Abstract
The diatom succession and sedimentation during 2011 in a subtropical, deep alpine lake (Lugu Hu, Southwest China) were studied. The seasonal patterns in diatom assemblages were related to light, nutrient availability, and thermal regime. Peaks in the concentration of the planktonic diatoms (Asterionella formosa and Cyclotella ocellata) occurred in the metalimnion during autumn when light penetration was greater and silica concentrations were higher. The spring peak of Cyclostephanos dubius appeared to be a result of the combination of a well-mixed water column, higher nutrient concentration, and low light availability. Its sedimentation in late spring suggests that the sinking rates of C. dubius increased rapidly with the onset of stratification. However, the minimal catches of A. formosa and C. ocellata in sediment traps during autumn suggest they were still actively growing with very low sedimentation losses. The catch of benthic Fragilaria species in sediment traps during early spring and autumn reflected their resuspension from shallow water and transfer into pelagic zone as thermal stratification breaks down. The results provide important information for the interpretation of paleolimnological records from the lakes in Southwest China.
Introduction
The ecology of planktonic diatoms is largely based on studies undertaken on temperate and boreal mono- and dimictic lakes (CitationReynolds, 1984, Citation1990) with an emphasis on the interaction of the physical environment and nutrient loading/availability and, to a lesser extent, the humic content (i.e., dissolved organic carbon [DOC]) (CitationJones, 1998). More recently, as the result of interest in the response of alpine lakes to global environmental change forcing (CitationCatalan et al., 2013), the number of studies of alpine lake phytoplankton has increased (CitationCatalan et al., 2002; CitationSaros et al., 2003, Citation2005), which has broadened our understanding of the dominant controls.
Studies of diatom abundance and sedimentation patterns in temperate lakes demonstrate that their growth and seasonal succession are influenced by the interaction of mixing intensity, light, temperature, and nutrient concentration (CitationMaberly et al., 1994). Reynolds (Citation1990) proposed that the succession of phytoplankton depended on the annual cycle of thermal stratification and its influence on an algal cell's access to light and nutrients. It was suggested that the succession of dominant species was related to the competitive advantages associated with differential sinking rates of non-motile species (diatoms) and behavioral adaptations of motile species, which provide different opportunities for them to exploit available nutrients. For example, the small-sized, fast-growing centric Cyclotella spp., characterized by low sinking rates, may be selectively favored in clear, oligotrophic waters that enable efficient nutrient uptake (CitationWinder et al., 2009), whereas for the large-sized, attenuated pennate species, access to light rather than nutrients is more likely to constrain their growth rate in eutrophic waters (CitationReynolds, 1990).
In contrast to temperate, eutrophic lakes, in alpine lakes the duration of ice cover as well as the onset of water column stratification has an important role in determining seasonal diatom succession (CitationLotter and Bigler, 2000; CitationCatalan et al., 2002). Hausmann and Pienitz (Citation2007) demonstrated the strong impact of spring and summer thermal conditions on diatom composition and productivity, which suggests that with regard to climate effects, the variable timing of seasonal processes is more important than annual mean temperatures. Moreover, thermal stratification and nutrient cycling in tropical and subtropical lakes are different from those at higher latitudes (CitationLewis, 1991). While excessive nutrient loading has altered diatom seasonality in lowland, temperate lakes, studies of the controls on the growth of planktonic diatoms in remote, oligotrophic alpine lakes in the western United States, have suggested that nitrogen enrichment is driving the widespread change in diatom community structure (CitationSaros et al., 2005).
Diatoms preserved in lake sediments provide a long-term record of ecological change in lakes and their catchments (CitationBattarbee, 1991). Transfer functions have been widely used as a basis for the interpretation of sediment diatom sequences, but they essentially reduce ecological complexity to a single variable (for temperate, meso-eutrophic lakes, normally total phosphorus) (e.g., CitationAnderson and Rippey, 1994). It is now clear, however, that transfer functions have fundamental problems (CitationJuggins, 2013), so there is an important need for using contemporary ecological data/knowledge to interpret sediment core profiles (CitationBattarbee et al., 2005). As many diatom species are considered to be globally distributed, their ecology is assumed to be transferable from region to region (i.e., temperate to tropical), and while this might be valid at a coarse ecological scale, the assumptions are rarely addressed in more specific terms, for example, of nutrient-climate relationships in contrasting environments. Although studies of diatoms in temperate alpine lakes have expanded of late, there are still few studies of high altitude [sub]tropical lakes.
A high-resolution fossil diatom sequence from Lugu Hu sediment core shows that (CitationWang et al., 2014; Wang et al., unpublished) the lakes have been dominated by four planktonic diatom species (Cyclostephanos dubius, Cyclotella rhomboideo-elliptica, Asterionella formosa, and Cyclotella ocellata) for more than 20,000 years, since the climate amelioration at the end of Last Glacial Maximum (LGM) (CitationWang et al., 2014). Asterionella was present at low abundance during 18,000–9000 cal. yr B.P. while C. ocellata was a codominant taxon from 14,000 cal. yr B.P. Since 2007, Asterionella has started to increase as indicated by diatom analysis of a 210Pb-dated sediment core (Wang, unpublished). It is suggested that increases of planktonic taxa (C. dubius, A. formosa, and C. ocellata) in sediment records from Lugu Hu reflect changes in the mixing depth, light, and nutrient regeneration from the hypolimnion, which is controlled by the strength and length of thermal stratification. The present study was initiated in part as an attempt to firmly establish this link and to understand the long-term dynamics of the main diatom species found in the sediment core (CitationWang et al., 2014), which are still present in the lake today.
Using limnological (thermal stratification, nutrients, monthly diatom concentration) and sediment trap studies of the subtropical, alpine Lugu Hu, a large, deep lake in Southwest China (), we first provide data on the diatom ecology of cosmopolitan species in a subtropical, alpine lake to test the legitimacy of extrapolating diatom ecology from temperate regions to these systems and second improve the understanding of seasonal succession of diatom species to assist with the interpretation of sedimentary records from this lake.
