating that species have the highest growth on the most nutrient-rich soils, rather than on their native soils. Taken together, these data suggest that water may be the limiting factor for species germination, and the differential nutrient availability across soil types has a strong influence on early seedling growth.
Introduction
Ecologists have long noted that species' distributions can be delineated by substrate boundaries (CitationWarming, 1909; CitationOosting, 1956). The absence of a species from a given soil type is often attributed either to competitive exclusion (CitationTansley, 1917; CitationKruckeberg, 1954; CitationMustart and Cowling, 1993) or to physiological constraints expressed in mature plants (CitationKruckeberg, 1954; CitationTyler, 1994; CitationGozlan and Gutterman, 1999; CitationPoot and Lambers, 2003; CitationRajakaruna et al., 2003). However, the absence of a species can also be due to its inability to germinate or to establish as a seedling under the given set of conditions (CitationRamakrishnan, 1965; CitationFossati, 1980; CitationAnderson, 1982; CitationSchauer et al., 1998; CitationMaschinski et al., 2004), as each species has a specific set of “safe site” requirements under which its establishment is optimal (CitationHarper, 1977). In particular, several studies have noted that acidic soil specialists have very low germination rates on calcitic soils or vice versa (CitationJefferies and Willis, 1964; CitationRamakrishnan, 1965; CitationRorison, 1967; CitationAnderson, 1982). Other studies have found that germination is unaffected by soil type, but plants establish better in their native soils (CitationMaschinski et al., 2004). To better understand why species composition shifts across edaphic boundaries it is important to determine at what life history stage species are excluded from a given substrate. As a first step it is therefore critical to know if species show differential germination and establishment across substrate boundaries.
Despite the large body of work investigating how adult plants segregate across soil types in alpine environments (CitationSharsmith, 1940; CitationScott and Billings, 1964; CitationPemble, 1970; CitationKörner, 1999; CitationMichalet et al., 2002), there are only a few studies that have considered the effect of rock type, or even microenvironments, on seed germination and seedling establishment in this environment (CitationFossati, 1980; CitationChapin and Bliss, 1989; CitationForbis, 2003). CitationFossati (1980) chose approximately 15 calcicole and 15 calcifuge species from the Alps and grew them in both siliceous and calcareous soils in lab and field environments. In the lab he found no differences in germination, but most species had higher establishment in their preferred soils. However, in the field, calcicole species showed lower germination on siliceous soils. Other studies have shown that soil moisture differences, also attributable to substrate shifts, are responsible for differences in germination. CitationChapin and Bliss (1989) compared pre-eruption soils and 1980-deposited tephra in a field experiment on Mount St. Helens, and they found that both study species had higher germination on the wetter pre-eruption soils, but that the relative difference in percent germination differed by species. Although comparing microsites within a single soil type, CitationForbis (2003) also showed that germination and survival differed across microsites with varying water availabilities in the Colorado Rockies, such that drier sites had lower germination but higher survival.
Despite several monographs on alpine plants that suggest they have poorer germination and establishment than plants at lower elevations (CitationBillings and Mooney, 1968; CitationBliss, 1971), the limited literature on alpine seeds suggests they behave similarly to other perennial plants (CitationFossati, 1980; CitationScherff et al., 1994; references in CitationKörner, 1999; CitationForbis, 2003). As such, seed germination is just as likely to be the stage at which plants fail to establish as in other ecosystems and should be investigated in more detail. In particular, studies have not determined seed germination and survival in California's Sierra Nevada, an especially warm and dry alpine environment (CitationWalter et al., 1975; CitationKörner and Paulsen, 2004).
In this community some species are widespread across multiple soil types, whereas others are restricted to certain soils, ostensibly by their nutrient and/or water availabilities (CitationWenk, 2005). In particular, many species' distributions are strongly correlated with or against high soil pH and calcium concentrations (CitationWenk, 2005). While some species' distributions are likely determined by adult physiological constraints or competitive exclusion, expressed as plants mature, others may be absent on one or more soil types due to their inability to germinate or establish. In the present study, four questions were addressed: First, can species germinate and survive for one growing season in soils in which they do not naturally occur? Second, do more dominant species have higher germination percentages than species less dominant across all soil types? Third, do seeds collected from the substrate type on which a species is dominant weigh more and/or display higher germination than seeds collected from soil types on which a species is less common? Fourth, in the absence of competition, is species' long-term growth higher on the substrates with which they most associate?
