
Abstract
The mite fauna of patches of High Arctic moss-grass tundra of Svalbard in Petuniabukta, Billefjord (moss, grasses, Salix polaris, bare soil), and adjacent to Vestpynten, Adventfjord (moss, moss mixed with grasses, Cassiope tetragona, S. polaris), were investigated. Our aim was to describe the mite fauna of this tundra with particular focus on the stage structure that is rarely investigated. We observed that the oribatid mites were distinctly more abundant and richer in species (22) than the gamasid mites (7) and their density and diversity varied between vegetation and location. Species diversity of Oribatida and Gamasida and the Shannon H′ index of mite communities were low, and Liochthonius sellnicki or Tectocepheus velatus dominated the Oribatida and Oppiella translamellata and Diapterobates notatus were relatively abundant, while the Gamasida were dominated by Arctoseius multidentatus, with Zercon forsslundi and Z. solenites relatively abundant. The juvenile densities of the Oribatida were usually greater than adults. Eniochthonius minutissimus and Metabelba sp. are new records for Svalbard.
Introduction
The High Arctic archipelago of Svalbard has a quite diverse invertebrate community and is among the best known for any region of the Arctic (CitationHodkinson, 2013). Bayartogtokh et al. (Citation2011) presented 81 species of oribatid mites for the archipelago, while Coulson et al. (Citation2014) gave 29 species of gamasid mites. However, considerable taxonomic confusion has accumulated during the 150-year history of mite research in Svalbard (CitationÁvila-Jiménez et al., 2011; CitationBayartogtokh et al., 2011; CitationCoulson et al., 2014), during which microscopic techniques and the taxonomy of these small arthropods has progressed greatly. Ávila-Jiménez et al. (Citation2011) considered critically the total list of 27 species then recorded from Svalbard archipelago and, based on an assessment of fresh soil samples collected between 2007 and 2009 from throughout Svalbard and from recent publications, they reduced this list to 22 species, since increased by subsequent material from new field collections to 29 (CitationCoulson et al., 2014, and references therein). A similar problem concerns the oribatid mites. It is unknown what level of taxonomic identification was used for many records of these mites, so potential synonyms exist, and since the original material has often been lost, this group is also in need of further investigation from fresh sampling. Moreover, ecological studies in Svalbard are generally lacking, except for Seniczak and Plichta (Citation1978) and Coulson et al. (Citation2003), who described the oribatid mites from their own microhabitats. The lists of species presented by other authors give only general information on the presence of species, including accidental ones that can be easily transported from quite distant habitats by birds (CitationLebedeva and Krivolutskiy, 2003; CitationLebedeva and Lebedev, 2008; CitationCoulson et al., 2009).
The ecology of oribatid mites in polar ecosystems such as Svalbard is important to study as these systems are unique and created by characteristic environmental conditions, mainly low annual temperature and precipitation, short summer, and a long, dark winter. The soil is frozen for the greater part of the year and therefore is still in an early stage of development, offering plants only a thin, nutrient-poor organic soil layer. Therefore, the tundra flora is composed of robust species, such as lichens, moss, grasses, and herbs (CitationJónsdóttir, 2005), which grow and develop in a short summer when the upper soil horizon is thawed and the soil fauna is active, decomposing the soil organic matter and releasing nutrients for plant growth. Some tree species also occur on the Svalbard tundra, but because of the High Arctic environment they are small with a prostrate growth form. Moreover, the extreme Arctic environment delays the succession of plants, and the community composition develops slowly, resembling the initial stages of plant succession near the glacier forelands observed in moderate climates. Finally, this tundra ecosystem is composed of clearly differentiated habitats, often at small scales of a few meters (CitationJónsdóttir, 2005), enabling clearly distinguishable soil mite communities to be readily identified (CitationGwiazdowicz and Coulson, 2011).
