
Abstract
Several factors may have interactive effects on natural 15N abundance of plant species. Some of these effects could be associated with different plant functional types, including mycorrhizal association type. Due to its high taxonomic and functional diversity, the alpine heath community in the Caucasus is a suitable object for studying 15N natural abundance of plants in relation to different functional/mycorrhizal groups, contrasting with the limited numbers of plant groups or species considered in previous studies of individual communities. The N concentration and δ15N were determined in leaves of 25 plant species from 8 functional/mycorrhizal groups from an alpine lichen heath in the Teberda Reserve, Northern Caucasus, Russia. Functional groups were represented by ericoid mycorrhizal species (ERI), ectomycorrhizal species (ECT), arbuscular mycorrhizal forbs (AM—FORB), arbuscular mycorrhizal grasses (AM—GRA), arbuscular mycorrhizal nodulated legumes (FAB—N), non-mycorrhizal graminoids (sedges and rushes) (NOM—GRA), non-mycorrhizal hemiparasites (NOM—SP), and orchids (ORC). We can summarize our results in two rankings for leaf N concentration (FAB—N > ORC > AM—FORB, ECT > NOM—SP, ERI ≥⃒ NOM—GRA, AM—GRA) and leaf δ15N signature (ORC > NOM—GRA, FAB—N > ECT ≥⃒ ERI ≥⃒ AM—FORB, NOM—SP, AM—GRA) of alpine heath species. We conclude that, within the alpine lichen heath in the Northern Caucasus, the δ15N signature of plant foliage is a relevant indicator of plant functional groups with relatively high 15N content (ORC, FAB-N, NOM-GRA), while the absence of a significant difference between relatively 15N-depleted groups (AM, ERI, and ECT species) isn't clear and may result from both processes, as the increased N isotope fractionation by arbuscular mycorrhizal fungi as the decreased role of ecto- and ericoid mycorrhizal fungi in the flux of N.
Introduction
Plant growth in alpine communities is often nitrogen (N) limited, and the various adaptations of plant species to cope with such limitation may be reflected in interspecific variation in leaf N concentrations and leaf δ15N signature, which give some information about N uptake and conservation strategies in infertile environments (CitationMichelsen et al., 1998; CitationCornelissen et al., 2001; CitationQuested et al., 2003; CitationHobbie and Hobbie, 2008). However, several underlying factors may have interactive effects on natural 15N abundance of plant species. Such factors include (1) the difference in δ15N of nitrogen sources (NH4 +, NO3 -, N2, organic N), (2) the difference in δ15N of the same sources in different soil horizons (as related to plant rooting depth), (3) isotope discrimination during N uptake and assimilation, and (4) the influence of mycorrhiza (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996; CitationHögberg, 1997; CitationEvans, 2001; CitationRobinson, 2001; CitationMiller and CitationBowman, 2002 ; CitationCraine et al., 2009). Some of these effects could be associated with different plant functional types, including mycorrhizal association type, and have been convincingly confirmed for co-occurring species in field studies.
The most evident differentiation in leaf δ15N has been shown for plants with different types of mycorrhiza. 15N depletion of ectomycorrhizal and ericoid mycorrhizal species was found for tundra, alpine, and forest plants from different northern regions and was initially attributed to the influence of soil nitrogen sources, that is, fungal uptake of 15N-depleted organic N compounds (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996; CitationMichelsen et al., 1996, Citation1998). However, subsequent studies indicated that ectomycorrhizal and ericoid mycorrhizal species are 15N-depleted by preferred transfer of 14N from mycorrhizal fungi to their host plants (CitationHögberg et al., 1999; CitationHobbie et al., 2000; CitationEmmerton et al., 2001; CitationHobbie and Colpaert, 2003).
All other effects (different nitrogen sources, source localization in the soil, and isotope fractionation during nitrogen uptake) have been accompanied with no or very special empirical field observations. For instance, the influence of N sources on 15N concentration in plant leaves has been indicated for spatially separated plants in agricultural studies where δ15N of plants directly reflected utilization of isotopically different N fertilizers (CitationChoi et al., 2003; CitationYun et al., 2006). Another example is the differences in δ15N for plants of different communities along a geochemical gradient for which a clear effect of 15N abundances of N sources was observed (CitationGarten, 1993). However, for co-occurring species within a community, the effect of different nitrogen sources was observed in special cases only, that is, legumes that obtain atmospheric N2 through fixation by symbiotic bacteria (CitationBowman et al., 1996; CitationMichelsen et al., 1996; CitationNadelhoffer et al., 1996; CitationKörner, 2003; CitationHobbie et al., 2005) or orchids that obtain N from their mycorrhizal fungi by transfer from hyphae (CitationGebauer and Meyer, 2003; CitationZimmer et al., 2007).
The influence of rooting depth has been confirmed experimentally even less. Earlier assumptions that higher δ15N values of arbuscular mycorrhizal Calamagrostis canadensis (CitationSchulze et al., 1994) and non-mycorrhizal Eriophorum vaginatum (CitationNadelhoffer et al., 1996) in central and northern Alaska ecosystems was a result of deeper roots seem incorrect because these species were compared with ericoid mycorrhizal and ectomycorrhizal species in which 15N depletion is a function of metabolic N fractionation by mycorrhizal fungi (CitationHögberg et al., 1999; CitationHobbie et al., 2000; CitationEmmerton et al., 2001). This difference could be also connected with different preferential uptake of N compounds. Furthermore, our recent study indicated that plants with deeper root systems can probably consume lighter rather than heavier NH4 +-N (CitationMakarov et al., 2008).
Due to its high taxonomic and functional diversity, the alpine heath community in the Caucasus is a suitable object for studying 15N natural abundance of plants in relation to different functional/mycorrhizal groups, contrasting with the limited numbers of plant groups or species considered in previous studies of individual communities. We hypothesized that δ15N of plant leaves within the alpine heath community is the result of several interacting drivers associated with their functional/mycorrhizal status. The objectives of this study were (1) to compare N concentration and δ15N in leaves of co-occurring species within an alpine heath community, and (2) to investigate if variation in N concentration and 15N natural abundance is characteristic for different plant functional/mycorrhizal groups.
