
Quantification of bacterial species from the highly diverse human gastrointestinal microflora (Citation1,Citation2) is a great technical challenge (Citation3). Since several diseases are associated with an altered composition of the gastrointestinal microflora (Citation2,Citation4,Citation5), methods for quantification of specific bacterial species or genera are of great importance to monitor the population dynamics within this microbial ecosystem. Various methods for analyzing the composition of complex microbiota have been developed (reviewed in Reference Citation6). As most intestinal bacterial species are yet uncultured, quantification by real-time PCR seems to be the method of choice. Real-time PCR allows quantification using SYBR® Green or probe-based systems, [e.g., TaqMan® or hybridization/fluorescent resonance emission transfer (FRET) probes] (Citation7). The major advantage of probes is the sequence-specific detection, whereas SYBR Green detection often results in a trade-off between sensitivity and specificity due to detection of unspecific amplification products.
Unfortunately, species- or genus-specific sequences of the 16S rRNA genes required for specific detection of bacteria are very short, thus preventing the use of DNA-based probes. Conventional DNA probes of approximately 20 bp in length are necessary to reach a suitable melting temperature (Tm). Thus, the quantification of species-specific bacterial DNA with probes requires species-specific amplification using two primer pairs for comparing two different bacterial species. Quantitative comparison of two PCR amplifications demands tremendous “fine-tuning” due to different PCR efficiencies resulting from varying binding kinetics of the two different primer pairs. Therefore, our aim was to establish an assay to quantify different bacterial species with probes from one 16S rDNA product, generated by one pair of eubacterial-conserved primers. This product spans different species-specific regions, allowing the quantification by individual specific probes. Such an approach using minor groove binding (MGB) TaqMan probes has recently been published (Citation8). The addition of the minor groove binding peptide to the TaqMan probes resulted in an increased Tm, thereby allowing for the design of very short species-specific probes. Alternatively, the incorporation of locked nucleic acid (LNA) oligonucleotides within the probes also results in an increased Tm due to their rigid conformational structure (Citation9). Several applications for LNA technology have been described within the last 2 years, including single nucleotide polymorphism (SNP) analysis, 5′-nuclease assay, expression profiling, and fluorescence in situ hybridization (FISH) analysis (Citation10–15).
We demonstrate here that LNA-containing oligonucleotides can also be used as hybridization/FRET probes for detection and quantification of short species-specific 16S rDNA sequences by real-time PCR. Based on this novel application, we established a sensitive assay for rapid quantification of bacterial 16S rDNA using hybridization/FRET probes containing LNA nucleotides.
The assay is based on the amplification of a 354-bp fragment of the 16S rRNA target gene from eubacterial DNA. DNA was isolated by a conventional phenol/chloroform extraction from human stool samples representating the complex gastrointestinal microbial community. Eubacterial-conserved primers for amplification used were as follows: sense, 5′-CATGGRTGTCGTCAGCTCGTG-3′ and reverse, 5′-TGACGGGCGGTGTGTACA-3′ (R = A or G, due to variations at this position). The product was detected with eubacterial-conserved hybridization/FRET probes (EuBdonor and EuBacceptor) containing LNA nucleotides: EuBdonor, 5′-ACGTCATCCCC-3′ labeled with fluorescein at the 3′ end and EuBacceptor, 5′-CCTTCCTC-3′ labeled with LC-705 at the 5′ end. The LNA nucleotides are shown in bold. The eubacterial-conserved regions within the 16S rRNA genes for designing primers and probes were chosen according to the analysis of domain-specific 16S primers by Baker and Cowan (sense, pos. 1054–1074; reverse, pos. 1391-1408; EuBdonor, pos. 1184-1194; EuBacceptor, pos. 1174-1182) (Citation16). The insertion of the respective LNA nucleotides resulted in a Tm shift from 33° to 63°C (EuBdonor) and below 26° to 65°C (EuBacceptor) compared to DNA oligonucleotides without LNAs. PCR was performed in a total volume of 20 µL containing 4 µL FastStart DNA MasterPLUS hybridization/FRET (Roche Diagnostics GmbH, Mannheim, Germany), 5 µL DNA, 0.25 µM primers, and 0.2 µM probes. Amplification in the LightCycler® instrument (Roche Diagnostics GmbH) was according to the following parameters: initial denaturation at 95°C for 10 min, 40 cycles of denaturation at 95°C for 0 s, annealing at 60°C for 10 s, and extension at 72°C for 15 s. Fluorescence intensity was determined during each cycle at the end of the annealing step. By performing 10-fold dilution steps of 100 ng target DNA, it could be seen that LNA-containing hybridization/FRET probes are well suited for specific product detection ().