In tundra mite communities, the saprophagous oribatid mites usually dominate and their densities depend highly on the spatial distribution of plants and the thickness of organic matter (CitationSeniczak and Plichta, 1978; CitationCoulson et al., 2003). As most plant species are distributed patchily, the mites also form aggregations, often displaying a relationship between plant and oribatid mite species. As with the flora, the oribatid mites of the tundra are composed mainly of pioneer and ubiquitous species that are well adapted to the polar climate. The gamasid mites of Svalbard are less known Citation(Ávila-Jiménez et al., 2011), being often omitted from studies involving other soil taxa, mainly due to the difficulties of working with this group (e.g., CitationHodkinson et al., 2003, Citation2004). Therefore, we hypothesize that the plant patches of moss-grass tundra of Svalbard regulate the density, species number, dominance, and stage structure of both oribatid and gamasid mites. Our aim is to describe the oribatid and gamasid mites for moss-grass High Arctic tundra, with particular focus on the stage structure of these mites that is rarely investigated in ecological studies due to the taxonomic challenges.
Study Areas, Material, and Methods
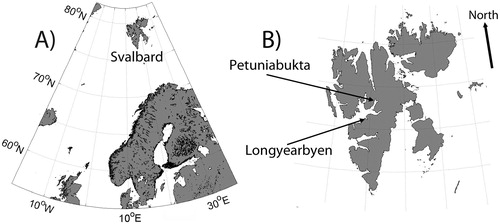
This study was conducted at two locations in central Spitsbergen: Petuniabukta (Billefjord, N78°42′, E16°40′) and Vestpynten close to the Svalbard airport at Vestpynten (N 78°15′, E15°25′), located about 60 km from Petuniabukta (). Both represent poorly developed sandy soils with moss-grass vegetation. In Petuniabukta, the greatest mean monthly air temperature in 2001–2003 was in July and August (+8.1 °C), and the lowest in January to March (-17.8 °C) (CitationRachlewicz and Styszyńska, 2007). At Vestpynten, the warmest monthly mean temperature was in July-August (+7.5 °C) and lowest in January-March (-18.1 °C) (CitationNorwegian Meteorological Institute, 2012). Mean annual precipitation in this region is about 200 mm (CitationRachlewicz, 2003).
Material was collected between 23 July and 7 August 2007 from various vegetation types: Petuniabukta—(1) moss, (2) grasses, (3) Salix polaris, and (4) bare soil (without higher plants and poor in organic matter); Vestpynten—(5) moss, (6) moss mixed with grasses, (7) Cassiope tetragona, and (8) S. polaris. Between one to seven samples of plants and litter (vegetation types 1–3 and 5–8) or bare soil (4), 1 dm2 in area and 5 cm deep in plant vegetation or 5 cm deep in bare soil, were taken (). Replication was partly limited by the strict environmental laws pertaining to Svalbard, which restricted the number of samples due to a slow regeneration time of the High Arctic tundra. Only one sample was taken from small patches of S. polaris and C. tetragona because of the need to limit disturbance to the vulnerable tundra environment.
Mites were extracted from the soil samples in Tullgren funnels in Svalbard, preserved in 70% ethanol, and identified to species, or genus, including all life stages. Names of oribatid species follow Subías (Citation2004) and partly Weigmann (Citation2006). The juveniles of Oribatida include larvae, protonymphs, deutonymphs, and tritonymphs, which were all identified separately using diagnostic characteristics given by Seniczak (Citation1980, Citation1991) and Seniczak and Seniczak (Citation2010a, Citation2012a) as well as undescribed material from existing collections. In total, 11,052 oribatid mites and 265 gamasid mites were collected, and the density presented as individuals m-2. The populations of oribatid and gamasid species were characterized by the density (A) and dominance (D) indices (CitationOdum, 1982), while the communities were described by total abundance and species number and compared by Shannon H′ index and Pielou and Hurlbert evenness (CitationBeisel et al., 2003). Further statistical analysis was precluded by the lack of necessary replication. We used the classes of dominance after Seniczak (Citation1978), including superdominants (D > 40), which are typical of extreme microhabitats.
FIGURE 1. Investigated regions of Svalbard (Spitsbergen). (A) Location of Svalbard in the Arctic Ocean; (B) locations of the town of Longyearbyen (close to Vestpynten) and Petuniabukta.
Stage structure is given for all species for which the juvenile stages are known, except for the Oppiidae and Suctobelbidae, which have soft juveniles that are poorly extracted from soil samples and which would distort the observed stage structure (CitationSeniczak, 1978). Investigation of the stage structure of the small oribatid mite, Liochthonius sellnicki (Thor, 1930) was possible because it was the only member of Brachychthoniidae in moss-grass tundra. All material identified is deposited at the University of Technology and Life Sciences, Bydgoszcz, Poland (Oribatida), or University of Life Sciences, Poznań, Poland (Gamasida), and is available for future comparisons.
