Abstract
γ-secretase complexes are multisubunit protease complexes that perform the intramembrane cleavage of more than 60 type-I transmembrane proteins, including Notch receptors. Since dysregulated Notch signaling has been implicated in the tumorigenesis and progression of breast cancer, small molecule γ-secretase inhibitors (GSIs) are being tested for their therapeutic potential in breast cancer treatment in several clinical trials. Here, the structure of γ-secretase complex and the development of GSIs are briefly reviewed, the roles of Notch and several other γ-secretase substrates in breast cancer are discussed, and the difference between γ-secretase inhibition and Notch inhibition, as well as the side effects associated with GSIs, are described. A better understanding of molecular mechanisms that affect the responsiveness of breast cancer to GSI might help to develop strategies to enhance the antitumor activity and, at the same time, alleviate the side effects of GSI.
Keywords:
Overview of γ-secretase complexes
γ-secretases are intramembrane multisubunit protease complexes that are composed of four core components (presenilin, nicastrin, presenilin enhancer 2 [Pen2], and anterior pharynx-defective 1 [Aph1]) and several associated proteins such as TMP21, CD147, and Rer1p.Citation1–Citation3 Two presenilin genes and two Aph1 genes have been identified in the human genome. Given that the Aph1a gene can be expressed in two isoforms due to alternative splicing,Citation4 there could be at least six different γ-secretase complexes. It has been shown that γ-secretase complexes containing different presenilin or Aph1 could exhibit distinct activities.Citation5,Citation6
The catalytic subunit presenilins are multipass transmembrane proteins with nine transmembrane domains. The catalytic active site lies between their sixth and seventh transmembrane domains, with two aspartates (D257 and D385 of presenilin 1) conferring the aspartate protease activity to the γ-secretase complex.Citation3 Nicastrin was originally proposed to be the “gatekeeper” of γ-secretase complex,Citation7 but a recent study suggested that it functions to stabilize γ-secretase complexes.Citation8 Pen2 is believed to be required for endoproteolytic processing of inactive full-length presenilin into the catalytically active heterodimer.Citation9 Aph1 was recently shown to associate with γ-secretase substrates even in the absence of intact γ-secretase complex, suggesting that Aph1 may be the substrate receptor for the protease complex.Citation10 The functions of other associated proteins are even less clear.
Substrates of γ-secretases are a growing family of single-pass type I transmembrane proteins with more than 60 identified members.Citation1 Notably, these substrates do not share a consensus sequence. The only known prerequisite for γ-secretase-mediated cleavage of these substrates is a short (<40 amino acids) extracellular domain. Therefore, a proteolytic processing within the ectodomain performed by a disintegrin and metalloproteinase (ADAM) or other sheddases at a site close to the transmembrane domain usually precedes the γ-secretase-mediated cleavage. While the cleavage of some substrates produces functional free ectodomain or intracellular domain (ICD), the γ-secretase-mediated intramembrane cleavage of other substrates might just serve as a clearance mechanism to regulate the abundance of these transmembrane proteins.Citation2,Citation3
γ-secretase inhibitor (GSI)
Due to their therapeutic potential in the treatment of Alzheimer’s disease where γ-secretase-mediated processing of β-amyloid precursor protein generates amyloid-β peptide, the key component of amyloid plaques in Alzheimer’s disease,Citation11 considerable effort has been made to develop potent and selective GSIs. As a result, more than 100 GSIs have been synthesized.Citation12,Citation13 They can be classified into transition-state analogs and non-transition-state analog inhibitors based on their structure and binding sites. The transition-state analog GSIs are peptide isosteres that mimic the transition state of substrate cleavage by γ-secretase and bind to the catalytic active site of presenilins. The best characterized first generation GSI of this type is L-685458.Citation14 On the other hand, the binding site for the non-transition-state analog GSIs has yet to be better defined, although a report suggested that they bind to the interface of a γ-secretase complex dimer.Citation15 The two most widely used first generation non-transition-state analog GSIs are N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT)Citation16 and Compound E.Citation17
Recently, new generations of GSIs with enhanced potency and/or improved drug-like properties (such as pharmacokinetics) have been developed and tested in animal studies and clinical trials.Citation13 The first potent GSI with drug-like properties is LY-411575, which is structurally similar to – but 100-fold more potent than – DAPT.Citation18,Citation19 However, LY-411575 has not been tested in clinical trials although it has been used in several animal studies.Citation20,Citation21 In contrast, two GSIs with a structural core derived from LY411575, LY-450139 (semagacestat)Citation22,Citation23 and RO4929097,Citation24 have been or are being tested in several clinical trials. In addition to these azepine-containing GSIs, two sulfonamide-containing non-transition-state analog GSIs, MRK003 and MK0752, have been developed by Merck & Co., Inc. (Whitehouse Station, NJ) and the latter is being tested in several clinical trials.