(A) Hybridization/FRET probes. (B) SYBR Green.
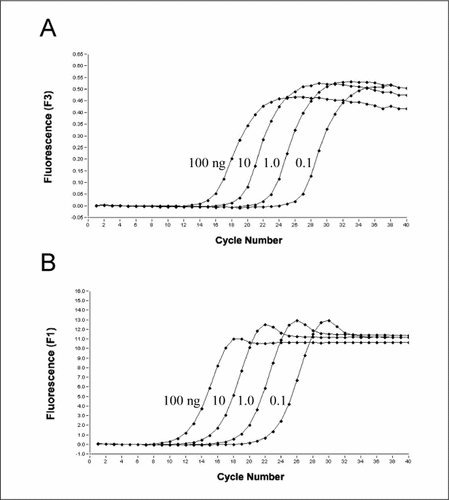
To compare this new detection method with a conventional detection method, we performed a similar PCR run with the same primers and template DNA using SYBR Green for detection (). A comparison of both assays showed similar results regarding fluorescence generation, demonstrating that LNA-containing oligonucleotides can be applied as hybridization/FRET probes for real-time PCR (). However, cycle threshold (Ct) values were found to be approximately 3.2 cycles lower with the SYBR Green assay, indicating a higher sensitivity of SYBR Green compared to hybridization/FRET probes, most likely due to the fact that a greater number of SYBR Green molecules are incorporated by the product.
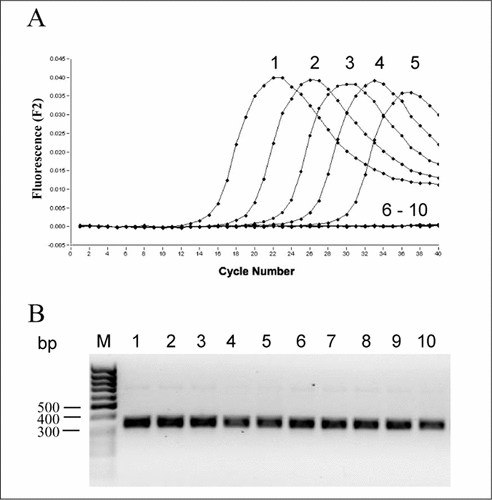
To show species-specific detection, we designed LNA/FRET probes specific for the Bacteroides genus as follows: 5′-LC-640-AGCCGACACCT-3′ and 5′-CGTTATGGCACTT-FL-3′. Amplification with the same conditions as described above, and Bacteroides-specific detection is shown in . Using 10-fold dilutions of Bacteroides thetaiotaomicron DNA yields strong signal generation (, lanes 1–5). In contrast, 100 ng of five control DNAs from Klebsiella planticola, Enterococcus faecalis, Bacillus cereus, Clostridium sporogenes, and Bifidobacterium longum, each containing three mismatches per probe, gave no fluorescence signal even after 40 cycles of amplification, indicating high specificity of the probes (, lanes 6–10). Successful amplification of all DNAs was confirmed by agarose gel electrophoresis ().
(A) Fluorescent signal generation using 100 ng (lane1), 10 ng (lane 2), 1 ng (lane 3), 0.1 ng (lane 4), and 0.01 ng (lane 5) of Bacteroides thetaiotaomicron DNA. To control specificity 100 ng DNA of Klebsiella planticola (lane 6), Enterococcus faecalis (lane 7), Bacillus cereus (lane 8), Clostridium sporogenes (lane 9), and Bifidobacterium longum (lane 10) were used. (B) Successful amplification was shown by agarose gel electrophoresis.
Taken together, the results demonstrate that LNA-based hybridization/FRET probes are suitable alternatives to the recently published MGB probes in quantitative real-time PCR of 16S rDNA and extend the potential of using short detection probes with respect to dye chemistry and PCR instruments. Employing LNA oligonucleotides may permit the use of significant shorter probes, as LNA-containing hybridization/FRET probes of 8–11 bp are working as well as MGB probes of 13–18 bp (Citation8). Furthermore, both assays should be able to discriminate between species that differ in only 1 or 2 bp due to the significant decrease in the Tm by either mismatched LNA-containing probes or MGB probes.