Results
DENSITY OF ORIBATIDA AND GAMASIDA, SPECIES NUMBER OF MITES, AND SHANNON AND EVENNESS INDICES
The Oribatida achieved the highest density in moss mixed with grasses at Vestpynten; their density was relatively high in patches of Cassiope tetragona and Salix polaris in Petuniabukta but was low in moss, grasses, and bare soil (, ). Generally, Vestpynten was inhabited by more abundant mites than Petuniabukta, but the density of mites depended on plant cover and which oribatid species dominated in the communities.
TABLE 1 Habitat (number of samples), mean density (thousand individuals m-2), species number (S) of Oribatida and Gamasida, Shannon H′ index and Pielou and Hurlbert evenness of communities of these mites in chosen microhabitats of moss-grass tundra in Svalbard.
Tectocepheus velatus (Michael, 1880) was distinctly more abundant at Vestpynten, whereas Liochthonius sellnicki achieved greatest densities at Petuniabukta (). The mean density of L. sellnicki at Petuniabukta was 10,600 individuals m-2, and at Vestpynten it was 1.7-fold greater than at Petuniabukta. Liochthonius sellnicki was sensitive to plant cover, being the most abundant in patches of C. tetragona and moss mixed with grasses, but less abundant in moss and S. polaris, and in other plant patches densities were low. The mean density of T. velatus at Vestpynten was 9300 individuals m-2, while near Petuniabukta it was 1.9-fold higher than at Vestpynten. Tectocepheus velatus was the most abundant in a patch of S. polaris in Petuniabukta, and decreased in density in the C. tetragona vegetation, while in other patches it was not abundant.
FIGURE 2. Density of Oribatida [thousand individuals m-2 in logarithmic scale (ln)] in investigated microhabitats of moss-grass tundra in Svalbard.
![FIGURE 2. Density of Oribatida [thousand individuals m-2 in logarithmic scale (ln)] in investigated microhabitats of moss-grass tundra in Svalbard.](/cms/asset/cd676a05-2e90-4776-aa31-fa937c1a1789/uaar_a_11957762_f0003.jpg)
In both regions Oppiella translamellata (Willmann, 1923) and Diapterobates notatus (Thorell, 1871) were relatively abundant and common, but the densities of each species were respectively 3.4-fold and 6.2-fold greater in Petuniabukta than at Vestpynten (). Oppiella translamellata preferred moss mixed with grasses at Vestpynten and was relatively abundant in other plant patches in this region, whereas at Petuniabukta this species was not abundant. Diapterobates notatus achieved maximum density in moss mixed with grasses at Vestpynten. This species was relatively abundant in patches of S. polaris at Petuniabukta and C. tetragona and moss at Vestpynten, but in other vegetation patches densities were low.
The remaining species—for example, Camisia horrida Hermann, 1804, Ceratoppias phaerica (L. Koch, 1879), Hermannia reticulata, and Oribatula cf. tibialis (Nicolet, 1855)—were more abundant at Vestpynten than Petuniabukta (). The two former species preferred moss mixed with grasses, H. reticulata was most abundant in patches of S. polaris, while O. cf. tibialis was most abundant in moss.
In both regions, the gamasid mites were distinctly less abundant than oribatid mites, being more abundant at Petuniabukta than at Vestpynten. For example, Zercon forsslundi Sellnick, 1958, occurred exclusively in moss in the former region. Zercon solenites Haarløv, 1942, was the most abundant gamasid mite in bare soil, and Arctoseius multidentatus Evans, 1955, preferred a patch of S. polaris at Petuniabukta ().
The diversity of oribatid mites depends on the type of vegetation and the number of replicated samples. Greatest species diversity occurred in moss in Petuniabukta, while the poorest habitat in terms of species number was in bare soil and S. polaris from the same region (). Moss in Petuniabukta was not rich in oribatid mites, but because of seven replicates the total species number was the highest. Also, bare soil in the same region was inhabited by a small population of oribatid mites, but probably because of the four replicates there was one oribatid species more than in the patch of S. polaris, where the density of these mites was nearly 50-fold higher than in bare soil.
