
Abstract
Standard methods for mitochondrial DNA (mtDNA) extraction do not provide the level of enrichment for mtDNA sufficient for direct sequencing and must be followed by long-range-PCR amplification, which can bias the sequencing results. Here, we describe a fast, cost-effective, and reliable method for preparation of mtDNA enriched samples from eukaryotic cells ready for direct sequencing. Our protocol utilizes a conventional miniprep kit, paramagnetic bead-based purification, and an optional, limited PCR amplification of mtDNA. The first two steps alone provide more than 2000-fold enrichment for mtDNA when compared with total cellular DNA (∼200-fold in comparison with current commercially available kits) as demonstrated by real-time PCR. The percentage of sequencing reads aligned to mtDNA was about 22% for non-amplified samples and greater than 99% for samples subjected to 10 cycles of long-range-PCR with mtDNA specific primers.
More mitochondrial DNA
Mitochondrial DNA (mtDNA) suffers a higher rate of mutation compared to nuclear DNA, due to factors including proximity to highly reactive oxygen species, lack of histone protection, and poor fidelity of mtDNA replication and repair. The result of this higher mutation rate is heterogeneity within the mtDNA population of a cell, a useful indicator of the cumulative effects of endogenous and environmental DNA damaging agents, as well as general cellular deterioration. Next-generation sequencing (NGS) is widely used to catalogue and analyze mtDNA sequence heterogeneity. However, even with multiple copies of mtDNA in each cell, mtDNA represents only a small fraction of the total DNA, necessitating enrichment schemes to isolate mtDNA prior to NGS. Enrichment techniques such as ultracentrifugation on CsCl gradients require special instrumentation, while PCR amplification of mtDNA from total DNA samples can potentially lead to unwanted amplification artifacts. As a simpler, faster, and more affordable alternative, A.Y. Maslov and his group at the Albert Einstein College of Medicine (New York, NY) describe a new method for preparing samples of highly enriched mtDNA suitable for direct sequencing. The method is composed of two steps: (i) mtDNA is isolated using a standard bacterial plasmid DNA miniprep kit, and (ii) the isolated DNA is purified by reversible binding on solid-phase paramagnetic beads. The use of DNA miniprep kits for mtDNA isolation is based the similarity of mtDNA size to that of plasmid DNA, a fact exploited in other mtDNA isolation techniques. However, when followed by the bead purification steo recommended by Maslov and his colleagues, the level of mtDNA purification, as measured by qPCR, is greatly enhanced, with a ∼2000-fold enrichment relative to total cellular DNA, compared to a ∼10-fold enrichment for 2 commercial mtDNA kits tested. mtDNA purified with this new method performed well in next-generation sequencing, resulting in 22% and 99% of reads aligning to mtDNA for unamplified samples and for samples enriched by 10 PCR cycles, respectively. While this new method is technically simple and rapid, it is the high level of enrichment that will truly help scientists with their studies of mtDNA.
See “Fast mitochondrial DNA isolation from mammalian cells for next-generation sequencing” on page 133.
Keywords::
Here we present a fast and cost-effective two-step protocol for isolation of mtDNA from mammalian cells that is suitable for NGS analysis: (i) extraction of an mtDNA-enriched fraction using a common plasmid miniprep kit and (ii) further purification of mtDNA using the Agencourt AMPure XP system. This method is capable of an up to ∼2000-fold enrichment of mtDNA, is superior to commercial kits, and costs a fraction of the price.
In mammalian cells, mitochondria are often present in thousands of copies, depending on the cell type. Mitochondrial genomes lack histone protection and reside in close proximity to reactive oxygen species. These factors, as well as limited fidelity of mtDNA replication and repair machineries, ensure a much higher mutation rate in the mitochondrial genome than in the nuclear genome (Citation1,Citation2), leading to heterogeneity within the mtDNA population (Citation3,Citation4). However, any deleterious effects of random mutations in mtDNA are compensated by the presence of multiple mitochondria in each cell. This decreased selection pressure allows mutated mtDNA to accumulate over time, making mtDNA a powerful indicator of detrimental effects of endogenous and environmental damaging agents, as well as overall somatic deterioration. It is also known that inherited mutations in the mtDNA can cause human diseases or mitochondrial disorders such as maternally inherited diabetes and deafness (Citation5,Citation6), mitochondrial myopathy (Citation7), and even accelerated aging (Citation8).
Currently, next-generation sequencing (NGS) approaches are widely used for analysis of mtDNA (Citation9,Citation10). However, despite the presence of multiple mitochondrial genomes in each cell, mtDNA only comprises a small portion of total cellular DNA, thus making it necessary to enrich samples for mtDNA before sequencing. Current methods for enrichment either require special equipment (ultra-centrifugation in CsCl density gradients), application of relatively expensive kits, or PCR amplification of mtDNA from total cellular DNA. This last and most commonly used method is relatively cheap and efficient, but may lead to artifacts as high number of PCR amplification cycles are often needed for sufficient enrichment. This can lead to misinterpretation of results and, ultimately, incorrect conclusions.
