
Abstract
Climate-based models predict global warming will cause mountain plants to migrate upward with local extirpation of species currently restricted to the highest elevations. Alpine monitoring studies have generally documented changes in alpine plant distributions consistent with these predictions; however, relatively few such studies have been reported from North America. I estimated canopy cover in 71 permanent microplots at two alpine moist-turf sites in Glacier National Park, Montana, U.S.A., three times between 1988 and 2011. Mean annual and summer temperatures were approximately 0.6 and 0.7 °C higher than the previous four decades, respectively. Plants more restricted to high elevations declined more than species with a broader elevational amplitude during this time. Dicots were more likely to have declined than monocots. These data support predictions and provide information that may help refine climate-envelope models.
Introduction
There is widespread agreement that increasing levels of atmospheric carbon dioxide and other greenhouse gases will result in global warming during the next 50 years (CitationHenderson-Sellers, 1990; CitationIPCC, 2007). Less rapid changes of comparable amplitude in the past altered the ranges of many species (CitationGraham and Grimm, 1990; CitationWebb, 1992; CitationJackson and Overpeck, 2000), and there is evidence that biota are currently shifting their geographic ranges upward in elevation or latitude (CitationRoot et al., 2003). High-mountain ecosystems are thought to be particularly vulnerable to the effects of warming (CitationGrabherr et al., 1995; CitationCannone et al., 2007; CitationNagy and Grabherr, 2009; CitationEngler et al., 2011), especially at higher latitudes (CitationKörner, 1999). In general, climate envelope models that employ the relation between current climate variables and present-day species distributions predict that species will migrate upward with local extirpation of species currently restricted to the highest elevations (CitationGottfried et al., 1998; CitationWalker et al., 2006). However, there are reasons to believe that some alpine species will persist by shifting microsites rather than elevational zones (CitationScherrer and Körner, 2010).
Many monitoring studies have documented changes in alpine plant distributions consistent with climate-warming predictions. The majority of these studies are based on presence/absence data in which a study area is surveyed for the presence of species at one point in time and then resurveyed at a subsequent time (CitationGrabherr et al., 1994; CitationKlanderud and Birks, 2003; CitationWalther et al., 2005; CitationParolo and Rossi, 2008; CitationErschbamer et al., 2012). In most cases, researchers have documented increases in species richness resulting from upward migration of lower-elevation species but have only infrequently found a loss of high-elevation species. Presence/absence measurements can be more repeatable than canopy cover estimates (CitationElzinga et al., 2001), but change is more likely to be detected earlier by accurate measures of canopy cover. Canopy-cover monitoring of alpine vegetation was begun on a large scale in Europe in 1994. One ten-year study in the Austrian Alps that monitored plant cover in permanent plots found an increase in the cover of “low-alpine” (= alpine) species but also documented a decline in “high-alpine” (i.e., nival and subnival) species (CitationPauli et al., 2007).
Several studies have indicated that a species' response to climatic warming will often be associated with its functional or life-form group (CitationArft et al., 1999). For example, experimental warming studies suggest that shrubs will increase at lower alpine zones in Europe (CitationCannone et al., 2007), Alaska (CitationEpstein et al., 2000; CitationWalker et al., 2006), and Asia (CitationKlein et al., 2007) at the expense of graminoids. Unfortunately, results of long-term monitoring studies have rarely been examined for life-form correlations. Although a good deal of experimental work has been done in North America, there have been relatively few published, long-term monitoring studies of alpine vegetation (CitationWalker et al., 1994). Here I report on a 24-year monitoring study of moist alpine turf in Glacier National Park in the Rocky Mountains of western North America. Canopy cover data are analyzed to determine if there is a signal similar to those found in Europe and which species and life-form groups have increased or declined.
