Abstract
In a warming climate, sexual reproduction may play an important role in plant community composition in arctic tundra. As temperatures increase and currently immobilized soil nutrients become available, flowering, fruiting, and germination conditions will likely improve. We examined how experimentally adding soil nutrients for 13 and 20 years affected species composition, flower and fruit production, seed dispersal, composition of the seed bank, and seedling establishment in a dry heath plant community in northern Alaska. Fertilizer addition significantly shifted adult community composition by decreasing lichens and evergreen shrubs and increasing abundance of a bunchgrass, Hierochloe alpina, and dwarf birch, Betula nana. More seeds were dispersed adjacent to nutrient amended plots, particularly of B. nana, and soil seed banks differed significantly between control and fertilized soils reflecting the adult communities in the field. Few seedlings were observed in any field plots. However, seeds of H. alpina likely played a role in the community shift because this species has few, small individuals in control plots, yet is densely packed in fertilized plots. B. nana, on the other hand, appears to be increasing in relative abundance via vegetative growth of existing individuals. Therefore, although sexual reproduction leading to seedling establishment is rare currently, as nutrients become more available in a warming climate, individuals may recruit from seed more often as long as space is available.
Introduction
Almost all the plants in the Arctic are slow-growing perennials that are adapted to survive stressful abiotic limitations such as low mean annual temperature and precipitation, a short growing season, soil moisture extremes, and low soil nutrient availability (CitationBabb and Whitfield, 1977; CitationChapin et al., 1978, Citation1987; CitationEllis, 1980; CitationBillings, 1987). These conditions are not favorable to the metabolic processes of soil microorganisms (CitationBliss, 1962; CitationClein and Schimmel, 1995; CitationMikan et al., 2002); therefore, while total stocks of N and P in soils may be substantial, a large proportion is bound in organic forms not directly available to plants (CitationShaver and Chapin, 1986; CitationBillings, 1987; CitationChapin et al., 1995). Most arctic plants direct substantial energy into clonal growth, allowing them to reproduce vegetatively and acquire and utilize nutrients in the highly heterogeneous and nutrient-limited landscape (CitationChester and Shaver, 1982; CitationJonsdottir et al., 1996).
Despite the prevalence of clonality, arctic plant species often invest in flowers and fruits to produce seeds. Seeds allow plants to introduce genetic variation within populations and communities and to colonize disturbed sites (CitationBaskin and Baskin, 2001). While arctic seed banks are often dense and diverse (CitationMcGraw and Vavrek, 1989; CitationAlsos et al., 2003; CitationCooper et al., 2004), species composition and abundance of arctic soil seed banks do not often well represent the adult community (CitationMcGraw, 1980; CitationFox, 1983; CitationDiemer and Prock, 1993; CitationMolau and Larsson, 2000). The absence of particular plant species as adults may be due to limited seed production, differences in dispersal capabilities among species, or an inability of seeds to persist in arctic soils (CitationAlsos et al., 2003; CitationCooper et al., 2004); graminoid seeds have been shown to persist in arctic soils longer than other plant groups (CitationBillings and Mooney, 1968; CitationMcGraw, 1980).
Despite the presence of a soil seed bank, germination and recruitment from seed are infrequently observed in many arctic tundra communities. Like growth of the adult plant community, germination of seeds in the Arctic is limited by biotic and abiotic factors. Laboratory experiments suggest that current arctic temperatures most likely limit germination in the field since high levels of germination occur under increased experimental temperature conditions (CitationBell and Bliss, 1980; CitationCooper et al., 2004). Competition for space may also inhibit germination. Natural disturbances, such as frost boils (areas of exposed mineral soil) and animal foraging, are important for opening space for potential seed germination (e.g., CitationGartner et al., 1986; CitationChang et al., 2001; CitationEskelinen and Virtanen, 2005; CitationGough, 2006; CitationLindgren et al., 2007). These results suggest that the bottleneck of germination is caused by limitations in soil temperature, available space, and ultimately whether or not viable seeds are present or can be dispersed into the area. Despite the usual absence of seedlings in the most common tundra communities in northern Alaska, evidence of substantial genetic diversity has been documented in arctic ecosystems (e.g., CitationAlsos et al., 2007), supporting the fact that viable seeds are produced, dispersed, and do successfully germinate in the tundra landscape, but likely quite infrequently (CitationBillings and Mooney, 1968; CitationMcGraw and Vavrek, 1989).
Global warming may change the ways in which flowering plants reproduce sexually and establish new individuals. Recent studies show that an air temperature increase of about 3 °C would significantly increase the reproductive efforts of arctic plants in heath tundra (CitationAlatalo and Totland, 1997; CitationKlanderud, 2005). A review by CitationAerts et al. (2006) of the general responses of cold, northern ecosystems to climate change indicated that plant phenology has shifted earlier with recent climate warming, and geographic ranges may be moving northward. Studies also suggest that increases in soil temperature enhance microbial activity and decomposition, which could potentially lead to higher inorganic N availability in the tundra (CitationRustad et al., 2001; CitationAerts et al., 2006). The newly available nutrients and warmer temperatures may allow adults to flower more, thus increasing seed production. Therefore, a warmer climate may benefit germination both directly, through increased temperatures, and indirectly, through soil nutrient availability affecting sexual reproduction rates.