TABLE 2 Mean density (thousand individuals m-2) of species of Oribatida (O) and Gamasida (G) in chosen microhabitats of moss-grass tundra in Svalbard. Oribatid species with a greatest density < 4.0 thousand individuals m-2, and gamasid species with a greatest density < 0.4 thousand individuals m-2 are listed below the table. Ad = adult; Juv = juveniles.
The diversity of the Gamasida was also distinctly lower than that of the Oribatida. The richest in gamasid species was the S. polaris community in Petuniabukta, and the poorest was the bare soil in this region (). The number of gamasid species also depended on the number of replicates, except for bare soil, where only one species was found in four samples. In all patches of moss-grass tundra, the Oribatida and Gamasida occurred together, and the greatest diversity of Gamasida occurred in the samples with the greatest oribatid species diversity.
The Shannon H′ index of mite communities of Oribatida and Gamasida in patches of moss-grass tundra was generally low and depended mainly on the species richness of mites. The index achieved the highest value in moss in Petuniabukta and the lowest value in bare soil from this region (). The Pielou and Hurlbert evenness indices were greatest in grasses and the lowest in a patch of C. tetragona.
DOMINANCE STRUCTURE OF MITES
In patches of moss-grass tundra, the greatest dominance index achieved was by either L. sellnicki or T. velatus (). The former species dominated in all plant patches, except for grasses and S. polaris in Petuniabukta where the latter species was the most abundant. In both regions, D. notatus and O. translamellata were relatively abundant and usually occupied higher dominance classes (eudominants or dominants) at Petuniabukta than at Vestpynten (dominants and subdominants) mainly because of lower dominance indices of L. sellnicki in the former region.
Table 3 Dominance structure of Oribatida (O) and Gamasida (G) mite communities in chosen microhabitats of moss-grass tundra in Svalbard. Dominance index of species is given in parentheses.
Among the non-abundant gamasid mites, Z. solenites was a eudominant in bare soil, Z. forsslundi a subdominant in moss, and A. multidentatus a recedent in moss, grass, and S. polaris in Petuniabukta (). At Vestpynten, only A. multidentatus was a recedent in a patch of S. polaris, while other species were subrecedents.
JUVENILE STAGES OF ORIBATIDA AND GAMASIDA
The patches of moss-grass tundra were generally rich in juvenile oribatid mites, but the proportion of juveniles was higher at Petuniabukta than at Vestpynten (). In most populations of oribatid species, all juvenile instars were relatively abundant, including the larvae (). For C. sphaerica, H. reticulata, D. notatus, and T. velatus the number in a particular development stage usually decreases from the larva to tritonymph.
The stage structure of L. sellnicki depended greatly on the vegetation (). In patches of S. polaris (Petuniabukta) and moss mixed with grasses (Vestpynten), the juveniles made up over 80% and 70% of population of this species, respectively, while in patches of C. tetragona and S. polaris at Vestpynten, the participation of juveniles was smaller (about 40% and 50% of population, respectively). In contrast, the stage structure of T. velatus depended on the location; in Petuniabukta the participation of juveniles was distinctly higher (70%–91%) than at Vestpynten (53%–73% of total populations). In some species, such as D. notatus, C. horrida, and C. sphaerica, the participation of juveniles in populations was also high (54%–88%, 72%–76%, and 60%–64%, respectively), whereas the participation of juveniles of O. translamellata in both regions was low (<10%). Generally, the mean proportion of juveniles of the larger species of oribatid mites (C. horrida, D. notatus, and H. reticulata) were greater (74%, 70%, and 84%, respectively) than that of small species, such as T. velatus and L. sellnicki (63% and 52%, respectively).
TABLE 4 Stage structure of some Oribatida (O) and Gamasida (G) species in chosen microhabitats of moss-grass tundra in Svalbard. LV = larva, PN = protonymph, DN = deutonymph, TN = tritonymph, Np = not present. Density in thousand individuals m-2 is given.
The proportion of juveniles of gamasid mites was generally low, except for A. multidentatus in a patch of S. polaris at Petuniabukta, where the juveniles were slightly more abundant than the adults ().