Study Area
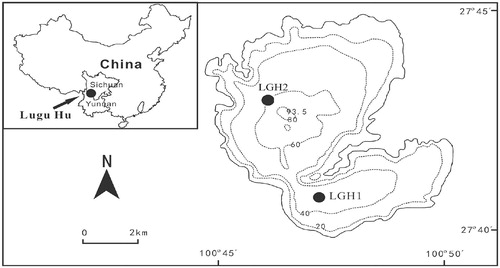
Lugu Hu is a seasonally closed, deep lake located on the boundary between Yunnan Province and Sichuan Province, Southwest China (). It is one of the deepest plateau freshwater lakes in the region (maximum water depth is ∼94 m and mean depth ∼40 m) with a surface area of ∼50.5 km2 (CitationJi and Fan, 1983). It lies at 2685 m above sea level with a catchment area of ∼171.4 km2. The lake sits within an intermontane basin with the highest altitude of 3870 m a.s.l. It has two main inflow rivers. The lake is hydrologically isolated from September to May and in general is connected to the outflow, the Gaizu River, only in the summer (wet) season (CitationJi and Fan, 1983). The estimated water retention time for the whole lake is ∼18 years but the southern basin flushes more quickly.
FIGURE 1. Location map of Lake Lugu showing sampling position of sediment trap and surface water columns that were collected. Depth contours are in metres (Wang and Dou, 1998).
The local climate is subtropical monsoon with distinct dry and wet seasons: the mean annual air temperature is 12.8 °C and mean annual precipitation is ∼940 mm. Precipitation during the wet season (May to September) accounts for more than 80% of the annual total. The terrestrial vegetation in the lake catchment area is dominated by deciduous broadleaved forest, mixed Cyclobalanopsis/Acer stands, and Pinus yunnanensis. Today, the lake is oligo-mesotrophic with secchi depth of 15 and 11 m in summer and autumn, respectively, and total phosphorus (TP) concentration of 28 μg L-1 in the open-water area but is undergoing cultural eutrophication due to tourist development and improved agricultural techniques in the catchment (CitationWu et al., 2008; CitationZhang et al., 2013).
Methods
SAMPLE COLLECTION AND LIMNOLOGICAL ANALYSES
A simple sediment trap made of two parallel transparent 60-cm-long polyvinyl chloride (PVC) tubes (total active area: 56 cm2) was deployed from mid-December 2010. The sediment traps were moored at a depth of 40 m in the southern basin of the lake (LGH1, ). The upper edges of the trap were located approximately 1 m above the lake sediment. Lugol's solution was added to the tubes at the beginning of each sampling interval to prevent grazing of algae by zooplankton (CitationHausmann and Pienitz, 2009). Sediment traps were collected monthly from January to December 2011, but the trap sample from December 2011 was lost due to vandalism by local fishermen.
During the period from January 2010 to December 2011, water samples (∼1 m water depth) were collected at monthly intervals in the southern basin (LGH1, ). These monthly samples were taken by the staff of the local research station. Water samples (at 10 m intervals) were collected at three-month intervals (January, April, July, and October 2011) from the north basin of Lugu Hu where water depth is >50 m (LGH2, ). These quarterly water samples from different depths are considered as representing winter, spring, summer, and autumn seasons. Water temperature, dissolved oxygen (DO), pH, and conductivity in the southern basin (LGH1, ) were measured in the field using a YSI 600XL data sonde (YSI Inc 1700/1725 Brannum Lane, Yellow Springs, Ohio 45387, U.S.A.). Water temperatures at the trap site were recorded at 2.0 h intervals in 2011 by Hobo data loggers circa 0.5 m below the lake surface, every 0.5 m above 20 m, 2 m intervals below 20 m, and at ∼40 m below the lake surface. Additional chemical variables were determined in the laboratory: total nitrogen (TN), total phosphorus (TP), chlorophyll a (Chl-a), concentration of HCO3- and CO3 2-, silica (Si), and DOC were analyzed according to national standards outlined in Monitoring and Analysis Method of Water and Waste Water (CitationState Environmental Protection Administration, 2002). All chemical analyses were completed at Nanjing Institute of Geography and Limnology.
DIATOM PREPARATION AND ANALYSIS
Diatom cells collected from the water column were concentrated and preserved in Lugol's iodine. Samples from sediment traps and water column for diatom analyses were processed with 10% HCl to remove carbonates, followed by cold 30% hydrogen peroxide to oxidize organic matter (CitationBattarbee, 1986). A known quantity of divinyl benzene (DVB) microspheres was added to the samples in order to estimate the total diatom concentrations (cells mL-1) in the water and sediment trap samples (CitationBattarbee and Kneen, 1982). Aliquots of the diatom-microsphere suspensions were transferred onto ethanol-cleaned cover glasses, and after drying at room temperature, mounted with Naphrax mounting medium onto microscope slides. A minimum of 500 diatom valves per slide were counted in each sample at a magnification of 1000× using differential interference contrast (DIC) optics on an Olympus BX51 microscope. Diatoms were identified to the species level or lower, primarily following standard European and North American references (CitationKrammer and Lange-Bertalot, 1986, Citation1988, Citation1991a, Citation1991b). Species relative abundances were expressed as percentage of the total count. For the sediment trap samples, the mean daily flux rate per sampling period was calculated from the total diatom numbers in the sediment trap divided by the number of days of exposure, in order to take into account the different time lags between sampling dates (CitationReynolds et al., 1982).
NUMERICAL ANALYSIS
Multivariate techniques were used to explore the relationships within and between the various data sets (water column and sediment trap samples) and to identify factors that could explain variation in the diatom data. The diatom data (abundances >1%) were used in the analysis. A detrended correspondence analysis (DCA) with detrending by segments and down-weighting of rare taxa helped to estimate the amount of diatom compositional change within the data (CitationTer Braak, 1995). The gradient length of the main DCA ordination axes indicated whether linear- or unimodal-based numerical techniques were appropriate for the analyses (CitationTer Braak and Prentice, 1988). DCA results showed that the gradient length for the first axis was 1.25 standard deviation (S.D.) units, indicating that linear analysis was appropriate (CitationBirks, 1995). Direct gradient ordination by redundancy analysis (RDA) with automatic forward selection was used to test the significance (P < 0.05) of those variables explaining variance in the diatom data. Monte Carlo permutation tests of selected variables and axes (n = 499 permutations, P < 0.05) were employed to test the significance of each variable. The partial-RDAs were also performed to evaluate to what extent the forward-selected variables explained the variation in the diatom assemblage independently. All such ordination techniques were carried out using the program CANOCO version 4.5 (CitationTer Braak and Šmilauer, 2002).