STUDY OBJECTIVES
Based on previous studies of nutrient manipulations in Alaskan dry heath tundra, we tested the following hypotheses regarding the role of sexual reproduction in plots that had received added soil nutrients for either 13 or 20 years. First, we expected the flowers and fruits produced in fertilized plots to reflect the adult species composition and thus differ significantly from control plots (H1). Prior results from this community (CitationGough et al., 2002) showed that graminoids and dwarf deciduous shrubs increase vegetative cover and biomass, and related studies suggest a positive correlation between vegetative growth and flower production (CitationGough et al., 2007). Second, we expected that the seed rain would reflect localized dispersal and would thus differ adjacent to fertilized and control plots (H2). Here we define dispersal as the process of a seed leaving the parent plant. The species in this community are predominantly wind-dispersed with the exception of the berry producers (e.g., Vaccinium uliginosum, Vaccinium vitis-idaea subsp. minus). Third, we expected the seed bank composition to also reflect the adult community when comparing control and fertilized plots (H3). Finally, we expected a higher establishment rate of seedlings of graminoid and deciduous species in fertilized treatments than in the control treatments, reflecting the shift in adult vegetation, seed production, and localized dispersal inherent in the previous hypotheses (H4). To test these four hypotheses we used a combination of field observations of long-term experimental manipulations and greenhouse experiments.
Methods
STUDY SITE AND EXPERIMENTAL DESIGN
Field studies were conducted at the Arctic Long Term Ecological Research (LTER) dry heath (DH) tundra plots located near the north end of Toolik Lake, Alaska (68°38′N, 149°43′W, elevation 760 m a.s.l.; CitationShaver and Chapin, 1991; CitationGough et al., 2002). DH tundra is a relatively common type of tundra vegetation found in North America, Greenland, Europe, and Asia, and is composed mostly of plants in the heath family, Ericaceae, and lichens (CitationBliss and Matveyeva, 1992; CitationShaver and Jonasson, 1999). At Toolik Lake, DH is dominated by lichens; deciduous dwarf shrubs, including Arctous alpina (previously Arctostaphylous alpina), Vaccinium uliginosum, and Salix phlebophylla; and evergreen dwarf shrub species, including Empetrum nigrum, Rhododendron tomentosum subsp. decumbens (previously Ledum palustre), Kalmia procumbems (previously Loiseleuria procumbens), and Vaccinium vitis-idaea subsp. minus (nomenclature follows CitationElven, 2010). Under ambient nutrient conditions, the graminoid Hierochloe alpina is rare with small individuals of 3–4 leaves, but in fertilized plots it grows into a tussock form. The dwarf deciduous shrub Betula nana is also present, but rare.
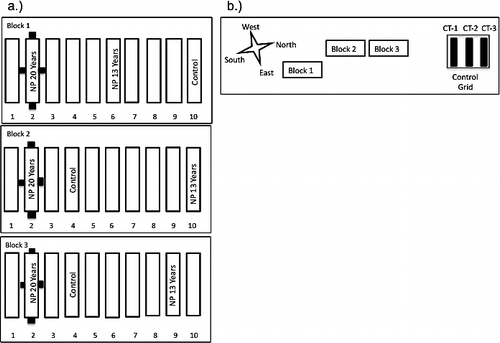
Three blocks of ten 5 × 20 m plots were initially established in 1989 as part of the LTER (see http:/ecosystems.mbl.edu/ARC) (). Control (CT) or N plus P (NP) treatments were randomly assigned to one plot each within the block design, thus each treatment was replicated 3 times. A similar experiment was initiated within the same experimental blocks in 1996 creating 3 additional replicates of control and NP plots. Annual spring (early June) application of both N (10 g m−2 as NH4NO3) and P (5 g m−2 as P2O5) in pellet form started in 1989 for the 20 years of NP treated plots, and 1996 for the 13 years of NP treated plots. The CT plots established in 1996 were used for all CT results presented here ().
FIGURE 1 Schematic map of the Long Term Ecological Research (LTER) Dry Heath experimental plots. Each plot has a treatment assigned, but only plots used in this experiment are labeled. (a) Plot layout within blocks; black squares are seed rain trap areas. (b) Site layout including Control Grid 35 m north of LTER blocks. Map courtesy of J. Laundre, Marine Biological Laboratory, Woods Hole, Massachusetts.
FIELD DATA
Community Composition
Permanent monitoring quadrats were randomly established in 1998 in each plot. Aerial percent cover was estimated in all three blocks of the CT, 13-year NP, and 20-year NP plots of the LTER DH in mid- to late July 2007 and 2008. A 1 × 1 m quadrat, divided into 20 × 20 cm squares, was placed 0.25 m inside the edge of the plot at a permanently marked location to allow resampling of the same area each year. Percent cover of moss, lichen, and all vascular plant species was estimated visually, with additional estimates of litter and animal activity. The quadrat was then moved to an adjacent 1 m2 area and observations were repeated; this was done a total of eight times to sample an area of 8 m2 in each plot. Relative percent cover of growth forms was calculated by dividing the summed cover of individuals in a growth form by the total plant cover (sum of all non-vascular and vascular species) for that 1 m2 plot.
Flower and Fruit Production
Flowers and fruits of all plant species were counted on a per area (not per individual) basis during the percent cover estimates in both 2007 and 2008 to provide an index of the number of sexual reproductive structures produced that season. The front 0.2 m2 of each 1 m2 quadrat used in percent cover (described above) was sampled per treatment per block to sample an area of 1.6 m2 in each plot. Although H. alpina flowers were counted in 2008, a different method was used that is not comparable, so these data are not reported.