Materials and Methods
Study Site
Lying within the alpine zone of the Sierra Nevada mountains in eastern California is Coyote Ridge (37°11.57′N 118°31.71′W), a broad, shallow crest lying between 3500 and 3600 m at the contact between the metamorphosed Paleozoic marine sediments that dominate the Great Basin province and the granitoid Sierra batholith (CitationElliott and McKee, 1982). The large variety in rock types present on Coyote Ridge has created soils that differ greatly in their nutrient and water availabilities, creating a diversity of microsites (CitationWenk, 2005). Along the ridgeline, soils on a given rock type are formed exclusively from the outcropping rock as there is no downslope movement and mixing, creating discrete soil environments. For this study the four rock/soil types considered (diorite, granite, hornfels, and marble) differ both in chemical composition and in the texture of the soils they form. Diorite and granite have lower nutrient availabilities than hornfels and marble, while granite and marble have lower water availabilities than diorite and hornfels, creating a mosaic of water and nutrient availabilities across the four substrates (CitationWenk, 2005). Marble soils also had much higher calcium and magnesium concentrations and a correspondingly high pH of 7.4. In comparison, the other three substrates have much lower pH values, with all measurements falling between 5.8 and 6.0 (CitationWenk, 2005). Diorite and granite formed coarser textured soils than did hornfels and marble, such that diorite and granite were both composed of greater than 70% sand, while hornfels and marble soils had, respectively, 49% and 54% sand-sized particles and greater percentages of silt-sized particles (CitationWenk, 2005). All soils had between 5 and 10% clay-sized particles.
Study Species
We recorded germination and measured seed mass (μg) on 12 dicot species, including both the dominant species across all soils types and congeners that differ in their substrate associations (; CitationWenk, 2005). The species are Arenaria kingii var. glabrescens (Caryophyllaceae), Erigeron clokeyi (Asteraceae), Erig. pygmaeus, Eriogonum ovalifolium var. nivale (Polygonaceae), Erio. rosense, Ivesia lycopodioides ssp. lycopodioides (Rosaceae), I. muirii, I. shockleyi, Oxytropis borealis var. viscida (Fabaceae), O. parryi, Phlox condensata (Polemoniaceae), and Raillardella argentea (Asteraceae). All nomenclature follows The Jepson Manual (CitationHickman, 1993). Since not all species occur on all rock types, for the remainder of this paper the phrase “all species on all rock types on which they occur” is used to acknowledge that seeds from some species were collected on all four rock types, while seeds from other species were available on fewer rock types (). There are a total of 27 species by “parent soil” combinations.
Table 1 Species percent cover across the four soil types. Seeds were not collected from species by parent soil combinations in parentheses. Blank cells indicate a species does not occur on a particular soil type. To determine species percent cover, we randomly selected twenty 0.5 × 0.5 m subplots within each 10 × 10 m plot and determined the percent cover of all species on each subplot.
Germination Experiment
To investigate whether species can germinate and establish in soils from which they are absent on Coyote Ridge, we performed a reciprocal transplant experiment using a completely randomized block design, where seeds collected from each of the 12 species on all soil types on which they occur were planted into all four soil types. We collected soil during early August 2001. On each of the four soil types, we dug two to four pits for soil excavation and collected soil to a depth of 30 cm, excepting only large rocks. We collected seeds, as they matured, over three weeks in August 2001. We sorted seeds to remove obviously empty fruits and stored them in coin envelopes at 4°C for four months before use. Just prior to planting, we weighed 180 seeds, in 36 groups of five from each species, by parent soil type “treatment” on a 6-point balance (MX5, Mettler Toledo, Columbus, Ohio). For four treatments, Erio. ovalifolium collected on marble, I. shockleyi collected on diorite, P. condensata collected on diorite, and R. argentea collected on granite, we had fewer than 180 seeds and weighed the seeds we had in groups of four. We then placed each set of seeds on damp filter paper for two days prior to planting. Within those two days, three A. kingii individuals had already germinated, and some seeds from several species very visibly swollen, but no other seed coats were broken.
The groups of four or five seeds were planted into tray cells, 1.0 inch (2.54 cm) diameter × 2.5 inches (6.35 cm) deep (Stuewe and Sons, Corvallis, Oregon), in two growth chambers (Conviron, Model BDR-16, Winnipeg, Canada) at the University of California, Berkeley. For most species, three replicates of each species by “parent soil” combination were planted into each soil type (“offspring soil”) within each of three blocks, for a total of nine replicates on each of the four rock types. Two of the blocks were within one growth chamber, each occupying half of the chamber, and the third was in a second growth chamber. However, there were insufficient seeds for three species by “parent soil” combinations: I. shockleyi collected on diorite, P. condensata collected on diorite, and R. argentea collected on granite. These three treatments were planted with less replication in block 3. We planted each set of seeds approximately 0.3 cm below the soil surface. Throughout the experiment the growth chambers were set to 14-hour days with daily temperatures ranging between 4°C at night and 20°C at midday, temperature fluctuations that reflect summertime conditions on Coyote Ridge (CitationWenk, 2005). The growth chamber lights were set to their maximum level, which corresponds to 160 PAR. We watered seeds every 1–2 days for the first 6 weeks and every 2–3 days for the following 10 weeks. This watering regime kept soils as wet as they would have been during either spring snowmelt or summer periods with daily rain events. As such, the seeds received high water availability for much longer periods of time than we witnessed on Coyote Ridge during four years of research. We have observed only five seedlings in the field over these four years, all following an anomalous 10-day rain event in July, suggesting that seedling recruitment on Coyote Ridge requires unusually high water availability. We tallied seedling emergence and seedling death each day. Throughout this paper we use the term germination to encompass both the process or germination, or the breaking open of the testa, and the germling stage, such that we consider a seed to have germinated once the cotyledons appear above the soil surface.