Results
LIMNOLOGY OF LUGU HU
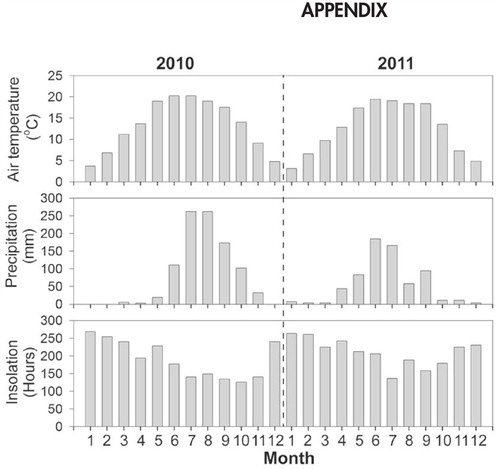
Precipitation, air temperature, and insolation duration data were obtained from the meteorological station at Ninglang, located 44 km to the south of the lake. During the study period, the mean precipitation at Ninglang meteorological station was 973 mm yr-1, with the lowest monthly values occurring between January and May in both 2010 and 2011 (see Appendix, Fig. A1). The 2010 winter period (January–March) was particularly dry, while summer (June–August) precipitation was above average (∼80 mm), with high lake outflow. Air temperatures were highest in June and July in both study years (2010, 2011). Insolation duration decreased during the summer in both years, presumably due to the dominance of cloudy weather at this time.
The water temperature profiles showed that Lugu Hu behaves as a warm monomictic lake (). In 2011, the lake started to stratify in April and was strongly stratified during summer and early autumn (May to September 2011) with a thermocline at ∼10 m. The Secchi depth increased from spring to autumn (), while from June to August (2011) surface water temperatures varied between 18 and 22 °C. With the decrease in air temperatures from August, the depth of thermocline gradually deepened from 10 m to ∼20 m, although the surface water temperature in the autumn was still around 18 °C. DO concentration was high during the circulation periods in winter and autumn, when water column stability was low (). Oxygen levels were low (∼4 mg L-1) in the bottom waters during summer. In the euphotic zone, DO concentration was the highest just below the thermocline in the summer and autumn of 2011 ().
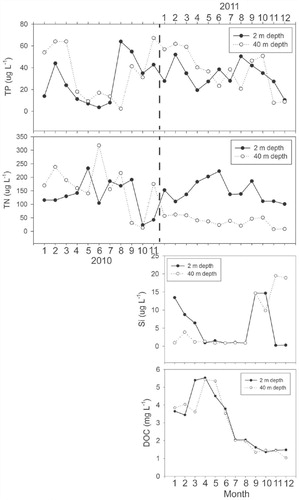
Epilimnetic TP concentration ranged from 4 to 64 µg L-1 with an average of 28 µg L-1 and TN from 23 to 233 µg L-1 with an average of 132 µg L-1 (), reflecting the ongoing eutrophication of the lake associated with shoreline development. TP concentration decreased from February to June in 2010 and February to April in 2011 as the lake thermally stratified. There was no trend in TN concentration from April to July in 2010 and actually an increasing trend in 2011. The Si concentration in the surface water declined from 13.4 to 0.97 mg L-1 from January to April 2011 but increased as the thermocline deepened in the late summer before declining to trace values again in November (). The Si concentration at 40 m was low apart from September to November 2011. DOC concentration peaked in early spring (March–April) at the end of the dry season at ∼5 mg L-1 but then declined steadily throughout the year reaching <1.5 mg L-1 in December (2011).
FIGURE 2. Temperature isopleths in °C for Lugu Lake from January to December 2011.
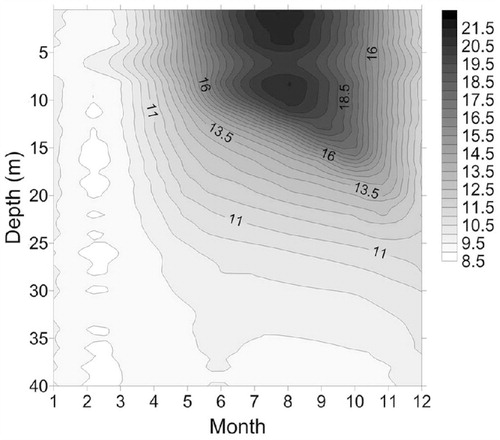
FIGURE 3. The seasonal variation of water temperatures and dissolved oxygen (DO) of stratified water column in Lugu Lake during late 2010 to autumn 2011. The dashed line shows the Secchi disk depth.
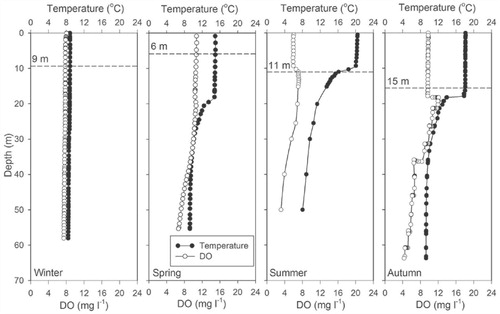
FIGURE 4. Seasonal variation of limnological parameters at Lugu Lake from January 2010 to December 2011. Total phosphorus (TP), total nitrogen (TN), Silica (Si) concentration, and dissolved organic carbon (DOC) from water column at 2 m and 40 m depth. Note that data are missing for December 2011 due to sample loss.
SEASONAL AND VERTICAL DISTRIBUTION OF DIATOMS IN WATER COLUMN
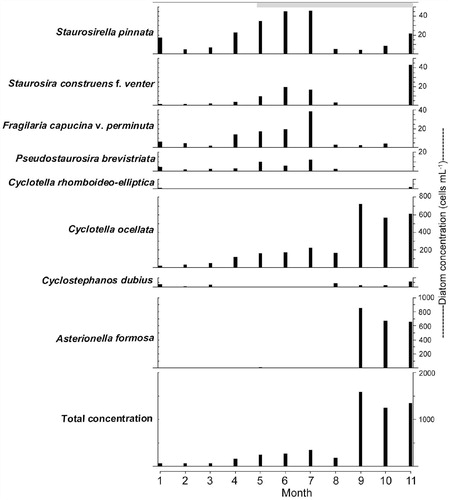
The planktonic diatoms Asterionella formosa Hassall and Cyclotella ocellata Pantocsek were conspicuous components during the autumn period of weakening thermal stratification (i.e., September–November) in both the south and north basins ( and ). In southern basin of Lugu Lake, the highest concentrations of A. formosa and C. ocellata were 8.5 × 102 and 7.2 × 102 cells mL-1, respectively. The total diatom concentration in the water column was the highest in autumn (1.7 × 103 cells mL-1). Maximum concentration of benthic Fragilaria spp. was observed from April to July 2011 (). As in the south basin, A. formosa and C. ocellata were the dominant species in the water column of the north basin during autumn (2011) (). As the thermocline deepened ( and ), A. formosa and C. ocellata were more abundant at depth with a maximum concentration in the metalimnion at 20–30 m (). The high concentration of Cyclostephanos dubius (Fricke) Round was observed in the late winter, that is, January 2011 ( and ). The concentration of C. dubius was highest at depth 40–50 m (hypolimnion) in winter and at 10–20 m (epilimnion) in spring, 2011 ().