Material and Methods
SITE DESCRIPTION
The research was conducted at the Teberda Biosphere Reserve (Northwestern Caucasus, Russia). The study site was located at Mount Malaya Khatipara (43°27′N, 41°42′E) at 2750 m a.s.l. in an alpine lichen heath, which has been the subject of diverse ecological studies during more than 20 years (CitationOnipchenko, 2004). The climate of the area is characterized by low air temperatures (mean annual temperature is -1.2 °C, mean July temperature is 7.9 °C) and high annual precipitation (1400 mm). The lichen heath covers snow-free wind-exposed ridges and upper slopes in the alpine landscape. The plant community is dominated by fruticose lichens (mainly Cetraria islandica), which cover about 30%-60% of the area. Vascular plants are represented by more than 40 species. The most common are Anemone speciosa, Antennaria dioica, Trifolium polyphyllum, Festuca ovina, Carex sempervirens, Carex umbrosa, Campanula tridentata, and Vaccinium vitisidaea. Detailed description of the site plant community, including species' nomenclature, is published elsewhere (CitationOnipchenko, 2002).
STUDIED SPECIES
Twenty-five species from eight functional/mycorrhizal groups (three species per group, except legumes which were represented by three nodulated and one non-nodulated [Trifolium polyphyllum] species) were studied. The different functional groups with respect to nutrient uptake strategy were represented by ericoid mycorrhizal species (ERI), ectomycorrhizal species (ECT), arbuscular mycorrhizal forbs (AM—FORB), arbuscular mycorrhizal grasses (AM—GRA), arbuscular mycorrhizal nodulated legumes (FAB—N), non-mycorrhizal graminoids (sedges and rushes) (NOM—GRA), non-mycorrhizal hemiparasites (NOM—SP), and orchids (ORC) (). Type of mycorrhiza and mycorrhizal infection rate for arbuscular mycorrhizal species (proportion of thin root mesoderm cells occupied by fungi) were studied earlier (CitationOnipchenko and Zobel, 2000; CitationOnipchenko, 2004).
Foliage from these plant species was sampled during one week at the beginning of August in five replications from a sampling site of 30 × 30 m in size. In general, leaves from five separate plants per species were collected, although in some cases several individuals of a species were pooled to make one replicate sample.
CHEMICAL ANALYSES
Total N was determined by dry combustion on an Elementar Vario EL elemental analyzer. Natural 15N abundance of plant samples was determined by dry combustion on a Carlo Erba NC 2500 elemental analyzer coupled with a Deltaplus continuous-flow isotope ratio mass spectrometer (Thermo Finnigan, Bremen, Germany) and expressed as δvalues, which were defined as:
STATISTICS
Nested design ANOVAs in general linear models were applied to estimate differences in nitrogen content and δ15N for plant species and functional types, with three species nested in each functional type (and excluding the non-nodulated legume). In case of significant effects, post hoc Tukey HSD test was used for comparison of means between individual species or functional groups. The calculations were made in Statistica 6.0 for Windows.
To compare nitrogen content and δ15N between three nodulated legumes and the non-nodulated legume Trifolium polyphyllum, one-way ANOVA was used. Correlation coefficients were calculated to test for a relationship between mycorrhizal infection rate and nitrogen content or δ15N for AM forbs and grasses.
Results
NITROGEN CONCENTRATION
The N concentration varied significantly and widely among alpine plant species and functional groups of lichen heath ( and , ). Festuca ovina (AM-GRA) had the lowest N concentration (0.97%), whereas Astragalus levieri (FAB-N) and Traunsteinera globosa (ORC) were the most N-rich (3.30% and 3.34%, respectively). The groups of N2-fixing legumes, orchids, arbuscular mycorrhizal forbs, and ectomycorrhizal species generally had higher N concentrations than other groups. Non-mycorrhizal hemiparasites and ericoid mycorrhizal species tended to have a somewhat higher N concentration among the four groups with relatively low N concentrations. Graminoids, irrespective of their mycorrhizal status, had the lowest N leaf concentration. So, we may represent the following ranking of the leaf N concentration among alpine heath species:
TABLE 1 Mycorrhizal and functional types of the studied plant species.
TABLE 2 Results of nested ANOVA (species are nested in functional type) for nitrogen concentration and δ15N.
TABLE 3 Leaf N concentration and δ15N for studied alpine species from different functional types (see for abbreviations).
FIGURE 1. Leaf N concentration for different functional types of alpine plants (mean and standard error) (see for abbreviations). Significant (p < 0.05) differences are shown by different letters.
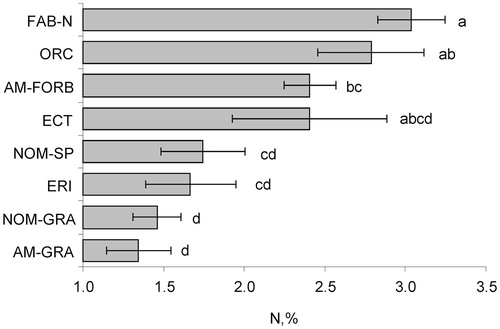
FIGURE 2. Leaf N concentration and δ15N for 3 nitrogen fixing and one non-fixing (Trifolium polyphyllum) legumes in the alpine lichen heath. Significant (p < 0.05) differences are shown by different letters.
