
Abstract
Recent studies have found that the biological features of primary tumors are faithfully recapitulated when a patient's tumor is processed and then maintained as a 3-D spheroid in specialized cell culture media. However, a major drawback for maintenance and routine passage of primary tumors as spheroids has been the high cost of custom-formulated media compared to regular serum-supplemented media. Here we report the formulation of a cost-effective, serum-free medium in which high-grade primary brain tumor (glioblastoma) explants can be established and maintained as spheroids. Based on DMEM, this formulation requires only supplementation with several amino acids, vitamins, synthetic EGF, and bFGF, with most of the cost being associated with the growth factors. A simple addition of BSA (fraction V) obviated the need for numerous other components (or human serum) commonly used in the specialized commercial media formulations.
This method describes a DMEM based formulation to establish and propagate spheroid (neurosphere) cultures from human glioblastoma resections. Lipid-fortified BSA (fraction V) was used as a substitute for several supplements present in commercial neurosphere culture media, including donor human serum; the resulting formulation is 1/6 the cost of commercially available neurosphere media.
Maintenance of primary human tumors as in vitro monolayer (2-D) cultures in the presence of bovine, equine, or human serum supplemented media has been an established laboratory practice for decades. However, it has become apparent that these culture conditions irreversibly modify both genotype and phenotype, with passaged tumor cell lines bearing little resemblance to the tumor isolated from the patient (Citation1). While cells derived from malignant brain tumors (glioblastoma) maintained in the presence of serum are tumorigenic in vivo, they tend to form well-circumscribed tumors in xenograft models that do not recapitulate the invasive (diffused) growth patterns observed in a clinical setting. In contrast, when tumors are established and maintained as 3-D spheroids using specialized serum-free media or media with minimum amounts of human serum, the biological features of the primary tumor is faithfully maintained, facilitating preclinical studies more relevant to the patient's tumor (Citation1, Citation2). A major drawback for such maintenance and routine passage of tumors as spheroids has been the high cost associated with this specialized media in comparison to “regular” media which are most often supplemented with fetal bovine serum (FBS), in the average tissue culture laboratory. Here, we report the formulation of a cost-effective, serum-free medium for routine use, in which primary malignant glioblastoma tissues can be established and maintained as spheroids (neurospheres).
Most commercial formulations for spheroid culture are proprietary and expensive (e.g., Neurobasal medium, Life Technologies Inc., Grand Island, NY; NeuroCult medium, StemCell Technologies Inc., Vancouver, Canada; CellGro COMPLETE medium, Mediatech Inc., Manassas, VA). In addition, some formulations also contain human serum as a component necessary for initiation and propagation of spheroids from human neuron or glial tissue explants derived from human patients. However, media formulations developed during pioneering studies several decades ago utilized either DMEM alone, or in combination with F12 media supplemented with various growth factor/steroid/hormone combinations to culture neurons and glial cells in vitro (Citation3–5).
We modified and supplemented the standard DME medium with components from the G-5 supplementation (Citation5), but using off-the-shelf components and insulin-transferrin-selenium premixes from low-cost commercial sources (see Supplementary Protocol). Our lab-made formulation did not include any human or bovine serum and, in addition to the standard components of high-glucose DME medium, included three additional amino acids (Ala, Asn, and Pro), Zn2+, biotin, vitamin B12, hydrocortisone, the insulin-transferrin-selenium mixture, and the growth factors EGF and bFGF, which are needed for propagation and maintenance of glial cells and tumor cells of astrocytic lineage as neurospheres or spheroids (). Since human plasma contains albumin at 3–4 g/dL (Citation6), our medium was supplemented with BSA preparations that maintain the fatty acid and growth factor traits of the original “fraction V” (cold-ethanol) preparations (Sigma Chemical Co., St. Louis, MO, or Life Technologies) at 4% (w/v) final concentration. Preliminary studies indicated that albumin was essential to the establishment and propagation of spheroids, serving as a carrier for cations, growth factors, and hormones, in addition to functioning as an osmoregulator. All other components listed in the proprietary B27-type (Citation4) supplement that are used in commercial media formulations were not necessary for establishment and propagation of spheroid cultures from glioblastoma explants.
Table 1. Comparison of supplements present in HMSF medium and the Neurobasal medium plus B27 supplement (4) that is routinely present in commercial spheroid or neurosphere culture media.