FIGURE 5. Seasonal distribution of the most abundant diatom taxa collected monthly (2011) from surface waters of the south basin. The horizontal gray bar at the top signifies the duration of thermal stratification.
Relationships between the diatom and environmental data were explored using RDA; 76.4% and 14.2% of the variance in the assemblages were explained by the first two RDA axes, respectively. In the partial-RDA of diatoms in the water column from 2011, only two forward-selected environmental variables (DOC and N:P ratio) were significant and accounted for 53.2% of the variance in the diatom assemblages. These analyses revealed that light and nutrient availability were statistically associated with changes in the diatom assemblages ().
SEASONAL SEDIMENTATION PATTERN OF DIATOMS IN SEDIMENT TRAPS
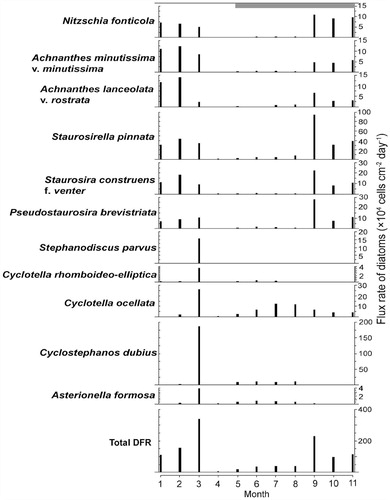
Diatom flux rates (DFR) varied considerably during the study period (). The calculated DFR was between 5.3×104 and 33.8×105 cells cm-2 day-1. The highest catch of planktonic diatoms occurred in March 2011, corresponding to the onset of stratification. During the rest of the year, DFR were lower and the trap catch showed a higher proportion of benthic and tychoplanktonic diatoms. C. dubius dominated in March 2011 reflecting its dominance in the water column immediately prior to this period, particularly in the north basin (). Other planktonic forms, for example, C. ocellata, C. rhomboideo-elliptica Skuja, A. formosa, and Stephanodiscus spp. were also present ().
Discussion
The four planktonic diatom species (C. dubius, C. rhomboideo-elliptica, A. formosa, and C. ocellata) were shown to dominate during the Last Glacial Maximum at Lugu Hu (CitationWang et al., 2014). Some of these species are typical of alpine lakes, for example, C. dubius and small Cyclotella spp. (CitationTapia et al., 2003), while others, for example, Asterionella, have been considered more representative of cultural eutrophication in lowland lakes elsewhere (e.g., CitationAnderson, 1990). C. rhomboideo-elliptica is a regionally endemic species typically observed in the Yunnan Plateau, for example, in Fuxian and Erhai Lakes (CitationQi, 1995; CitationZhang et al., 2005; CitationLi et al., 2012), but it is declining and disappearing at present due to anthropogenic disturbance and nutrient enrichment (CitationLi et al., 2012). Here, we compare the contemporary, seasonal changes at Lugu Hu with those observed in eutrophic lakes in northwest Europe and alpine lakes in North America, with the aim of explaining the changing abundance of planktonic diatoms in the sediment core (CitationWang et al., 2014), which are still extant in the lake today.
FIGURE 6. The diatom concentration from selected depths in the water column of the north basin for four seasons: winter 2010, spring—autumn 2011.
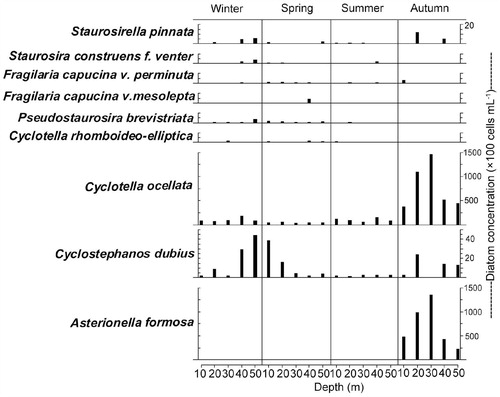
FIGURE 7. Redundancy analysis (RDA) biplot showing the two significant environmental variables selected by forward selection and key diatom taxa. Diatom taxa abbreviations: ASFO, Asterionella formosa; CYDU, Cyclostephanos dubius; CYOC, Cyclotella ocellata; CYRH, Cyclotella rhomboideo-elliptica; FRCAPE, Fragilaria capucina v. perminuta; PSBR, Pseudostaurosira brevistriata; STPI, Staurosirella pinnata; STCOV, Staurosira construens f. venter.
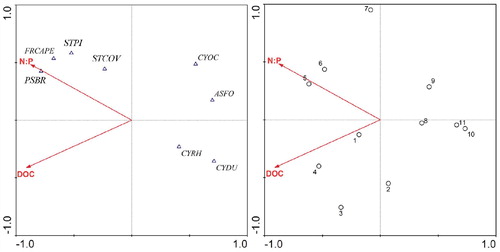
FIGURE 8. Seasonal distribution of the most abundant diatom taxa collected monthly (2011) in the sediment trap.
