
Abstract
Novel disturbance regimes (e.g., introduced herbivores and fire) are among the major drivers of degradation in island ecosystems. High-elevation ecosystems (HEEs) on islands might be especially vulnerable to these disturbances due to high endemism. Here, data from an 11-year exclosure experiment in the HEE of La Palma (Canary Islands) are presented where mammalian herbivores have been introduced. We investigate the combined effect of herbivory and fire on total species richness, seedling richness, and seedling establishment on the whole system and a subset of highly endangered species (target species). Total species richness, seedling species richness, and seedling establishment decreased with herbivory. Five out of eight target species were exclusively found inside the exclosures indicating the negative impact of introduced herbivores on endemic high elevation flora. Target species were generally affected more negatively by introduced herbivores and were subject to significantly higher browsing pressure, probably owing to their lack of defense strategies. A natural wildfire that occurred six years before data sampling substantially increased total species richness and seedling richness in both herbivory exclosure and reference conditions. We conclude that species composition of the HEE has been severely altered by the introduction of non-native herbivores, even though fire seems to have a positive effect on this system.
Introduction
Anthropogenic influences have created novel disturbances as well as altered natural disturbance regimes (CitationWhite and Jentsch, 2001; CitationHobbs et al., 2006; CitationCatford et al., 2012). Indeed, changes in disturbance regimes are widely recognized as major threats to island biota and ultimately to global biodiversity (CitationCaujapé-Castells et al., 2010). Mammalian herbivores (CitationCourchamp et al., 2003; CitationCampbell and Donlan, 2005), altered fire regimes (CitationPerry et al., 2012; CitationRamirez et al., 2012), along with land use change, habitat degradation, or climate change (CitationCaujapé-Castells et al., 2010) are of special importance among the threats to native and endemic island biota and insular ecosystems in general. Conservation efforts on islands should have particularly high-elevation (CitationKier et al., 2009) because of the outstanding position of islands as hotspots of endemic plant species diversity (CitationKreft et al., 2008).
A high degree of endemic species (CitationSteinbauer et al., 2012) and their ephemeral evolutionary position (CitationFernández-Palacios et al., 2014) characterize especially high-elevation ecosystems (HEEs) within islands. These ecosystems experience strong environmental (due to increasing climatic harshness with elevation) and geographical isolation (because of increasing distance to comparable ecosystems on other islands or the continent; CitationSteinbauer et al., 2013). As the ocean is an effective filter for terrestrial mammals, most oceanic islands did not possess mammalian herbivores before human colonization. Thus, mammalian herbivory has not influenced the evolution of endemic species on islands (CitationTraveset et al., 2009). As a consequence, endemic plant species have often lost the particular herbivore defense mechanisms and strategies of their continental ancestors due to this lack of selection pressure (CitationAtkinson, 2001). Indeed, evolutionary effects of missing herbivore pressure on plant traits can become apparent even within only decades (CitationDidiano et al. 2014).
Introduced herbivores often profit from the absence of natural enemies, diseases, and pests (CitationShea and Cheeson, 2002). Thus, introduced mammalian herbivores have not only inflicted damage in the past but continue to inflict great damage on natural island vegetation and endemic plant species (e.g., CitationCampbell and Donlan, 2005; CitationCaujapé-Castells et al., 2010; CitationIrl et al., 2012; CitationRamirez et al., 2012). Important examples are the feral goat (Capra hircus) and the European rabbit (Oryctolagus cuniculus), both considered among the world's 100 most invasive species (CitationLowe et al., 2000).
As an additional driver of change and degradation, human-induced alterations of fire regime (i.e., increased fire frequency or magnitude) can negatively affect island vegetation, even though it may be naturally adapted to occasional wildfires (CitationArévalo et al., 2001; CitationPerry et al., 2012, CitationGarzón-Machado et al., 2012; CitationCrausbay et al., 2014). Especially combined changes in both disturbances, introduced mammalian herbivores and altered fire regimes, will likely create negative impacts on vegetation dynamics or species richness. Population structure and community composition can be substantially modified via reduced regeneration success after fire events (CitationJohansson et al., 2010; CitationRamirez et al., 2012).
In contrast to lower elevation ecosystems, HEEs in oceanic islands are still believed to be largely in a natural or at least semi-natural state. This is reflected in different forms of nature reserves and protection status. High-elevation nature reserves have been established, for instance, on La Palma and Tenerife (Canary Islands, Spain), Pico (Azores, Portugal), Fogo (Cape Verdes), La Réunion (Mascarene Islands, France), and Hawaii and Maui (Hawaii Islands, United States). The exclusion of intensive land use, settlements and infrastructure and the limited direct human impact in general makes it possible to investigate the dynamics of introduced herbivore pressure and altered fire regimes.
La Palma is a subtropical volcanic-oceanic island with strong environmental gradients (Irl and CitationBeierkuhnlein, 2011). It is inhabited by several globally relevant introduced mammalian herbivores such as the feral goat (C. hircus) and the European rabbit (O. cuniculus; CitationGarzón-Machado et al., 2010; CitationIrl et al., 2012) but also the recently introduced Barbary sheep (Ammotragus lervia) originally from northern Africa. The selective grazing of these herbivores has been shown to cause the virtual mono-dominance of a single shrub species (i.e., the single-island endemic subspecies Adenocarpus viscosus subsp. spartioides), even though climatically better-adapted shrub species are present in the natural HEE flora (CitationIrl et al., 2012). Fires occur naturally in the subalpine scrub due to the close vicinity to the endemic fire-promoting Canary Pine forest that forms the treeline (CitationHöllermann, 2000; Citationdel Arco Aguilar et al., 2010). Both disturbances, fire and herbivory, have been individually studied on the Canary Islands and specifically on La Palma (see CitationHöllermann, 2000, but also CitationGarzón-Machado et al., 2010, or CitationIrl et al., 2012), but an integrative approach to identify the combined effects of both introduced herbivores and altered fire regime with focus on the unique characteristics of HEEs is still missing.
We aimed at exploring the effects of two major disturbances—herbivory and fire—on the high elevation vegetation of La Palma, Canary Islands. Using an 11-year fenced exclosure experiment and nonfenced counterparts we take the HEEs of La Palma and the transition zone with the Canary Pine forest as a case study. We hypothesize that (1) introduced herbivores negatively affect total species richness, seedling species richness, and seedling establishment; (2) additional fire reduces total species richness, seedling richness, and seedling establishment; and (3) introduced herbivores selectively browse on endemics due to their lack of adaptation to herbivory.