Study Sites
Permanent monitoring transects were established at two locations above timberline in Glacier National Park in the Logan Pass area: Lunch Creek and Mount Reynolds (). Both sites are on gentle south-facing slopes, subirrigated throughout at least part of the growing season by snowmelt originating upslope. These unusual sites mimic arctic tundra by having a long growing season due to the south exposure but perennially cold and moist soil. These moist turf sites harbor several arctic plant species that are at the southern margin of their range (CitationLesica and Steele, 1996; CitationLesica and McCune, 2004). Parent materials of both sites are limestones and argillites of the Precambrian Belt Series, and both sites have coarse to fine colluvium overlain by highly organic soil of varying thickness (sometimes absent) that is saturated or nearly so for much of the growing season. The Mount Reynolds site is on the south-facing side of Mount Reynolds, 4 km SSW of the Logan Pass visitor center at an elevation of 2285 m. There is a late-season snowfield at the base of the near-vertical slope ∼50 m above the transects. The Lunch Creek site is in Lunch Creek Cirque in the valley created by Polock Mountain and Piegan Mountain, 2 km NNE of the Logan Pass visitor center at an elevation of 2165 m. There are permanent snowfields ∼150 m farther up the cirque.
Climate data are available for several sites at both high and low elevations in Glacier National Park, but only discontinuous data are available from high-elevation recording stations. Annual precipitation at Sperry Glacier, at 2440 m and 5–10 km south of the study sites exceeds 250 cm. Precipitation occurs as rainfall during June through September and as snowfall the remainder of the year (CitationFinklin, 1986). Summer climate is cool and moist; during the 1959 growing season, average daily temperature and relative humidity were 10 °C and 74% (CitationChoate and Habeck, 1967). Mean annual daily high and low temperatures were estimated to be 6.6 and -4.9 °C, respectively, between 1980 and 2003, with a mean annual precipitation of 159 cm (CitationMalanson et al., 2012). Continuous long-term (i.e., >50 yr) climatic data have been collected only at West Glacier at 975 m. Climate at West Glacier over the two-decade course of the study was warmer but not wetter than the previous four decades (). Snowpack across the Rocky Mountains has been melting earlier over the past three decades due to warmer spring temperatures (CitationPederson et al., 2011), suggesting that climate trends at West Glacier mirror those at higher elevations.
FIGURE 1. Location of Lunch Creek and Mount Reynolds study areas in Glacier National Park, Montana, U.S.A. Aerial photograph was taken in late summer, 1990. Note the snowfields just above the two sites.
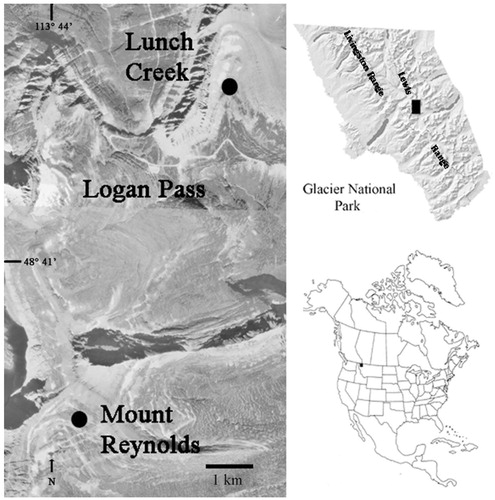
TABLE 1 Mean annual and summer (June through September) temperature and mean annual precipitation at West Glacier.
Methods
FIELD METHODS
Two (Mount Reynolds) or three (Lunch Creek) permanent transects were established at the study sites in 1988. Transects were subjectively located to pass through the middle of longitudinal strips of moist turf habitat running perpendicular to the slope. I estimated canopy cover for all vascular plant species in 0.1 m2 plots (50 × 20 cm) placed 1 m apart along the transect lines. There was a total of 35 and 36 sample plots at Mount Reynolds and Lunch Creek sites, respectively. Canopy cover (CitationDaubenmire, 1959) was estimated to the nearest 5%, but estimates less than 2% cover were recorded as 1%. Canopy cover of vascular plants was recorded at Mount Reynolds in 1988, 2000, 2011, and at Lunch Creek in 1988, 2000, and 2010. There is a subjective aspect to canopy cover estimation (CitationElzinga et al., 2001) that can introduce unwanted error when data collected by different observers are compared. All canopy-cover estimates were made by the author for all three recording periods. Although the estimates may not be perfectly accurate, the change between estimates at different times by the same observer will usually result in acceptable precision (CitationElzinga et al., 2001). Vascular plant nomenclature follows Lesica (Citation2002).
DATA ANALYSIS
I used repeated-measures analysis of variance (ANOVA) followed by a linear polynomial contrast test to ascertain the presence of trend for each of the common species (recorded in >10 sample plots). Plots were considered independent samples. The data met requirements for normality of residuals and homogeneity of variance. Site was used as a factor in the ANOVA model if a species occurred at both Lunch Creek and Mount Reynolds, and sites were analyzed separately if there was a significant Year × Site interaction (P ≤ 0.05).