DIATOM SEASONALITY AND ITS CONTROLS
The growth of diatoms in temperate mono- and dimictic lakes is usually associated with periods prior to stratification in the spring or during deeper mixing phases in the autumn (CitationReynolds, 1990; CitationInterlandi et al., 1999). The main feature of diatom succession at Lugu Hu in 2011 was the dominance of A. formosa and C. ocellata during the autumn and the occurrence of metalimnion peaks at depths of 20–30 m ( and ). The autumn period was characterized by increased light penetration and weakening of thermal stratification during autumnal cooling ( and ). The metalimnetic peaks of Asterionella and C. ocellata were presumably related to their response to changes in the physical and chemical characteristics of the water column. Lund et al. (Citation1963) showed that the reduction in nutrient availability, particularly lake water silicate, was an important factor leading to sudden decline of A. formosa at Windermere. In Lugu Lake, Si was replenished in September (>14 mg L-1) as water temperatures declined (although water temperatures were still ∼17 °C) and the thermocline deepened ( and ). Although Si was depleted in the epilimnion by December it was still abundant at 40 m (). While the dominance of A. formosa and C. ocellata can be broadly attributed to the changing Si concentration and a deeper mixed layer, the improved light climate (Secchi disk depth increased from 6 to 15 m) at the end of summer due to reduced cloud cover and declining DOC concentration ( and Appendix, Fig. A1), will also have contributed. Some support for this conclusion is provided by the RDA (), which indicated that DOC and N:P ratio were the most important factors influencing the diatom assemblages. The increased light penetration during autumn therefore allowed metalimnetic peaks of algae to utilize the nutrients (Si) available at depth (). Maberly et al. (Citation1994), analyzing the 45-year record of Asterionella in Windermere, suggested that light played a critical role in the timing and growth of both spring and autumn populations.
Another factor influencing diatom succession at Lugu Hu is the reduced flushing rate during autumn and winter, which will have reduced cell loss rates via the outflow, certainly in the southern basin (LGH1, ). Phytoplankton population increase is a function of growth minus loss: for diatoms, these are mainly sedimentation and washout; grazing is generally assumed to be of minor importance, particularly for Asterionella (CitationThackeray et al., 2008).
A. formosa is a characteristic member of the phytoplankton in many temperate mesotrophic and eutrophic lakes where it often has a pronounced vernal population when lakes are isothermal with a secondary, autumnal population (CitationReynolds and Irish, 2000), which contrasts with its seasonal succession in the Lugu Hu where it dominated during autumn (September–November 2011). Reynolds (Citation1990) noted that stratification causes the loss of A. formosa from the water column because it depends on well-mixed water for suspension. However, its sinking rate changes dramatically when growth ceases, increasing its susceptibility to sinking losses. It has been suggested that enhanced sinking rate of Asterionella can aid nutrient uptake in clear and oligotrophic waters (CitationReynolds, 1990).
COMPARISON WITH ALPINE LAKES ELSEWHERE
The seasonal succession of A. formosa and C. ocellata in alpine lakes contrasts to that in lowland, temperate lakes in that summer metalimnetic populations seem to be more important (CitationSaros et al., 2005, Citation2013). At Lugu Hu, A. formosa and C. ocellata had their highest concentrations in metalimnion layer during autumn (). The declining DOC concentration during autumn allowed light to penetrate deeper into the metalimnion as nutrients diffused upward from the hypolimnion (CitationWebb et al., 1997), contributing to the metalimnetic peak of planktonic diatoms during the autumn period (). Anneville et al. (Citation2004) have showed that autumn community contains R—strategists, such as A. formosa, which are tolerant to low light intensities and therefore adapted to autumnal mixing conditions. However, in oligotrophic alpine lakes in the western United States, it has been shown that the growth of A. formosa has been stimulated by N additions due to enhanced atmospheric N deposition (CitationSaros et al., 2003). The distribution of A. formosa was positively correlated with the N:P ratio, suggesting that its abundance peaks when P availability is very low and the supply of N and Si are moderate to high (CitationSaros et al., 2005).
Cyclotella species are common members of phytoplankton in Arctic and alpine lakes (CitationLotter and Bigler, 2000). The changing abundance of Cyclotella species was considered to indicate changes in stratification patterns or shifts in light:nutrient ratios (CitationInterlandi et al., 1999). This genus with high surface area to volume ratio has high light requirements and low sinking rates, allowing it to remain in the epilimnion during strong summer stratification (CitationBradbury, 1988). Saros et al. (Citation2012) conducted comparative lake surveys and growth rate experiments, which revealed that Cyclotella species are influenced by lake thermal structure when nitrogen is sufficient. Increases in the relative abundances of small Cyclotella species are often interpreted as signals for warming-induced changes in the length of ice-free season or depth of mixed layer (CitationWinder et al., 2009). Depth of the mixed surface layer and hence the stability of thermal stratification influences Cyclotella abundance; C. ocellata, for example, is associated with shallower summer mixing depths (CitationWhitlock et al., 2012). Nutrient availability, however, is an important aspect of the response of Cyclotella species to changes in lake mixing depths (CitationSaros et al., 2012).
The studies discussed above are similar to the observations at Lugu Hu where Asterionella and Cyclotella spp. dominated during the autumn period as a metalimnetic peak as a result of the increased light and nutrient availability during the autumnal mixing period. This is instructive for the interpretation of fossil sequences from Lugu Hu. For example, the rapid increases of Cyclotella and A. formosa around 14,500 cal. yr B.P. were presumably indicative of increased nutrient availability, associated with the changes in the lake's mixing regime (CitationWang et al., 2014). A better understanding of the seasonal succession of planktonic diatoms at Lugu Hu today therefore improves the paleoenvironmental and paleoclimatic interpretation of lacustrine diatom records from this area.
TEMPORAL VARIABILITY IN DIATOM SEDIMENTATION AT LUGU HU
Although the diatom plankton was dominated by Asterionella and C. ocellata in the autumn of 2011 (), C. dubius was more important during the late winter of 2010 and early spring 2011 (). This is also reflected in a peak in the catch of C. dubius in the sediment traps in March 2011 (). Water temperature started to increase during this period and the lake started to stratify (). During the spring period (January–March 2011), light penetration was comparatively low (Secchi disk = 6 m) as DOC was close to its maximum (>5 mg L-1) and Si was declining (6.4 mg L-1) ().
Although C. dubius was dominant in the late autumn and winter at Lugu Hu, it was observed during late-summer and/or autumn period in shallow, nutrient-rich Danish lakes where C. dubius competed well for nutrient resources under low light conditions (CitationBradshaw and Anderson, 2003). Bradbury et al. (Citation1994) commented on its success during periods of water column mixing, high nutrient concentrations, and decreasing light availability in lakes. The presence of C. dubius during the winter of 2010–2011 in Lugu Hu, confirms that this species may be well adapted to the conditions of a well-mixed water column, abundant nutrients, but relatively low light availability. The result strongly supports the interpretation of C. dubius dominance in the Lugu Hu sediment core as indicative of high nutrient conditions related to the altered thermal structure, reflecting rising temperatures and development of thermal stratification patterns similar to today (CitationWang et al., 2014). The differences in concentration of C. dubius between the north and south basins suggest that it grows deeper in the metalimnion layer (), and its apparent low concentration in the south basin reflects only sampling surface waters () (see Methods).