Material and Methods
STUDY AREA
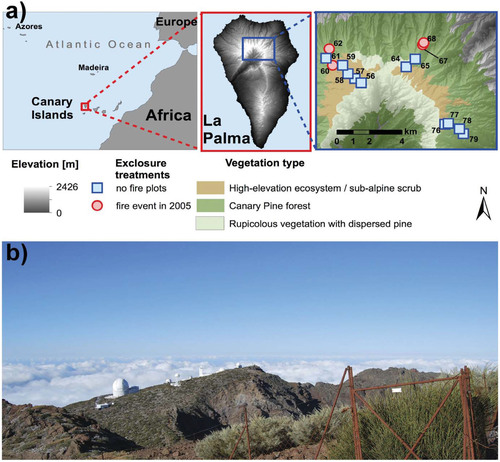
The Canary Islands are situated off the northwestern coast of Africa in the Atlantic Ocean. All islands are of volcanic origin (CitationCarracedo et al., 2002). La Palma is located in the NW of the archipelago. At approx. 1.7 Ma it is the second youngest island, spanning an area of 706 km2 (CitationCarracedo et al., 2002). The HEE of La Palma—also referred to as the subalpine summit scrub (CitationIrl and Beierkuhnlein, 2011; CitationIrl et al., 2012) or summit broom scrub (CitationGarzón-Machado et al., 2011)—is most prominent on the outer flanks of the Caldera de Taburiente complex, ranging from about 2000 to 2400 m a.s.l. It covers an area of 14.3 km2 (see ). The treeline ecotone consists of a transitional zone of the HEE and the Canary Pine forest and ranges between 1800 and 2000 m a.s.l. The inner cliffs of the caldera are too steep for closed vegetation cover. Above the trade-wind induced cloud bank the climatic conditions are characterized by high solar radiation, extensive arid periods of up to 6 months in summer, high diurnal temperature amplitudes, and harsh winter conditions with snow and ice storms at irregular frequencies and magnitude (CitationGarzón-Machado et al., 2013).
No mammalian herbivores exist or have ever existed naturally on the Canary Islands (CitationTraveset et al., 2009). However, aboriginal inhabitants brought the feral goat (C. hircus) to the Canary Islands more than 2000 years ago as archaeological studies have shown (CitationZöller et al., 2003), whereas the Castilian conquerors introduced the European rabbit (O. cuniculus) in the 15th and 16th century to the archipelago. Today, rabbits are partially controlled by hunting (CitationCabrera-Rodriguez, 2006) but are still relatively abundant throughout the whole island. As recently as the 1970s, the Barbary sheep (Ammotragus lervia) was deliberately introduced to the Caldera de Taburienta National Park (NP) (CitationNogales et al., 2006) by the National Institution for Environmental Conservation (ICONA) to stimulate big game hunting as a new economical field.
Forest fires spread upslope from the fire-adapted Canary Pine forest into the HEE at irregular intervals and intensities (Citationdel Arco Aguilar et al., 2010), generally affecting the lower HEE regions more than the higher ones. Fire frequency has increased in the last decades (CitationCliment et al., 2004; CitationPalomares Martínez et al., 2004; CitationGarzón-Machado et al., 2012).
The HEE of La Palma is dominated by endemic legume shrub species (; Citationdel Arco Aguilar et al., 2010). It has already been shown that, although climatically better adapted legume shrub species such as Genista benehoavensis, Spartocytisus subpranubius, or Chamaecytisus proliferus subsp. proliferus are potentially present, the selective grazing of introduced herbivores strongly favors the legume Adenocarpus viscosus subsp. spartioides (CitationIrl et al., 2012). In order to halt extinction dynamics of several very rare and highly endangered endemic plant species the administration of the Caldera de Taburiente NP has established a variety of conservation measures specifically for these endangered endemics, including large-scale fenced exclosures and massive aerial seed dispersal (CitationPalomares Martínez et al., 2004). These endangered endemic species are considered our target species (). All other species are non-target species.
SAMPLING DESIGN
A paired approach of thirty 20 × 20 m plots (15 pairs) in the presence and absence of introduced herbivores was implemented in this study to assess total species richness, seedling species richness, and establishment success of all vascular plant species of the HEE and transitional zone of La Palma (). The plots are situated outside of the actual Caldera de Taburiente NP but administrated by the park authorities on the outer flanks of the caldera. The first plot of each pair was a fenced exclosure in order to prohibit the access of introduced herbivores (herbivory exclosure plot; fence height 1.20 m), The exclosures were established by the Caldera de Taburiente NP in 2000. During the establishment of the herbivory exclosure plots, all aboveground vegetation was removed and eight target species were sown within this area. Target species were sown in equal quantities per plot but differing quantities per species because germination success varied greatly between species (see experimental germination tests done by CitationPalomares Martínez, 1993). See Appendix A1 for the sown seed quantity per species. We assumed that seeds of species, which were not sown, were present in the soil seed bank as these species are more or less widely distributed throughout the study area.
FIGURE 1. (a) Map of the Canary Islands, La Palma, and the study area. (b) A view of the rim of the Caldera de Taburiente from the highest plot (56) at 2400 m a.s.l. The study area is located on the northern part of the island (a, central panel). The symbols depict the sites of exclosures (herbivory exclosure) and their respective herbivory reference pairs: squares = unaffected by 2005 fire and circle = affected by 2005 fire (created with ArcGIS 10, ESRI Inc.). The species inside the exclosure are two of our target species, i.e. Spartocytisus supranubius (dark green) and Genista benehoavensis (grayish green). Around the telescopes a mono-dominant stand of Adenocarpus viscosus subsp. spartioides can be seen. Photo by Gesche Blume-Werry.
The reference plot of each pair was an unfenced plot permitting full access for introduced herbivores (herbivory reference plot). The herbivory reference plot was situated directly next to the herbivory exclosure counterpart and possessed the same size. It was established under comparable site conditions (inclination, soil conditions, fire regime; in the transitional zone, this also included tree cover, etc.), although vegetation was not removed in the herbivory reference plots. Each herbivory reference plot is situated downslope from its herbivory exclosure pair to ensure effective and continuous seed dispersal from within the herbivory exclosure plots. Dispersal ability differs between species, but the Caldera de la Taburiente NP undertook an additional seed rain in 2006 throughout large parts of the study area due to massive aerial seed dispersal of target species. Most target species reach their reproductive age after 2 to 5 years (CitationPalomares Martínez, 1993), indicating that natural seed dispersal from the herbivory exclosure plots to the herbivory reference plots has been possible for a maximum of 6 to 9 yr.