Six species were kept for later physiology experiments, and for these species we transplanted seedlings to 2-inch (5.08-cm) cone-tainers (Stuewe and Sons, Corvallis, Oregon) after approximately six months. We estimated total above-ground biomass after either one year (A. kingii and P. condensata) or two years (Erig. clokeyi, Erig. pygmaeus, I. lycopodioides, I. muirii) of growth to determine what long-term effects offspring soil type had on plant growth. Plant size was determined by a different set of measurements for each species, based on their growth form. For A. kingii we multiplied plant width, depth, height, and percent green material as viewed from above. For P. condensata we counted total leaf number. For the remaining species we determined total leaf or shoot length by measuring the length of each green leaf (Erigeron spp.) or each shoot (Ivesia spp.). We did not harvest these individuals for actual biomass determination because we had only enough individuals for the ongoing physiological experiments. The other species that were part of the germination experiment were not kept due to lack of growth chamber space.
Statistical Analyses
To test whether either parent or offspring soil type affected percent germination, we used a fixed-factor analysis of variance model with seed germination as the dependent variable; and parent soil, offspring soil, and block, and all interaction terms as independent variables. These tests had to be done individually for each species, because not all species occur on all parent soils. For species naturally occurring on only one soil type, we omitted the parent soil term. For all tests, we have expressed final seed germination as a proportion of seeds planted in each tray cell. The proportions were arcsin-transformed to increase homogeneity of variance (CitationSokal and Rohlf, 1995). Data were visually inspected for normality and tested for homogeneity of variance using Cochran's C-test (p > 0.05). Post hoc comparisons were made using a Tukey HSD test. Since there were too few seeds for complete replication within block 3 for I. shockleyi, P. condensata, and R. argentea, we omitted block 3 from all tests including these treatments. We used an identical analysis of variance model to test for variation in seedling establishment. Seedling establishment is expressed as the proportion of seeds planted that germinated and were still alive at the end of the 16-week study period.
To test whether species differed in their percent germination, we used a single analysis of variance model with proportion germination as the dependent variable and seed type, offspring soil, block, and their interactions as independent variables, where “seed type” refers to the 27 species by “parent soil” combinations. Significant differences between seed type therefore encompass the effects of species and parent soil, two factors that cannot be distinguished with this model. I. shockleyi collected on diorite, P. condensata collected on diorite, and R. argentea collected on granite are omitted from this model since we had insufficient seeds for block 3. All data were approximately normally distributed, and variances were equal across treatments after the proportion germination data were arcsin-transformed (Cochran's C-test, p > 0.05). Post hoc comparisons were made using a Tukey HSD test. To test whether more dominant species had higher percent germination, we regressed germination on species percent cover.
For each species that naturally occurred on multiple soils in the field, we determined whether seed mass differed by parent soil using an analysis of variance model with seed mass as the dependent variable and parent substrate as the independent variable. For species with different numbers of seeds collected across “parent soils,” we equalized sample sizes by randomly removing replicates while ensuring that the mean of replicates retained equaled that of all replicates. All data were approximately normally distributed, and variances were equal across parent soil types for six of the eight species (Cochran's C-test, p > 0.05). For the other two species variances showed just slight deviations from homogeneity, but since large analysis of variance models are robust to slight deviations from homogeneity, we analyzed these data using the same model (CitationUnderwood, 1997). Post hoc comparisons were made using a Tukey HSD test.
To determine whether either parent soil or offspring soil affected species size for the six species grown for additional years, we used an analysis of variance model with plant size as the dependent variable; and parent soil, offspring soil, and their interaction as independent variables. For the species occurring on only one parent material, that term and the interaction were omitted. All data were approximately normally distributed, and variances were equal across treatments after data were log-transformed (Cochran's C-test, p > 0.05). Post hoc pairwise comparisons were made with a Tukey HSD test. All statistical tests were performed using SYSTAT 10.2 (SYSTAT Software Inc., Point Richmond, California). Tests are considered significant when p ≤ 0.05.
Results
Eleven of the 12 species showed between 9 and 60% germination, and over 90% of all germinated individuals were still alive at the end of 16 weeks, indicating they were established (, ). In this table, seedling establishment is given as the percent of all seeds planted that were alive after 16 weeks. All data for the 12th species, I. shockleyi, are omitted both from figures and from statistical tests as this was the only species that displayed negligible germination (of the 360 seeds planted, only three individuals emerged, for a percent germination of less than 1%).
Table 2 Species average germination, mortality, and establishment percentages at the end of 16 weeks.
Figure 1 Final percent germination for species at end of 16 weeks, showing differences between both parent and offspring soils. Each species by parent soil by offspring soil treatment had 9 replicates distributed in two growth chambers. Values are mean ± one standard error. Bar shading denotes the offspring soil into which seeds were planted. See text for species name abbreviations.