It should be noted that information regarding the specificity of most GSIs is not publicly available. It has been shown that the cytotoxicity of Z-LLNle-CHO, commonly referred as GSI-I from Calbiochem® (EMD Millipore, Billerica, MA), in breast cancer cells is due to proteasome inhibition, but not γ-secretase inhibition.Citation25 Another commonly used GSI from Calbiochem, Z-IL-CHO (GSI-XII), can also suppress the activity of proteasome at concentrations that show cytotoxicity in breast cancer cells (authors’ unpublished observation). Although it is unlikely that the new generation non-transition-state analog GSIs that are tested in animal studies and clinical trials have proteasome-inhibitory activity, it has been reported that both L-685458 and LY411575 – but not DAPT – can inhibit the activity of signal peptide peptidases (SPPs), a family of aspartyl proteases that is closely related to γ-secretase complex.Citation26 Fewer than ten SPP substrates have been identified with the significance of SPP-mediated processing largely unknown.Citation27 However, a recent study reported that SPP-like 2-mediated tumor necrosis factor-α signal peptide clearance could modulate the response of breast cancer cells to tumor necrosis factor-α signaling.Citation28 Therefore, observations from treatment with GSIs with unknown specificity should be interpreted cautiously.
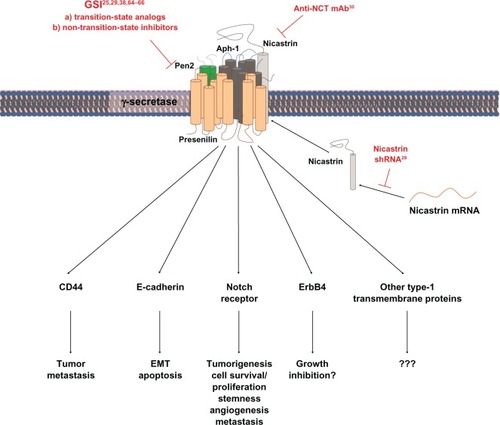
In addition to GSIs, γ-secretase activity has also been downregulated by using ribonucleic acid (RNA) interference to knock down nicastrin or by using neutralizing antibody against nicastrin in vitro ().Citation29,Citation30 These approaches can avoid affecting the activity of γ-secretase-related proteases such as SPP and, therefore, are good complementary methods to examine the outcome of γ-secretase inhibition. Nonetheless, GSI might still be the first choice when it comes to in vivo studies and eventually clinical use due to its convenience to administer.
Notch as γ-secretase substrates in breast cancer
To date, the interest in targeting γ-secretase for cancer treatment stems almost exclusively from the fact that Notch receptors are γ-secretase substrates. Notch receptors are a family of evolutionarily conserved transmembrane proteins and humans have four Notch homologs, Notch1–4. They share similar structural domains, activation process, and mechanism of target gene regulation. Humans also have five classical Notch ligands: Jagged 1, Jagged 2, Delta-like 1 (Dll1), Dll3, and Dll4. In the absence of ligand engagement, cytomembrane-anchored Notch receptors adopt a conformation that masks its ADAM cleavage site. Upon ligand binding, physical force generated from ligand endocytosis exposes the ADAM cleavage site. The C-terminal product of the ADAM-mediated cleavage is a constitutive substrate of γ-secretase complex, which releases the active Notch ICD from the cell membrane. Free Notch ICD translocates into the nucleus to interact with the sequence-specific DNA binding protein, CSL (named after C-promoter binding factor 1, Su[H], and Lag-1), mastermind or mastermind-like proteins, and other transcription coactivators to activate the transcription of Notch target genes.Citation31 In addition to the well-characterized canonical Notch signaling pathway, noncanonical Notch activities have also been reported.Citation32
Notch signaling has been implicated in the tumorigenesis and progression of breast cancer and, consequently, has been proposed to be a promising therapeutic target for breast cancer treatment.Citation33–Citation36 However, discrepancies among key observations raise concerns over the therapeutic potential of blocking Notch signaling, especially through GSIs, in breast cancer.Citation37 Encouragingly, a recent study provided compelling evidence that the Notch pathway is dysregulated in a small subset of breast cancers and these cancers are responsive to γ-secretase inhibition.Citation38 Using paired-end transcriptome sequencing, the authors detected Notch1 or Notch2 involving chromosomal translocation in three out of 38 breast cancer tissues and five out of 41 breast cancer cell lines. Among these eight translocations, three found in cell lines and one found in breast cancer tissues are expected to result in dys-regulated Notch activity. Indeed, all three cell lines showed much higher Notch transcriptional activity. Most importantly, DAPT treatment inhibited the proliferation of the two cell lines that express truncated Notch1 with short extracellular domains C-terminal to the ADAM cleavage site, but not that of the cell line that expresses a truncated Notch2 with no γ-secretase cleavage site as expected.