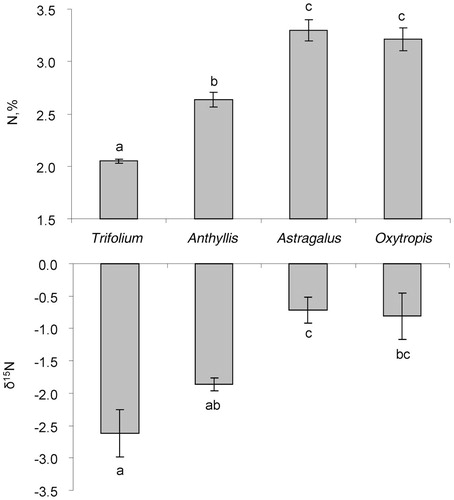
However, within separate functional groups the variation was also high, and in each group there were species differing significantly in N concentration from each other. For example, ectomycorrhizal Polygonum viviparum had more than twice the N concentration (3.12%) of another ectomycorrhizal species-Juniperus communis (1.48%). Within other groups, pronounced differences were also observed (). A separate one-way ANOVA for legumes indicated that leaf N concentration in non-fixing Trifolium polyphyllum was significantly lower than the N concentrations for other legumes ().
FIGURE 3. Relationship between N leaf concentration and rate of mycorrhizal infection (%) for 6 arbuscular mycorrhizal (AM) plant species.
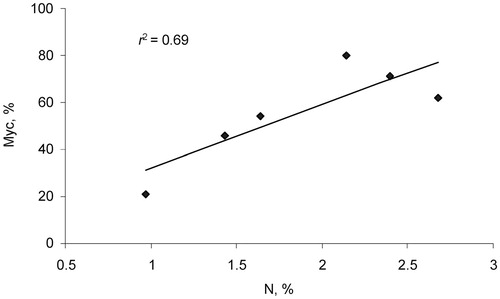
There was a highly significant positive correlation (r 2 = 0.70) for six AM species (forbs and grasses) between N leaf concentration and rate (degree) of AM infection (). The same tendency is apparent within three species of grasses, but not within forbs.
15N ABUNDANCE
Natural abundance of 15N in the plant leaves also varied widely among species within the lichen heath community, ranging from -5.5‰ in Bromus variegatus (AM-GRA) to +2.3‰ Traunsteinera globosa (ORC). All orchid species had about zero or positive δ15N; their mean of +0.8‰ was significantly higher than that in any other group, excluding the marginally 15N-depleted non-mycorrhizal graminoids (sedges and rushes) and nodulated legumes (, ). In contrast, there was no single most 15N-depleted group. Pronounced 15N depletion was typical for arbuscular mycorrhizal forbs and grasses, non-mycorrhizal hemiparasites, and ericoid mycorrhizal species, with δ15N values ranging from -3.8‰ to -2.9‰ and not significantly differing from each other. Ectomycorrhizal species were intermediate. We can summarize these results in the following ranking of leaf 15N enrichment among alpine heath species:
Similar to N concentration, δ15N varied widely within separate functional groups, often by more than 2‰. For example, Helictotrichon versicolor and Bromus variegatus (AM-GRA species) has δ15N values of -2.2‰ and -5.5‰, respectively (). Within all legumes, non-nodulated Trifolium polyphyllum had the lowest δ15N (-2.6‰), which was close to the values for the ERI and ECT mycorrhizal groups (). This value was significantly lower than δ15N of two other legume species-Oxytropis kubanensis and Astragalus levieri-but there was no significant difference between δ15N for Trifolium polyphyllum and Anthyllis vulneraria ().
In contrast to N concentration, δ15N did not show any relationships with mycorrhizal infection rate for six AM species.
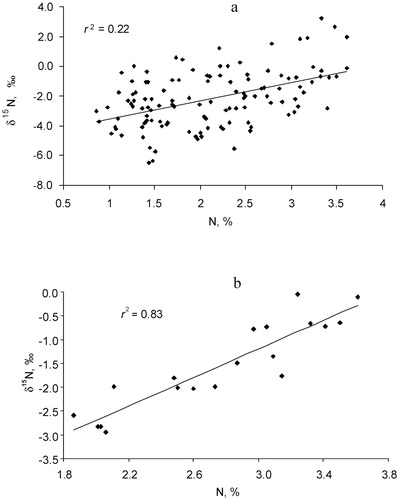
Across all samples, foliar δ15N increased with increasing N concentrations (r 2 = 0.22, P < 0.001) (, part a). The group of non-mycorrhizal sedges, being strongly N-depleted and relatively 15N-enriched, was the main functional group that decreased the strength of this correlation. A highly significant correlation (r 2 = 0.83, P < 0.001) between N concentration and δ15N was observed within the group of legume species (, part b).
FIGURE 4. δ15N for different functional types of alpine plants (mean and standard error) (see for abbreviations). Significant (p < 0.05) differences are shown by different letters.
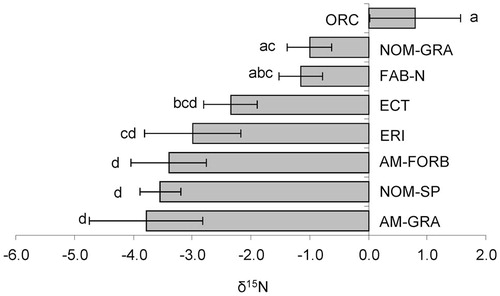
FIGURE 5. Relationships between N concentration and δ15N in alpine plants: (a) all species; (b) legume species.
Discussion
NITROGEN CONCENTRATION IN ALPINE PLANTS
Leaf N concentrations in our study for all but four of the studied species were lower than the mean values for alpine plants in the Alps (2.87%) and the northern Scandes (3.18%) (CitationKörner, 1989). The mean value for the Caucasian species was 2.11%, which is lower than the N concentration in leaves of plant species even from low altitude in the Alps (2.40%) (CitationKörner, 1989). Monson et al. (Citation2001) reported mean foliar N concentration in alpine forbs in Niwot Ridge (Colorado) of about 3.1%, which is higher than our data for AM forbs (2.4%). Relatively low N concentrations in the species of the lichen heath may be explained by a general tendency of decreasing N concentration in alpine plants with decrease of latitude (CitationKörner, 2003). Also, nitrogen availability in lichen heath is the lowest among the plant communities in the same region (CitationMakarov et al., 2003), as evidenced by a doubling of productivity after N fertilization (CitationSoudzilovskaia et al., 2005).