Seedling Surveys
Non-destructive seedling censuses were conducted at the same time flowers and fruits were counted, using different areas of 0.2 m2 of each 1 m2 quadrat surveyed for percent cover (described above) each year to avoid duplicate recordings. A seedling was defined as a single individual with less than 10 leaves that was physically separate from established adult plants (minimum distance from adult plant >5 cm) to avoid considering a new clonally produced ramet as a seedling. Litter and canopy vegetation were shifted, but not removed during searches. Seedlings were counted and identified to species when possible, and grouped by growth form as graminoid, ericaceous, deciduous, or forb.
Seed Rain Experiment
To assess seed availability in unmanipulated vegetation, in late July 2007, a 3 × 3 control grid was set up 35 m north of the three LTER blocks (). Three parallel rows numbered CT-1, CT-2, and CT-3 were set up 10 m apart from each other, with three 2 × 2 m trap areas 12 m away from each other within each row (n = 9). To assess seed availability in the immediate vicinity of fertilized plots, four 2 × 2 m “NP” trap areas were set up within the LTER research site on the edge of the plots that received 20 years of NP treatment in each of the three blocks (one in each cardinal direction; n = 12; ).
In early August 2007, seed rain traps modified from Molau (CitationMolau and Molgaard, 1996; CitationMolau and Larsson, 2000) were placed in each trap area and left over winter. Three 0. 5 × 0.5 m (0.25 m2) artificial turf mats were placed in a triangle for each trap area. Each mat was placed 1 m from the other within the 2 × 2 m trap area. The mats were then fixed to the ground using 13 cm nails in two opposing corners, and GPS coordinates of the plots were taken. Artificial grass mats are best for capturing seeds 0.3 mm or larger, which includes seeds of most of the species reported here (CitationMolau and Larsson, 2000).
On 11 June 2008, the final snowmelt of the spring, each mat was collected and allowed to air dry in a field laboratory. On 21 July 2008, seeds were removed from the mats, then placed in a 40 °C drying oven for seven days. Samples were transported back to University of Texas (UT) Arlington where individual seed trap samples were separated using four different mesh size soil sieves. Seeds and berries were identified and counted in the lab using an Olympus SZ30 stereo microscope. A reference collection of seeds and fruits used for identification was collected from dry heath, moist acidic and moist non-acidic tussock tundra sites in the summer of 2007.
GREENHOUSE DATA
Seedbank Experiment
On 3 August 2006, 5-cm-deep soil cores were taken adjacent to CT and within 13-year NP plots in the LTER DH site using a 5-cm-diameter corer. In each of the three blocks, four cores were randomly located in an area designated for destructive sampling in the 13-year NP treatment plots, and directly south of the CT plots (n = 12 cores per treatment; n = 24). The cores were shipped to UT Arlington and placed in cold storage (4 °C) for 6 months. In February 2007, the four cores from each treatment and block combination were homogenized after removing live mosses, vascular plants, roots, rhizomes, and rocks greater than 1 cm in diameter. Clear plastic deli cups (4.3 cm deep × 11.7 cm diameter) with three drainage holes were filled half full with a mixture of equal parts potting soil and sand. The homogenized sample was used to generate 8 replicate subsamples by placing 1 cm layer of homogenized soil on top of the potting soil mix. Eight deli cups with the potting soil/sand mixture only were used as blanks to allow for identification of non-target seedlings in the greenhouse.
The deli cups were placed in the greenhouse facility at UT Arlington on 2 February 2007. Temperature was set to follow a 20 °C daily maximum and a 15 °C nightly minimum temperature (day/night: 16/8 hour) cycle. To simulate a 24-hour growing season photoperiod, supplemental lighting was provided during both day and night by overhead fluorescent lighting. Replicates were watered with tap water three to four times a week, and monitored daily until the first seed germinated. All cups were rearranged randomly each week to avoid possible temperature gradients within the greenhouse.
Seedlings were monitored weekly for eight weeks and marked using colored toothpicks. Non-target seedlings, grasses, and forbs not of Alaskan origin were removed once identified. Alaskan seedlings were gently removed and repotted in a potting soil mixture for later identification. Transplanted seedlings that dried up prior to identification to species were categorized as graminoid, ericaceous, or unknown.
STATISTICAL ANALYSIS
All statistical analyses were carried out using SAS v. 9.1.3 for Windows (SAS Institute, Cary, North Carolina, USA). Community composition was compared among the three levels of fertilization treatment using a MANOVA including a block factor to reflect the experimental design; each 1 m2 quadrat was nested within block. We only report data analysis from 2008 to focus on the longest running data; patterns in 2007 were quite similar. The dependent variables included deciduous shrubs, evergreen shrubs, graminoids, lichen, and moss. Additional cover categories including litter, bare ground, and “animal category” were analyzed separately. Animal category included all observed and quantified activity by voles and caribou (i.e. caribou feces, grass litter created by voles, vole trails, and vole feces) within each treatment. Data were arcsine-square-root transformed prior to analysis to meet parametric model assumptions. Differences among fertilization treatments were examined using Wilk's Lambda as the test statistic.