For each species, I used the slope of the best-fit linear regression model of canopy cover across the three years of the study (see previous section) divided by the mean canopy cover for the three years as a measure of trend. This adjusted regression slope (hereafter “trend index”) allowed me to compare trend for common species with those that were less common. I used a recent floristic treatment for Montana (CitationLesica, 2012) to assign an “alpine index number” to each of the 33 common species found at the study sites. This index number reflects the degree to which each species is restricted to high elevations in Montana. Species that occur only above treeline were assigned an alpine index of (4), both alpine and subalpine (3), montane through alpine (2), and valleys through alpine (1). I used linear regression analysis to assess the relationship between the trend index and the alpine index number, that is, whether species restricted to high elevations declined more or increased less than less restricted species. I used a two-sample t-test to determine whether adjusted regression slope (trend) differed between monocots and dicots over the course of the study.
Results
Vegetation at the Lunch Creek and Mount Reynolds sites was similar. The plant community at Lunch Creek was dominated by the dwarf shrubs Salix reticulata (mean canopy cover across all 3 years = 11%), and Dryas octopetala (9%). Carex scirpoidea (18%) and C. capillaris (7%) were the dominant graminoids. The most common forbs were Solidago multiradiata (7%), Hedysarum sulphurescens (7%), Polygonum viviparum (6%), Silene acaulis (5%), and Anemone parviflora (3%). Tundra at the Mount Reynolds site was also dominated by Dryas octopetala (26%) and Salix nivalis (8%). Carex scirpoidea (18%), Kobresia simpliciuscula (17%), and C. rupestris (5%) were the dominant graminoids. The most common forbs were Polygonum viviparum (9%), Silene acaulis (3%), and Solidago multiradiata (3%). The cover of bare rock was 5% at the Lunch Creek site and 3% at Mount Reynolds.
A total of 63 vascular plant species were recorded at the Lunch Creek and Mount Reynolds sites over the course of the study. Of these, 33 species were recorded in more than ten plots (). Canopy cover of Carex scirpoidea, Potentilla fruticosa, and Sedum rosea (at Lunch Creek) showed statistically significant increases over the course of the study, while cover of Gentianella propinqua, Kobresia simpliciuscula (at Mount Reynolds), Polygonum viviparum, Salix arctica, S. reticulata, and Sedum rosea (at Mount Reynolds) declined significantly (). All six of the significantly declining species, but only one of the three increasing species, are restricted to alpine and subalpine habitats in Montana ().
There was a marginally significant negative association between alpine index and trend index indicating a tendency for species restricted to higher elevations (greater alpine index) to have declined more in canopy cover (more negative trend index) than species with greater ecological amplitude over the two decades of the study (R 2 = 0.09, P = 0.089, ). In addition, canopy cover of the 21 dicots declined at a greater rate compared to the 11 monocots (t = 2.1, P = 0.045, ). Only three species of woody plants were common enough for analysis. All three were in the genus Salix (). The two dwarf shrubs (S. arctica and S. reticulata) both declined, while the taller S. vestita remained stable.
Discussion
There was a declining trend in the canopy cover of species restricted to high elevations compared to species with a broader elevational amplitude over the past two decades in the moist-turf study sites in Glacier National Park. During this same period, mean annual daily and summer daily temperatures increased 0.6 and 0.7 °C, respectively, at the West Glacier recording station compared to the previous 40 years, while annual precipitation was virtually unchanged. Results from this study are descriptive in nature rather than experimental, so I cannot rigorously infer that climatic warming caused the observed changes. However, my results are consistent with forecasts for alpine areas in Europe and elsewhere (CitationGottfried et al., 2002; CitationNagy and Grabherr, 2009).