TABLE 1 Total species list including the abbreviations used in the and , a classification of their endemism status (SIE = single-island endemic; AE = archipelago endemic; other status = native or non-native), growth form, and the species status as target species.
Besides herbivory, a second treatment was present. A wildfire consumed the vegetation of several plot pairs in 2005 (n = 4 herbivory exclosure plots; n = 4 herbivory reference plots), thus enabling the assessment of the combined effects of herbivory and fire on total species richness, seedling species richness, and seedling establishment success (). This resulted in a two-factorial sampling design ().
The plots are distributed throughout the whole HEE reaching from 1800 to 2400 m a.s.l.. They are located at three major aspects (i.e., NW, NE, E) as shown in . The distribution of the plots follows the main elevational and aspect gradients and aims at reaching from the transitional zone to the caldera rim. Data sampling took place in April 2011.
Table 2 All possible combinations of the two-factorial design implemented in our sampling design. The number of plots per treatment type is indicated by “n.”
BIOTIC MEASUREMENTS
Total species richness was recorded within each of the thirty 20 × 20 m plots. All additional biotic measurements took place in 10 × 1 m2 randomly distributed subplots within each of the thirty 20 × 20 m plots (see ). In each of the ten 1 m2 subplots seedling identity and abundance of all seedling individuals per species were recorded. Here, we define seedling as a plant individual that has not yet lignified (except for P. canariensis, where maximum growth height of 10 cm was chosen as threshold, and the herbaceous Lactuca palmensis, which was classified to be juvenile if only juvenile leaves were present). In addition, the absence of flower buds or branch ramifications was mandatory to be considered as seedling. Seedling species richness was calculated by summing up all seedling species. Seedling density (as a measure of seedling establishment success) was calculated by dividing the number of seedlings per species through the total subplot area (i.e., 10 × 1 m2).
In addition, we measured species-specific browsing intensity of introduced herbivores. Rabbits were able to enter temporarily due to storm-associated fence damage (plot # 56, 59, 77, 78; ) in four herbivory exclosure plots. Temporary rabbit presence did not significantly change total species or seedling richness when compared to the other herbivory exclosure plots (Wilcoxon signed rank test). All plant individuals (i.e., seedlings and adults) were used to identify browsing intensity. Browsing intensity was classified by estimating the percentage of total browsed plant tissue ().
STATISTICAL ANALYSIS
For comparison of total species richness (measured on the 20 × 20 m plot level) and seedling species richness (i.e., integrating all subplots of one plot) between herbivore and no herbivore plots, paired Wilcoxon signed rank tests (R-package stats, version 3.1.0) were used. The effect of fire treatments on total species richness and seedling species richness was tested using a mixed-effects model (package lme4 version 1.1.6; CitationBates et al., 2014) with herbivory as random effect.
TABLE 3 Classification of browsing intensity used in .
Seedling density was compared between herbivory exclosure and herbivory reference plots for all species separately by using mixed-effects models. Subplots were grouped within pairs by adding a random site factor. The analysis was repeated for all species combined with the factor “species” as an additional random effect.
Model assumptions for mixed-effects models (normality of residuals, homogeneity of variances among treatments) were tested by visual inspection and were not met in all cases. The models were thus repeated with rank transformed data. Results did not differ qualitatively. In the following, results based on the untransformed data are reported.
Browsing intensity between target species and non-target species was tested using the Mann-Whitney U-test.
Illustrations were created with SigmaPlot 7.0 (CitationSystat Software Inc., 2001) and R version 3.1.0 (CitationR Development Core Team, 2014). The latter was used for all statistical analyses.
Results
EFFECTS OF INTRODUCED HERBIVORES ON SEEDLING ESTABLISHMENT
A total of 24 species was identified in the study sites; 79% (n = 19) of species in the data set were archipelago endemics, 38% La Palma endemics (n = 9; ). The majority of the species were woody perennials (67%).
Total species richness was significantly higher inside the herbivory exclosure plots (7.8 ± 5.1; mean ± standard deviation [SD]), when compared to the herbivory reference plots (4.9 ± 4.2; paired Wilcoxon signed rank test p < 0.01; , part a). The difference is even more distinct for seedling species richness (paired Wilcoxon signed rank test p = 0.002; , part b; herbivory exclosure: 5.1 ± 3.5; herbivory reference: 3.1 ± 3.4).
Plots facing the 2005 fire event were characterized by a significant increase in the total species richness and seedling species richness inside and outside the exclosures (total species richness: 12.3 ± 4.8; seedling species richness: 8.5 ± 2.9) compared to non-fire plots (total species richness: 4.2 ± 2.6; seedling species richness: 2.5 ± 2.1; mixed-effects model p < 0.001 in both cases, ). However, the effect of fire is statistically not distinguishable from vegetation type (summit scrub vs. transition zone) or elevation because all burnt areas are situated in or close to the transition zone with the fire-prone pine forest at lower elevations.
FIGURE 2. (a) Total species richness and (b) seedling species richness varied significantly according to herbivory (herbivory reference = herb; herbivory exclosure = no herb) and fire treatment. Note that the effect of herbivory was tested in a paired t-test (including all plots) and the effect of fire in a mixed-effects model (herbivory as random effect, including all plots).
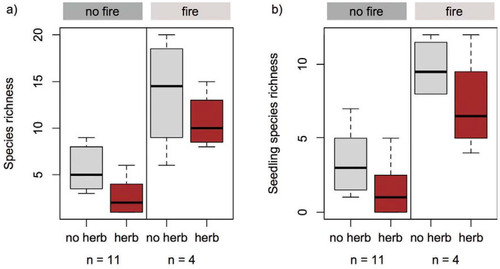
FIGURE 3. Seedling density per target species and nontarget species in the 1 m2 subplots of the herbivory exclosure (black bar; n = 15 plots, 150 subplots) and herbivory reference plots (gray bars; n = 15 plots, 150 subplots). For species abbreviations see . Significant differences (indicated as asterisks with p-values as p < 0.05 = *, p < 0.01 = ** and p < 0.001 = ***) were assessed using mixed effect models on the subplot data comparing each species individually with a random site factor.