Species richness and diversity were analyzed separately using an ANOVA to test for fertilizer effect with block and quadrat as described above. Because the data were marginally not normally distributed, we used the more conservative Scheffe's test for pairwise differences and interpret these results cautiously. To test whether there were differences in number of flowers and fruits among fertilization treatments, we used a Chi square test of independence on the 2007 flower and fruit data, where flowers and fruits were summed within plots for each species to represent total number of sexual reproductive units per plot for that year. To determine effects of proximity to the experimental plots on seed dispersal patterns, we first analyzed the seed rain results for total seeds and B. nana separately for the CT trap areas only with a one-way ANOVA with “block” as a factor of interest reflecting distance from the experimental plots, including trap area and mat nested in trap area to reflect the sampling design. We then analyzed the seed rain results as a one-way ANOVA with fertilization treatment as the main effect, and included block as a blocking factor, trap area as a subsample, and mat nested within trap area. The seed bank results were analyzed as a one-way ANOVA with fertilization as the main effect, cores nested within block, and replicates nested within cores.
Results
PERCENT COVER, FLOWERS AND FRUITS
Community composition shifted significantly with fertilization, as indicated by an overall significant treatment effect in the MANOVA (F10,84 = 51.30, P < 0.0001). Moss (F2,46 = 19.30, P < 0.0001), litter (F2,46 = 28.78, P < 0.0001), and graminoid cover (F2,46 = 137.06, P < 0.0001) increased significantly with nutrient addition (). Hierochloe alpina, although rare in control plots, had the greatest abundance of all graminoids observed in fertilized plots (). Both lichen (F2,46 = 365.45, P < 0.0001) and evergreen cover (F2,46 = 122.84, P < 0.0001) decreased significantly with the addition of nutrients. Vaccinium vitis-idaea, K. procumbens, and R. tomentosum comprised the majority of the evergreen cover in CT plots, but were either significantly reduced or eliminated from fertilized plots (). Deciduous cover did not change significantly with fertilization; however, a shift from A. alpina to B. nana occurred when fertilizer was added (). Bare ground and animal activity also did not differ between the treatments (). Species richness decreased significantly in the fertilized plots with more species lost with additional years of nutrient addition (; F2,46 = 21.19, P < 0.0001). Community diversity also decreased significantly over time with fertilizer addition (; F2,46 = 42.79, P < 0.0001), suggesting evenness was slower to respond than richness.
FIGURE 2 Relative cover of the adult dry heath tundra community in 2008. Different letters indicate significant differences within a particular growth form.
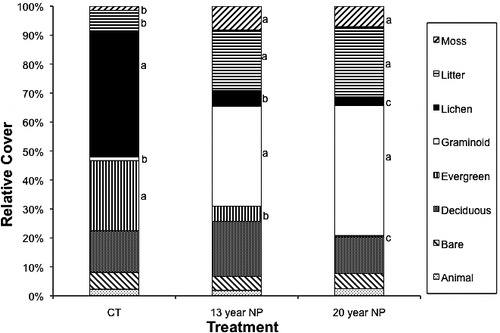
TABLE 1 Summary of plant species in dry heath tundra near the Arctic Long Term Ecological Research (LTER) plots at Toolik Lake, Alaska. Nomenclature follows CitationElven (2010). Quantities are mean ± SE per m2. Percent cover, seed bank, and seedling data were sampled within plots; seed rain was assessed outside plots to reflect dispersal (). CT is control; NP is N plus P.
TABLE 2 Species richness (S) and Diversity (H′) in dry heath tundra assessed in 2008. Results reported as mean ± SE at 1 m2 scale. Different superscript letters indicate significant differences between treatments. CT is control; NP is N plus P.
Flower and fruit data for the following four species are reported here: B. nana and H. alpina, because of their presence in the fertilized adult community, and R. tomentosum and K. procumbens, because of their high seed number in the seed rain experiment. Other species did not show a trend in dispersed seed numbers and in many cases had very low abundance as adults. There was a significant effect of fertilizer on flowering and fruiting of the four plant species (χ2 6 = 2413.60, P < 0.0001). Rhododendron tomentosum and K. procumbens, both evergreen dwarf shrubs, had greater numbers of flowers and fruits per m2 in the control plots than in the 13-year NP treatment plots (). Flowers and fruits of these species were absent in the 20-year NP treatment plots for both years because the adults were either rare or absent in the fertilized community. In contrast, B. nana and H. alpina had greater numbers of flowers and fruits in both 13- and 20-year NP treatments compared with CT.
TABLE 3 Number of fruit and flower totals per m2 in dry heath tundra over two years. In 2008, H. alpina flowers were counted differently and are not reported here. CT is control; NP is N plus P.
SEED RAIN
Seeds recovered from both CT and 20-year NP treatment plot trap areas were similar in overall species composition. Betula nana, K. procumbens, and R. tomentosum were the most abundant species and were collected in almost all trap areas in both treatments (). No significant differences were found between total seed numbers collected from control mats and those close to fertilized plots, although there was a suggestion of more total seeds and more B. nana seeds on mats adjacent to fertilized plots (F1,48 = 3.37, P = 0.07; and ).
FIGURE 3 Mean total number of seeds collected per 0.25 m2 in the seed rain experiment separated by species. NP-1 indicated the seed rain mat trap areas in 20 years of NP addition in Block 1 (See ). CT-1 indicates the seed rain mat trap areas in the row closest to the LTER plots (See ). Error bars represent ±1 SE. NP is N plus P treatments; CT is control treatments.