In addition to regulating cancer cell survival/proliferation cell-autonomously, Notch signaling could also affect the tumor phenotypes through non-cell autonomous mechanisms. The best known example is through angiogenesis, where excessive but nonfunctional angiogenesis triggered by Notch inhibition can suppress tumor growth.Citation39,Citation40 A recent study provided another example of non-cell autonomous effects of Notch signaling on breast cancer.Citation41 Sethi et al showed that tumor-expressed Jagged 1 could promote breast cancer bone metastasis by activating Notch receptors expressed in osteoblasts and osteoclasts. Higher Notch activity in osteoblasts stimulated the release of interleukin-6 from osteoblasts, which in turn promoted the growth of metastatic cancer cells. On the other hand, Notch activation in osteoclasts caused severe osteolysis and consequently provided space for metastatic breast cancer cells to proliferate.Citation41 This inspiring study raises an intriguing possibility that tumor-expressed Notch ligands might also activate Notch signaling in other tumor stromal cells, such as fibroblasts and immune cells, which in turn promote the survival and proliferation of breast cancer cells. Although this hypothesis remains to be rigorously tested, it is consistent with the observation that the expression level of Notch ligands rather than that of Notch receptors in the breast cancer tissues correlates with clinical outcomes.Citation41,Citation42
Other γ-secretase substrates in breast cancer
In addition to Notch receptors and ligands, more than 50 transmembrane proteins have been identified as substrates for γ-secretease.Citation1 Among them, several proteins such as ErbB4, E-cadherin, and CD44 are known to have important functions in breast cancer ().
ErbB4 receptor tyrosine kinase is a member of epidermal growth factor receptor (EGFR) family. In contrast to the well-documented proproliferative effects of other members of EGFR family, the functions of ErbB4 in breast cancer are open to debate.Citation43,Citation44 In some breast cancer cell lines, activation of ErbB4 by its ligand heregulin resulted in its intramembrane cleavage and concomitant growth inhibition.Citation45,Citation46 The intramembrane cleavage of ErbB4 is performed by γ-secretase as the expression of dominant-negative presenilin or treatment with Compound E could prevent the generation of free ErbB4 ICD and reverse the growth inhibition induced by heregulin.Citation46 The significance of γ-secretase in ErbB4 signaling was further confirmed by later studies.Citation47,Citation48 By expressing a mutant ErbB4 (ErbB4-V673I) that does not undergo γ-secretase-mediated cleavage, it was found that some of the ErbB4-induced gene expression and proapoptotic activity were substantially abolished.Citation47 However, it should be noted that ErbB4-V673I still retained some of the ErbB4 activities,Citation47 which might even be enhanced due to a possibly higher level of membrane-anchored ErbB4 when its cleavage is blocked. In addition, the abundance of cell surface ErbB4 might affect the signaling of other members of EGFR family by changing their dimeric association with available EGFR family members. Therefore, it cannot yet be concluded whether γ-secretase inhibition is therapeutically disadvantageous from the ErbB4 perspective.
E-cadherin, a cell–cell adhesion molecule, is also a γ-secretase substrate.Citation49 As a potent tumor suppressor, partial or complete loss of E-cadherin expression has been associated with the onset of metastasis and poor prognosis in breast cancer patients.Citation50 The γ-secretase-mediated cleavage of E-cadherin between Leu731 and Arg732 promotes the disassembly of adherens junctions and increases the level of free cytosolic β-catenin, with the former possibly promoting epithelial-mesenchymal transition and the latter enhancing the Wnt signaling activity.Citation49 Significantly, a later study found that C-terminal fragment 2 of E-cadherin generated from γ-secretase-mediated cleavage could also translocate into the nucleus where it modulates the p120/Kaiso-mediated gene regulation and protects cells from staurosporine-induced apoptosis.Citation51 Furthermore, it was found in another study that exposure of breast cancer T47D cells to cadmium (a toxic heavy metal) induced γ-secretase-mediated E-cadherin cleavage.Citation52 Interestingly, this cadmium-induced γ-secretase-mediated E-cadherin cleavage can be blocked by N-acetylcysteine (an antioxidant), suggesting that other reactive oxygen species-producing reagents might also be able to induce γ-secretase-mediated E-cadherin cleavage. In the later study, it was also observed that cadmium exposure enhanced T47D cell motility and migration, an effect that could be inhibited by DAPT. Taking all these observations together, it appears that blocking γ-secretase activity is therapeutically advantageous from the E-cadherin perspective.