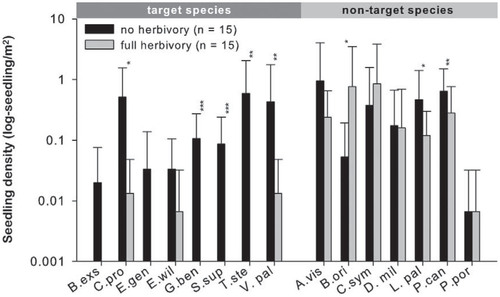
In general, the seedling density (as a proxy for seedling establishment) was higher inside the exclosures than outside (p < 0.001; mixed-effects model on subplot level including all species). On average, the herbivory exclosure subplots have a higher seedling density of 0.33 seedlings per m2 than the herbivory reference subplots. For the majority of target species, this negative effect of herbivory on seedling density was also detectable if analyzed individually (). The same trend is visible for the target species where no significant difference was detectable. However, significance is likely not reached due to very low numbers of individuals. Seedlings of five out of eight target species were exclusively found inside the herbivory exclosure plots ().
SELECTIVE BROWSING ON ENDANGERED ENDEMIC PLANT SPECIES
The target species T. stenopetala and S. supranubius showed highest browsing damage, followed by C. proliferus subsp. proliferus and G. benehoavensis (). Non-target species such as B. origanifolius and L. palmensis showed a high variability in browsing damage, from no visible browsing marks up to serious damage. Herbivores did not browse A. viscosus subsp. spartioides. Target species were significantly more heavily browsed than non-target ones (Mann-Whitney U-test: W = 1067.5, p < 0.001).
Discussion
NEGATIVE IMPACT OF INTRODUCED HERBIVORES ON SEEDLING RICHNESS
Our findings indicate that total species richness, seedling species richness as well as seedling density in insular HEE decreases significantly if introduced herbivores are present (i.e., feral goats, rabbits, or Barbary sheep). Even if plant individuals have reached a certain growth height (i.e., most species of the HEE have a shrubby growth form), rabbit browsing can still cause severe damage, although it remains mostly non-lethal (see ). However, under severe drought conditions when other food sources are lacking, the rabbits can turn to browsing the bark of adult shrub individuals (e.g., G. benehoavensis, C. proliferus subsp. proliferus, B. exstipulata, S. supranubius, T. stenopetala subsp. sericea) even causing the shrub's death (CitationPalomares Martínez et al., 2011).
Both endangered target and the other non-target species are mainly single-island or—at least—archipelago endemics, in that case mostly restricted exclusively to the two corresponding Canarian HEEs (CitationAcebes Ginovés et al., 2010; Citationdel Arco Aguilar et al., 2010). This high degree of endemism is typical for HEEs within an archipelago (CitationFernández-Palacios et al., 2014). However, some of the non-target species such as A. viscosus subsp. spartioides possess a variety of alkaloids possibly functioning as herbivore deterrents (CitationIrl et al., 2012, and references therein), while others are rich in essential oil (e.g. B. origanifolius [CitationEconomou and Nahrstedt, 1991] and P. canariensis [CitationDob et al., 2005]), or other compounds potentially suitable as herbivore deterrents (e.g., C. symphytifolius [CitationCalabuig et al., 1981]). Therefore, these species might have a competitive advantage over the target species. However, recent studies indicate that intraspecific herbivore defense decreases with elevation due to low temperatures and less herbivore pressure (CitationPellissier et al., 2014).
FIGURE 4. Selective grazing indicated by species-specific browsing intensity of introduced herbivores on target and nontarget species present in the herbivory exclosure plot temporarily entered by rabbits. Whiskers indicate the 0.95-confidence interval. Browsing intensity was significantly higher for target species than nontarget species (Mann-Whitney U-test: W = 1067.5, p < 0.001). For browsing classification, see ; for abbreviations, .
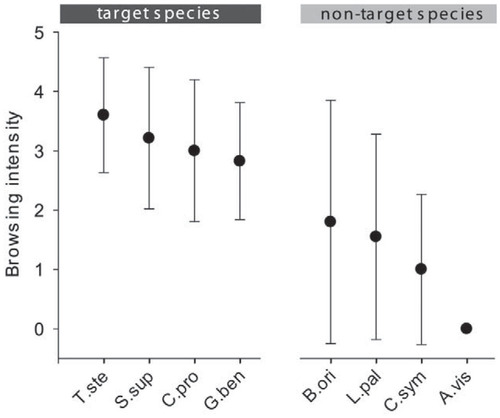
FIGURE 5. Example of selective browsing of introduced herbivores on the single-island endemic Genista benehoavensis. The individual at the top is protected from herbivores situated within an exclosure and displaying a vital and healthy growth (growth height about 3.5 m), while the lower individual of the same species has been heavily browsed, reacting with compensational growth (growth height about 0.5 m). Photos by (a) Severin D.H. Irl and (b) Carl Beierkuhnlein.

Similar to the results given by Garzón-Machado et al. (Citation2010) for the Canary Pine forest of La Palma, a large share of target species was exclusively found inside the exclosures in our study area, indicating that introduced herbivores reduce seedling establishment to nearly zero. On the one hand, it cannot be fully excluded that the sampling design (i.e., total vegetation removal and sowing of target species only in the herbivory exclosure plots but not in the herbivory reference twin) might bias our results. On the other hand, aerial seed dispersal by the NP administration (CitationPalomares Martínez et al., 2004) as well as many years of possible seed rain of target species from inside the exclosures to the herbivory reference plots in direct proximity to the exclosures (CitationPalomares Martínez, 1993) should have ensured the possibility of target species to establish also outside of the exclosures. Obviously, this is not the case. In addition, the nonremoval of vegetation in the herbivory reference plots should actually facilitate seedling establishment during the critical period of establishment compared to the herbivory exclosure plots because the removal of the vegetation in the herbivory exclosure plots increases solar radiation and drought stress, and excludes facilitative effects common for high stress HEEs (CitationAnthelme and Dangles, 2012), all affecting seedling establishment and survival negatively.