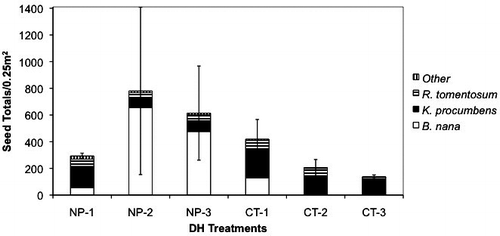
Within the 20-year NP treatments, the number of B. nana seeds varied greatly among seed rain mats in two of the three blocks (NP-2: max = 5835, min = 3; NP-3: max = 4161, min = 10), driving the high variance in total seeds among the blocks. In NP-1, seeds of the grass, Calamagrostis canadensis, made up a majority of the “other” category (173 seeds out of 184 total “other” seeds). C. canadensis seeds were also present in NP-2, but were fewer in number. H. alpina seeds were slightly greater in number in the 20-year NP treatment mats, but only accounted for a small percentage of the “other” category, and were not examined separately statistically. Seeds of the boreal species of fireweed, Chamerion angustifolium (previously Epilobium angustifolium), were present in low numbers in both CT and 20-year NP treatment seed traps ().
SEED BANK EXPERIMENT
In the soil seed bank study, more seedlings emerged overall from fertilized soil cores when compared with control cores collected the same year (; F1,22 = 5.48, P = 0.03). The germination of graminoid seeds was higher in soils after 13 years of fertilization. Ericaceous seeds germinated in both treatments at similar rates. Hierochloe alpina only germinated in the nutrient-amended soils, while no B. nana seeds germinated from any core.
SEEDLING SURVEY
Seedling numbers in both years were extremely low. One unidentified graminoid seedling was recorded from 24 plots in the 20-year NP treatment in 2007; no seedlings were encountered in any of the surveyed plots in 2008.
Discussion
DRAMATIC CHANGE IN COMMUNITY COMPOSITION
Our results support similar changes documented in a previous study in the dry heath tundra plots examining the effects of fertilization after 17 years (CitationGough et al., 2008). The shift from an evergreen dwarf shrub– and lichen-dominated community to one dominated by grasses follows those of other fertilization experiments in dry heath tundra (CitationPress et al., 1998; CitationCornelissen et al., 2001; CitationGrellmann, 2002; CitationRichardson et al., 2002), as well as the transition from heathlands to grasslands in northern Europe as a result of atmospheric N deposition (CitationRoelofs, 1986; CitationVitousek et al., 1997; CitationBerendse et al., 2001; CitationHeijmans et al., 2001). The increase in plant litter in nutrient amended plots is also consistent with other studies in dry heath (CitationPress et al., 1998; CitationGough et al., 2002, Citation2008), as well as in other tundra communities (CitationAerts and Berendse, 1988; CitationBret-Harte et al., 2008). The greater quantity of plant litter can potentially have multiple effects on the plant community, including shading of smaller stature plants (CitationFoster and Gross, 1998; CitationXiong and Nilsson, 1999), trapping moisture (CitationFowler, 1986), and providing nutrients upon decomposition (CitationFacelli and Pickett, 1991).
There was an overall positive effect of nutrient addition on fruits and flowers for graminoids and deciduous species, providing support for our first hypothesis (H1). Although we did not analyze species independently, it is apparent that flower and fruit total numbers of H. alpina and B. nana were greater under increased nutrient levels, most likely a result of greater relative abundance within the fertilized adult community. CitationGough et al. (2007) found a significant increase in H. alpina flowering measured on individual plants under fertilized conditions in dry heath compared to control conditions. Similarly, a trend toward greater flowering of B. nana due to increased nutrient availability at dry heath tundra was also observed on individual plants (Gough, unpublished data). The two evergreen species examined had greater productivity of flowers and fruits under ambient conditions, but fewer total flower and fruit numbers in fertilized plots, reflecting their lowered abundance in the amended adult communities. Thus relative abundance of adults was well correlated with flower and fruit abundance for the examined species.
SEED DISPERSAL REFLECTED ABUNDANCE IN THE ADULT COMMUNITY
Species composition of dispersed seeds was similar adjacent to nutrient-amended and control plots; we therefore reject the second hypothesis (H2) overall, but note the differences across species below. Because of the differences in the adult community between control and fertilized, we expected the species found on the seed traps near each treatment to reflect that adult community. However, ericaceous species that were abundant in control plots yet absent from fertilized plots as adults, particularly R. tomentosum and K. procumbens, had similar seed abundance in both treatments. In contrast, adjacent to the fertilized plots, we expected and found more seeds of H. alpina and C. canadensis (two grasses), B. nana, and the forb C. angustifolium, all of which are found in the nutrient-amended adult community. In the control plot seed traps closest to the fertilized treatments (CT-1, 35 m away), the number of B. nana seeds was higher than in those further away. Since B. nana is present but rare in dry heath tundra under current climate conditions (CitationGough et al., 2007), these results suggest that the greater abundance of B. nana seeds found in the control plots must have come from another source, such as the fertilized treatments. The small size and low weight of these seeds make them well suited for wind dispersal over long distances, as seen in a previous seed rain study in northern Sweden (CitationMolau and Larsson, 2000). Thus while there appears to be some localized dispersal from the adult plants within each treatment type, there also appears to be some longer distance dispersal, specifically from B. nana.