Discussion
The densities of oribatid and gamasid mites in moss-grass tundra was highly heterogeneous (2300-208,700 individuals m-2), and were probably related to the patchy vegetation cover. In mite communities, the Oribatida dominated. These are mainly saprophages and therefore their density strongly depends on quantity and quality of the organic matter. Some species of oribatid mites achieved different densities in plant patches with similar appearances, raising interesting questions about microhabitat requirements among species. For example, the density of Liochthonius sellnicki was similar in patches of Salix polaris in the two investigated locations, while that of Tectocepheus velatus and Diapterobates notatus was distinctly higher in Petuniabukta than at Vestpynten, although all three species extract well from soil cores. Answering these questions requires more microhabitat data, including those linking biotic distributions to soil chemistry (CitationMagalhães et al., 2012) and at a molecular level (CitationStevens and Hogg, 2006).
The oribatid mite communities of moss-grass tundra were poor in species with a low Shannon H′ index, similar to mite communities from Mediterranean areas (CitationSeniczak and Seniczak, 2006, Citation2010b, Citation2012b; CitationSeniczak et al., 2009, Citation2011, Citation2012) and indicates a low soil fertility (CitationThienemann, 1939). These biocenotic principles themselves refer to aquatic ecosystems, but they also hold true for land vascular plants and the oribatid mites (CitationPuchalski and Prusinkiewicz, 1975), where the density and species diversity depend greatly on the amount and form of plant litter. However, while those communities in the Mediterranean region are stressed by a high temperature and low precipitation, the oribatid mite communities of moss-grass tundra are limited by a low temperature and high soil moisture, which is consistent with the Shelford principle (CitationShelford, 1931) that either a lack or excess of particular environmental factors, such as temperature and moisture, can limit the development of mite communities.
Liochthonius sellnicki or T. velatus dominated in the Svalbard soils and Oppiella translamellata and D. notatus were relatively abundant; all species are typical of tundra, and most of them were abundant in moss-lichen-tundra (CitationSeniczak and Plichta, 1978; CitationCoulson et al., 2003). However, the oribatid mite communities investigated here differ slightly from those investigated by Seniczak and Plichta (Citation1978) by greater numbers of D. notatus in the latter tundra. The oribatid mite communities of moss-grass tundra have similarities to those inhabiting the glacial foreland zones at Hardangerjøkulen (Norway) (CitationSeniczak et al., 2006). In the youngest moraine, T. velatus and L. sellnicki dominated, and their densities decreased along with the age of moraines (CitationHågvar et al., 2009).
An interesting habitat for further examination is the bare soil in Petuniabukta, which was poor in organic matter, but was inhabited by two oribatids, L. sellnicki and D. notatus, and a gamasid, Z. solenites. For all three species the juveniles were present, which suggests that these species develop there, although immigration from neighboring vegetated patches cannot be excluded.
Juveniles dominated adult densities in most of the oribatid species studied, and the mean proportion of juveniles in the larger species of oribatid mites was higher than in small species, which can be explained by shorter time of development of small mites. Tectocepheus velatus is a small (the adult is approximately 0.3 mm) and ubiquitous species with a rather short time of development. In temperate climates, it has three to five generations per year Citation(Niedbała, 1980), but in the High Arctic its development is probably much slower. The larger Arctic oribatid species Ameronothrus lineatus (Thorell, 1871) requires 5 years or more to reach maturity (CitationSøvik et al., 2003). The juveniles of some species, such as O. translamellata, have a thin cuticle and soft body and do not survive the extraction process (CitationSeniczak, 1978), which explains the lower juvenile-to-adult ratios observed for this species.
A high juvenile-to-adult ratio for oribatid mites increases nutrient cycling because these stages transform organic matter more actively than the adults (CitationBerthet, 1963), probably due to more abundant and active gut microflora that is able to decompose even decay-resistant substances, such as cellulose, lignin, and chitin (CitationStefaniak and Seniczak, 1976, Citation1981). In temperate climates, the juveniles of soil oribatid mites usually form one-third of the total population of these mites (CitationNiedbała, 1980), but in oribatid mite communities that are species-poor, the juveniles are often more abundant than the adults (CitationSeniczak and Seniczak, 2006, Citation2010b, Citation2012b; CitationSeniczak et al., 2009, Citation2011, Citation2012), such as observed in this study. With the lack of lumbricid earthworms and generally low diversity of Enchytraeidae in Svalbard, the role of the microarthropod fauna in nutrient cycling may be enhanced (CitationCoulson et al., 2014).