Extremely few to no seedlings of the target species were found in the herbivory reference plots, indicating the poor adaptation of island species to introduced herbivory (CitationBowen and Van Vuren, 1997). Interestingly, seedlings of some non-target species (e.g., B. origanifolius) are significantly more abundant or do not differ (A. viscosus subsp. sparitoides, C. symphytifolius, Descurainia millefolia, and Pterocephalus porphyranthus) if introduced herbivores are present, probably profiting from chemical compounds acting as herbivore deterrents, reduced interspecific competition, or increased resource availability outside the exclosures. However, some non-target species were also negatively affected (Lactuca palmensis and P. canariensis).
Our data suggest that the environmental niches made available by reducing plant species diversity through introduced herbivores are filled by native and endemic species, and not by non-native plant species commonly associated with disturbed systems at lower elevations (CitationMerlin and Juvik, 1992). Thus, even though this ecosystem is subject to substantial changes in plant species composition and diversity, there is no indication that non-native species abundant in lower elevations of the Canary Islands (CitationArévalo et al., 2005) are able to fill the available niches. Exceptions are roadside communities often identified as invasion pathways for non-native plant species (CitationArévalo et al., 2005; CitationPauchard et al., 2009) but also for endemics (CitationIrl et al., 2014) into high elevations.
POST-FIRE EFFECTS WITH AND WITHOUT PRESENCE OF INTRODUCED HERBIVORES
Our findings indicate that the legacy of fire has an overall positive effect on total species richness and seedling richness. The negative effects of herbivory seem to be comparable to non-fire conditions. This can result from ameliorated growing conditions owing to the enhanced availability of soil nutrients and the opening of the canopy associated with post-fire conditions, the reduced habitat suitability for introduced herbivores, or population decline of herbivores during the fire (CitationKeeley and Fotheringham, 2000). However, the positive effect on total plant species richness and seedling species richness cannot be separated from elevation or vegetation type.
Herbivore impact is still effective as only 37.5% of the target species that occur in no herbivore conditions were actually recorded in the herbivory reference plots and then only in very low abundances in post-fire conditions. A single shrub species (i.e., A. viscosus subsp. spartioides) creates almost mono-dominant stands if introduced herbivores are present (CitationIrl et al., 2012). Our data suggest that the effect of introduced herbivores acts on the complete species composition of the entire HEE flora. In addition, fire probably has a positive impact on plant species richness, even though the fire regime is considered an anthropogenically altered disturbance regime mainly due to its elevated reoccurrence frequency (CitationGarzón-Machado et al., 2012).
IMPLICATIONS FOR CONSERVATION AND RESTORATION MANAGEMENT
Under natural conditions, that is, in the absence of large mammalian herbivores, other species than A. viscosus subsp. spartioides will probably have played a more important role in the HEE of La Palma, but the reconstruction of pre-human ecosystems remains speculative. Indeed, recent discoveries suggest that a Juniperus cedrus woodland might have existed above the Canary Pine forest, which would have encompassed large parts of the current summit scrub (CitationNogales et al., 2014). Also, even small reductions in species abundance and distributions are likely to have a big impact on the function and structure of species-poor systems such as the HEE of La Palma (sensu the insurance hypothesis; CitationNaeem and Li, 1997).
The definition of a potential natural state of vegetation and ecosystems is difficult if not impossible when environmental conditions and pressures have changed (CitationChiarucci et al., 2010). Garzón-Machado et al. (Citation2011) assumed that G. benehoavensis might be an important species next to A. viscosus subsp. spartioides in this system, which is confirmed by our data. However, other species such as C. proliferus ssp. proliferus and T. stenopetala subsp. sericea, performed well, too. This indicates that these species should also play a vital role in future considerations of conservation and habitat restoration for hypothetical HEEs without introduced herbivores. Also S. supranubius, which is very common in the HEE of Tenerife (CitationKyncl et al., 2006), might be an important species for the highest elevations of La Palma because it seems to be optimally adapted to the harshest HEE conditions (CitationIrl et al., 2012).
Due to their specific growth form and morphology, the La Palma endemics E. gentianoides and E. wildprettii subsp. trichosiphon are not able to dominate the vegetation structure. Yet, their conservation should have top priority because their global distribution is confined to the relatively small area of the HEE of La Palma (CitationAcebes Ginovés et al., 2010; Citationdel Arco Aguilar et al., 2010). Interestingly, A. viscosus subsp. spartioides does not seem to play such a dominant role without introduced herbivores.
All in all, insular HEEs are islands within islands dominated by range-restricted endemics (CitationFernández-Palacios et al., 2014), and therefore these ecosystems and their species should be considered with high conservation priority. This might become especially relevant because additional pressure is likely exerted on the diversity of island HEEs and their endemic species by anthropogenic climate change, which has the potential to increase the extinction risk of these species (CitationGreenwood and Jump, 2014).
STRONG SELECTIVE BROWSING INFLUENCES PLANT COMMUNITY DYNAMICS
In the HEE of La Palma, introduced herbivores selectively browse on the different species present. Selective browsing is characteristic for many mammalian herbivores (e.g., CitationRafferty and Lamont, 2007; CitationTraveset et al., 2009; CitationJohansson et al., 2010). In non-adapted systems, it can induce substantial alterations of plant community dynamics and structure (CitationRamirez et al., 2012). In the HEE of La Palma, several plant species, especially the endangered ones, are heavily browsed, while others are less damaged or even remain completely unaffected. Plant species with defense mechanisms possess a competitive advantage, because unprotected species, even though they are rare, are more likely to be browsed severely than abundant species possessing defense mechanisms (CitationBryant et al., 1991). In our model system on La Palma this phenomenon can be observed as well. Rare species such as T. stenopetala subsp. sericea, S. supranubius, C. proliferus subsp. proliferus, or G. benehoavensis are preferred by introduced herbivores, while common and widespread species are significantly less damaged. Current rarity, however, reflects the long-lasting legacy of introduced herbivore pressure.
We suppose that the vegetation we see today in the HEE of La Palma strongly deviates from the former “natural” vegetation. As HEEs are very isolated, no introduced or invasive plant species has entered these communities. However, shifting dominance patterns and a local decrease of diversity have likely caused impoverished and almost mono-dominant stands of plants that can cope with current herbivore pressure. Today, many species that can be expected to have been abundant under “natural” conditions (i.e., without mammal herbivory) only occur in refugial locations (e.g., on cliffs or rocky outcrops; nowadays also in large exclosures established for conservation purposes by the Caldera de Taburiente NP) and are highly threatened to become extinct (CitationPalomares Martínez et al., 2004).