SEEDS IN THE SOIL SEED BANK REFLECT THE ADULT COMMUNITIES
Under lab conditions, the abundance of germinated seeds from the soil seed bank was greater from nutrient-amended than control plots. Species composition of the seed bank was similar between the treatment soils, therefore we reject the third hypothesis overall, but note how the different groups differed in response. Both control and nutrient-amended soil seed banks had graminoid and ericaceous species germinate. However, the presence of H. alpina seedlings in nutrient-amended soil seed banks suggests that this species is producing viable seeds that are locally dispersing into the landscape, which may be an indirect effect of the increased available nutrients. The lack of B. nana seed germination in either the control or fertilized seed bank was surprising given the results from the seed rain experiment. However, in a lab germination study, only 37% of B. nana seeds germinated under optimal control conditions (CitationMoulton, 2009), suggesting low seed viability for this species. Additional experiments, such as a seed sowing experiment in the field, would help tease apart the mechanisms driving the germination differences found here.
SEEDLING RECRUITMENT IN THE FIELD IS RARE
Numbers of seedlings were so low in both nutrient amended and control plots in dry heath tundra that the fourth hypothesis was rejected (H4). Although flower, fruit, and seed production was greater for graminoids and deciduous shrubs in the nutrient-amended plots, actual germination from seed and subsequent seedling establishment in the field is severely limited. Seedling counts from a 2006 biomass harvest from other plots in this dry heath tundra support these findings: one E. nigrum in the control and one H. alpina in the fertilized, out of twelve 0.04 m2 plots for each treatment (Gough, unpublished data). These results may stem from multiple causes. Viability of seeds likely initially limits germination for at least some species, as noted above for B. nana. Seedling establishment may be further limited by available space and associated resources in the landscape either by increased litter production or adult plant biomass. A seed sowing study in Finland found that fertilization negatively affected rates of colonization, although the removal of biomass in nutrient amended plots allowed for seeds to establish and colonize (CitationEskelinen, 2010; also see CitationGough, 2006). Therefore, seedling establishment despite more seeds being available appears to be limited by multiple factors including initial seed viability, presence of litter and available space in the adult community.
ROLE OF SEEDS MAY CHANGE IN WARMED CLIMATE
Based on the findings of this study, we conclude that additional soil nutrients shift the composition and availability of seeds in dry heath tundra, but that a severe bottleneck on new recruitment in the field is present after more than a decade of experimental fertilization. We believe, however, that this bottleneck was not as important in the first few years after the experiments began, because at that time, litter and increased plant growth had not yet limited the space needed for germination. We speculate that the shift in the adult community from one dominated by lichens and evergreen dwarf shrub species to that of a grassland included seedling recruitment for the grass H. alpina. This species consists of a few, widely scattered individuals under control conditions, and increases in both size and density of individuals when fertilized. The other species that appears to benefit from increased nutrients, B. nana, is able to do so almost entirely vegetatively, as it has in other Alaskan tundra communities (e.g., CitationBret-Harte et al., 2001).
As currently immobilized soil nutrients in tundra communities become more readily available as climate change continues, species that are capable of utilizing these nutrients could potentially outcompete the current dominant plant species, as shown in these manipulative experiments. Our results suggest that species which are better able to take advantage of increased nutrients may vegetatively expand into the landscape and produce more flowers, and subsequently seeds, potentially allowing a further advantage by increasing population-level genetic diversity. This process will depend on several of the factors discussed above, including seed viability and available space. Disturbances such as the removal of litter or adult vegetation by herbivores or soil frost heaving may be required to open up space where germination can occur. Species not currently found in dry heath tundra, such as C. angustifolium, could potentially immigrate by seed and flourish under increased soil nutrients, driving additional community changes. Chamerion angustifolium is wind dispersed and has been recorded in dry heath plots that have received nutrient addition for almost 20 years, as well as in other fertilized plots in different, nearby tundra communities (Gough, unpublished data). This species does well in disturbed areas such as the gravel pad at Toolik Field Station and along the Dalton Highway, but is not currently found in undisturbed tundra. As environmental changes associated with global warming continue in the Arctic, the role of sexual reproduction is likely to shift in complex and somewhat unpredictable ways, differing substantially among individual species.
Acknowledgments
This research was conducted under National Science Foundation grant OPP-0435827 to L. Gough. We thank C. Rogers and D. Kirkland for their excellent assistance with fieldwork, and the Arctic LTER for logistical support. We thank P. Wookey and two anonymous reviewers for comments on an earlier draft of the manuscript.
Related Research Data
References Cited
- Aerts, R. and F. Berendse . 1988. The effects of increased nitrogen availability on vegetation dynamics in wet heathlands. Plant Ecology 76:63–69.
- Aerts, R. , J. H. C. Cornelissen , and E. Dorrepaal . 2006. Plant performance in a warmer world: general responses of plants from cold, northern biomes and the importance of winter and spring events. Plant Ecology 182:65–77.
- Alatalo, J. M. and O. Totland . 1997. Response of simulated climatic change in an alpine and subarctic pollen-risk strategist, Silene acaulis . Global Change Biology 3:74–79.
- Alsos, I. G. , S. Spjelkavik , and T. Engelskjon . 2003. Seed bank size and composition of Betula nana , Vaccinium uliginosum , and Campanula rotundifolia habitats in Svalbard and northern Norway. Canadian Journal of Botany 81:220–231.
- Alsos, I. G. , P. B. Eidesen , D. Ehrich , I. Skrede , K. Westergaard , G. H. Jacobsen , J. Y. Landvik , P. Taberlet , and C. Brochmann . 2007. Frequent long-distance plant colonization in the changing Arctic. Science 316:1606–1608.