To overcome these limitations, we have designed a fast, cost-effective, and reliable method for preparation of samples highly enriched for mtDNA (). This method includes two steps—isolation of total cellular DNA enriched for supercoiled mtDNA using a conventional bacterial miniprep kit followed by additional purification using solid phase reversible immobilization on paramagnetic beads.
Total cellular DNA is isolated for control purposes.
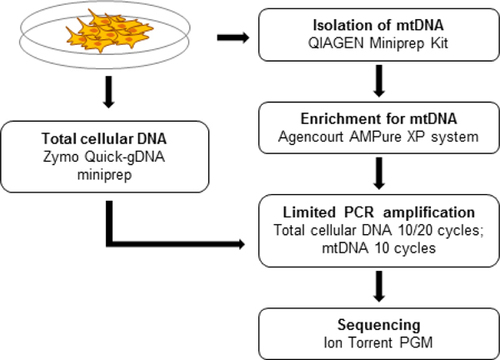
As proof of principle, we applied our method for the isolation of mtDNA-enriched cellular DNA from mouse embryonic fibroblasts (MEFs). MEFs were harvested from E13.5 C57Bl/6 mouse embryos and maintained in culture using standard conditions. Cultured cells were collected into 10 mL of PBS using cell lifters to avoid excessive damage associated with the application of trypsin. Next, MEFs were pelleted by centrifugation at 500 × g for 5 min. Approximately 17 million cells were used for each mtDNA extraction.
We assumed, as have others (Citation11,Citation12), that since mtDNA properties are similar to those of bacterial DNA (i.e., it is supercoiled and its size is in the range of conventional plasmids), a common miniprep kit can be used for extraction and enrichment of mtDNA from eukaryotic cells. In our experiments we applied the QIAprep Spin Miniprep Kit (Qiagen, Germantown, MD) as the first step for mtDNA isolation (). The procedure was performed according to the manufacturer's recommendations, and DNA was eluted into 100 μL of elution buffer. Next, the mtDNA-enriched fraction was purified using Agencourt AMPure XP system (Beckman Coulter, Brea, CA). The beads were added in a 0.4× proportion by volume, collected on the magnetic stand, and washed twice with freshly prepared 70% ethanol. After air-drying the beads, the mtDNA was resuspended in 25 μL of 0.1× TE buffer. In parallel, mtDNA was isolated using two dif ferent commercial kits, the Mitochondrial DNA Isolation Kit (BioVision, Milpitas, CA) and the Mitochondria Isolation Kit (MACS) (Miltenyi Biotec Inc., Auburn, CA). In the latter case, DNA from isolated mitochondria was extracted using Quick-gDNA Blood miniprep kit (Zymo Research, Irvine, CA). Additionally, total cellular DNA was extracted from the cell pellets using this last kit. DNA obtained by these methods was also subjected for an additional purification step using 0.4× AMPure beads as described above.
To compare the levels of mtDNA enrichment obtained with the different procedures we performed real-time PCR using genomic GAPDH as a reference. The primers used were:
GAPDH (amplicon size 84 bp):
5’-GGCTCCCTAGGCCCCTCCTG-3’
and
5′-TCCCAACTCGGCCCCCAACA-3′ mtDNA (amplicon size 119 bp):
5′-CCCAGCTACTACCATCATTCAAGT-3′
and
5′-GATGGTTTGGGAGATTGGTTGATGT-3′
The relative level of enrichment was calculated using the formula RQ = 2(−ΔΔCt). The results indicate that our method provides a considerable level of enrichment for mtDNA, as compared with the two commercial kits tested (). We were able to achieve more than 3 orders of magnitude of enrichment as compared with total genomic DNA, whereas standard kits provided roughly 10× enrichment. The additional bead purification step had an effect only when applied to DNA samples extracted with the QIAGEN kit. Interestingly, the QIAGEN miniprep kit applied without bead purification still achieved much better results than any other specialized kit.
Table 1. Enrichment of mtDNA over nuclear DNA using different isolation methods as measured by real-time PCR.
To test the suitability of the mtDNA enriched with the new method, we performed direct next-generation sequencing, using the Ion Torrent PGM platform (Life Technologies, Grand Island, NY), of (i) total cellular DNA, both un-amplified and enriched for mtDNA by 10 cycles of PCR amplification; (ii) mtDNA isolated by the two commercially available kits; and (iii) mtDNA isolated by our method. PCR amplification of the total mitochondrial genome was performed using Q5 High-Fidelity DNA polymerase (New England Biolabs, Ipswich, MA), and two primer sets.
First set (amplicon size 8.6 Kb):.