CD44 is another γ-secretase substrate that has been associated with breast cancer tumorigenesis and progression.Citation53,Citation54 As a major cell adhesion molecule for binding hyaluronan in the extracellular matrix, CD44 has been implicated in cell proliferation, differentiation, and migration in both physiological and pathological conditions.Citation55 In breast cancer, CD44 is generally believed to promote tumor metastasis, although the opposite has also been observed.Citation56 Although there is no report on the functional consequence of γ-secretase-mediated CD44 cleavage in breast cancer cells yet, the free ICD (CD44 ICD) generated from such cleavage has been shown to translocate into the nucleus to regulate gene expression,Citation54,Citation57 and be capable of transforming rat fibroblast cells.Citation58 These observations are corroborated by a recent study, which reported that CD44 ICD enhanced cyclic adenosine monophosphate response element-binding-mediated cyclin D1 transcription and, consequently, cell proliferation in thyroid tumor cells.Citation59 Most importantly, γ-secretase inhibition suppressed the proliferation of thyroid cancer cells, which could be reversed by ectopic expression of CD44 ICD. If these results could be replicated in breast cancer cells, it will undoubtedly provide a strong argument for using GSIs to achieve therapeutic benefits.
In addition to the three proteins discussed above, γ-secretase also regulates many other molecules that might have potential roles in tumorigenesis and disease progression. For example, the ICD of amyloid precursor protein has been shown to bind directly to the promoter of EGFR, a receptor tyrosine kinase that is involved in the tumorigenesis and progression of breast cancer, and to suppress its expression.Citation60 In addition, cleavage of low-density lipoprotein receptor-related protein 1B by γ-secretase was shown to suppress the anchorage-independent growth of glioma cells,Citation61 and cleavage of voltage-gated sodium channel β2-subunit by γ-secretase was reported to regulate cell adhesion and migration.Citation62 Although the functional consequences of γ-secretase-mediated cleavage of these substrates in breast cancer cells have not been reported, they should be taken into consideration when developing therapeutics that target γ-secretase.
Targeting γ-secretase versus targeting Notch
Given that several γ-secretase substrates other than Notch receptors also play important roles in the survival, proliferation, and migration of breast cancer cells, it will not be surprising to see that the outcomes of γ-secretase inhibition in breast cancer cells might be different from that of downregulating Notch activity. Indeed, RNA interference-mediated knockdown of individual Notch receptors or ligands produced stronger cytotoxicity/proliferation inhibition in a large collection of breast cancer cell lines than GSIs,Citation25,Citation29,Citation38,Citation63–Citation66 suggesting that blocking the cleavage of some γ-secretase substrates neutralized the tumor-suppressing effect of Notch inhibition. If this is true, identification of this/these substrate(s) may help to improve the therapeutic potential of GSI. However, before this/these γ-secretase substrate(s) can begin to be identified, it is imperative to verify that the difference between Notch inhibition and γ-secretase inhibition is real, as RNA interference-mediated knockdown often suffers from off-target effects.Citation67,Citation68 This can be addressed by testing whether breast cancer cells can be rescued from small interfering RNA-induced proliferation inhibition with expression of small interfering RNA-resistant complementary DNA.
A good start to search for the γ-secretase substrates that functionally antagonize prosurvival Notch signaling, if it turns out to be necessary, is to examine the status of γ-secretase substrates in the two breast cancer cell lines that harbor Notch1 involving translocation.Citation38 Since these two cell lines are responsive to GSI treatment, the expression and/or activity of the γ-secretase substrates, whose reduced cleavage by GSI treatment provides prosurvival/proliferation signal in the GSI nonresponsive breast cancer cell lines, should be significantly downregulated prior to GSI treatment so that its reduced processing by GSI would no longer result in sufficient prosurvival/proliferation signal in these two cell lines. One noticeable feature of these two cell lines is that they grow as weakly adherent culture or in suspension, a sign of reduced expression of cell–cell and/or cell–matrix adhesion molecules. Therefore, it will be interesting to see whether the expression levels of E-cadherin and CD44 are reduced in these two cells.