Hemiparasitic plants usually have higher N and other nutrient concentrations than their hosts (CitationPress et al., 1999). Our results for three Scrophulariaceae species (annual Euphrasia ossica and perennial Pedicularis comosa and P. caucasica) showed that N leaf concentration was lower than those values for AM-FORB but higher than for NOM-GRA (both groups can serve as hosts for studied hemiparasites [CitationPopova, 1966]). We can suppose that the results indicate relatively low or facultative root parasitic activity of the studied species.
Nitrogen concentration in leaves of Trifolium polyphyllum was significantly lower (2.05%) than in other studied legumes (2.64%-3.30%). This result confirms our visual observations of the absence of nodules in the root system of Trifolium polyphyllum. The non-nitrogen fixing status of Trifolium polyphyllum was also confirmed by its positive responses to N fertilization (CitationSoudzilovskaia et al., 2005; CitationSoudzilovskaia and Onipchenko, 2005; CitationSoudzilovskaia et al., 2012) and by the acetylene reduction method (CitationMakarov et al., 2011). Trifolium polyphyllum is therefore the first known temperate non-fixing legume besides previously reported tropical examples (CitationSprent, 2005). The N concentrations of the three other legume species that do have root nodules correspond to those reported from high mountain ecosystems in Colorado and the Alps (CitationBowman et al., 1996; CitationJacot et al., 2000), confirming the adaptiveness of this strategy at high altitude.
Arbuscular mycorrhizal fungi are known to provide plant access to organic soil phosphates and improve mainly phosphate acquisition of plants (CitationCavagnaro et al., 2003; CitationSmith and Read, 2008). Nitrogen capture from organic material and transferring N to plants via AM is a recently discovered phenomenon (CitationNakano et al., 2001; CitationJin et al., 2005; CitationLeigh et al., 2009). However, to our knowledge there is no example in the literature of positive relationships between rate of AM infection and leaf N concentration. For example, the alpine species Ranunculus adoneus had no relationship between AM root colonization and leaf N concentration (CitationMonson et al., 2001). In spite of the limited number of AM species in the present research, the strong positive correlation between these parameters () is consistent with AM fungi influencing plant nitrogen uptake as well.
15N NATURAL ABUNDANCE IN ALPINE PLANTS
The δ15N variation among plant species could be due to several factors. Many surveys highlighted the influence of the mycorrhizal status of species (CitationSchulze et al., 1994; CitationMichelsen et al., 1996, Citation1998; CitationNadelhoffer et al., 1996; CitationHögberg et al., 1999; CitationHobbie et al., 2000; CitationEmmerton et al., 2001; CitationHobbie and Colpaert, 2003), while other factors (species-specific differences in isotopic fractionation, differences in plant preference for N forms and root placement at different soil depths) could also be important (CitationSchulze et al., 1994; CitationEmmerton et al., 2001; CitationMiller and Bowman, 2002).
Relatively high 15N abundances of plant leaves of ORC, FAB-N, and NOM-GRA species from an alpine lichen heath correspond to previously published results. For instance, the highest δ15N values of orchids were also observed in forest and some grassland sites in Bavaria and southern France and were explained by utilization of fungi-derived 15N-enriched nitrogen (CitationGebauer and Meyer, 2003).
N2-fixing legumes that partly acquire atmospheric N2 have higher δ15N (close to naught) relative to non-fixing species growing in N-limited conditions of tundra and alpine ecosystems (CitationBowman et al., 1996; CitationMichelsen et al., 1996; CitationNadelhoffer et al., 1996; CitationKörner, 2003; CitationHobbie et al., 2005). This difference in 15N signature between N2-fixing and non-fixing species is often used to assess the degree of N2 fixation by symbiotic bacteria using the dual-sources model (CitationShearer and Kohl, 1986). Although this method requires care in selecting non-fixing reference plants, which ideally should be characterized by the same preference of soil N and the same temporal and spatial N uptake pattern as N-fixing species, it was successfully applied for alpine ecosystems (CitationBowman et al., 1996). The studied alpine lichen heath provides a unique test case for revealing N2-fixation with the δ15N method. The non-nodulated legume Trifolium polyphyllum provides a taxonomically close reference species for N2-fixing legumes. The lowest N concentration and δ15N in Trifolium polyphyllum leaves () and highly significant correlation between N concentration and δ15N for all legume species (, part b) allow us to conclude that the 15N natural abundance method can provide an acceptable estimation of N2-fixation. Applying the dual-source model and the result of Bowman et al. (Citation1996) that 15N enrichment of the legume grown solely with atmospheric N2 is close to 0‰, we calculated that the percentage of plant N that is fixed from the atmosphere varied from 27% in Anthyllis vulneraria to 69% and 73% for Oxytropis kubanensis and Astragalus levieri, respectively (CitationMakarov et al., 2011). The latter results are in close correspondence with the large proportions of atmospherically fixed N2 (from 70% up to 100%) to satisfy N requirements of legumes in other mountains (CitationBowman et al., 1996; CitationJacot et al., 2000).
The 15N enrichment of non-mycorrhizal graminoids (sedges and rushes) of alpine lichen heath corresponds to the high 15N content in plant species from this functional group within two plant communities in northern Swedish Lapland where Carex spp. and Luzula arcuata had δ15N values between -0.5‰ and +2.5‰ (CitationMichelsen et al., 1996). Non-mycorrhizal graminoids in Alaskan tundra (Eriophorum vaginatum and Carex bigelowii) and in Niwot Ridge alpine tundra (Kobresia myosuroides, Carex rupestris, Luzula spicata) were also among the species with the highest δ15N (up to +2.3‰) (CitationNadelhoffer et al., 1996; CitationMiller and Bowman, 2002; CitationKörner, 2003). According to Körner (Citation2003), a probable reason for high 15N enrichment of Cyperaceae species (up to +3.5‰) could be their access to 15N-enriched stable forms of organic N. However, the more recent finding of considerable 15N enrichment of DON (CitationPörtl et al., 2007) indicated that the reason could also be access to labile forms of organic N. In particular, Raab et al. (Citation1996, Citation1999) demonstrated that non-mycorrhizal alpine Cyperaceae species had uptake rates for glycine similar to or substantially greater than those for inorganic N.