Very likely the present ecosystems have been altered by centuries of goat herding and cyclic burning. Such kinds of intense past impacts and land use history can have a long-lasting legacy in local plant species occurrence even after its abolishment (CitationHeubes et al., 2011). In addition, the presence of introduced herbivores has been shown to prevent the recuperation of natural vegetation (CitationMerlin and Juvik, 1992).
Conclusion
The results presented from an 11-yr exclosure experiment offer the unique opportunity to identify the long-term impact of introduced mammalian herbivores on an insular HEE in combination with reoccurring fire events. Presumably, the presence of introduced herbivores has substantially altered the species richness and vegetation composition, leading to a strong degradation of this system. This resulted, among others, in a massive coordinated conservation program initiated by the Caldera de Taburiente NP administration to protect the various endangered single-island or archipelago endemics of this system (CitationPalomares Martínez et al., 2004).
Now, the long-term goal should be to preserve the endangered species, create habitats and “safe sites” for these species to expand their current ranges, and, if possible, restore the HEE to a natural state. A necessary step in this direction would be to monitor and strictly control the population of all major herbivores (especially feral goats, rabbits, and Barbary sheep) as well as create awareness among the population about the damages caused by human-induced fires. However, no matter how successful total eradication programs have previously been in enabling the full recovery of natural island vegetation (e.g., CitationCaujapé-Castells et al., 2010, and references therein), it seems unrealistic to achieve this conservation goal on such a large and inhabited island as La Palma that relies heavily on rural and traditional agricultural practices. Nevertheless, total eradication of introduced herbivores from the whole island of La Palma should be the long-term goal from a conservation perspective to sustainably protect the unique native endemic flora and enable its survival and reestablishment. In the meantime, however, more and larger fenced exclosures might be a promising conservation method to preserve the most endangered species and create patches of restored natural vegetation.
Acknowledgments
We would like to thank the employees of the Caldera de Taburiente NP for establishing and maintaining the exclosures during the last 11 years, the Elite study program Global Change Ecology (GCE) within the Elite Network of Bavaria (ENB) for its financial support, and Dr. Felix Medina from the Consejería de Medio Ambiente del Cabildo Insular de La Palma for providing us with helpful information. A special thank you goes to all organizers of the great conference “Vulnerable Islands in the Sky: Science and Management of Tropical Islands Alpine and Sub-alpine Ecosystems” in Waimea, Hawaii, in August 2012 and the financial support provided for the two main authors, enabling them to participate. We would especially like to express our gratitude to the four reviewers for their constructive and knowledgeable comments and remarks.
Related Research Data
References Cited
- Acebes Ginovés, J. R. , León Arencibia, M. C. , Rodríguez Navarro, M. L. , del Arco Aguilar, M. , García Gallo, A. , Pérez de Paz, P. L. , Rodríguez Delgado, O. , Martín Osorio, V. E. , Wildpret de la Torre, W. , 2010: Spermatophyta. In Arechavaleta, M. , Rodríguez, S. , Zurita, N. , and García, A. (eds.), Lista de especies silvestres de Canarias. Hongos, plantas y animales terrestres. La Laguna: Consejería de Politica Territorial y Medio Ambiente del Gobierno de Canarias, 122–172.
- Anthelme, F. , and Dangles, O. , 2012: Plant-plant interactions in tropical alpine environments. Perspectives in Plant Ecology, Evolution and Systematics , 14: 363–372.
- Arévalo, J. R. , Delgado, J. D. , Otto, R. , Naranjo, A. , Salas, M. , and Fernández-Palacios, J. M. , 2005: Distribution of alien vs. native plant species in roadside communities along an altitudinal gradient in Tenerife and Gran Canaria (Canary Islands). Perspectives in Plant Ecology, Evolution and Systematics , 7: 185–202.
- Arévalo, J. R. , Fernández-Palacios, J. M. , Jiménez, M. J. , and Gil, P. , 2001: The effect of fire intensity on the understorey species composition of two Pinus canariensis reforested stands in Tenerife (Canary Islands). Forest Ecology and Management 148: 21–29.
- Atkinson, I. A. E. , 2001: Introduced mammals and models for restoration. Biological Conservation , 99: 81–96.
- Bates, B. , Maechler, M. , and Bolker, B. , 2014: lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-42, http://CRAN.R-project.org/package=lme4 .
- Bowen, L. , and Van Vuren, D. , 1997: Insular endemic plants lack defenses against herbivores. Conservation Biology , 11: 1249–1254.
- Bryant, J. P. , Provenza, F. D. , Pastor, J. , Reichardt, P. B. , Clausen, T. P. , and du Toit, J. T. , 1991: Interactions between woody plants and browsing mammals mediated by secondary metabolites. Annual Reviews in Ecology and Systematics , 22: 431–446.
- Cabrera-Rodriguez, F. , 2006: Microhabitat selection of the European rabbit on La Palma, Canary Islands, Spain. Acta Theriologica , 51: 435–442.
- Calabuig, M. T. , Cortés, M. , Francisco, C. G. , Hernández, R. , and Suárez, E. , 1981: Labdane diterpenes from Cistus symphytifolius. Phytochemistry , 20: 2255–2258.
- Campbell, K. , and Donlan, C. J. , 2005: Feral goat eradications on islands. Conservation Biology , 19: 1362–1374.
- Carracedo, H. C. , Pérez Torrado, F. J. , Ancochea, E. , Meco, J. , Hernán, F. , Cubas, C. R. , Casillas, R. , Rodriguez, E. , and Ahijado, A. , 2002: Cenozoic volcanism II: the Canary Islands. In Gibbons, W. , and Moreno, T. (eds.), The Geology of Spain. London: The Geological Society, 439–472.
- Catford, J. A. , Daehler, C. C. , Murphy, H. T. , Sheppard, A. W. , Hardesty, B. D. , Westcott, D. A. , Rejmánek, M. , Bellingham, P. J. , Pergl, J. , Horvitz, C. C. , and Hulme, P. E. , 2012: The intermediate disturbance hypothesis and plant invasions: implications for species richness and management. Perspectives in Plant Ecology, Evolution, and Systematics , 14: 231–241.