Side effects from γ-secretase inhibition
Most of the side effects associated with GSI treatment in animal studies or in clinical trials to date can be attributed to reduced Notch activity. Diarrhea was one of the earliest observed side effects of systemic GSI administration in mice.Citation18 Notch activity is essential for maintaining the population of undifferentiated intestinal stem cells and their differentiation into adsorptive enterocytes.Citation69 As a result, Notch inhibition resulted in a rapid increase in the number of secretory goblet cells at the expense of adsorptive cells, and consequently diarrhea. Consistent with animal studies, gastrointestinal tract disorders were also a prominent side effect observed in a phase II clinical trial of semagacestat in Alzheimer’s patients.Citation70 Encouragingly, recent studies have demonstrated that the occurrence of diarrhea could be greatly reduced by intermittent dosing or coadministration of glucocorticoid while the antitumor effect was still maintained in T-cell acute lymphoblastic leukemia mouse models.Citation71,Citation72 Therefore, it is hopeful that gastrointestinal tract disorders would not be a big barrier to using GSI in patients.
Notch plays multiple roles in regulating immune system development and function, including T-cell lineage commitment, marginal zone B-cell development, and activation of peripheral T helper 2 cells.Citation73 Therefore, it is expected that blocking Notch signaling will adversely affect the immune defense. However, although GSI treatment caused several immune-related side effects in mice,Citation18 no increase in the upper respiratory tract and urinary tract infection or seasonal allergy was reported in the phase II clinical trial mentioned above.Citation70 Nonetheless, the frequency of immune system disorders in patients receiving GSI treatment should be closely monitored as immunosuppression might help the spread of tumor cells.
The critical role Notch plays in angiogenesis and vascular homeostasis not only can be exploited for therapeutic benefits, but can also cause severe side effects when Notch signaling is blocked for a prolonged time. Dou et al showed in a mouse model that loss of Notch activity accelerated choroidal neovascularization, a common symptom of wet age-related macular degeneration that results in vision loss in elderly people.Citation74 Additionally, chronic treatment with Dll4 antibody or gradual loss of Notch1 activity through loss of heterogeneity in mouse models triggered development of vascular tumors and lethal hemorrhage.Citation75,Citation76 Of note, vision deterioration occurred in two out of 36 patients receiving semagacestat while none was reported in patients receiving placebo in the phase II clinical trial mentioned above,Citation70 although whether vision deterioration was due to abnormal angiogenesis in retina was not reported.
An even more problematic side effect of blocking Notch signaling for cancer treatment is paradoxically an increase in the risk of developing cancers. It has been well established that loss of Notch activity could trigger skin squamous cell carcinoma (SCC) development through creating a tumor-promoting microenvironment.Citation77,Citation78 Consistent with mouse studies, an increase in the risk of developing skin cancers was observed in two prematurely terminated phase III clinical trials that tested semagacestat in more than 2600 patients.Citation79 In addition to skin SCC, blocking Notch signaling might also increase the risk of developing lung SCC as a recent study suggested that loss-of-function Notch receptor mutation might also contribute to a small population of lung SCC.Citation80
Another notable side effect of semagacestat in the failed clinical trials of Alzheimer’s disease is the worsening of cognition.Citation79 Whether this side effect is specific to Alzheimer’s patients or to the general population is not known, but warrants close monitoring. γ-secretase substrates other than Notch might be responsible for this side effect as Notch activity has not been associated with cognitive symptoms in genetic studies before.
Future direction
A recent publication reported by far the most encouraging therapeutic effects of GSI treatment.Citation81 The authors found that in vitro exposure of primary mammosphere cultures from mammary carcinomas that developed in a transgenic mouse model to MRK003 for 4 days resulted in a dose-dependent decrease in the frequency and the sizes of tumors formed when these primary cultures were injected subcutaneously into syngeneic immune-competent mice. Notably, in cases when a mammosphere that was exposed to a higher dose of MRK003 did develop a tumor, cells prepared from such tumors failed to produce secondary tumors when injected into new hosts, indicating that exposure to MRK003 for 4 days in vitro completely eliminated the long-term tumor initiating cells. More encouragingly, when mice bearing subcutaneous tumors that developed from injected nontreated primary mammosphere culture were treated with MRK003 for two cycles (3 days on followed by 4 days off), tumors in most treated mice completely regressed without recurrence for up to 1 year while the tumors in control mice continued to grow. Although two previous animal studies did not observe such a strong therapeutic effect of MRK003,Citation82,Citation83 this recent success indicates that GSI can be effective if the cases are properly selected, and that some yet-to-be-identified factors affect the responsiveness of breast cancer to GSI treatment and need to be investigated. Through identifying – and modulating the activities of – these contributing factors, it is hopeful that the antitumor activity of GSIs can be potentiated, and its side effects alleviated, so a good therapeutic window can be found for its use in breast cancer patients.