Isotopic nitrogen fractionation during ammonium and nitrate plant assimilation should essentially complicate the measure of the δ15N of available N in the soil (CitationEmmerton et al., 2001). However, 15N discrimination during nitrogen uptake will only be observed when plant nitrogen demand is relatively low, compared with the nitrogen available in the solution, and will probably decrease to negligible under conditions of low N concentration and highly efficient N uptake typical for most natural ecosystems. Hence, in natural N-limited ecosystems the δ15N of non-mycorrhizal plants is a good approximation of δ15N of the available N (CitationHögberg et al., 1999; CitationHobbie et al., 2005). This assumption was confirmed by very similar δ15N values of non-mycorrhizal plants (between -2‰ and +2‰) (CitationMichelsen et al., 1996) and of NH4 +-N (between -1.5‰ and +1.4‰) (CitationMakarov et al., 2008) from the surface soil horizon in tundra ecosystems in northern Sweden. Similar δ15N were also found for non-mycorrhizal plants, total soil N, ammonium, and nitrate N in the arctic tundra ecosystem in Alaska (CitationHobbie and Hobbie, 2006).
Conditions of N-limitation are quite applicable to the alpine lichen heath where inorganic N concentrations in the soil solution are typically low at about 0.05 mg L-1 both N-NH4 + and N-NO3 - (CitationMakarov et al., 2003). However, in our case, alpine non-mycorrhizal sedges were much more 15N-enriched (between -0.2‰ and -1.5‰) than the dominant form of inorganic soil N-exchangeable NH4 +-N (between -2.6‰ and -5.1‰) (CitationMakarov et al., 2008). It seems likely that the 15N abundance of alpine non-mycorrhizal plants is the result of uptake of a mixture of N sources.
15N AND TYPE OF MYCORRHIZA
In spite of the prevalent opinion that there is the great difference between ericoid mycorrhizal and ectomycorrhizal species on the one hand (more negative δ15N values), and non-mycorrhizal or arbuscular mycorrhizal species on the other hand (less negative δ15N values) (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996; CitationMichelsen et al., 1996, Citation1998; CitationKörner, 2003; CitationCraine et al., 2009), it was not confirmed in our study in an alpine heath community, where 15N abundance of ericoid mycorrhizal and especially ectomycorrhizal species tended to be rather higher than that of arbuscular mycorrhizal forbs and grasses (). Some other previous results also did not correspond to this prevalent pattern. For example, the isotopic similarity within ecto- and arbuscular mycorrhizal species in forest sites in Bavaria was explained by utilization of isotopically similar N compounds (CitationGebauer and Meyer, 2003). Also, Hobbie et al. (Citation2005) demonstrated that some arbuscular mycorrhizal species (Luetkea pectinata) had similar δ15N values with ecto- and ericoid mycorrhizal plants in the Cascade Mountains of Washington, U.S.A. Last, our study of AM species in subarctic meadow communities in the Khibiny Mountains (NW Russia) and Abisko region (N Sweden) indicated that these plants were characterized by low δ15N values close to ERI and ECT species (not published).
These findings could be connected with increased N isotope fractionation by arbuscular mycorrhizal fungi and preferential transfer of 15N-depleted compounds from fungi to host plants in a similar fashion to the well-established transfer in ectomycorrhizal symbioses (CitationHobbie and Hobbie, 2008; CitationCraine et al., 2009). However, it remains unclear why it occurs in meadow communities where nitrogen is probably more available to plants in comparison with heaths for which much higher δ15N values of AM species are typical (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996; CitationMichelsen et al., 1996, Citation1998). In addition, culture studies investigating the effects of arbuscular mycorrhizal colonization on plant δ15N indicated only modest changes and sometimes resulted in 15N enrichment rather than depletion (CitationHandley et al., 1993, Citation1999; CitationAzcon-Aguilar et al., 1998).
On the other hand, the rate of 15N fractionation by ecto- and ericoid mycorrhizal fungi probably depends on N availability. In conditions of strongly pronounced N limitation, plants take up the most part of N through mycorrhiza and thus become 15N-depleted, while for ecosystems with higher N availability and lower importance of mycorrhiza in plant N nutrition, similarity between ecto-, ericoid, and arbuscular mycorrhizal species is more typical (CitationHobbie et al., 2000, Citation2005). The alpine lichen heath, while being the most N limited among alpine ecosystems in the Teberda Reserve, is probably not as limited as arctic and subarctic tundra ecosystems, because annual net N mineralization rate in the heath soil (0.6 g m-2) (CitationMakarov et al., 2003) is higher than in soils of Arctic ecosystems (0.05–0.5 g m-2) (CitationGiblin et al., 1991; CitationSchmidt et al., 2002). Probably this is the reason of relatively high δ15N values of ERI and ECT plant species, which isn't comparable with values close to -8‰ or -9‰, as it was observed when studying subarctic communities (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996; CitationMichelsen et al., 1996, Citation1998).
Therefore we assume that the reason of absence of a significant difference between AM, ERI, and ECT species in the Caucasian alpine heath community isn't clear and may result from both processes: the increased N isotope fractionation by arbuscular mycorrhizal fungi and the decreased role of ecto- and ericoid mycorrhizal fungi in the flux of N.
VARIATION IN δ15N VALUES WITHIN FUNCTIONAL GROUPS
The variability of δ15N within separate plant functional groups, as discussed above for legumes, can be due to species-specificity in isotopic fractionation (CitationEmmerton et al., 2001), differences in plant preference for N forms (CitationSchulze et al., 1994; CitationMiller and Bowman, 2002), and their root placement at different soil depths (CitationSchulze et al., 1994; CitationNadelhoffer et al., 1996).