- Babb, T. A. and T. W. A. Whitfield . 1977. Mineral nutrient cycling and limitation of plant growth in the Truelove lowland ecosystem. In Bliss, L. C. (ed.). Truelove Lowland, Devon Island, Canada: a High Arctic Ecosystem. Edmonton University of Alberta Press. 589–606.
- Baskin, C. C. and J. M. Baskin . 2001. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. San Diego Academic Press.
- Bell, K. L. and L. C. Bliss . 1980. Plant reproduction in a high arctic environment. Arctic and Alpine Research 12:1–10.
- Berendse, F. , N. van Vreemen , H. Ryden , A. Buttler , M. Heijmans , M. R. Hoosbeek , J. A. Lee , E. Mitchell , T. Saarinen , H. Vasander , and B. Wallen . 2001. Raised atmospheric CO2 levels and increased nitrogen deposition cause shift in plant species composition and production on bog vegetation in the Netherlands. Journal of Ecology 89:268–279.
- Billings, W. D. 1987. Constraints to plant growth, reproduction and establishment in arctic environments. Arctic and Alpine Research 19:357–357.
- Billings, W. D. and H. A. Mooney . 1968. The ecology of arctic and alpine plants. Biological Review 43:481–529.
- Bliss, L. C. 1962. Adaptations of arctic and alpine plants to environmental conditions. Arctic 15:117–144.
- Bliss, L. C. and N. V. Matveyeva . 1992. Circumpolar arctic vegetation. In Chapin III, F. S. , R. L. Jefferies , J. F. Reynolds , G. R. Shaver , and J. Svoboda . (eds.). Arctic Ecosystems in a Changing Climate: an Ecophysiological Perspective. San Diego Academic Press. 59–90.
- Bret-Harte, M. S. , G. R. Shaver , J. P. Zoerner , J. F. Johnstone , J. L. Wagner , A. S. Chavez , R. F. Gunkelman , S. C. Lippert , and J. A. Laundre . 2001. Developmental plasticity allows Betula nana to dominate tundra subjected to an alternative environment. Ecology 82:18–32.
- Bret-Harte, M. S. , M. C. Mack , G. R. Goldsmith , D. B. Sloan , J. DeMarco , G. R. Shaver , P. M. Ray , Z. Biesinger , and F. S. Chapin III . 2008. Plant functional types do not predict biomass responses to removal and fertilization in Alaskan tussock tundra. Journal of Ecology 96:713–726.
- Chang, E. R. , R. L. Jefferies , and T. J. Carleton . 2001. Relation between vegetation and soil seed banks in an arctic coastal marsh. Journal of Ecology 89:367–384.
- Chapin III, F. S. , R. S. Barsdate , and D. Barel . 1978. Phosphorus cycling in Alaskan coastal tundra: a hypothesis for the regulation of nutrient cycling. Oikos 31:189–199.
- Chapin III, F. S. , A. J. Bloom , C. B. Field , and R. H. Waring . 1987. Plant responses to multiple environmental factors. BioScience 37:49–57.
- Chapin III, F. S. , G. R. Shaver , A. E. Giblin , K. J. Nadelhoffer , and J. A. Laundre . 1995. Responses of arctic tundra to experimental and observational changes in climate. Ecology 76:694–711.
- Chester, A. L. and G. R. Shaver . 1982. Reproductive effort in cotton grass tussock tundra. Holarctic Ecology 5:200–206.
- Clein, J. S. and J. P. Schimmel . 1995. Microbial activity of tundra and taiga soils at sub-zero temperatures. Soil Biology and Biochemisty 27:1231–1234.
- Cooper, E. J. , I. G. Alsos , D. Hagen , F. M. Smith , S. J. Coulson , and I. D. Hodkinson . 2004. Plant recruitment in the High Arctic. Journal of Vegetative Science 15:115–224.
- Cornelissen, J. H. C. , T. V. Callaghan , J. M. Alatalo , A. Michelsen , E. Graglia , A. E. Hartley , D. S. Hik , S. E. Hobbie , M. C. Press , C. H. Robinson , G. H. R. Henry , F. S. Chapin III , U. Molau , C. Neill , J. A. Lee , J. M. Melillo , B. Sveinbjornsson , and R. Aerts . 2001. Global change and arctic ecosystems: is lichen decline a function of increases in vascular plant biomass? Journal of Ecology 89:984–994.
- Diemer, M. and S. Prock . 1993. Establishment of alpine seed bank side in two central European and one Scandinavian subarctic plant communities. Alpine and Arctic Research 25:194–200.
- Ellis, S. 1980. An investigation of weathering in some arctic-alpine soils on the northeastern flank of Okaskolten, northern Norway. Journal of Soil Science 31:371–385.
- Elven, R. (ed.). 2010. Checklist of the Panarctic Flora (PAF) Vascular Plants Version 2010 provided by Murray, D. A., University of Alaska, Fairbanks. <http://www.binran.ru/infsys/paflist/index.htm>.
- Eskelinen, A. 2010. Resident functional composition mediates the impacts of nutrient enrichment and neighbour removal on plant immigration rates. Journal of Ecology 98:540–550.
- Eskelinen, A. and R. Virtanen . 2005. Local and regional processes in low-productive mountain plant communities: the roles of seed and microsite limitation in relation to grazing. Oikos 110:360–368.
- Facelli, J. M. and S. T. A. Pickett . 1991. Plant litter: its dynamics and effects on plant community structure. The Botanical Review 57:1–32.