Although the observed changes in species canopy cover were associated with increased temperature rather than declines in precipitation, I believe that changes in snowpack mediated by increased temperatures are the most likely cause of the observed vegetation shifts at my study sites because the duration of snowmelt and disposition of meltwater strongly affect the pattern of alpine plant communities (CitationWalker et al., 1993; CitationMalanson et al., 2012). Tundra plants often have detrimentally high base metabolic rates in abnormally warm environments (CitationBillings and Mooney, 1968; CitationSpomer and Salisbury, 1968; CitationMcNulty et al., 1988; CitationKörner, 1999, p. 192), and there is evidence that species from cold, moist sites have less ability to adapt to warming temperatures (CitationCooper, 2004). Moist tundra vegetation at the two study sites is subirrigated by cold meltwater, which presumably keeps the roots and, to some extent, the above-ground vegetation cooler than ambient temperatures. Warmer temperatures in spring and summer in Glacier National Park have resulted in a diminution of summer snowpack since the mid-1980s (Pederson et al., 2010; P. Lesica, observation), presumably leading to drier and warmer soil downslope. I suggest that the warmer and drier soil of these sites later in the growing season is causing a loss of species adapted to cold, moist habitats in favor of those adapted to warmer, drier, lower-elevation environments.
Studies of plant distributions in alpine sites have frequently documented upward migration of low-elevation species but less commonly declines in high-elevation species (CitationGrabherr et al., 1995; CitationCannone et al., 2007; CitationNagy and Grabherr, 2009; CitationEngler et al., 2011), while my study suggests both processes are occurring. Most alpine dominants are long-lived perennials, so detecting a decline of species will take a longer time than detecting immigration of lower-elevation plants (CitationGrabherr, 2003). Furthermore, many monitoring studies have been conducted on exposed alpine summits with drier soils and hydrology that is less affected by persistent snowfields. Alpine plants in these environments are expected to be less sensitive to drying, so the main effect on the plants will be a longer growing season and warmer ambient temperatures that allow immigration by lower-elevation species. Most alpine monitoring has been done using presence/absence data from large plots or zones (CitationGrabherr et al., 1994; CitationKlanderud and Birks, 2003; CitationWalther et al., 2005; CitationParolo and Rossi, 2008; CitationErschbamer et al., 2012). Monitoring canopy cover in numerous small sample plots will usually be more sensitive than monitoring presence/absence over large areas because extirpation in a large area will take more time than a diminution of cover. Pauli et al. (Citation2007), who also measured canopy cover in the Austrian Alps, documented an increase in low-elevation species but also a decline in high-elevation species, similar to my results.
TABLE 2 Common species (# of plots > 10) at Lunch Creek and Mount Reynolds sites and their mean canopy cover over the course of the study, trend index (see Methods), alpine index (see Methods), and significant change as determined by repeated-measures ANOVA followed by a linear polynomial contrast test.
Some researchers have suggested that the response of arctic species to climatic warming will depend on the plant's growth form (i.e., functional group) (CitationChapin and Shaver, 1996; CitationChapin et al., 1996; CitationArft et al., 1999; CitationYang et al., 2011; but see CitationKlanderud, 2008). Common alpine graminoids increased during the warmest decade on record in the Austrian Alps (CitationBahn and Körner, 2003), and Walker et al. (Citation2006) found that arctic graminoids increased under experimental warming. My study suggests that the same may be true for alpine plant communities. Populations of graminoids and other monocots (5 of 11 species in the Cyperaceae) tended to increase or remain stable more than broad-leaved forbs and shrubs over the course of the study. Greater resilience to drier conditions by monocots compared to dicots may be the result of anatomical differences such as structural root mass or size of conductive tissues (CitationDickison, 2000; CitationCarlquist, 2012) or differences in physiology (CitationEdwards et al., 2007). Similar observations were made on Great Plains grasslands during the great drought of the 1930s. Weaver (Citation1968) reported that drought caused only a shift in the dominance of the grass species, but resulted in the disappearance of many species of forbs (CitationAlbertson and Weaver, 1944).
Climate-change experimental research on arctic plants also suggests that shrubs will increase with a warming climate, but dwarf (i.e., prostrate) shrubs are likely to decline (CitationKlanderud and Totland, 2005; CitationWipf et al., 2005; CitationWalker et al., 2006; but see CitationBahn and Körner, 2003). Although the sample size is small, my results are consistent with these reports. The two prostrate willow species (Salix arctica, S. reticulata) declined significantly, but there was no detectable change in the cover of the taller shrub, Salix vestita (), which grows 20–100 cm tall in Montana (CitationLesica, 2012).