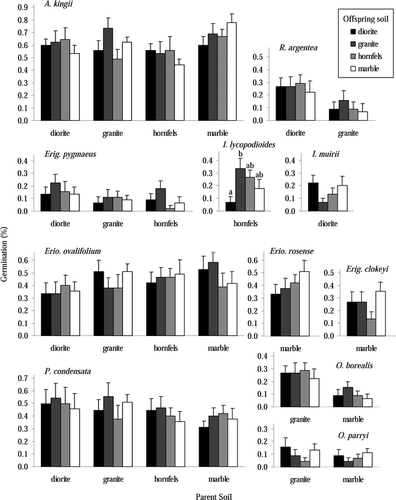
Among the remaining 11 species, I. lycopodioides was the only species to show significantly different germination across offspring soil types, with a higher germination rate on granite than on diorite (F = 7.20, p < 0.01, , ), a result that bears little relation to its field distribution pattern. I. lycopodioides occurs on only the wettest hornfels site on Coyote Ridge, although it is very common in streamside environments outside the study area (CitationWenk, 2005). Although no other species showed statistically different germination percentages across offspring soil types, there were trends within some other species: Erig. pygmaeus often had higher germination on granite, I. muirii had much lower germination on granite, and O. parryi had lower germination on granite and hornfels. In addition, only O. borealis' germination percentage was affected by the seed source; seeds collected from granite populations had much higher germination than those from marble populations (F = 19.49, p < 0.001, , ). There were no significant parent soil by offspring soil interactions. Several species had significant block or block by parent soil terms, usually reflecting that germination was lower in block 3, either for all seeds or for seeds collected on certain parent soils.
Table 3 Analyses of variance for effects of parent soil, offspring soil, and block on seed germination of each plant species. Significant differences between treatments are indicated as *p < 0.05, **p < 0.01, ***p < 0.001. MS = mean square, df = degrees of freedom.
Since more than 90% of germinants survived until the end of the experiment, the patterns for seedling establishment were very similar to those for seed germination (). However, a large number of A. kingii seedlings on hornfels died, such that A. kingii's percent establishment was significantly lower on hornfels than on granite (; results of Tukey test of pairwise comparisons not shown), even though A. kingii is most abundance on hornfels in the field. Most of the seedlings died over a several-day period 20 days into the experiment when insufficient water was administered on one day and the surface of the hornfels soil quickly formed a very dry, hard surface layer. In addition, higher mortality of Erig. pygmaeus collected on hornfels, measured as the percent of germinants that died before the end of the study, meant that Erig. pygmaeus seeds collected on diorite had higher establishment than those from the hornfels population (F = 3.16, p < 0.05, ).
Table 4 Analyses of variance for effects of parent soil, offspring soil, and block on seedling establishment of each plant species. Significant differences between treatments are indicated as: *p < 0.05, **p < 0.01, ***p < 0.001.
Although species generally did not respond to different offspring soils with higher or lower percent germination or establishment across offspring soils, different species varied significantly in their percent germination regardless of offspring soil type, and they germinated at different rates after planting (, ; ). The analysis of variance comparing all “seed types” showed a strongly significant “seed type” effect (F = 29.49, p < 0.001, ) as well as a significant “seed type” by block interaction (F = 2.13, p < 0.001). The significant “seed type” by block interaction term reflects that some species showed depressed germination in block 3 while others did not. The post hoc pairwise comparisons of all “seed types” demonstrates both how little variation there was among populations of a certain species, and how different species can be from one another, as the multiple populations of a given species never have significantly different germination percentages from one another (). In addition, these results show that the species dominant on a given site had higher germination than less dominant species (; , , ). The four populations of A. kingii had the four highest germination percentages, followed by the nine populations of P. condensata and Eriogonum spp. seeds. A. kingii and P. condensata are the two dominant species across all soil types on Coyote Ridge (). Erio. ovalifolium is a dominant species on diorite, granite, and hornfels, while on marble Erio. rosense becomes a dominant species. Falling at the other end of the spectrum are Erig. pygmaeus (the fourth most widespread species but one that is never dominant), I. lycopodioides, and O. parryi ().
Table 5a Analysis of variance for effects of seed type (species by parent soil combinations), offspring soil, and block on seed germination among the 11 plant species. Treatments are significantly different from one another if the p-value shown is less than 0.05.
Table 5b Tukey test probabilities comparing percent germination between different “seed types”. Parent soils indicated in parentheses.
Table 6 Analysis of variance for effect of parent substrate of seed mass (µg) for each species. Mean and one standard error, in parentheses, given for each soil type. Significant differences between treatments are indicated as: *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 2 Germination curves for species across time. All “parent” and “offspring” soil treatments are averaged.