Competing Interests Statement
The authors declare no competing interests.
Acknowledgments
This work was supported by the Bundesministerium für Bildung und Forschung (BMBF, CAPNetz, Project C5) and the Deutsche Forschungsgemeinschaft (DFG, project GO 363/10-1).
Additional information
Funding
References
- Boriello, S.P. 1986. Microbial flora of the gastrointestinal tract, p. 2–16. In M.J.Hill (Ed.), Microbial Metabolism in the Digestive Tract. CRC Press, Boca Raton.
- Kalliomaki, M. and E.Isolauri. 2003. Role of intestinal flora in the development of allergy. Curr. Opin. Allergy Clin. Immunol.3:15–20.
- von Wintzingerode, F., U.B.Gobel, and E.Stackebrandt. 1997. Determination of microbial diversity in environmental samples: pitfalls of PCR-based rRNA analysis. FEMS Microbiol. Rev.21:213–229.
- Guarner, F. and J.R.Malagelada. 2003. Gut flora in health and disease. Lancet361:512–519.
- Seksik, P., L.Rigottier-Gois, G.Gramet, M.Sutren, P.Pochart, P.Marteau, R.Jian, and J.Dore. 2003. Alterations of the dominant faecal bacterial groups in patients with Crohn’s disease of the colon. Gut52:237–242.
- Zoetendal, E.G., C.T.Collier, S.Koike, R.I.Mackie, and H.R.Gaskins. 2004. Molecular ecological analysis of the gastrointestinal microbiota: a review. J. Nutr.134:465–472.
- Wittwer, C.T., K.M.Ririe, R.V.Andrew, D.A.David, R.A.Gundry, and U.J.Balis. 1997. The LightCycler: a microvolume multisample fluorimeter with rapid temperature control. BioTechniques22:176–181.
- Ott, S.J., M.Musfeldt, U.Ullmann, J.Hampe, and S.Schreiber. 2004. Quantification of intestinal bacterial populations by real-time PCR with a universal primer set and minor groove binder probes: a global approach to the enteric flora. J. Clin. Microbiol.42:2566–2572.
- Koshkin, A.A., K.S.Sanajay, P.Nieslen, K.V.Rajwanshi, R.Kumar, M.Meldgaard, C.E.Olsen, and J.Wengel. 1998. LNA (locked nucleic acid): synthesis of the adenin, cytosin, guanine, 5-methylcytosin, thymidin and uracil bicyclonucleosid monomers,oligomerisationand unprecedented nucleic acid recognition. Tetrahedron54:3607–3630.
- Johnson, M.P., L.M.Haupt, and L.R.Griffiths. 2004. Locked nucleic acid (LNA) single nucleotide polymorphism (SNP) genotype analysis and validation using real-time PCR. Nucleic Acids Res.32:e55.
- Letertre, C., S.Perelle, F.Dilasser, K.Arar, and P.Fach. 2003. Evaluation of the performance of LNA and MGB probes in 5′-nuclease PCR assays. Mol. Cell Probes17:307–311.
- Silahtaroglu, A.N., N.Tommerup, and H.Vissing. 2003. FISHing with locked nucleic acids (LNA): evaluation of different LNA/DNA mixmers. Mol. Cell Probes17:165–169.
- Simeonov, A. and T.T.Nikiforov. 2002. Single nucleotide polymorphism genotyping using short fluorescently labeled locked nucleic acid (LNA) probes and fluorescence polarization detection. Nucleic Acids Res.30:e91.
- Tolstrup, N., P.S.Nielsen, J.G.Kolberg, A.M.Frankel, H.Vissing, and S.Kauppinen. 2003. OligoDesign: optimal design of LNA (locked nucleic acid) oligonucleotide capture probes for gene expression profiling. Nucleic Acids Res.31:3758–3762.
- Ugozzoli, L.A., D.Latorra, R.Pucket, K.Arar, and K.Hamby. 2004. Real-time genotyping with oligonucleotide probes containing locked nucleic acids. Anal. Biochem.324:143–152.
- Baker, G.C., J.J.Smith, and D.A.Cowan. 2003. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods55:541–555.