Conclusion
The results of my study support the general prediction for alpine vegetation that lower-elevation species will expand their range, while species restricted to the highest elevations will decline in abundance with climatic warming. Furthermore, it appears that dicots may be more prone to decline than monocots. Finally, four of the common recorded species, Pinguicula vulgaris, Tofieldia pusilla, Kobresia simpliciuscula, and Gentianella propinqua, are at or near the southern edge of their arctic-alpine distribution. Of these, the latter two (50%) declined compared to only 18% of the common species overall, suggesting that peripheral populations may also be at greater risk from climate change (CitationLesica and McCune, 2004). These results may help make climate-based models more biologically realistic and accurate in predicting the fate of alpine plants on a warming planet.
Acknowledgments
Kathy Ahlenslager, Teri Allendorf, Jennifer Asebrook, Levi Besaw, Tara Carolin, Ann DeBolt, Pete Del Zotto, Jennifer Hintz, Laurie Kurth, Bruce McCune, and Diane Taylor assisted in the field. Rober Pal provided helpful comments on the manuscript. This study was funded by the U.S. National Park Service.
Related Research Data
References Cited
- Albertson, F. W. , and Weaver, J. E. , 1944: Nature and degree of recovery of grassland from the great drought of 1933 to 1940. Ecological Monographs, 14: 393–479.
- Arft, A. M. , Walker, M. D. , Gurevitch, J. , Alatalo, J. M. , Bret-Harte, M. S. , Dale, M. , Diemer, M. , Gugerli, F. , Henry, H. R. , Jones, M. H. , Hollister, R. D. , Jónsdóttir, I. S. , Laine, K. , Lévesque, E. , Marion, G. M. , Molau, U. , Mogaard, P. , Nordenhäll, U. , Raszhivin, V. , Robinson, C. H. , Starr, G. , Stenström, A. , Stenström, M. , Totland, Ø . , Turner, P. L. , Walker, L. J. , Webber, P. J. , Welker, J. M. , and Wookey, P. A. , 1999: Responses of tundra plants to experimental warming: meta-analysis of the international tundra experiment. Ecological Monographs, 69: 491–511.
- Bahn, M. , and Körner, C. , 2003: Recent increases in summit flora caused by warming in the Alps. In Nagy, L. , Grabherr, G. , Körner, C. , and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer-Verlag, 437–441.
- Billings, W. D. , and Mooney, H. A. , 1968: The ecology of Arctic and alpine plants. Biological Review, 43: 481–529.
- Cannone, N. , Sgorbayi, S. , and Guglielmin, M. , 2007: Unexpected impacts of climate change on alpine vegetation. Frontiers in Ecology and Environment, 5: 360–364.
- Carlquist, S. , 2012: Monocot xylem revisited: new information, new paradigms. Botanical Review, 78: 87–153.
- Chapin, F. S. , and Shaver, G. R. , 1996: Physiological and growth responses of Arctic plants to a field experiment simulating climatic change. Ecology, 77: 822–840.
- Chapin, F. S. , Bret-Harte, M. S. , Hobbie, S. E. , and Zhong, H. , 1996: Plant functional types as predictors of transient responses of Arctic vegetation to global change. Journal of Vegetation Science, 7: 347–358.
- Choate, C. M. , and Habeck, J. R. , 1967: Alpine plant communities at Logan Pass, Glacier National Park. Proceedings of the Montana Academy of Sciences, 27: 36–54.
- Cooper, E. J. , 2004: Out of sight, out of mind: thermal acclimation of root respiration in Arctic Ranunculus. Arctic, Antarctic, and Alpine Research, 36: 308–313.
- Daubenmire, R. , 1959: A canopy-coverage method of vegetation analysis. Northwest Science, 33: 43–64.
- Dickison, W. G. , 2000: Integrative Plant Anatomy. San Diego: Academic Press, 533 pp.
- Edwards, E. J. , Still, C. J. , and Donoghue, M. J. , 2007: The relevance of phylogeny to studies of global change. Trends in Ecology and Evolution, 22: 243–249.
- Elzinga, C. L. , Salzer, D. W. , Willoughby, J. W. , and Gibbs, J. P. , 2001: Monitoring Plant and Animal Populations. Oxford: Blackwell Science, 360 pp.