The catch of planktonic diatoms in the sediment traps was dominated by C. dubius in March 2011 (), suggesting that sinking and loss rates of C. dubius increased rapidly when the lake started to stratify. Although A. formosa and C. ocellata were dominant during autumn 2011, there was minimal catches of these species in the sediment traps at this time ( and ), suggesting that they were still actively growing with very low sedimentation losses. The ability of A. formosa to remain in suspension in water column depends in part on mixing and turbulence (CitationReynolds, 1990). Its sinking rate has been observed to increase dramatically after growth ceases (CitationReynolds, 1998). A. formosa has a higher surface area to volume ratio, suggesting that it has a higher form resistance to sinking and would be lost less quickly from a mixed-water column (CitationThackeray et al., 2008, and references therein). This suggests that A. formosa has lower sinking losses than C. dubius and can still maintain a positive net growth rate. There is considerable interannual variability in phytoplankton, as shown by long-term studies (e.g., CitationMaberly et al., 1994), and our attempts to explain the contrasting abundances of C. dubius and A. formosa in Lugu Hu would benefit from an extended study.
BREAKDOWN OF STRATIFICATION AND RESUSPENSION OF FRAGILARIA
Small Fragilaria and other benthic species (e.g., Nitzschia fonticola [Grunow] Grunow in Van Heurck) entered the sediment trap mainly during early spring and autumn of 2011 (). At this time the lake was either isothermal or was mixed to depths >17 m, and the increased catch rates of Fragilaria cells primarily reflect their resuspension from shallow water and transfer into the pelagic. The spatial distribution of diatom assemblages from surface sediments taken at different water depths in Lugu Hu showed that high concentrations of Fragilaria were mainly observed in water depths <10 m (CitationWang et al., 2012). The resuspension of diatoms and sediment from the littoral zone during the breakdown of thermal stratification contributes to the sediment accumulation of benthic taxa in deep water sediments. Our results are also supported by the sediment trap study by Köster and Pienitz (Citation2006) who found that Fragilaria species were associated with stronger water column circulation and resuspension of sediments. The sediment trap catches of Fragilaria peaked during spring and autumn periods as stratification weakens (), which can be used to track the paleoenvironmental changes during the past 30,000 cal. yr B.P. The increased abundance of small Fragilaria species during the LGM may reflect weaker stratification and longer isothermal conditions during the climatic cooling and possible development of seasonal ice cover at this time.
THE RESPONSE OF A. FORMOSA TO ENVIRONMENTAL CHANGES
The recent increase of A. formosa at Lugu Hu, as at Windermere (CitationPennington, 1943) is its response to the eutrophication associated with the development of the lake as a tourist resort. Nutrient loading to the lake has increased dramatically as uncontrolled hotel and restaurant building has proliferated over the last 10 years (CitationMattison, 2010). Asterionella is often associated with the early phases of cultural eutrophication (CitationAnderson et al., 1995) but has also been observed in some oligotrophic lakes (CitationSaros et al., 2005). It is recognized that the ecology of Asterionella at an oligotrophic, alpine lake (Lugu Hu) seems to be a conflation of the main processes that have been identified elsewhere—nutrients (Si, N:P ratio), light, and the thermal regime—but its seasonality is very different, with peak numbers in autumn and winter. This seasonality represents the interplay between the physical characteristics of Lugu Hu and the regional weather conditions that influence seasonal light and nutrient availability. The results of the present study broaden our understanding of Asterionella and small Cyclotella and contribute to a better paleoclimatological interpretation of diatom records from subtropical deep alpine lakes. The present study highlights the need for more regional studies of diatom succession and the need for some circumspection when extrapolating ecological inferences from temperate lakes to sites in the subtropics. The dominant controlling factors are, however, not that dissimilar to those observed by Lund et al. (Citation1963).
Acknowledgments
This work was supported by the Research Funding Plan for Post-doctoral fellowships in Jiangsu Province (1301006C), the National Basic Research Program of China (2010CB950201), and the Jiangsu Collaborative Innovation Center for Climate Change, China. Anderson acknowledges the support of the Royal Society (London) and the Chinese Academy of Sciences (visiting professorship scheme).
References Cited
- Anderson, N. J. , 1990: The biostratigraphy and taxonomy of small Stephanodiscus and Cyclostephanos species (Bacillariophyceae) in a eutrophic lake, and their ecological implications. British Phycological Journal , 25: 217–235.
- Anderson, N. J. , and Rippey, B. , 1994: Monitoring lake recovery from point-source eutrophication: the use of diatom-inferred epilimnetic total phosphorus and sediment chemistry. Freshwater Biology , 32: 625–639.
- Anderson, N. J. , Renberg, I. , and Segerstrom, U. , 1995: Diatom production responses to the development of early agriculture in a boreal forest lake-catchment (Kassjon, Northern Sweden). Journal of Ecology , 83: 809–822.
- Anneville, O. , Souissi, S. , Gammeter, S. , and Straile, D. , 2004: Seasonal and inter-annual scales of variability in phytoplankton assemblages: comparison of phytoplankton dynamics in three peri-alpine lakes over a period of 28 years. Freshwater Biology , 49: 98–115.
- Battarbee, R. W. , 1986: Diatom analysis. In Berglund, B. G. (eds.), Handbook of Holocene Paleoecology and Paleohydrology. London: Wiley and Sons, 527–570.
- Battarbee, R. W. , 1991: Recent paleolimnology and diatom-based environmental reconstruction. In Shane, L. C. K. , and Cushing, E. J. (eds.), Quaternary Landscapes. Minneapolis: University of Minnesota Press, 129–174.
- Battarbee, R. E. , Anderson, N. J. , Jeppesen, E. , and Leavitt, P. R. , 2005: Combining palaeolimnological and limnological approaches in assessing lake ecosystem response to nutrient reduction. Freshwater Biology , 50: 1772–1780.
- Battarbee, R. W. , and Kneen, M. J. , 1982: The use of electronically counted microspheres in absolute diatom analysis. Limnology and Oceanography , 27: 184–188.
- Birks, H. J. B. , 1995: Quantitative palaeoenvironmental reconstructions. In Maddy, D. , and Brew, J. S. (eds.), Statistical Modelling of Quaternary Science Data. Cambridge: Quaternary Research Association, 161–254.