In the regions of Petuniabukta and Vestpynten, the seven gamasid species were relatively common and are all typical for tundra vegetation (CitationGwiazdowicz and Gulvik, 2008; CitationÁvila-Jiménez et al., 2011; CitationGwiazdowicz and Coulson, 2011). Their density was low, so it is difficult to identify microhabitat preferences. Most gamasid mites are considered to be predators of small invertebrates, including the oribatid mites, and although the immediate connections between particular species of Gamasida and Oribatida are poorly known, the occurrence of the greatest gamasid diversity in the more diverse communities of Oribatida studied here possibly reflects the trophic relation between these groups. Thus, the presence of gamasid mites in tundra undoubtedly increases the activity of soil biology. Eniochthonius minutissimus (Berlese, 1903) and Metabelba sp. are recorded for the first time from this archipelago. Both taxa are considered cosmopolitan (CitationSubías, 2004; CitationWeigmann, 2006).
Conclusions
We observed that (1) the oribatid mites were distinctly more abundant and richer in species than the gamasid mites, and their density and diversity varied between vegetation and location; (2) diversity of Oribatida and Gamasida and the Shannon H′ index of mite communities were low; (3) Liochthonius sellnicki or T. velatus dominated the Oribatida, and O. translamellata and D. notatus were relatively abundant, while the Gamasida were dominated by A. multidentatus, with Z. forsslundi and Z. solenites relatively abundant; and (4) juvenile densities of the Oribatida were usually greater than adults.
Acknowledgments
We thank the anonymous reviewers for the thorough review of this manuscript and all suggestions that were very professional and considerably improved the value of this paper. We also thank the Associate Editor, Prof. Scott Armbruster, for all editorial suggestions.
Related Research Data
References Cited
- Ávila-Jiménez, M. L. , Gwiazdowicz, D. J. , and Coulson, S. J. , 2011: On the gamasid (Acari: Parasitiformes) mite fauna of Svalbard: a revised checklist of a High Arctic archipelago. Zootaxa , 3091: 33–41.
- Bayartogtokh, B. , Schatz, H. , and Ekrem, T. , 2011: Distribution and diversity of the soil mites of Svalbard, with redescriptions of three known species (Acari: Oribatida). International Journal of Acarology , 37: 467–484.
- Beisel, J.-N. , Usseglio-Polatera, Ph. , Bachmann, V. , and Moreteau, J.-C. , 2003: A comparative analysis of evenness index sensitivity. International Review of Hydrobiology , 88: 3–15.
- Berthet, P. , 1963: Mesure de la consummation d'oxygene des Oribates (Acariens) de la litieredesforêts. In Doeksen, J., and Drift, J. V. D. (eds.), Soil Organisms. Amsterdam: North Holland Publisher, 18–31.
- Coulson, S. J. , 2007: Terrestrial and freshwater invertebrate fauna of the High Arctic Archipelago of Svalbard. Zootaxa , 1448: 41–58.
- Coulson, S. J. , Hodkinson, I. D. , and Webb, N. R. , 2003: Microscale distribution patterns in High Arctic soil microarthropod communities: the influence of plant species within the vegetation mosaic. Ecography , 26: 801–809.
- Coulson, S. J. , Moe, B. , Monson, F. , and Gabrielsen, G. W. , 2009: The invertebrate fauna of High Arctic seabird nests: the microarthropod community inhabiting nests on Spitsbergen, Svalbard. Polar Biology , 32: 1041–1046.
- Coulson, S. J. , Convey, P. , Aakra, K. , Aarvik, L. , Ávila-Jiménez, M. L. , Babenko, A. , Biersma, E. , Boström, S. , Brittain, J. , Carlsson, A. M. , Christoffersen, K. S. , De Smet, W. H. , Ekrem, T. , Fjellberg, A. , Füreder, L. , Gustafsson, D. , Gwiazdowicz, D. J. , Hansen, L. O. , Holmstrup, M. , Kaczmarek, L. , Kolicka, M. , Kuklin, V. , Lakka, H-K. , Lebedeva, N. , Makarova, O. , Maraldo, K. , Melekhina, E. , Ødegaard, F. , Pilskog, H. E. , Simon, J. C. , Sohlenius, B. , Solhøy, T. , Søli, G. , Stur, E. , Tanaevitch, A. , Taskaeva, A. , Velle, G. Zawierucha, K. , and Zmudczyńska-Skarbek, K. , 2014: The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya. Soil Biology and Biochemistry , 68: 440–470.