In brief, a tumor tissue section (0.25–0.5 g wet weight, the size of a pencil-tip-eraser) was resuspended immediately after surgical resection in 10 mL ice-cold sterile PBS (or saline) in a 60 mm or 100 mm diameter tissue culture dish, and then sliced into approximately 1–2 mm3 fragments by using a cross-cut pattern with a sterile scalpel. The tissue fragments, along with the PBS or saline, were aspirated into a 15 mL conical tube and allowed to settle for 1 min. The spent PBS (or saline) containing blood and serum components was aspirated off, and the tissue fragments were washed while in the tube in the same manner as above, with 10 mL aliquots of cold PBS until the wash was visibly clear of blood (approximately 2–3 additional washes are needed depending on the vascularity of the tumor). Then, tissue fragments were aspirated and transferred onto a fresh 100 mm tissue culture dish, and all residual PBS was aspirated off. Five mL of AccuMax (Innovative Cell Technologies, San Diego, CA), a cell-dissociation enzyme formulation of crustacean origin that is more mild than trypsin, was added to the plate, and the plate placed on a rocker platform for 30 min to 1 h (but no longer) at room temperature until at least half of the tissue fragments had sloughed off as cell clumps consisting of 2–10 cells when visualized under a low-power (40×) inverted microscope. Next, the mixture was placed back in a 15 mL conical tube and residual undigested tissue fragments were allowed to settle for 1 min. The supernatant was aspirated into a new 15 mL conical tube and the cell clumps then allowed to “gravity-settle” for 10 min. The supernatant from this step was discarded; although some tumor cells are lost at this step, almost 90% of the erythrocytes that contaminate primary tumor cell preparations can be eliminated from the tissue digest. Ten mL of our “home-made serum free” (hereafter referred to as HMSF medium) medium formulated as described above (see Supplementary Protocol) was added to the tube. The suspension was then aspirated into a 100 mm tissue culture dish and placed in a 37°C CO2 tissue culture incubator (5% CO2-air). After 24 h, the culture was aspirated into a 15 mL conical tube and centrifuged in a swing-bucket rotor at 100× g, in 1 min steps until most of the glial cells were pelleted while residual erythrocytes remained in the supernatant (the stepwise centrifugations are carried out until the “whitish” cell pellet just starts to become covered with a “reddish layer” of erythrocytes). Again, supernatant was discarded, and the cell pellet resuspended in a fresh 10 mL aliquot of HMSF medium and placed in a new 100 mm tissue culture dish. This process was repeated every 72 h, until tumor spheroids were clearly visible under low-power microscopy (1–2 weeks based on tumor grade, with highly aggressive tumors giving rise to spheroids within a week). Media changes were continued once every three days during long-term propagation of the spheroids.
Once the spheroids reached a diameter of roughly 100 µm (approximately 100 cells/spheroid), they were spun down at 100× g for 5 min, washed once in 10 mL PBS, and then resuspended in either 2.5 mL of Accutase (another mild trypsin substitute of crustacean origin; Innovative Cell technologies) or 2.5 mL of Accumax. This suspension was incubated at room temperature for up to 10 min with occasional gentle mixing until most of the spheroids had dispersed into a fine suspension of cell clumps consisting of 2 to 10 cells. Ten mL HMSF medium was added to the mixture and centrifuged as before. The pellet was resuspended in 10 mL fresh HMSF medium, roughly enumerated using a hemocytometer or with a Coulter counter, and the equivalent of 1 × 106 cells (in 10 mL) plated per 100 mm diameter tissue culture plate for passaging the cells as spheroids (see Supplementary Protocol for alternate non-enzymatic fragmentation of spheroids for routine passage).
To date, we have established and maintained spheroid cultures from eight primary glioblastoma explants using our HMSF media (with one failure, where the tumor explant grew as attached cells in both HMSF and commercial neurosphere media). It is important to note that even established glioma cell lines such as U87MG and U251MG generate spheroids in HMSF within 1–2 weeks ().
(A) U87-MG glioma in regular DMEM supplemented with 10% FBS; (B) U87-MG glioma after passaging in HMSF for two weeks, to induce spheroid formation; (C) human glioma spheroids established from a grade IV glioma (glioblastoma multiforme) after maintenance and passage in HMSF for six months; (D) spheroids established from a primary tumor tissue (grade III astrocytoma) after four weeks of passage; and (E) after two months of passage.