- Foster, B. L. and K. L. Gross . 1998. Species richness in a successional grassland: effects of nitrogen enrichment and plant litter. Ecology 79:2593–2602.
- Fowler, N. L. 1986. Microsite requirements for germination and establishment of three grass species. American Midland Naturalist 115:131–145.
- Fox, J. F. 1983. Germinable seed banks of interior Alaskan tundra. Arctic and Alpine Research 15:405–411.
- Gartner, B. L. , F. S. Chapin , and G. R. Shaver . 1986. Reproduction of Eriophorum vagninatum Eriophorum vagninatum by seed in Alaskan tussock tundra. Journal of Ecology 74:1–18.
- Gough, L. 2006. Neighbor effects on germination, survival, and growth in two arctic tundra plant communities. Ecography 29:44–56.
- Gough, L. , P. A. Wookey , and G. R. Shaver . 2002. Dry heath tundra responses to long-term nutrient and light manipulation. Arctic, Antarctic, and Alpine Research 34:211–218.
- Gough, L. , E. A. Ramsey , and D. R. Johnson . 2007. Plant-herbivore interactions in Alaskan arctic tundra change with soil nutrient availability. Oikos 116:407–418.
- Gough, L. , K. Shrestha , D. R. Johnson , and B. Moon . 2008. Long-term mammalian herbivory and nutrient addition alters lichen community structure in Alaskan dry heath tundra. Arctic, Antarctic, and Alpine Research 40:65–73.
- Grellmann, D. 2002. Plant responses to fertilization and exclusion of grazers on an arctic tundra heath. Oikos 98:190–204.
- Heijmans, M. M. P. D. , F. Berendse , W. J. Arp , A. K. Masselink , H. Klees , W. de Visser , and N. van Breeman . 2001. Effects of elevated carbon dioxide and increased nitrogen deposition on bog vegetation in the Netherlands. Journal of Ecology 89:268–279.
- Jonsdottir, I. S. , T. V. Callaghan , and A. D. Headley . 1996. Resource dynamics within arctic clonal plants. Ecological Bulletins 45:53–64.
- Klanderud, K. 2005. Climate change effects on species interactions in an alpine plant community. Journal of Ecology 93:127–137.
- Lindgren, A. , O. Eriksson , and J. Moen . 2007. The impact of disturbance and seed availability on germination of alpine vegetation in the Scandinavian mountains. Arctic, Antarctic, and Alpine Research 39:449–454.
- McGraw, J. B. 1980. Seed bank size and distribution of seeds in cottongrass tussock tundra, Eagle Creek, Alaska. Canadian Journal of Botany 58:1608–1611.
- McGraw, J. B. and M. C. Vavrek . 1989. The role of buried viable seeds in arctic and alpine plant communities. In Leck, M. A. , V. T. Parker , and R. L. Simpson . (eds.). Ecology of Soil Seed Banks. San Diego Academic Press. 91–105.
- Mikan, C. J. , J. P. Schimel , and A. P. Doyle . 2002. Temperature controls of microbial respiration in arctic tundra soils above and below freezing. Soil Biology and Biogeochemistry 34:1785–1795.
- Molau, U. and E. L. Larsson . 2000. Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland. Canadian Journal of Botany 78:728–747.
- Molau, U. and P. Molgaard . (eds.). 1996. ITEX Manual. Copenhagen Danish Polar Centre.
- Moulton, C. 2009. How Soil Nutrient Availability Affects Plant Sexual Reproduction and Seedling Recruitment in Alaskan Dry Heath Tundra: Implications for Response to Climate Change. MS thesis, University of Texas at Arlington.
- Press, M. C. , J. A. Potter , M. J. W. Burke , T. V. Callaghan , and J. A. Lee . 1998. Responses of a subarctic dwarf shrub heath community to simulated environmental change. Journal of Ecology 86:315–327.
- Richardson, S. J. , M. C. Press , A. N. Parsons , and S. E. Hartley . 2002. How do nutrients and warming impact on plant communities and their insect herbivores? A 9-year study from a sub-arctic heath. Journal of Ecology 90:544–556.
- Roelofs, J. G. M. 1986. The effect of airborne sulphur and nitrogen deposition on aquatic and heathland vegetation. Cellular and Molecular Life Science 42:372–377.
- Rustad, L. E. , J. L. Campbell , G. M. Marion , R. J. Norby , M. J. Mitchell , A. E. Hartley , J. H. C. Cornelissen , and J. Gurevitch . GCTE-NEWS 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126:543–562.
- Shaver, G. R. and F. S. Chapin . 1986. Effects of fertilizer on production and biomass of tussock tundra, Alaska, U.S.A. Arctic and Alpine Research 18:261–268.
- Shaver, G. R. and F. S. Chapin . 1991. Production: biomass relationships and element cycling in contrasting arctic vegetation types. Ecological Monographs 61:1–34.
- Shaver, G. R. and S. Jonasson . 1999. Response of arctic ecosystems to climate change: results of long term field experiments in Sweden and Alaska. Polar Research 18:245–252.
- Vitousek, P. M. , J. D. Aber , R. W. Howarth , G. E. Likens , P. A. Matson , D. W. Schinder , W. H. Schlesinger , and D. G. Tilman . 1997. Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications 7:737–750.
- Xiong, S. and C. Nilsson . 1999. The effects of plant litter on vegetation: a meta-analysis. Journal of Ecology 87:984–994.