- Engler, R. , Randin, C. F. , Thuiller, W. , Dullinger, S. , Zimmermann, N. E. , Araújo, M. B. , Pearman, P. B. , LeLay, G. , Piedallu, C. , Albert, C. H. , Choller, C. , Coldea, G. , DeLamo, X. , Dirnböck, T. , Gégout, J. , Gómez-García, D. , Grytnes, J. , Heegaard, E. , Hoistad, F. , Nogués-Bravo, D. , Normand, S. , Purcarr, M. , Ebastià, M. , Stanisci, A. , Theurillat, J. , Trivedi, M. R. , Vittoz, P. , and Guisan, A. , 2011: 21st century climate change threatens mountain flora unequally across Europe. Global Change Biology, 17: 2330–2341.
- Epstein, H. E. , Walker, M. D. , Chapin, F. S. , and Starfield, A. M. , 2000: A transient, nutrient-based model of Arctic plant community response to climatic warming. Ecological Applications, 10: 824–841.
- Erschbamer, B. , Kiebacher, T. , Mallaun, M. , and Unterluggauer, P. , 2012: Short-term signals of climate change along an altitudinal gradient in the South Alps. Plant Ecology, 202: 79–89.
- Finklin, A. I. , 1986: A climatic handbook for Glacier National Park with data for Waterton Lakes National Park. Ogden, Utah: USDA Forest Service Intermountain Research Station General Technical Report INT-204.
- Gottfried, M. , Pauli, H. , and Grabherr, G. , 1998: Prediction of vegetation patterns at the limits of plant life: a new view of the alpine-nival ecotone. Arctic and Alpine Research, 30: 207–221.
- Gottfried, M. , Pauli, H. , Reiter, K. , and Grabherr, G. , 2002: Potential effects of climate change on alpine and nival plants in the Alps. In Körner, C. , and Spehn, E. M. (eds.), Mountain Biodiversity-A Global Assessment. London: Parthenon Publishing, 213–223.
- Grabherr, G. , 2003: Alpine vegetation dynamics and climate change-A synthesis of long-term studies and observations. In Nagy, L. , Grabherr, G. , Körner, C. , and Thompson, D. B. A. (eds.), Alpine Biodiversity in Europe. Berlin: Springer-Verlag, 399–409.
- Grabherr, G. , Gottfired, M. , and Pauli, H. , 1994: Climate effects on mountain plants. Nature, 369: 448.
- Grabherr, G. , Gottfried, M. , Gruber, A. , and Pauli, H. , 1995: Patterns and current changes in alpine plant diversity. In Chapin, F. S. , and Körner, C. (eds.), Arctic and Alpine Biodiversity. Patterns, Causes and Ecosystem Consequences. Berlin: Springer, 167–181.
- Graham, R. W. , and Grimm, E. C. , 1990: Effects of global climate change on the patterns of terrestrial biological communities. Trends in Ecology and Evolution, 5: 289–311.
- Henderson-Sellers, A. , 1990: Modeling and monitoring ‘greenhouse' warming. Trends in Ecology and Evolution, 5: 270–275.
- IPCC [Intergovernmental Panel on Climate Change], 2007: Summary for policymakers. In Solomon, S. , Qin, D. , Manning, M. , Chen, Z. , Marquis, M. , Averyt, K. B. , Tignor, M. , and Miller, H. L. (eds.), Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the IPCC. Cambridge, Cambridge University Press, 1–18.
- Jackson, S. T. , and Overpeck, J. T. , 2000: Responses of plant populations and communities to environmental changes of the late Quaternary. Paleobiology, 26: 194–220.
- Klanderud, K. , 2008: Species-specific responses of an alpine plant community under simulated environmental change. Journal of Vegetation Science, 19: 363–372.
- Klanderud, K. , and Birks, H. J. B. , 2003: Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. The Holocene, 13: 1–6.
- Klanderud, K. , and Totland, Ø. , 2005: Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology, 86: 2047–2054.
- Klein, J. A. , Harte, J. , and Zhao, X. , 2007: Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecological Applications, 17: 541–557.
- Körner, C. , 1999: Alpine Plant Life. Functional Ecology of High Mountain Ecosystems. Berlin: Springer, 344 pp.
- Lesica, P. , 2002: Flora of Glacier National Park. Corvallis, Oregon State University Press, 512 pp.
- Lesica, P. , 2012: Manual of Montana Vascular Plants. Fort Worth: Botanical Research Institute of Texas, 771 pp.
- Lesica, P. , and McCune, B. , 2004: Decline of arctic-alpine plants at the southern margin of their range following a decade of climatic warming. Journal of Vegetation Science, 15: 679–690.