- Gwiazdowicz, D. J. , and Coulson, S. J. , 2011: High Arctic gamasid mites (Acari, Mesostigmata); community composition on Spitsbergen, Svalbard. Polar Research , 30: 1–7.
- Gwiazdowicz, D. J. , and Gulvik, M. E. , 2008: Mesostigmatid mites (Acari, Mesostigmata) in Svalbard. In Materiały, Międzynarodowego Sympozjum Polarnego, 32nd, Wrocław, Poland: 32–34.
- Hågvar, S. , Solhøy, T. , and Mong, C. E. , 2009: Primary succession of soil mites (Acari) in a Norwegian glacier foreland, with emphasis on oribatid mites. Arctic, Antarctic, and Alpine Research , 41: 219–227.
- Hodkinson, I. D. , 2013: Terrestrial and freshwater invertebrates. In Meltofte, H. (ed.), Arctic Biodiversity Assessment. Status and Trends in Arctic Biodiversity. Akureyri, Iceland: Conservation of Arctic Flora and Fauna, 193–223.
- Hodkinson, I. D. , Coulson, S. J. , and Webb, N. R. , 2003: Community assembly on proglacial chronosequences in the High Arctic: vegetation and soil development in north west Svalbard. Journal of Ecology , 91: 651–653.
- Hodkinson, I. D. , Coulson, S. J. , and Webb, N. R. , 2004: Invertebrate community assembly along proglacial chronosequences in the High Arctic. Journal of Animal Ecology , 73: 556–568.
- Jónsdóttir, I. S. , 2005: Terrestrial ecosystems on Svalbard: heterogeneity, complexity and fragility from an Arctic island perspective. Proceedings of the Royal Irish Academy , 105: 155–165.
- Lebedeva, N. V. , and Krivolutskiy, D. A. , 2003: Birds spread soil microarthropods to Arctic Islands. Doklady Biological Sciences , 391: 392–332.
- Lebedeva, N. V. , and Lebedev, V. D. , 2008: Transport of oribatid mites to the polar areas by birds. In Bertrand, M. , et al. (eds.), Integrative Acarology. Proceedings of the 6th European Congress. Montpellier: European Association of Acarologists, 359–367.
- Magalhães, C. , Stevens, M. I. , Cary, S. C. , Ball, B. A. , Storey, B. C. , Wall, D. H. , Türk, R. , and Ruprecht, U. , 2012: At limits of life: multidisciplinary insights reveal environmental constraints on biotic diversity in continental Antarctica. PloS One, 7(9): e44578.
- Niedbała, W. , 1974: Studies on the family Brachychthoniidae. Seria biologia, 4. Poznań, Poland: Adam Mickiewicz University Press,1–65 (in Polish).
- Niedbała, W. , 1980: Oribatida-The mites of terrestrial ecosystems. Warszawa: Państwowe Wydawnictwo Naukowe, 255 pp. (in Polish).
- Norwegian Meteorological Institute , 2012: eKlima, <http://www.eklima.no>, accessed July 2012.
- Odum, E. P. , 1982: Fundamentals of Ecology. Warszawa: Państwowe Wydawnictwo Rolnicze i Leśne, 551 pp. (in Polish).
- Puchalski, T. , and Prusinkiewicz, Z. , 1975: Ecological Principles of Forest Habitats. Warszawa: Państwowe Wydawnictwo Rolnicze i Leśne, 463 pp. (in Polish).
- Rachlewicz, G. , 2003: Climate conditions of Petunia Gulf (Middle Spitsbergen) in summer 2000 and 2001. Problemy Klimatologii Polarnej , 13: 127–138 (in Polish).
- Rachlewicz, G. , and Styszyńska, A. , 2007: Comparison of the course of air temperature in Petuniabukta and Svalbard-Lufthavn (Isfjord, Spitsbergen) in the years 2001–2003. Problemy Klimatologii Polarnej , 17: 121–134.
- Seniczak, A. , Solhøy, T. , and Seniczak, S. , 2006: The oribatid mites (Acari, Oribatida) in the Glacier Foreland at Hardangerjokulen (Norway). Biological Letters, Poznań, 43(2): 231–225.
- Seniczak, S. , 1978: Juvenile Stages of Oribatid Mites (Acari, Oribatei) as an Essential Part of Communities of These Mites Transforming the Soil Organic Matter. Toruń: Rozprawy UMK, 171 pp. (in Polish).