- Bradbury, J. P. , 1988: Fossil diatoms and Neogene paleolimnology. Palaeogeography, Palaeoclimate, Palaeoecology , 62: 299–316.
- Bradbury, J. P. , Bezrukova, Ye. V. , Chernyaeva, G. P. , Colman, S. M. , Khursevich, G. , King, J. W. , and Likoshway, Ye. V. , 1994: A synthesis of post-glacial diatom records from Lake Baikal. Journal of Paleolimnology , 10: 213–252.
- Bradshaw, E. G. , and Anderson, N. J. , 2003: Environmental factors that control the abundance of Cyclostephanos dubius (Bacillariophyceae) in Danish lakes, from seasonal to century scale. European Journal of Phycology , 38: 265–276.
- Catalan, J. , Ventura, M. , Brancelj, A. , Granados, I. , Thies, H. , Nickus, U. , Korhola, A. , Lotter, A. F. , Barbieri, A. , Stuchlík, E. , Lien, L. , Bitušík, P. , Buchaca, T. , Camarero, L. , Goudsmit, G. H. , Kopácek, J. , Lemcke, G. , Livingstone, D. M. , Müller, B. , Rautio, M. , Šiško, M. , Sorvari, S. , Šporka, F. , Strunecký, O. , and Toro, M. , 2002: Seasonal ecosystem variability in remote mountain lakes: implications for detecting climatic signals in sediment. Journal of Paleolimnology , 28: 25–46.
- Catalan, J. , Pla-Rabés, S. , Wolfe, A. P. , Smol, J. P. , Rühland, K. M. , Anderson, N. J. , Kopáček, J. , Stuchlík, E. , Schmidt, R. , Koinig, K. A. , Camarero, L. , Flower, R. J. , Heiri, O. , Kamenik, C. , Korhola, A. , Leavitt, P. R. , Psenner, R. , and Renberg, I. , 2013: Global change revealed by palaeolimnological records from remote lakes: a review. Journal of Paleolimnology , 49: 513–535.
- Hausmann, S. , and Pienitz, R. , 2007: Seasonal climate inferences from high-resolution modern diatom data along a climate gradient: a case study. Journal of Paleolimnology , 38: 73–96.
- Hausmann, S. , and Pienitz, R. , 2009: Seasonal water chemistry and diatom changes in six boreal lakes of the Laurentian Mountains (Québec, Canada): impacts of climate and timber harvesting. Hydrobiologia , 635: 1–14.
- Interlandi, S. J. , Kilham, S. S. , and Theriot, E. C. , 1999: Responses of phytoplankton to varied resource availability in large lakes of the Greater Yellowstone Ecosystem. Limnology and Oceanography , 44: 668–682.
- Ji, J. , and Fan, Y. Q. , 1983: Preliminary analysis on the hydrologic characteristics of Lake Lugu. In The Comprehensive Scientific Expedition to the Qinghai-Xizang Plateau, Chinese Academy of Sciences. Qinghai-Xizang Plateau Research, Hengduan Mountains Expedition, Series One. Kunming: Yunnan People's Publishing House, 214–225 (in Chinese).
- Jones, R. I. , 1998: Phytoplankton, primary production and nutrient cycling. In Hessen, D. O. , and Tranvik, L. J. (eds.), Aquatic Humic Substances: Ecology and Biogeochemistry. Berlin: Springer, 145–195.
- Juggins, S. , 2013: Quantitative reconstructions in palaeolimnology: new paradigm or sick science? Quaternary Science Reviews , 64: 20–32.
- Köster, D. , and Pienitz, R. , 2006: Seasonal diatom variability and paleolimnological inferences—a case study. Journal of Paleolimnology , 35: 395–416.
- Krammer, K. , and Lange-Bertalot, H. , 1986: Bacillariophyceae, Teil 1, Naviculaceae. Band 2/1: Süßwasserflora von Mitte-leuropa (begründet von A. Pascher). Stuttgart: Gustav Fischer, 876 pp.
- Krammer, K. , and Lange-Bertalot, H. , 1988: Bacillariophyceae, Teil 2, Epithemiaceae, Bacillariaceae, Surirellaceae. Band 2/2: Süßwasserflora von Mitteleuropa (begründet von A. Pascher). Stuttgart: Gustav Fischer, 611 pp.
- Krammer, K. , and Lange-Bertalot, H. , 1991a: Bacillariophyceae. 3: Teil: Centrales, Fragilariaceae, Eunotiaceae. Unter Mitarbeit von H. Håkannson und M. Nörpel. In Ettl, H. , Gärtner, G. , Gerloff, J., Heynig, H. , and Mollenhauer, D. (eds.), Süßwasserfloravon Mitteleuropa, 2/3. Stuttgart: Gustav Fischer Verlag, 576 pp.
- Krammer, K. , and Lange-Bertalot, H. , 1991b: Bacillariophyceae. 4: Teil: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis. In Ettl, H. , Gärtner, G. , Gerloff, J. , Heynig, H. , and Mollenhauer, D. (eds.), Süßwasserflora von Mitteleuropa, 2/4. Stuttgart: Gustav Fischer Verlag, 437 pp.
- Lewis, W. M., Jr. , 1991: Comparisons of phytoplankton biomass in temperate and tropical lakes. Limnology and Oceanography , 35: 1838–1845.
- Li, Y. L. , Gong, Z. J. , and Shen, J. , 2012: Effects of eutrophication and temperature on Cyclotella rhomboideo-elliptica Skuja, endemic diatom to China. Phycological Research , 60(4): 288–296.
- Lotter, A. F. , and Bigler, C. , 2000: Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquatic Science , 62: 125–141.
- Lund, J. W. G. , Mackereth, F. J. H. , and Mortimer, C. H. , 1963: Changes in depth and time of certain chemical and physical conditions and of the standing crop of Asterionella formosa Hass. in the north basin of Windermere in 1947. Philosophical Transactions of the Royal Society of London B , 246: 255–290.
- Maberly, S. C. , Hurley, M. A. , Corry, J. E. , Heaney, S. I. , Irish, A. E. , Jaworski, G. H. M. , Lund, J. W. G. , Reynolds, C. S. , and Roscoe, J. V. , 1994: The rise and fall of Asterionella formosa in the South Basin of Windermere: analysis of a 45-year series of data. Freshwater Biology , 31: 19–34.
- Mattison, S. M. , 2010: Economic impacts of tourism and erosion of the visiting system among the Mosuo of Lugu Lake. The Asia Pacific Journal of Anthropology: Ethnicity and Economies in Yunnan , 11(2): 159–176.