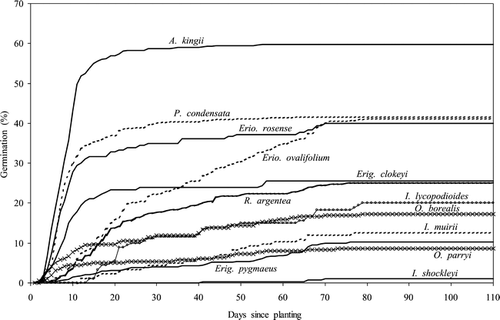
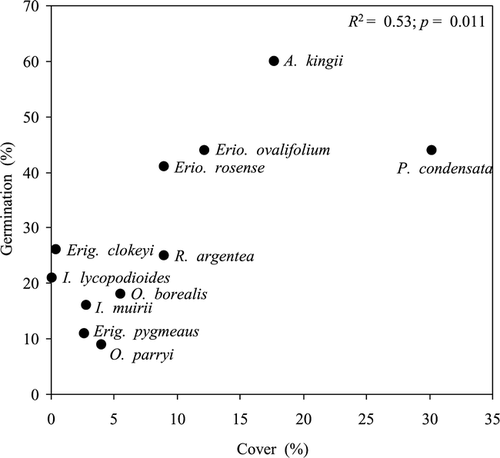
Figure 3 Relationship between percent cover on sites on which a species in present and percent germination, across all offspring soil types.
Although the experimental design had insufficient degrees of freedom for repeated measures of analysis of variance, the plot of seed percent germination by time provides additional information (, ). Some species began germinating much more quickly than others, with A. kingii reaching over 80% of its final percent germination within two weeks of planting (). Other species, such as Erio. ovalifolium, eventually reached a high percent germination, but the seeds required a much longer period in wet soil to emerge fully. It is apparent that many of the species present only on wetter sites are those that began germinating later, such that many of these species' seeds do not germinate until they have had high soil water for more than a month.
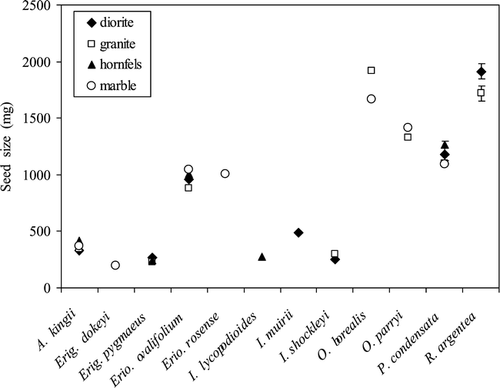
While parent soil only affected germination or establishment in O. borealis (, ), seed mass did differ significantly between parent soils for all eight species that occur across multiple soil types (, ). Two patterns emerge from these data. These data show that seeds that came from the substrates with higher nutrient availability were larger, such that most species had the largest seeds on either hornfels or marble even if they did not have the highest percent cover on these substrates (; , ). Notably, O. borealis, the species with the largest differences in seed mass between parent soil types (F = 23.44, p < 0.001), was the species for which parent soil affected germination rate.
Figure 4 Seed masses of species on each soil type on which they naturally occur. Values are mean ± one standard error. Error bars are only shown where larger than symbols.
While some species showed limited long-term growth differences based on the seed origin, all species showed large plant size differences based on offspring soil (, ). Five of six species had the greatest size on hornfels, the soil with the highest nutrient availability, and the second greatest size on marble (). I. muirii plants growing on marble had greater size than those on hornfels (). These results demonstrate that all species were able to continue growing on all substrates and responded to increased nutrients with higher growth. In addition there was a significant “parent soil” effect for A. kingii and P. condensata, such that seeds collected on marble produced the largest plants, even though those were not the largest seeds for either species (,
Figure 5 Plant biomass estimates after either one year (A. kingii and P. condensata) or two years (other species) of growth. Values are mean ± one standard error. Bar shading denotes the offspring soil into which seeds were planted.
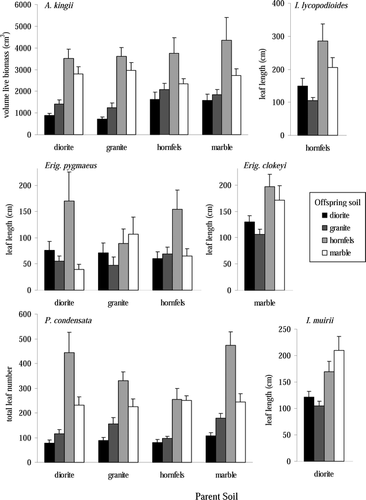
Table 7 Analysis of variance for effects of offspring soil type on plant biomass in growth chambers. Biomass for A. kingii is volume of live material (cm3), for P. condensata is total leaf number, and for the other species is total leaf length (mm). Mean and one standard error, in parenthesis, given for each offspring soil type. Significant differences between treatments are indicated as: + p < 0.10, *p < 0.05, *p < 0.01, ***p < 0.001.