- Seniczak, S. , 1980: The morphology of juvenile stages of moss mites of the subfamily Trichoribatinae (Acari: Oribatei), II. Annales Zoologici, Warszawa, 35(15): 221–231.
- Seniczak, S. , 1991: The morphology of juvenile stages of moss mites of the family Camisiidae (Acari: Oribatida), VI. Zoologischer Anzeiger , 227(5/6): 331–342.
- Seniczak, S. , and Plichta, W. , 1978: Structural dependence of moss-mites populations (Acari: Oribatei) on patchiness of vegetation in moss-lichen tundra at the north coast of Hornsund, West Spitsbergen. Pedobiologia , 18: 145–152.
- Seniczak, S. , and Seniczak, A. , 2006: The oribatid mites (Acari) of some habitats of Rhodes Island (Greece). Biological Letters, Poznań, 43 (2): 215–219.
- Seniczak, S. , and Seniczak, A. , 2010a: Differentiation of body form of Gustavioidea (Acari: Oribatida) in the light of ontogeny of three species. Zoologischer Anzeiger , 249: 95–112.
- Seniczak, S. , and Seniczak, A. , 2010b: Oribatid mites (Acari, Oribatida) of various habitats in southern Andalusia (Spain). Biological Letters, Poznań, 47(1): 29–35.
- Seniczak, S. , and Seniczak, A. , 2012a: Differentiation of external morphology of Oribatulidae (Acari: Oribatida) in light of the ontogeny of three species. Zootaxa , 3184: 1–34.
- Seniczak, S. , and Seniczak, A. , 2012b: Oribatid mites (Acari, Oribatida) of yew, cypress and pine litter in southern Italy. Biological Letters, Poznań, 49(1): 27–34.
- Seniczak, S. , Kaczmarek, S. , and Seniczak, A. , 2009: Oribatid mites (Acari, Oribatida) of steppe vegetation on cape Tarhankut in Crimea (Ukraine). Biological Letters, Poznań, 46(2): 97–103.
- Seniczak, S. , Kaczmarek, S. , and Seniczak, A. , 2011: Oribatid mites (Acari, Oribatida) of bushy stripes in steppe vegetation of cape Tarhankut in Crimea (Ukraine). Biological Letters, Poznań, 48(2): 177–183.
- Seniczak, S. , Kaczmarek, S. , Seniczak, A. , and Graczyk, R. , 2012: Oribatid mites (Acari, Oribatida) of open and forested habitats of Korčula Island (Croatia). Biological Letters, Poznań, 49(1): 27–34.
- Shelford, V. E. , 1931: Some concepts of bioecology. Ecology , 12(3): 455–467.
- Søvik, G. , Leinaas, H. P. , Ims, R. A. , and Solhøy, T. , 2003: Population dynamics and life history of the oribatid mite Ameronothrus lineatus (Acari: Oribatida) on the High Arctic archipelago of Svalbard. Pedobiologia , 47: 257–271.
- Stefaniak, O. , and Seniczak, S. , 1976: The microflora of the alimentary canal of Achipteria coleoptrata (Acarina, Oribatei). Pedobiologia , 16: 185–194.
- Stefaniak, O. , and Seniczak, S. , 1981: The effect of fungal diet on the development of Oppia nitens (Acari, Oribatei) and on the microflora of its alimentary tract. Pedobiologia , 21: 202–210.
- Stevens, M. I. , and Hogg, I. D. , 2006: Contrasting levels of mitochondrial DNA variability between mites (Penthalodidae) and springtails (Hypogastruridae) from the Trans-Antarctic Mountains suggest long-term effects of glaciations and life history on substitution rates, and speciation processes. Soil Biology & Biochemistry , 38: 3171–3180.
- Subías, L. S. , 2004: Listadosistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) delmundo (1758–2002). Graellsia , 60: 3–305.
- Thienemann, A. , 1939: Grundzüge einer allgemainen Oecologie. Archive für Hydrobiologie , 35: 267–285.
- Weigmann, G. , 2006: Hornmilben (Oribatida). In Dahl, F. (ed.), Die Tierwelt Deutschland und der angrenzenden Meeresteile. Vol. 76. Keltern, Germany: Goecke & Evers, 520 pp.