- Caujapé-Castells, J. , Tye, A. , Crawford, D. J. , Santos-Guerra, A. , Sakai, A. , Beaver, K. , Lobin, W. , Florens, F. B. V. , Moura, M. , Jardim, R. , Gómes, I. , and Kueffer, C. , 2010: Conservation of oceanic island floras: present and future global challenges. Perspectives in Plant Ecology, Evolution, and Systematics , 12: 107–130.
- Chiarucci, A. , Araujo, M. B. , Decocq, G. , Beierkuhnlein, C. , and Fernandez-Palacios, J. M. , 2010: The concept of potential natural vegetation: an epitaph? Journal of Vegetation Science , 21: 1172–1178.
- Climent, J. , Tapias, R. , Pardos, J. A. , and Gil, L. , 2004: Fire adaptations in the Canary Islands pine (Pinus canariensis). Plant Ecology , 171: 185–196.
- Courchamp, F. , Chapouis, J. L. , and Pascual, M. , 2003: Mammal invaders on islands: impacts, control and control impact. Biological Reviews , 78: 347–383.
- Crausbay, S. , Genderjahn, S. , Hotchkiss, S. , Sachse, D. , Kahmen, A. , and Arndt, S. K. , 2014: Vegetation dynamics at the upper reaches of a tropical montane forest are driven by disturbance over the past 7300 years. Arctic, Antarctic, and Alpine Research , 46: 787–799.
- Del Arco Aguilar, M. J. , González-González, R. , Garzón-Marchado, V. , and Pizarro-Hernández, B. , 2010: Actual and potential natural vegetation on the Canary Islands and its conservation status. Biodiversity and Conservation , 19: 3089–3140.
- Didiano, T. J. , Turley, N. E. , Everwand, G. , Schaefer, H. , Crawley, M. J. , and Johnson, M. T. J. , 2014: Experimental test of plant defence evolution in four species using long-term rabbit exclosure. Journal of Ecology , 102: 584–594.
- Dob, T. , Berramdane, T. , Dahmane, T. , and Chelghoum, C. , 2005: Chemical composition of the needles oil of Pinus canariensis from Algeria. Chemistry of Natural Compounds 41: 165–167.
- Economou, D. , and Nahrstedt, A. , 1991: Chemical, physiological, and toxicological aspects of the essential oil of some species of the genus Bystropogon. Planta Medica , 57: 347–351.
- Fernández-Palacios, J. M. , Otto, R. , Thebaud, C. , and Price, J. , 2014: Overview of habitat history in subtropical oceanic island summit ecosystems. Arctic, Antarctic and Alpine Research , 46: 801–809.
- Garzón-Machado, V. , del Arco-Aguilar, M. J. , and Pérez-de-Paz, P. L. , 2011: A tool set for description and mapping vegetation on protected natural areas: an example from the Canary Islands. Biodiversity and Conservation , 20: 3605–3625.
- Garzón-Machado, V. , del Arco Aguilar, M. J. , Valdés González, F. , and Pérez-de-Paz, P. L. , 2012: Fire as a threatening factor for endemic plants of the Canary Islands. Biodiversity and Conservation , 21: 2621–2632.
- Garzón-Machado, V. , González-Mancebo, J. M. , Palomares-Martínez, A. , Acevedo-Rodríguez, A. , Fernández-Palacios, J. M. , Del-Arco-Aguilar, M. , and Pérez-de-Paz, P. L. , 2010: Strong negative effect of alien herbivores on endemic legumes of the Canary pine forest. Biological Conservation , 143: 2685–2694.
- Garzón-Machado, V. , Otto, R. , and del Arco Aguilar, M. J. , 2013: Bioclimatic and vegetation mapping of a topographically complex oceanic island applying different interpolation techniques. International Journal of Biometeorology (early view). http://dx.doi.org/10.1007/s00484-013-0670-y.
- Greenwood, S. , and Jump, A. S. , 2014: Consequences of treeline shifts for the diversity and function of high altitude ecosystems. Arctic, Antarctic and Alpine Research , 46: 829–840.
- Heubes, J. , Retzer, V. , Schmitdlein, S. , and Beierkuhnlein, C. , 2011: Historical land use explains current distribution of calcareous grassland species. Folia Geobotanica 46: 1–16.
- Hobbs, R. J. , Arico, S. , Aronson, J. , Baron, J. S. , Bridgewater, P. , Cramer, V. A. , Epstein, P. R. , Ewel, J. J. , Klink, C. A. , Lugo, A. E. , Norton, D. , Ojima, D. , Richardson, D. M. , Sanderson, E. W. , Valladares, F. , Vilà, M. , Zamora, R. , and Zobel, M. , 2006: Novel ecosystems: theoretical and management aspects of the new ecological world order. Global Ecology and Biogeography , 15: 1–7.
- Höllermann, P. , 2000: The impact of fire in Canarian ecosystems 1983-1998. Erdkunde , 54: 70–75
- Irl, S. D. H. , and Beierkuhnlein, C. , 2011: Distribution of endemic plant species on an oceanic island-a geospatial analysis of La Palma (Canary Islands). Procedia Environmental Sciences , 7: 170–175.
- Irl, S. D. H. , Steinbauer, M. J. , Babel, W. , Beierkuhnlein, C. , Blume-Werry, G. , Messinger, J. , Palomares Martínez, Á. , Strohmeier, S. , and Jentsch, A. , 2012: An 11-yr exclosure experiment in a high-elevation island ecosystem: introduced herbivore impact on shrub species richness, seedling recruitment and population dynamics. Journal of Vegetation Science , 23: 1114–1125.
- Irl, S. D. H. , Steinbauer, M. J. , Epperlein, L. , Harter, D. E. V. , Jentsch, A. , Pätz, S. , Wohlfahrt, C. , and Beierkuhnlein, C. , 2014: The hitchhiker's guide to island endemism-biodiversity and endemic perennial plant species in roadside and surrounding vegetation. Biodiversity and Conservation , 23: 2273–2287, http://dx.doi.org/10.1007/s10531-014-0722-6.
- Johansson, M. , Rooke, T. , Fetene, M. , and Granström, A. , 2010: Browser selectivity alters post-fire competition between Erica arborea and E. trimera in the sub-alpine heathlands of Ethiopia. Plant Ecology , 207: 149–160.
- Keeley, J. E. , and Fotheringham, C. J. , 2000: Role of fire in regeneration from seed. In Fenner, M. , (ed.), Seeds: The Ecology of Regeneration in Plant Communities. 2nd edition. New York: CABI, 311–330.