Disclosure
The authors report no conflicts of interest in this work.
Figure 1 Schematic description of approaches that have been used to suppress γ-secretase activity in breast cancer studies and of signaling pathways that are possibly affected by γ-secretase inhibition.
Abbreviations: Aph-1, anterior pharynx-defective 1; EMT, epithelial-mesenchymal transition; GSI, γ-secretase inhibitor; mAb, monoclonal antibody; mRNA, messenger ribonucleic acid; NCT, nicastrin; Pen2, presenilin enhancer 2; shRNA, small hairpin ribonucleic acid.
References
- McCarthyJVTwomeyCWujekPPresenilin-dependent regulated intramembrane proteolysis and gamma-secretase activityCell Mol Life Sci20096691534155519189053
- De StrooperBAnnaertWNovel research horizons for presenilins and γ-secretases in cell biology and diseaseAnnu Rev Cell Dev Biol20102623526020604710
- JorissenEDe StrooperBGamma-secretase and the intramembrane proteolysis of NotchCurr Top Dev Biol20109220123020816396
- GuYChenFSanjoNAPH-1 interacts with mature and immature forms of presenilins and nicastrin and may play a role in maturation of presenilin, nicastrin complexesJ Biol Chem200327897374738012471034
- LaiMTChenECrouthamelMCPresenilin-1 and presenilin-2 exhibit distinct yet overlapping gamma-secretase activitiesJ Biol Chem200327825224752248112684521
- SerneelsLVan BiervlietJCraessaertsKGamma-secretase heterogeneity in the Aph1 subunit: relevance for Alzheimer’s diseaseScience2009324592763964219299585
- ShahSLeeSFTabuchiKNicastrin functions as a gamma-secretase-substrate receptorCell2005122343544716096062
- ZhaoGLiuZIlaganMXKopanRGamma-secretase composed of PS1/Pen2/Aph1a can cleave notch and amyloid precursor protein in the absence of nicastrinJ Neurosci20103051648165620130175
- TakasugiNTomitaTHayashiIThe role of presenilin cofactors in the gamma-secretase complexNature2003422693043844112660785
- ChenACGuoLYOstaszewskiBLSelkoeDJLaVoieMJAph-1 associates directly with full-length and C-terminal fragments of gamma-secretase substratesJ Biol Chem201028515113781139120145246
- HaassCSteinerHAlzheimer disease gamma-secretase: a complex story of GxGD-type presenilin proteasesTrends Cell Biol2002121255656212495843
- OlsonREAlbrightCFRecent progress in the medicinal chemistry of gamma-secretase inhibitorsCurr Top Med Chem200881173318220929
- KreftAFMartoneRPorteARecent advances in the identification of gamma-secretase inhibitors to clinically test the Abeta oligomer hypothesis of Alzheimer’s diseaseJ Med Chem200952206169618819694467
- ShearmanMSBeherDClarkeEEL-685,458, an aspartyl protease transition state mimic, is a potent inhibitor of amyloid beta-protein precursor gamma-secretase activityBiochemistry200039308698870410913280
- ClarkeEEChurcherIEllisSIntra- or intercomplex binding to the gamma-secretase enzyme. A model to differentiate inhibitor classesJ Biol Chem200628142312793128916899457
- DoveyHFJohnVAndersonJPFunctional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brainJ Neurochem200176117318111145990
- SeiffertDBradleyJDRomingerCMPresenilin-1 and -2 are molecular targets for gamma-secretase inhibitorsJ Biol Chem200027544340863409110915801
- WongGTManfraDPouletFMChronic treatment with the gamma-secretase inhibitor LY-411,575 inhibits beta-amyloid peptide production and alters lymphopoiesis and intestinal cell differentiationJ Biol Chem200427913128761288214709552
- LewisHDPerez RevueltaBINadinACatalytic site-directed gamma-secretase complex inhibitors do not discriminate pharmacologically between Notch S3 and beta-APP cleavagesBiochemistry200342247580758612809514
- LanzTAHosleyJDAdamsWJMerchantKMStudies of Abeta pharmacodynamics in the brain, cerebrospinal fluid, and plasma in young (plaque-free) Tg2576 mice using the gamma-secretase inhibitor N2-[(2S)-2-(3,5-difluorophenyl)-2-hydroxyethanoyl]-N1-[(7S)-5-methyl-6-oxo-6,7-di hydro-5H-dibenzo[b,d]azepin-7-yl]-L-alanin-amide (LY-411575)J Pharmacol Exp Ther20043091495514718585