We have no experimental data to evaluate the first two determinants, while the effect of rooting depth was not confirmed in our study, because δ15N varied widely among species of the same functional group with similar rooting depths. For example, arbuscular mycorrhizal grasses Helictotrichon versicolor and Bromus variegatus, both have rooting depths mainly within the upper 5–10 cm of soil but very different δ15N values of -2.2‰ and -5.5‰, respectively. Similarly, Anemone speciosa and Campanula tridentata (AM-FORB), both with deeper (15–20 cm) root systems (CitationOnipchenko, 1987), have δ15N values of -2.2‰ and -4.4‰, respectively.
The dependence of δ15N of plant leaves on rooting depth is one of the least confirmed in field observations. We do not know of any studies that experimentally demonstrate any relationships between δ15N of plant leaves and rooting depth, although Schulze et al. (Citation1994) and Nadelhoffer et al. (Citation1996) hypothesized that deep rooting system can be responsible for the δ15N increase of forest and Arctic tundra plant species (see Introduction). Similar to our study, considerable foliar δ15N differences within functional groups were found for forbs, grasses, and sedges (rushes) in two dry meadow alpine tundra sites in the Rocky Mountains, Colorado (CitationMiller and Bowman, 2002), while there were no differences in the δ15N of bulk soils within the upper 15 cm where roots of dry meadow species are concentrated. Therefore, the authors concluded that the observed variation in the foliar δ15N was due to factors other than rooting depth and was attributed to the acquisition of different N forms. At the same time, δ15N of non-mycorrhizal and ectomycorrhizal fine roots in European forest soils usually increased with soil depth (CitationHögberg et al., 1996), and the idea of Taylor et al. (Citation1997) that δ15N values of ectomycorrhizal fungi may be a function of the soil depth at which they utilize the N compounds was subsequently confirmed by Wallander et al. (Citation2004) for the ectomycorrhizal mycelia, the δ15N of which increased with soil depth in two forest types.
Though we assumed that low δ15N values of the studied AM species may result from the increased N isotope fractionation by arbuscular mycorrhizal fungi, it also remains unclear whether arbuscular mycorrhizal fungi contribute to variation in foliar δ15N, because we did not find any relationships with mycorrhizal colonization rate for six AM species.
RELATION BETWEEN FOLIAR N AND δ15N
Positive relationships between foliar δ15N and N concentration were reported at the global (CitationCraine et al., 2009) and landscape (regional) scale (CitationHobbie et al., 2000), indicating the effect of N availability. Hobbie et al. (Citation2000) determined δ15N in plant materials across six sites representing different postdeglaciation ages at Glacier Bay, Alaska. When compared across all sites and species, the foliar δ15N values and N concentrations were generally lower at sites of low N availability, suggesting either an increased fraction of N obtained from mycorrhizal uptake, or a reduced proportion of mycorrhizal N transferred to vegetation. However, within our alpine lichen heath, higher foliar N concentrations and δ15N values are mostly connected with atmospheric N2 fixation by legumes (see also CitationHobbie et al., 2000, for Glacier Bay) or utilization of fungi-derived 15N-enriched N by orchids and accompanying species.
Conclusion
We conclude that, within the alpine lichen heath in the Northern Caucasus, the δ15N signature of plant foliage is a relevant indicator of plant functional groups with relatively high 15N content (ORC, FAB-N, NOM-GRA), while the lack of clear difference between relatively 15N-depleted groups (AM, ERI and ECT species) isn't clear and may result from both processes: the increased N isotope fractionation by arbuscular mycorrhizal fungi and the decreased role of ecto- and ericoid mycorrhizal fungi in the flux of N. These findings are important pieces of the large puzzle of understanding belowground competition and complementarity in N-limited ecosystems.
Acknowledgments
This work was sponsored by the Netherlands Organization for Scientific Research (NWO grants 047.011.2004.005, 047.017.010, and 047.018.003) and the Russian Foundation for Basic Research (10-04-00780 and 11-04-01215).
Related Research Data
References Cited
- Azcon-Aguilar, G. R. , Handley, L. L. , and Scrimgeour, C. M. , 1998: The δ15N of lettuce and barley are affected by AM status and external concentration of N. New Phytologist , 138: 19–26.
- Bowman, W. D. , Schardt, J. C. , and Schmidt, S. K. , 1996: Symbiotic N2-fixation in alpine tundra: ecosystem input and variation in fixation rates among communities. Oecologia , 108: 345–350.
- Cavagnaro, T. R. , Smith, F. A. , Ayling, S. M. , and Smith, S. E. , 2003: Growth and phosphorus nutrition of a Paris-type arbuscular mycorrhizal symbiosis. New Phytologist , 157: 127–134.
- Choi, W. J. , Ro, H. M. , and Hobbie, E. A. , 2003: Patterns of natural 15N in soils and plants from chemically and organically fertilized uplands. Soil Biology and Biochemistry , 35: 1493–1500.
- Cornelissen, J. H. C. , Aerts, R. , Cerabolini, B. , Werger, M. J. A. , and van der Heijden, M. G. A. , 2001: Carbon cycling traits of plant species are linked with mycorrhizal strategy. Oecologia , 129: 611–619.
- Craine, J. M. , Elmore, A. J. , Aidar, M. P. M. , Bustamante, M. , Dawson, T. E. , Hobbie, E. A. , Kahmen, A. , Mack, M. C. , McLauchlan, K. K. , Michelsen, A. , Nardoto, G. B. , Pardo, L. H. , Peñuelas, J. , Reich, P. B. , Schuur, E. A. G. , Stock, W. D. , Templer, P. H. , Virginia, R. A. , Welker, J. M. , and Wright, I. J. , 2009: Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytologist , 183: 980–992.
- Emmerton, K. S. , Callaghan, T. V. , Jones, H. E. , Leake, J. R. , Michelsen, A. , and Read, D. J. , 2001: Assimilation and isotopic fractionation of nitrogen by mycorrhizal and nonmycorrhizal subarctic plants. New Phytologist , 151: 513–524.
- Evans, R. D. , 2001: Physiological mechanisms influencing plant nitrogen isotope composition. Trends in Plant Science , 6: 121–126.