- Kier, G. , Kreft, H. , Lee, T. M. , Jetz, W. , Ibisch, P. L. , Nowicki, C. , Mutke, J. , and Barthlott, W. , 2009: A global assessment of endemism and species richness across island and mainland regions. Proceedings of the National Academy of Science , 106: 9322–9327.
- Kreft, H. , Jetz, W. , Mutke, J. , Kier, G. , and Barthlott, W. , 2008: Global diversity of island floras from a macroecological perspective. Ecology Letters , 11: 116–127.
- Kyncl, T. , Suda, J. , Wild, J. , Wildova, R. , and Herben, T. , 2006: Population dynamics and clonal growth of Spartocytisus supranubius (Fabaceae), a dominant shrub in the alpine zone of Tenerife, Canary Islands. Plant Ecology , 186: 97–108.
- Lowe, S. , Browne, M. , Boudjelas, S. , De Poorter, M. , 2000: 100 of the world's worst invasive alien species-a selection from the Global Invasive Species Database. Aliens , 12: 1–12.
- Merlin, M. D. , and Juvik, J. O. , 1992: Relationships among native and alien plants on Pacific islands with and without significant human disturbance and feral ungulates. In Alien Plant Invasions in Native Ecosystems of Hawai‘i: Management and Research. Manoa: Cooperative National Park Resources Studies Unit, University of Hawai‘i, 597–624.
- Naeem, S. , and Li, S. , 1997: Biodiversity enhances ecosystem reliability. Nature , 390: 507–509.
- Nogales, M. , Rodriguez-Luengo, J. L. , and Marrero, P. , 2006: Ecological effects and distribution of invasive non-native mammals on the Canary Islands. Mammal Review , 36: 49–65.
- Nogales, M. , Rumeu, B. , de Nascimento, L. , and Fernández-Palacios, J. M. , 2014: Newly discovered seed dispersal system of Juniperus cedrus questions the pristine nature of the high elevation scrub of El Teide (Tenerife, Canary Islands). Arctic, Antarctic, and Alpine Research , 46: 853–858.
- Palomares Martínez, Á. , 1993: El matorral de alta montaña en la isla de La Palma, seguimiento debido al peligro de extinción de la especie Genista benehovensis. Ponencias y comunicaciones. Congreso forestal español. Lourizán 2: 421–425.
- Palomares Martínez, Á. , Méndez Carvajal, C. , López Soriano, I. , and Delgado Luzardo, A. , 2004: Plan de Conservación del Hábitat de las Cumbres del Norte de La Palma. Internal document of the National Park Caldera de Taburiente, El Paso, La Palma, Canary Islands.
- Palomares Martínez, Á. , López Graciano, C. , Freixes Montes, F. , Gómez Gómez, M. , Moral del Barrio, M. F. , León Pérez, A. M. , Lorenzo Pérez, J. H. , Balsera Pinar, G. , Méndez Carvajal, C. , Acevedo Rodríguez, A. , Ibáñez de Aldecoa, F. B. , Rodríguez Lerin, A. , and Fernandez San Martín, J. M. , 2011: Memoria anual del Parque Nacional de La Caldera de Taburiente de 2009. Ministerio de Medio Ambiente, y Medio Rural, y Marino, Madrid.
- Pauchard, A ,. Kueffer, C. , Dietz, H. J. , Daehler, C. , Alexander, J. , Edwards, P. J. , Arévalo, J. , Cavieres, L. A. , Guisan, A. , Haider, S. , Jakobs, G. , McDougall, K. , Millar, C. , Naylor, B. , Parks, C. , Rew, L. , and Seipel, T. , 2009: Ain't no mountain high enough: plant invasions reaching new elevations. Frontiers in Ecology and in the Environment , 7: 479–486.
- Pellissier, L. , Roger, A. , Bilat, J. , and Rasmann, S. , 2014: High elevation Plantago lanceolata plants are less resistant to herbivory than their low elevation conspecifics: is it just temperature? Ecography 37: http://dx.doi.org/10.1111/ecog.00833.
- Perry, G. L. W. , Wilmshurst, J. M. , McGlone, M. S. , McWethy, D. B. , and Whitlock, C. , 2012: Explaining fire-driven landscape transformation during the Initial Burning Period of New Zealand's prehistory. Global Change Biology , 18: 1609–1621.
- R Development Core Team , 2014: R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. http://www.R-project.org.
- Rafferty, C. , and Lamont, B. B. , 2007: Selective herbivory by mammals on 19 species planted at two densities. Acta Oecologia , 32: 1–13.
- Ramirez, A. R. , Pratt, R. B. , Jacobsen, A. L. , and Davis, S. D. , 2012: Exotic deer diminish post-fire resilience of native shrub communities on Santa Catalina Island, southern California. Plant Ecology , 213: 1037–1047.
- Shea, K. , and Cheeson, P. , 2002: Community ecology theory as a framework for biological invasions. Trends in Ecology & Evolution , 17: 170–176.
- Steinbauer, M. J. , Irl, S. D. H. , and Beierkuhnlein, C. , 2013: Elevation-driven ecological isolation promotes diversification on Mediterranean islands. Acta Oecologica , 47: 52–56.
- Steinbauer, M. J. , Otto, R. , Naranjo-Cigala, A. , Beierkuhnlein, C. , and Fernández-Palacios, J. M. , 2012: Increase of island endemism with altitude—speciation processes on oceanic islands. Ecography , 34: 1–10.
- Systat Software, Inc. , 2001: SigmaPlot 7.0. http://www.sigmaplot.com. San Jose, California.
- Traveset, A. , Nogales, M. , Alcover, J. A. , Delgado, J. D. , López-Darias, M. , Godoy, D. , Igual, J. M. , and Bover, P. , 2009: A review on the effects of alien rodents in the Balearic (Western Mediterranean Sea) and Canary Islands (Eastern Atlantic Ocean). Biological Invasions , 11: 1653–1670.
- White, P. S. , and Jentsch, A. , 2001: The search for generality in studies of disturbance and ecosystem dynamics. Progress in Botany , 63: 399–449.
- Zöller, L. , von Suchodoletz, H. , and Küster, N. , 2003: Geoarchaeological and chronometrical evidence of early human occupation on Lanzarote (Canary Islands). Quaternary Science Reviews , 22: 1299–1307.
APPENDIX
TABLE A1 List of sown seed quantity for each of the eight target species.