- HydeLAMcHughNAChenJStudies to investigate the in vivo therapeutic window of the gamma-secretase inhibitor N2-[(2S)-2-(3,5-difluorophenyl)-2-hydroxyethanoyl]-N1-[(7S)-5-methyl-6-oxo-6,7-di hydro-5H-dibenzo[b,d]azepin-7-yl]-L-alaninamide (LY411,575) in the CRND8 mouseJ Pharmacol Exp Ther200631931133114316946102
- SiemersESkinnerMDeanRASafety, tolerability, and changes in amyloid beta concentrations after administration of a gamma-secretase inhibitor in volunteersClin Neuropharmacol200528312613215965311
- HenleyDBMayPCDeanRASiemersERDevelopment of semagacestat (LY450139), a functional gamma-secretase inhibitor, for the treatment of Alzheimer’s diseaseExpert Opin Pharmacother200910101657166419527190
- LuistroLHeWSmithMPreclinical profile of a potent gamma-secretase inhibitor targeting notch signaling with in vivo efficacy and pharmacodynamic propertiesCancer Res200969197672768019773430
- HanJMaIHendzelMJAllalunis-TurnerJThe cytotoxicity of gamma-secretase inhibitor I to breast cancer cells is mediated by proteasome inhibition, not by gamma-secretase inhibitionBreast Cancer Res2009114R5719660128
- WeihofenALembergMKFriedmannETargeting presenilin-type aspartic protease signal peptide peptidase with gamma-secretase inhibitorsJ Biol Chem200327819165281653312621027
- GoldeTEWolfeMSGreenbaumDCSignal peptide peptidases: a family of intramembrane-cleaving proteases that cleave type 2 trans-membrane proteinsSemin Cell Dev Biol200920222523019429495
- YanDQinNZhangHExpression of TNF-alpha leader sequence renders MCF-7 tumor cells resistant to the cytotoxicity of soluble TNF-alphaBreast Cancer Res Treat200911619110218618239
- DongYLiAWangJWeberJDMichelLSSynthetic lethality through combined Notch-epidermal growth factor receptor pathway inhibition in basal-like breast cancerCancer Res201070135465547420570903
- FilipovicAGronauJHGreenARBiological and clinical implications of nicastrin expression in invasive breast cancerBreast Cancer Res Treat20111251435320224929
- KopanRIlaganMXThe canonical Notch signaling pathway: unfolding the activation mechanismCell2009137221623319379690
- HeitzlerPBiodiversity and noncanonical Notch signalingCurr Top Dev Biol20109245748120816404
- Al-HussainiHSubramanyamDReedijkMSridharSSNotch signaling pathway as a therapeutic target in breast cancerMol Cancer Ther201110191520971825
- RizzoPOsipoCForemanKGoldeTOsborneBMieleLRational targeting of Notch signaling in cancerOncogene200827385124513118758481
- ShiWHarrisALNotch signaling in breast cancer and tumor angiogenesis: cross-talk and therapeutic potentialsJ Mammary Gland Biol Neoplasia2006111415216947085
- HarrisonHFarnieGBrennanKRClarkeRBBreast cancer stem cells: something out of notching?Cancer Res201070228973897621045140
- HanJHendzelMJAllalunis-TurnerJNotch signaling as a therapeutic target for breast cancer treatment?Breast Cancer Res201113321021672271
- RobinsonDRKalyana-SundaramSWuYMFunctionally recurrent rearrangements of the MAST kinase and Notch gene families in breast cancerNat Med201117121646165122101766
- RidgwayJZhangGWuYInhibition of Dll4 signalling inhibits tumour growth by deregulating angiogenesisNature200644471221083108717183323
- Noguera-TroiseIDalyCPapadopoulosNJBlockade of Dll4 inhibits tumour growth by promoting non-productive angiogenesisNature200644471221032103717183313
- SethiNDaiXWinterCGKangYTumor-derived JAGGED1 promotes osteolytic bone metastasis of breast cancer by engaging notch signaling in bone cellsCancer Cell201119219220521295524
- XingFOkudaHWatabeMHypoxia-induced Jagged2 promotes breast cancer metastasis and self-renewal of cancer stem-like cellsOncogene201130394075408621499308
- SundvallMIljinKKilpinenSSaraHKallioniemiOPEleniusKRole of ErbB4 in breast cancerJ Mammary Gland Biol Neoplasia200813225926818454307
- ChuuCPChenRYBarkingeJLCiaccioMFJonesRBSystems-level analysis of ErbB4 signaling in breast cancer: a laboratory to clinical perspectiveMol Cancer Res20086688589118567793
- SartorCIZhouHKozlowskaEHer4 mediates ligand-dependent antiproliferative and differentiation responses in human breast cancer cellsMol Cell Biol200121134265427511390655