- Pennington, W. , 1943: Lake sediments: the bottom deposits of the north basin of Windermere, with special reference to the diatom succession. New Phytologist , 42(1): 1–27.
- Qi, Y. Z. , 1995: Flora Algarum Sinicarum Aquae Dulcis, Tomus IV, Bacillariophyta, Centricae. Beijing: Science Press, 104 pp (in Chinese).
- Reynolds, C. S. , 1984: The Ecology of Freshwater Phytoplankton. Cambridge: Cambridge University Press.
- Reynolds, C. S. , 1990: Temporal scales of variability in pelagic environments and response of phytoplankton. Freshwater Biology , 23: 25–53.
- Reynolds, C. S. , 1998: What factors influence the species composition of phytoplankton in lakes of different trophic status? Hydrobiologia , 369/370: 11–26.
- Reynolds, C. S. , and Irish, A. E. , 2000. The phytoplankton of Windermere (English Lake District). Ambleside, U.K.: Freshwater Biological Association.
- Reynolds, C. S. , Morison, H. R. , and Butterwick, C. , 1982: The sedimentary flux of phytoplankton in the south basin of Windermere. Limnology and Oceanography , 27: 1162–1175.
- Saros, J. E. , Interlandi, S. J. , Wolfe, A. P. , and Engstrom, D. R. , 2003: Recent changes in the diatom community structure of lakes in the Beartooth Mountain Range (USA). Arctic, Antartic, and Alpine Research , 35: 18–23.
- Saros, J. E. , Michel, T. J. , Interlandi, S. J. , and Wolfe, A. P. , 2005: Resource requirements of Asterionella formosa and Fragilaria crotonensis in oligotrophic alpine lakes: implications for recent phytoplankton community reorganizations. Canadian Journal of Fisheries and Aquatic Sciences , 62: 1681–1689.
- Saros, J. E. , Stone, J. R. , Pederson, G. T. , Slemmons, K. E. H. , Spanbauer, T. , Schliep, A. , Cahl, D. , Williamson, C. E. , and Engstrom, D. R. , 2012: Climate-induced changes in lake ecosystem structure inferred from coupled neo- and paleoecological approaches. Ecology , 93(10): 2155–2164.
- Saros, J. E. , Strock, K. E. , Mccue, J. , Hogan, E. , and Anderson, N. J. , 2013: Response of Cyclotella species to nutrients and incubation depth in Arctic lakes. Journal of Plankton Research , 36(2): 450–460.
- State Environmental Protection Administration , 2002: Monitoring and Analysis Method of Water and Waste Water. Fourth edition. Beijing: China Environment Science Press, 778 pp.
- Tapia, P. M. , Fritz, S. C. , Baker, P. A. , Seltzer, G. O. , and Dunbar, R. B. , 2003: A Late Quaternary diatom record of tropical climatic history from Lake Titicaca (Peru and Bolivia). Palaeogeography, Palaeoclimatology, Palaeoecology , 194: 139–164.
- Ter Braak, C. J. F. , 1995: Ordination. In Jongman, R. H. G. , ter Braak, C. J. F. , and van Tongeren, O. F. R. (eds.), Data Analysis in Community and Landscape Ecology. Cambridge: Cambridge University Press, 91–169.
- Ter Braak, C. J. F. , and Prentice, I. C. , 1988: A theory of gradient analysis. Advances in Ecological Research , 18: 271–317.
- Ter Braak, C. J. F. , and Šmilauer, P. , 2002: CANOCO Reference Manual and CanoDraw for Windows User's Guide: Software for Canonical Community Ordination (Version 4.5). Ithaca, New York: Microcomputer Power, 500 pp.
- Thackeray, S. J. , Jones, I. D. , and Maberly, S. C. , 2008: Long-term change in the phenology of spring phytoplankton: species-specific response to nutrient enrichment and climatic change. Journal of Ecology , 96(3): 523–535.
- Wang, Q. , Yang, X. D. , Hamilton, P. B. , and Zhang, E. L. , 2012: Linking spatial distributions of sediment diatom assemblages with hydrological depth profiles in a plateau deep-water lake system of subtropical China. Fottea , 12: 59–73.
- Wang, Q. , Yang, X. D. , Anderson, N. J. , Zhang, E. L. , and Li, Y. L. , 2014: Diatom response to climate forcing of a deep, alpine lake (Lugu Hu, Yunnan, SW China) during the Last Glacial Maximum and its implications for understanding regional monsoon variability. Quaternary Science Reviews , 86: 1–12.
- Webb, D. J. , Robarts, R. D. , and Prepas, E. E. , 1997: Influence of extended water column mixing during the first 2 years of hypolimnetic oxygenation on the phytoplankton community of Amisk Lake, Alberta. Canadian Journal of Fisheries and Aquatic Sciences , 54: 2133–2145.
- Whitlock, C. , Dean, W. E. , Fritz, S. C. , Stevens, L. R. , Stone, J. R. , Power, M. J. , Rosenbaum, J. R. , Pierce, K. L. , and Bracht-Flyr, B. B. , 2012: Holocene seasonal variability inferred from multiple proxy records from Crevice Lake, Yellowstone National Park, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 331–332: 90–103.
- Winder, M. , Reuter, J. E. , and Schladow, G. , 2009: Lake warming favours small-sized planktonic diatom species. Proceedings of the Royal Society B , 276: 427–435.
- Wu, G. , Zhang, Q. , Zheng, X. , Mu, L. , and Dai, L. , 2008: Water quality of Lugu Lake: changes, causes and measurements. International Journal of Sustainable Development & World Ecology , 15: 10–17.
- Zhang, E. L. , Cao, Y. M. , Langdon, P. , Wang, Q. , Shen, J. , and Yang, X. D. , 2013: Within-lake variability of subfossil chironomid assemblage in a large, deep subtropical lake (Lugu Lake, Southwest China). Journal of Limnology , 72(1): 117–126.
- Zhang, X. , Xie, P. , Chen, F. Z. , Li, Y. L. , Li, S. X. , Guo, N. C. , and Qin, J. H. , 2005: Present status and changes of the phytoplankton community after invasion of Neosalanx taihuensis since 1982 in a deep oligotrophic plateau lake, Lake Fuxian in the subtropical China. Journal of Environmental Sciences , 17(3): 389–394.
FIGURE A1. Meteorological data recorded at the meteorological station of Ninglang.