Discussion
Specific Goals Of Study
The results of this experiment demonstrate that offspring soil type did not directly affect seed germination. The demonstrated ability of all other species to germinate across all offspring soil types, often with high total germination, suggests that when given ample water over long periods of time, offspring soil type does not limit seed germination in these Coyote Ridge species. However, interspecific differences in percent germination and timing of germination can be important in explaining patterns of species dominance across all four soil types and species ability to persist on the driest sites. In addition, differences in growth and seed size across the four soil types reflect their nutrient availabilities, suggesting that all species would thrive on hornfels and marble in absence of water limitation or competitive effects.
The lack of an offspring soil type effect on seed germination and seedling establishment () counters that reported in several other studies, especially across acidic-calcite soil boundaries (CitationJefferies and Willis, 1964; CitationRamakrishnan, 1965; CitationRorison, 1967; CitationAnderson, 1982). However, the results of the present experiment are consistent with the assessment that, although many species are substrate specialists on Coyote Ridge, none of the study species are listed as substrate specialists in the local floras (CitationHickman, 1993; CitationWeeden, 1996). The germination results therefore suggest that each species has specific nutrient and/or water requirements, and on Coyote Ridge these soil requirements are closely correlated with substrate. However, appropriate “safe sites” for seedlings may be present on additional substrates at lower elevations or in more protected locations (CitationPemble, 1970; CitationWenk, 2005). Therefore, under the moister, more protected environment created in the growth chambers, all four substrates provided the necessary germination conditions for these species. This result is also consistent with many studies that suggest moisture availability is the main factor limiting seedling establishment in alpine environments (CitationUrbanska and Schutz, 1986, and references therein; CitationChapin and Bliss, 1989).
Although the results of this experiment demonstrate that the substrate specialists are not restricted to certain offspring soils by germination requirements, they do suggest that patterns of species dominance across all soil types may be partially explained by germination characteristics (, ; ). The correlation between percent cover and percent germination () suggests that species may be more dominant on a given soil type because they have higher percent germination than other species. Species present on marble, the driest soil under field conditions, had a more rapid onset of germination than species present only on wetter soils (). This pattern suggests that rapid recruitment is important to establishing on the drier sites, especially in an environment where surface soils do not stay moist for very long. Indeed, over the four years we conducted physiological studies on Coyote Ridge, only once did we see seedlings, and that was after an unusual 10-day rain event in early July 2001. At that time we noted three A. kingii seedlings and two Erio. ovalifolium seedlings. In an alpine environment as dry as the Sierra Nevada, we expect seedlings will establish during unusually wet summers, although they may also germinate during spring runoff. Under such circumstances the large differences in time to germination could be important in determining species recruitment patterns. Moreover, recent evidence suggests the Sierra Nevada will warm dramatically over the next century with slight to moderate decreases in growing season rainfall (CitationHayhoe et al., 2004), such that seedling recruitment may become even rarer in the future and species with rapid germination rates may become more dominant across all sites.
Seed mass measured across parent soil types () and plant growth measured across offspring soil types () demonstrate that these species respond to the different nutrient availabilities across the soil types. All species, regardless of field substrate association, had the highest long-term growth on the most nutrient-rich substrates (CitationWenk, 2005); this demonstrated that species growth was not highest on the substrate with which they most associate, but rather when they were provided with the highest nutrient levels. These findings again suggest biotic interactions or effects of water availability, which we were unable to mimic with this experiment, determine species' field distributions. Seed masses were also usually highest on hornfels and marble, suggesting that parent plants could allocate more resources to seed production on more nutrient-rich soils. However, only for one species, O. borealis, did this increased maternal allocation result in higher percent germination or seedling establishment. CitationTyler and Zohlen (1998) showed that despite large differences in soil chemistry and adult plant leaf chemistry between acidic and calcic soils, seed chemistry was more constant across the gradient, implying that parent plants were ensuring that seeds received the correct balance of necessary nutrients. It is possible that despite some difference in embryo size, all seeds have similar elemental concentrations and that parent seed source therefore makes little difference to seed germination.
Broader Application of Results to Alpine Research
In addition to addressing the specific questions outlined for this study, the results add to the limited knowledge about alpine germination patterns, especially in the Sierra Nevada alpine environment with its warm, dry summers. The results agree with the findings of several studies and reviews suggesting alpine germination and establishment rates are high when provided with the appropriate conditions (CitationSöyrinki, 1938; CitationUrbanska and Schutz, 1986; CitationChapin and Bliss, 1989; CitationChambers et al., 1990; CitationKörner, 1999; CitationForbis, 2003). However, we do not wish to suggest that we expect natural recruitment rates on Coyote Ridge to approach the levels observed by CitationForbis (2003) on Niwot Ridge in the Colorado Rockies, as mid-season rain events are rare in the Sierra. If such rains do occur, then the results of the study shown here indicate that a large percent of seeds set are viable and constitute therefore an important seed bank.