To compare the characteristics of glioma cells established and maintained as either spheroids or as monolayer cultures, we utilized 3 primary glioblastoma cell isolates: (i) GBM1, initially established as spheroids in commercial (NeuroCult) media and then maintained in HMSF media (as spheroids) or in DMEM supplemented with 10% FBS (monolayer culture) for 12 months, (ii) GBM2, a recent tumor explant, directly established as a monolayer culture in DMEM supplemented with 10% FBS and maintained in the same medium for 2 months, and (iii) GBM3, another recent tumor explant, but established and maintained as spheroids in HMSF media for 2 months.
We then placed each isolate in HMSF or 10% FBS supplemented DMEM for 2 additional months, to maintain (GBM1 and 3) or establish (GBM2) spheroids, or to establish (GBM 3) or maintain (GBM 1 and 2) monolayer cultures, respectively. Invasive capacity of each cell culture was examined using Matrigel-based invasion assays (). In separate experiments, each isolate was also placed in HMSF or commercial medium (NeuroCult), again for two additional months and allowed to maintain (GBM1 and 3) or form spheroids (GBM2). Proliferative capacity was measured for each isolate (). We also assessed CD133 expression for each GBM isolate using flow cytometry (). CD133, a known marker for tumor hypoxia, is the most commonly used marker to identify the “stemness” of glioblastoma explants (Citation7).
(A) Invasive capacity of primary glioma spheroids versus monolayer cultures. Assays were carried out in Boyden chambers essentially as described before (Citation9). Matrigel was diluted 1:3 in Optimem-1 media (Life Technologies) supplemented with 4% FBS. Glioma single cell suspensions were prepared by treating with Accutase and 4 × 104 cells plated per well-insert in 0.5 mL of Optimem-1 media as above. 0.75 mL of DMEM/F12 media supplemented with 10% FBS was placed in bottom wells as the chemo-attractant. Twenty-four hours post-plating, the well-inserts were placed in a new set of wells containing 4 µg/mL Calcein AM (Caymen Chemical, MI) in 0.5 mL PBS. The bottom surface of wells was read at 517 nm (494 nm excitation). P-values (± SD, Student's t-test) were: GBM1, 0.049; GBM2, 0.347; GBM3, 0.068. (B) Proliferative rates of glioma spheroids in NeuroCult versus HMSF media. Three to ten cell spheroids maintained in NeuroCult or HMSF media were plated at approximately 1.2 × 106 cells per well in 60 mm diameter plates in 5 mL of each medium, respectively. The media were changed every three days for two weeks. Single cell suspensions were prepared from each spheroid culture and plated in 6 well plates pre-coated in poly L-lysine (10 µg/mL) and allowed to attach overnight in Optimem-1 media supplemented with 4% FBS. MTT assays were carried out on the cells as described previously (Citation8). P-values (± SD, Student's t-test) were: GBM1, 0.959; GBM2, 0.008; GBM3, <0.001. (C) CD133 expression in glioma spheroids versus monolayer cultures. Single cell suspensions were prepared using Accutase and washed in Optimem-1 media supplemented with 4% FBS. The cells were suspended in PBS supplemented with 0.2% BSA and 2 mM EDTA at 107 cells per 80 µl. Twenty µl of FcR blocking reagent and 10 µl of phycoerythrin coupled CD133 antibody (Miltenyi Biotech, CA) were added to each sample and incubated in the dark for 30 min at room temperature. The cells were then washed (2 mL) and resuspended in the same PBS buffer as above and analyzed in a BD LSRII flow cytometer (BD Biosciences, San Jose, CA). FlowJo ver. 9.3.2 software (TreeStar, Inc., Ashland, OR) was used for analysis.