- Garten, C. T., Jr. , 1993: Variation in foliar 15N abundance and the availability of soil nitrogen on Walker Branch Watershed. Ecology , 74: 2098–2113.
- Gebauer, G. , and Meyer, M. , 2003: 15N and 13C natural abundance of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal association. New Phytologist , 160: 209–223.
- Giblin, A. E. , Nadelhoffer, K. J. , Shaver, G. R. , Laundre, J. A. , and McKerrow, A. J. , 1991: Biogeochemical diversity along a riverside toposequence in arctic Alaska. Ecological Monographs , 61: 415–435.
- Handley, L. L. , Daft, M. J. , Wilson, J. , Scrimgeour, C. M. , Ingleby, K. , and Sattar, M. A. , 1993: Effects of the ecto- and VA-mycorrhizal fungi Hydnagium carneum and Glomus clarum on the δ15N and δ13C values of Eucalyptus globules and Ricinus communis. Plant, Cell and Environment , 16: 375–382.
- Handley, L. L. , Azcon, R. , Lozano, J. M. R. , and Scrimgeour, C. M. , 1999: Plant δ15N associated with arbuscular mycorrhization, drought and nitrogen deficiency. Rapid Communications in Mass Spectrometry , 13: 1320–1324.
- Hobbie, E. A. , and Colpaert, J. V. , 2003: Nitrogen availability and colonization by mycorrhizal fungi correlate with nitrogen isotope patterns in plants. New Phytologist , 157: 115–126.
- Hobbie, J. E. , and Hobbie, E. A. , 2006: 15N in symbiotic fungi and plants estimates nitrogen and carbon flux rates in Arctic tundra. Ecology , 87: 816–822.
- Hobbie, E. A. , and Hobbie, J. E. , 2008: Natural abundance of 15N in nitrogen-limited forests and tundra can estimate nitrogen cycling through mycorrhizal fungi: a review. Ecosystems , 11: 815–830.
- Hobbie, E. A. , Macko, S. A. , and Williams, M. , 2000: Correlations between foliar δ15N and nitrogen concentrations may indicate plant-mycorrhizal interactions. Oecologia , 122: 273–283.
- Hobbie, E. A. , Jumpponen, A. , and Trappe, J. , 2005: Foliar and fungal 15N:14N ratios reflect development of mycorrhizae and nitrogen supply during primary succession: testing analytical models. Oecologia , 146: 258–268.
- Högberg, P. , 1997: 15N natural abundance in soil-plant systems. New Phytologist , 137: 179–203.
- Högberg, P. , Högbom, L. , Schinkel, H. , Högberg, M. , Johannisson, C. , and Wallmark, H. , 1996: 15N abundance of surface soils, roots and mycorrhizas in profiles of European forest soils. Oecologia , 108: 207–215.
- Högberg, P. , Högberg, M. N. , Quist, M. E. , Ekblad, A. , and Näsholm, T. , 1999: Nitrogen isotope fractionation during nitrogen uptake by ectomycorrhizal and non-mycorrhizal Pinus sylvestris. New Phytologist , 142: 569–576.
- Jacot, K. A. , Lüscher, A. , Nösberger, J. , and Hartwig, U. A. , 2000: Symbiotic N2 fixation of various legume species along an altitudinal gradient in the Swiss Alps. Soil Biology and Biochemistry , 32: 1043–1052.
- Jin, H. , Pfeffer, P. E. , Douds, D. D. , Piotrowski, E. , Lammers, P. J. , and Shachar-Hill, Y. , 2005: The uptake, metabolism, transport and transfer of nitrogen in an arbuscular mycorrhizal symbiosis. New Phytologist , 168: 687–696.
- Körner, C. , 1989: The nutritional status of plants from high altitudes: a worldwide comparison. Oecologia , 81: 379–391.
- Körner, C. , 2003: Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin, Heidelberg: Springer-Verlag.
- Leigh, J. , Hodge, A. , and Fitter, A. H. , 2009: Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytologist , 181: 199–207.
- Makarov, M. I. , Glaser, B. , Zech, W. , Malysheva, T. I. , Bulatnikova, I. V. , and Volkov, A. V. , 2003: Nitrogen dynamics in alpine ecosystems of the Northern Caucasus. Plant and Soil , 256: 389–402.
- Makarov, M. I. , Malysheva, T. I. , Cornelissen, J. H. C. , van Logtestijn, R. S. P. , and Glaser, B. , 2008: Consistent patterns of 15N distribution through soil profiles in diverse alpine and tundra ecosystems. Soil Biology and Biochemistry , 40: 1082–1089.
- Makarov, M. I. , Malysheva, T. I. , Ermak, A. A. , Onipchenko, V. G. , Stepanov, A. L. , and Menyailo, O. V. , 2011: Symbiotic nitrogen fixation in the alpine community of a lichen heath of the northwestern Caucasus region (the Teberda Reserve). Eurasian Soil Science , 44: 1381–1388.
- Michelsen, A. , Schmidt, I. K. , Jonasson, S. , Quarmby, C. , and Sleep, D. , 1996: Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of nitrogen. Oecologia , 105: 53–63.
- Michelsen, A. , Quarmby, C. , Sleep, D. , and Jonasson, S. , 1998: Vascular plant 15N natural abundance in heath and forest tundra ecosystems is closely correlated with presence and type of mycorrhizal fungi in roots. Oecologia , 115: 406–418.
- Miller, A. E. , and Bowman, W. D. , 2002: Variation in nitrogen-15 natural abundance and nitrogen uptake traits among co-occurring alpine species: do species partition by nitrogen form? Oecologia , 130: 609–616.
- Monson, R. K. , Mullen, R. , and Bowman, W. D. , 2001: Plant nutrient relations. In : Bowman, W. D. , and Seastedt, T. R. (eds.), Structure and Function of an Alpine Ecosystem: Niwot Ridge, Colorado. Oxford: Oxford University Press, 198–221.
- Nadelhoffer, K. , Shaver, G. , Fry, B. , Giblin, A. , Johnson, L. , and McKane, R. , 1996: 15N natural abundances and N use by tundra plants. Oecologia , 107: 386–394.