5′-AGCAAAAGCCCACTTCGCCA-3′.
and.
5′-GGTTGGCCCCCAATTCAGGT-3′.
.Second set (amplicon size 8 Kb):.
5′-ACCTGAATTGGGGGCCAACC-3′.
and.
5′-TGGCGAAGTGGGCTTTTGCT-3′.
PCR reactions were performed separately for each primer pair. Once the reaction was completed, products were combined and purified using AMPure beads (1.5 ×) as described above.
Bar-coded sequencing libraries were prepared using the NEBNext Fast DNA Fragmentation & Library Prep Set for the Ion Torrent (New England Biolabs) and sequenced on the Ion Torrent PGM using the Ion PGM Sequencing 200 Kit v2 (Life Technologies) and the Ion 318 chip (Life Technologies) (). Results obtained for the sample purified with our new procedure show that after 10 cycles of amplification for mtDNA, 99% of reads aligned to the mitochondrial genome, whereas the un-amplified sample had about 22% of the reads aligning to the mitochondrial genome (). In sharp contrast, we observed an alignment of ∼10% and ∼35% for total cellular DNA and mtDNA extracted with the MACS, respectively, but only after 10 cycles of amplification. Of note, direct sequencing after MACS enrichment remains unfeasible.
Table 2. Sequencing quality.
Table 3. Summary of sequencing results.
Taken together, these results show that our new method is a convenient, fast, and cost-effective means of preparing mtDNA, with or without limited PCR amplification, for next-generation sequencing. To our knowledge, this is currently the only approach allowing direct sequence analysis of mtDNA after enrichment from total cellular DNA.
Competing interests
The authors declare no competing interests.
Acknowledgments
This research was supported by NIH grant P01 AG017242 (JV) and by the Ministry of Education and Science of Russian Federation grant 14.B37.21.1966 (VNP and AYM). This paper is subject to the NIH Public Access Policy.
Additional information
Funding
References
- Brown, W.M., M.George, Jr., and A.C.Wilson. 1979. Rapid evolution of animal mitochondrial DNA.Proc. Natl. Acad. Sci. USA76:1967–1971.
- Baklouti-Gargouri, S., M.Ghorbel, A.Ben Mahmoud, E.Mkaouar-Rebai, M.Cherif, N.Chakroun, A.Sellami, F.Fakhfakh, and L.Ammar-Keskes. 2013. Mitochondrial DNA mutations and polymorphisms in asthenospermic infertile men.Mol Biol Rep.40:4705–4712.
- Greaves, L.C., A.K.Reeve, R.W.Taylor, and D.M.Turnbull. 2012. Mitochondrial DNA and disease.J. Pathol.226:274–286.
- Larsson, N.G. and D.A.Clayton. 1995. Molecular genetic aspects of human mitochondrial disorders.Annu. Rev. Genet.29:151–178.
- Mezghani, N., M.Mnif, E.Mkaouar-Rebai, N.Kallel, N.Charfi, M.Abid, and F.Fakhfakh. 2013. A maternally inherited diabetes and deafness patient with the 12S rRNA m.1555A>G and the ND1 m.3308T>C mutations associated with multiple mitochondrial deletions.Biochem. Biophys. Res. Commun.431:670–674.
- van den Ouweland, J.M., H.H.Lemkes, R.C.Trembath, R.Ross, G.Velho, D.Cohen, P.Froguel, and J.A.Maassen. 1994. Maternally inherited diabetes and deafness is a distinct subtype of diabetes and associates with a single point mutation in the mitochondrial tRNA(Leu(UUR)) gene.Diabetes43:746–751.
- Bindoff, L.A. and B.A.Engelsen. 2012. Mitochondrial diseases and epilepsy.Epilepsia53(Suppl 4):92–97.
- Khrapko, K. and J.Vijg. 2009. Mitochondrial DNA mutations and aging: devils in the details?Trends Genet.25:91–98.
- Ameur, A., J.B.Stewart, C.Freyer, E.Hagstrom, M.Ingman, N.G.Larsson, and U.Gyllensten. 2011. Ultra-deep sequencing of mouse mitochondrial DNA: mutational patterns and their origins.PLoS Genet.7:e1002028.
- Mondal, R. and S.K.Ghosh. 2013. Accumulation of mutations over the complete mitochondrial genome in tobacco-related oral cancer from northeast India.Mitochondrial DNA.24:432–439.
- Defontaine, A., F.M.Lecocq, and J.N.Hallet. 1991. A rapid miniprep method for the preparation of yeast mitochondrial DNA.Nucleic Acids Res.19:185.
- Peloquin, J.J., D.M.Bird, and E.G.Platzer. 1993. Rapid miniprep isolation of mitochondrial DNA from metacestodes, and free-living and parasitic nematodes.J. Parasitol.79:964–967.