- Lesica, P. , and Steele, B. M. , 1996: A method for monitoring long-term population trends: an example using rare arctic-alpine plants. Ecological Applications, 6: 879–887.
- Malanson, G. P. , Bengtson, L. E. , and Fagre, D. B. , 2012: Geomorphic determinants of species composition of alpine tundra, Glacier National Park, U.S.A. Arctic, Antarctic, and Alpine Research, 44:197–209.
- McNulty, A. K. , Cummins, W. R. , and Pellizzar, A. , 1988: A field survey of respiration rates in leaves of Arctic plants. Arctic, 41: 1–5.
- Nagy, L. , and Grabherr, G. , 2009: The Biology of Alpine Habitats. Oxford: Oxford University Press, 376 pp.
- Parolo, G. , and Rossi, G. , 2008: Upward migration of vascular plants following a climate warming trend in the Alps. Basic and Applied Ecology, 9: 100–107.
- Pauli, H. , Gottfried, M. , Reiter, K. , Klettner, C. , and Grabherr, G. , 2007: Signals of range expansions and contractions of vascular plants in the high Alps: observations (1994–2004) at the GLORIA master site Schrankogel, Tyrol, Austria. Global Change Biology, 13: 147–156.
- Pederson, G. T. , Graumlich, L. J. , Fagre, D. B. , Kipfer, T. , and Muhfeld, C. C. , 2011: A century of climate and ecosystem change in western Montana: what do temperature trends portend? Climatic Change, 98: 133–154.
- Root, T. L. , Price, J. T. , Hall, K. R. , Schneider, S. H. , Rosenzweigk, C. , and Pounds, J. A. , 2003: Fingerprints of global warming on wild animals and plants. Nature, 421: 57–60.
- Scherrer, D. , and Körner, C. , 2010: Infra-red thermometry of alpine landscapes challenges climatic warming projections. Global Change Biology, 16: 2602–2613.
- Spomer, G. G. , and Salisbury, F. B. , 1968: Ecophysiology of Geum turbinatum and implications concerning alpine environments. Botanical. Gazette, 129: 33–49.
- Walker, D. A. , Halfpenny, J. C. , Walker, M. D. , and Wessman, C. A. , 1993: Long-term studies of snow-vegetation interactions. BioScience, 43: 287–301.
- Walker, M. D. , Webber, P. J. , Arnold, E. H. , and Ebert-May, D. , 1994: Effects of interannual variation on the aboveground phytomass in alpine vegetation. Ecology, 75: 393–408.
- Walker, M. D. , Wahren, C. H. , Hollister, R. D. , Henry, G. H. R. , Ahlquist, L. E. , Alatalo, J. M. , Bret-Harte, M. S. , Calef, M. P. , Callaghan, T. V. , Carroll, A. B. , Epstein, H. E. , Jónsdóttir, I. S. , Klein, J. A. , Magnusson, B. , Molau, U. , Oberbauer, S. F. , Rewa, S. P. , Robinson, C. H. , Shaver, G. R. , Suding, K. N. , Thompson, C. C. , Tolvanen, A. , Totland, Ø. , Turner, P. L. , Tweedie, C. E. , Webber, P. J. , and Wookey, P. A. , 2006. Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences of the United States, 103: 1342–1346.
- Walther, G. , Sascha, B. , and Burga, C. A. , 2005: Trends in the upward shift of alpine plants. Journal of Vegetation Science, 16: 541–548.
- Weaver, J. E. , 1968: Prairie Plants and Their Environment, a Fifty-year Study in the Midwest. Lincoln: University of Nebraska Press, 276 pp.
- Webb, T. , 1992: Past changes in vegetation and climate: lessons for the future. In Peters, R. L. , and Lovejoy, T. E. (eds.), Global Warming and Biological Diversity. New Haven: Yale University Press, 59–75.
- Wipf, S. , Stoeckli, V. , and Bebi, P. , 2005: Winter climate change in alpine tundra: plant responses to changes in snow depth and snowmelt timing. Climatic Change, 94: 105–121.
- Yang, Y. , Wang, G. , Klanderud, K. , and Yang, L. , 2011: Responses in leaf functional traits and resource allocation of a dominant alpine sedge (Kobresia pygmaea) to climate warming in the Qinghai-Tibetan Plateau permafrost region. Plant and Soil, 349: 377–387.