Many studies report that alpine plants require vernalization to germinate (CitationSöyrinki, 1938; CitationAmen, 1966; CitationFossati, 1980), and we expect the same requirement exists for the species occurring on Coyote Ridge. All seeds in this experiment were subjected to four months of cold storage before germination, but as we do not have germination data for seeds that were not placed in cold storage, we do not know whether the cold treatment was necessary for germination. For a separate pilot experiment we noted that after soil cores were stored at −2°C for six weeks and then watered, they displayed high germination, while no germination was observed prior to the cold storage.
The average seed mass values of seeds collected on Coyote Ridge fell within the range observed in other alpine systems (CitationKörner, 1999). In addition, interspecific differences match expectations for seed mass differences among families (CitationKörner, 1999). Members of Fabaceae (Oxytropis spp.) and Polygonaceae (Eriogonum spp.) usually have larger seeds, while species in Caryophyllaceae (Arenaria) tend to have smaller seeds. However, while Rosaceae species often have large seeds (CitationKörner, 1999), the three Ivesia species on Coyote Ridge had some of the smallest seeds.
Despite previous studies suggesting that many alpine seeds have very short dispersal distances (CitationScherff et al., 1994; CitationKörner, 1999), we do not believe that dispersal limitations have created the species disjunctions across substrate boundaries. The four soil types are interdigitated across Coyote Ridge, yet the same species disappear and reappear every time certain substrate boundaries occur. In addition we have observed seeds freely moving considerable distances on many windy days.
Conclusions
Overall, this study suggests that under the high moisture conditions of the growth chamber, seeds did not respond to offspring soil type differences. We cannot be sure that under the drier field conditions on Coyote Ridge the chemical or moisture differences across soil types are not limiting species distribution at early life history stages. Species that grow on drier soils in the field had a more rapid onset of germination than species present only on wetter soils. This result fits nicely with other research we have conducted on many of the same species that shows species' differential abilities to tolerate water deficit is an important determinant of performance and therefore distribution in this alpine community (CitationWenk, 2005). These results are similar to those observed by CitationChapin and Bliss (1989) in another dry alpine environment.
Acknowledgments
We would like to thank Vanessa Schmidt, Primrose Boynton, Neil Hausmann, and Sharon Ng for their help with this experiment. A National Science Foundation pre-doctoral research grant and a graduate student research grant from the White Mountain Research Station, Bishop, California, provided funding for this project.
Related Research Data
References Cited
- Amen, R. D. 1966. Extent and role of seed dormancy in alpine plants. Quarterly Review of Biology 41:271–281.
- Anderson, C. A. 1982. The effect of calcium on the germination, growth and mineral-nutrition of acidic and calcareous populations of Eucalyptus obliqua Lherit. Plant and Soil 69:213–223.
- Billings, W. D. and H. A. Mooney . 1968. The ecology of arctic and alpine plants. Biological Reviews 43:481–529.
- Bliss, L. C. 1971. Arctic and alpine plant life cycles. Annual Review of Ecology and Systematics 2:405–438.
- Chambers, J. C. , J. A. MacMohan , and R. W. Brown . 1990. Alpine seedling establishment: the influence of disturbance type. Ecology 71:1323–1341.
- Chapin, D. M. and L. C. Bliss . 1989. Seedling growth, physiology, and survivorship in a subalpine, volcanic environment. Ecology 70:1325–1334.
- Elliott, G. S. and E. H. McKee . 1982. Geologic map of the Coyote SE and Table Mountain roadless areas, Inyo County, California. scale 1:62,500. Reston, Virginia U.S. Geological Survey.
- Forbis, T. A. 2003. Seedling demography in an alpine ecosystem. American Journal of Botany 90:1197–1206.
- Fossati, A. 1980. Keimverhalten und frühe Entwicklungsphasen einiger Alpenpflanzen. Veröfentlichungen des Geobotanischen Institutes ETH Zürich 73.
- Gozlan, S. and Y. Gutterman . 1999. Dry storage temperatures, duration, and salt concentrations affect germination of local and edaphic ecotypes of Hordeum spontaneum (Poaceae) from Israel. Biological Journal of the Linnean Society 67:163–180.
- Harper, J. L. 1977. The Population Biology of Plants London, U.K Academic Press.
- Hayhoe, K. , D. Cayan , C. B. Field , P. C. Frumhoff , E. P. Maurer , N. L. Miller , S. C. Moser , S. H. Schneider , K. N. Cahill , E. E. Cleland , L. Dale , R. Drapek , R. M. Hanemann , L. S. Kalkstein , J. Lenihan , C. K. Lunch , R. P. Neilson , S. C. Sheridan , and J. H. Verville . 2004. Emissions pathways, climate change, and impacts on California. Proceedings of the National Academy of Sciences of the United States of America 101:12422–12427.
- Hickman, J. C. 1993. The Jepson Manual: Higher Plants of California Berkeley, California University of California Press. 1400.
- Jefferies, R. L. and A. J. Willis . 1964. Studies on the calcicole-calcifuge habit. 2. The influence of calcium on the growth and establishment of 4 species in soil and sand cultures. Journal of Ecology 52:691–707.
- Körner, C. 1999. Alpine plant life : functional plant ecology of high mountain ecosystems Berlin, New York Springer. ix + 338.