- Nakano, A. , Takahashi, K. , Koide, R. T. , and Kimura, M. , 2001: Determination of nitrogen source for arbuscular mycorrhizal fungi by 15N application to soil and plants. Mycorrhiza , 10: 267–273.
- Onipchenko, V. G. , 1987: Mechanisms of ecological niches separation in terrestrial plants. Zhurnal obshchei biologii , 48: 687–695.
- Onipchenko, V. G. , 2002: Alpine Vegetation of the Teberda Reserve, the Northwest Caucasus. Zurich: Veroffentlichungen des Geobotanischen Institutes der ETH, Stiftung Rubel, H. 130.
- Onipchenko, V. G. (ed.), 2004: Alpine Ecosystems in the Northwest Caucasus. Dordrecht, Boston, London: Kluwer Academic Publishers.
- Onipchenko, V. G. , and Zobel, M. , 2000: Mycorrhiza, vegetative mobility and responses to disturbance of alpine plants in the Northwestern Caucasus. Folia Geobotanica , 35: 1–11.
- Popova, T. N. , 1966: On parasitic Caucasian Pedicularis. Biologicheskie nauki , 2: 113–118 (in Russian).
- Pörtl, K. , Zechmeister-Boltenstern, S. , Wanek, W. , Ambus, P. , and Berger, T. W. , 2007: Natural 15N abundance of soil N pools and N2O reflect the nitrogen dynamics of forest soils. Plant and Soil , 295: 79–94.
- Press, M. S. , Scholes, J. D. , and Watling, J. R. , 1999: Parasitic plants: physiological and ecological interactions with their hosts. In Press, M. C. , Scholes, J. D. , and Barker, M. G. (eds.), Physiological Plant Ecology. Oxford: Blackwell Science, 175–197.
- Quested, H. M. , Cornelissen, J. H. C. , Press, M. C. , Callaghan, T. V. , Aerts, R. , Trosien, F. , Riemann, P. , Gwynn-Jones, D. , Kondratchuk, A. , and Jonasson, S. , 2003: Decomposition of sub-arctic plants with differing nitrogen economies: a functional role for hemiparasites. Ecology , 84: 3209–3221.
- Raab, T. K. , Lipson, D. A. , and Monson, R. K. , 1996: Non-mycorrhizal uptake of amino acids by roots of the alpine sedge Kobresia myosuroides: implications for the alpine nitrogen cycle. Oecologia , 108: 488–494.
- Raab, T. K. , Lipson, D. A. , and Monson, R. K. , 1999: Soil amino acid utilization among species of the Cyperaceae: plant and soil processes. Ecology , 80: 2408–2419.
- Robinson, D. , 2001: δ15N as an integrator of the nitrogen cycle. Trends in Ecology and Evolution , 16: 153–162.
- Schmidt, I. K. , Jonasson, S. , Shaver, G. R. , Michelsen, A. , and Nordin, A. , 2002: Mineralization and distribution of nutrients in plants and microbes in four arctic ecosystems: responses to warming. Plant and Soil , 242: 93–106.
- Schulze, E.-D. , Chapin, F. S., III , and Gebauer, G. , 1994: Nitrogen nutrition and isotope differences among life forms at the northern treeline of Alaska. Oecologia , 100: 406–412.
- Shearer, G. , and Kohl, D. H. , 1986: N2-fixation in field settings: estimation based on natural 15N abundance. Australian Journal of Plant Physiology , 13: 699–756.
- Smith, S. E. , and Read, D. J. , 2008: Mycorrhizal Symbiosis. 3rd edition. San Diego, California: Academic Press.
- Soudzilovskaia, N. A. , and Onipchenko, V. G. , 2005: Experimental investigation of fertilization and irrigation effects on an alpine heath, Northwest Caucasus, Russia. Arctic, Antarctic, and Alpine Research , 37: 602–610.
- Soudzilovskaia, N. A. , Onipchenko, V. G. , Cornelissen, J. H. C. , and Aerts, R. , 2005: Biomass production, N:P ratio and nutrient limitation in a Caucasian alpine tundra plant community. Journal of Vegetation Science , 16: 399–406.
- Soudzilovskaia, N. A. , Aksenova, A. A. , Makarov, M. I. , Onipchenko, V. G. , Logvinenko, O. A. , ter Braak, C. J. F. , and Cornelissen, J. H. C. , 2012: Legumes affect alpine tundra community composition via multiple biotic interactions. Ecosphere, 3: article 33, doi: http://dx.doi.org/10.1890/ES11-00357.1.
- Sprent, J. I. , 2005: Biological nitrogen fixation associated with angiosperms in terrestrial ecosystems. In BassiriRad, H. (ed.), Nutrient Acquisition by Plants: An Ecological Perspective. Ecological Studies, vol. 181. Berlin, New York: Springer, 89–115.
- Taylor, A. F. S. , Högbom, L. , Högberg, M. , Lyon, A. J. E. , Näsholm, T. , and Högberg, P. , 1997: Natural 15N abundance in fruit bodies of ectomycorrhizal fungi from boreal forests. New Phytologist , 136: 713–720.
- Wallander, H. , Göransson, H. , and Rosengren, U. , 2004: Production, standing biomass and natural abundance of 15N and 13C in ectomycorrhizal mycelia collected at different soil depths in two forest types. Oecologia , 139: 89–97.
- Yun, S.-I. , Ro, H.-M. , Choi, W.-J. , and Chang, S. X. , 2006: Interactive effects of N fertilizer source and timing of fertilization leave specific N isotopic signatures in Chinese cabbage and soil. Soil Biology and Biochemistry , 38: 1682–1689.
- Zimmer, K. , Hynson, N. A. , Gebauer, G. , Allen, E. B. , Allen, M, F. , and Read, D. J. , 2007: Wide geographical and ecological distribution of nitrogen and carbon gains from fungi in pyroloids and monotropoids (Ericaceae) and in orchids. New Phytologist , 175: 166–175.