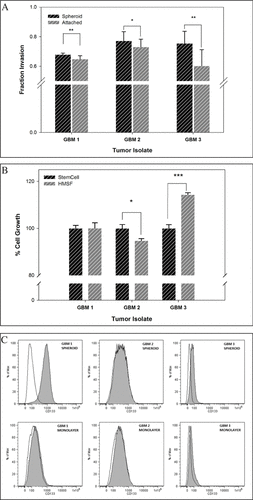
While the invasion assays indicated slightly higher (but statistically significant) penetrance of Matrigel by cells maintained as spheroids, no clear difference in proliferation rates was observed for spheroids maintained in commercial versus HMSF medium. Markedly higher expression of CD133 was observed in GBM1 spheroids over GBM1 cells maintained as monolayers. GBM2, initially established as a monolayer culture, either lost or did not harbor CD133 expression; GBM2 did not gain CD133 expression upon spheroid formation either. However, the GBM3 cell isolates that were established and then maintained as spheroids from inception displayed a small, but statistically significant, population of CD133 positive cells that diminished upon monolayer culture. Thus, our studies indicate that “stemness” is selected for and maintained, upon prolonged culture of primary glioblastoma explants as spheroids in HMSF medium.
BSA fraction V is the only “non-standard” component in our HMSF media formulation that may show batch-to-batch variability. However, similar variation will be present in donor human sera that are provided as separate supplements in most commercial neurosphere culture media. To date, we have tested four batches of BSA, three from Sigma Chemical Co., and one from Life Technologies, finding no variance in spheroid formation capabilities or in growth properties (data not shown).
In summary, our media formulation and protocols represent cost-effective and reproducible methods for the initiation, propagation, and maintenance of primary human glioma cultures as spheroids that maintain the phenotypic characteristics of the original tumor, allowing for more applicable clinical extrapolation.
Competing interests
The authors declare no competing interests.
Formulation of Serum-Free Media for Spheroid / Neurosphere Culture
Download PDF (242.2 KB)Acknowledgments
Research support for the corresponding author was provided by a grant from the National Cancer Institute/National Institute of Health (CA 116257), the Fund for Medical Research and Education (FMRE), Wayne State University, and a gift from the Marvin E. Klein, M.D., Charitable Trust. The authors wish to thank Eric Van Buren and Jessica Back from the Cytometry Resources Core (supported in part by NIH Center grant CA022453) at Wayne State University for the flow analyses. This paper is subject to the NIH Public Access Policy.
Supplementary data
To view the supplementary data that accompany this paper please visit the journal website at: www.tandfonline.com/doi/suppl/10.2144/000114066
Additional information
Funding
References
- Lee, J., S.Kotliarova, Y.Kotliarov, A.Li, Q.Su, N.M.Donin, S.Pastorino, B.W.Purow, et al.. 2006. Tumor stem cells derived from glioblastomas cultured in bFGF and EGF more closely mirror the phenotype and genotype of primary tumors than do serum-cultured cell lines.Cancer Cell9:391–403.
- Fael Al-Mayhani, T.M., S.L.Ball, J.W.Zhao, J.Fawcett, K.Ichimura, P.V.Collins, and C.Watts. 2009. An efficient method for derivation and propagation of glioblastoma cell lines that conserves the molecular profile of their original tumours.J. Neurosci. Methods176:192–199.
- Brewer, G.J. and C.W.Cotman. 1989. Survival and growth of hippocampal neurons in defined medium at low density: advantages of a sandwich culture technique or low oxygen.Brain Res.494:65–74.
- Brewer, G.J., J.R.Torricelli, E.K.Evege, and P.J.Price. 1993. Optimized survival of hippocampal neurons in B27-supplemented Neurobasal, a new serum-free medium combination.J. Neurosci. Res.35:567–576.
- Bottenstein, J.E. and G.Sato. 1985. Cell culture in the neurosciences. Plenum Press, New York.
- Young, D.S. and E.J.Huth. 1998. SI units for clinical measurement. American College of Physicians, Philadelphia, PA.
- Brescia, P., C.Richichi, and G.Pelicci. 2012. Current strategies for identification of glioma stem cells: adequate or unsatisfactory?J. Oncol.2012:376894.
- Saifer, A. and L.Goldman. 1961. The free fatty acids bound to human serum albumin.J. Lipid Res.2:268–270.
- Colen, C.B., Y.Shen, F.Ghoddoussi, P.Yu, T.B.Francis, B.J.Koch, M.D.Monterey, M.P.Galloway, et al.. 2011. Metabolic targeting of lactate efflux by malignant glioma inhibits invasiveness and induces necrosis: an in vivo study.Neoplasia13:620–632.