- Körner, C. and J. Paulsen . 2004. A world-wide study of high altitude treeline temperatures. Journal of Biogeography 31:713–732.
- Kruckeberg, A. R. 1954. The ecology of serpentine soils: III. Plant species in relation to serpentine soils. Ecology 35:267–274.
- Maschinski, J. , J. E. Baggs , and C. F. Sacchi . 2004. Seedling recruitment and survival of an endangered limestone endemic in its natural habitat and experimental reintroduction sites. American Journal of Botany 91:689–698.
- Michalet, R. , C. Gandoy , D. Joud , J. P. Pages , and P. Choler . 2002. Plant community composition and biomass on calcareous and siliceous substrates in the northern French Alps: comparative effects of soil chemistry and water status. Arctic and Alpine Research 34:102–113.
- Mustart, P. J. and R. M. Cowling . 1993. The role of regeneration stages in the distribution of edaphically restricted Fynbos Proteaceae. Ecology 74:1490–1499.
- Oosting, H. J. 1956. The Study of Plant Communities San Francisco, California W.H. Freeman and Co. 440.
- Pemble, R. H. 1970. Alpine vegetation in the Sierra Nevada of California as lithosequence and in relation to local site factors Ph.D. thesis. Davis Department of Botany, University of California.
- Poot, P. and H. Lambers . 2003. Are trade-offs in allocation pattern and root morphology related to species abundance? A congeneric comparison between rare and common species in the south-western Australian flora. Journal of Ecology 91:58–67.
- Rajakaruna, N. , M. Y. Siddiqi , J. Whitton , B. A. Bohm , and A. D. M. Glass . 2003. Differential responses to Na+/K+ and Ca2+/Mg2+ in two edaphic races of the Lasthenia californica (Asteraceae) complex: a case for parallel evolution of physiological traits. New Phytologist 157:93–103.
- Ramakrishnan, P. S. 1965. Studies on Edaphic Ecotypes in Euphorbia thymifolia L. 1. Seed-germination. Journal of Ecology 53:157–162.
- Rorison, I. H. 1967. A seedling bioassay on some soils in Sheffield area. Journal of Ecology 55:725–741.
- Schauer, A. J. , B. K. Wade , and J. B. Sowell . 1998. Persistence of subalpine forest-meadow ecotones in the Gunnison Basin, Colorado. Great Basin Naturalist 58:273–281.
- Scherff, E. J. , C. Galen , and M. L. Stanton . 1994. Seed dispersal, seedling survival and habitat affinity in a snowbed plant—Limits to the distribution of the snow buttercup, Ranunculus adoneus . Oikos 69:405–413.
- Scott, D. and W. D. Billings . 1964. Effects of environmental factors on standing crops and productivity of an alpine tundra. Ecological Monographs 34:243–270.
- Sharsmith, C. W. 1940. A contribution to the history of the alpine flora of the Sierra Nevada. Ph.D. thesis. Berkeley Department of Botany, University of California.
- Sokal, R. R. and F. J. Rohlf . 1995. Biometry: the Principles and Practice of Statistics in Biological Research. Third edition New York W. H. Freeman and Company.
- Söyrinki, N. 1938. Studien uber die generative und vegetative Vermehrung der Samenpflanzen in der alpinen Vegetation Petsamo-Lapplands. Annales Botanici Societatis Zoologicae Botanicae Fennicae Vanamo (Helsinki) 11:1–311.
- Tansley, A. G. 1917. On competition between Galium saxatile L. (G. hercynicum Weig.) and Galium sylvestre Poll. (G. asperum Schreb.) on different types of soil. Journal of Ecology 5:173–179.
- Tyler, G. 1994. A new approach to understanding the calcifuge habit of plants. Annals of Botany 73:327–330.
- Tyler, G. and A. Zohlen . 1998. Plant seeds as mineral nutrient resource for seedlings—A comparison of plants from calcareous and silicate soils. Annals of Botany 81:455–459.
- Underwood, A. J. 1997. Experiments in ecology: their logical design and interpretation using analysis of variance Cambridge, U.K Cambridge University Press.
- Urbanska, K. M. and M. Schutz . 1986. Reproduction by seed in alpine plants and revegetation research above timberline. Botanica Helvetica 96:43–60.
- Walter, H. , E. Harnickell , and D. Mueller-Dombois . 1975. Climate-diagram maps of the individual continents and the ecological climatic regions of the earth Berlin Springer-Verlag.
- Warming, E. 1909. Oecology of Plants: an Introduction to the Study of Plant-Communities Oxford, U.K Oxford University Press. 422.
- Weeden, N. F. 1996. A Sierra Nevada Flora. Fourth edition Berkeley, California Wilderness Press. 259.
- Wenk, E. H. 2005. Physiology and distribution of plants in relation to substrate on Coyote Ridge in the alpine Sierra Nevada of California Ph.D. thesis. Berkeley Department of Integrative Biology, University of California. 297.