- NiCYMurphyMPGoldeTECarpenterGGamma-secretase cleavage and nuclear localization of ErbB-4 receptor tyrosine kinaseScience200129455492179218111679632
- VidalGANareshAMarreroLJonesFEPresenilin-dependent gamma-secretase processing regulates multiple ERBB4/HER4 activitiesJ Biol Chem200528020197771978315746097
- FengSMSartorCIHunterDThe HER4 cytoplasmic domain, but not its C terminus, inhibits mammary cell proliferationMol Endocrinol20072181861187617505063
- MarambaudPShioiJSerbanGA presenilin-1/gamma-secretase cleavage releases the E-cadherin intracellular domain and regulates disassembly of adherens junctionsEMBO J20022181948195611953314
- BerxGVan RoyFThe E-cadherin/catenin complex: an important gatekeeper in breast cancer tumorigenesis and malignant progressionBreast Cancer Res20013528929311597316
- FerberECKajitaMWadlowAA role for the cleaved cytoplasmic domain of E-cadherin in the nucleusJ Biol Chem200828319126911270018356166
- ParkCSKimOSYunSMJoSAJoIKohYHPresenilin 1/gamma-secretase is associated with cadmium-induced E-cadherin cleavage and COX-2 gene expression in T47D breast cancer cellsToxicol Sci2008106241342218791180
- LammichSOkochiMTakedaMPresenilin-dependent intramembrane proteolysis of CD44 leads to the liberation of its intracellular domain and the secretion of an Abeta-like peptideJ Biol Chem200227747447544475912223485
- MurakamiDOkamotoINaganoOPresenilin-dependent gamma-secretase activity mediates the intramembranous cleavage of CD44Oncogene200322101511151612629514
- MarhabaRZollerMCD44 in cancer progression: adhesion, migration and growth regulationJ Mol Histol200435321123115339042
- LouderboughJMSchroederJAUnderstanding the dual nature of CD44 in breast cancer progressionMol Cancer Res20119121573158621970856
- OkamotoIKawanoYMurakamiDProteolytic release of CD44 intracellular domain and its role in the CD44 signaling pathwayJ Cell Biol2001155575576211714729
- PelletierLGuillaumotPFrecheBGamma-secretase-dependent proteolysis of CD44 promotes neoplastic transformation of rat fibroblastic cellsCancer Res20066673681368716585194
- De FalcoVTamburrinoAVentreSCD44 proteolysis increases CREB phosphorylation and sustains proliferation of thyroid cancer cellsCancer Res20127261449145822271686
- ZhangYWWangRLiuQZhangHLiaoFFXuHPresenilin/gamma-secretase-dependent processing of beta-amyloid precursor protein regulates EGF receptor expressionProc Natl Acad Sci U S A200710425106131061817556541
- LiuCXRanganathanSRobinsonSStricklandDKGamma-secretase-mediated release of the low density lipoprotein receptor-related protein 1B intracellular domain suppresses anchorage-independent growth of neuroglioma cellsJ Biol Chem2007282107504751117227771
- KimDYInganoLACareyBWPettingellWHKovacsDMPresenilin/gamma-secretase-mediated cleavage of the voltage-gated sodium channel beta2-subunit regulates cell adhesion and migrationJ Biol Chem200528024232512326115833746
- YamaguchiNOyamaTItoENOTCH3 signaling pathway plays crucial roles in the proliferation of ErbB2-negative human breast cancer cellsCancer Res20086861881188818339869
- RizzoPMiaoHD’SouzaGCross-talk between notch and the estrogen receptor in breast cancer suggests novel therapeutic approachesCancer Res200868135226523518593923
- OsipoCPatelPRizzoPErbB-2 inhibition activates Notch-1 and sensitizes breast cancer cells to a gamma-secretase inhibitorOncogene200827375019503218469855
- YongTSunAHenryMDMeyersSDavisJNDown regulation of CSL activity inhibits cell proliferation in prostate and breast cancer cellsJ Cell Biochem201111292340235121520243
- JacksonALBartzSRSchelterJExpression profiling reveals off-target gene regulation by RNAiNat Biotechnol200321663563712754523
- ScacheriPCRozenblatt-RosenOCaplenNJShort interfering RNAs can induce unexpected and divergent changes in the levels of untargeted proteins in mammalian cellsProc Natl Acad Sci U S A200410171892189714769924
- van EsJHvan GijnMERiccioONotch/gamma-secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cellsNature2005435704495996315959515
- FleisherASRamanRSiemersERPhase 2 safety trial targeting amyloid beta production with a gamma-secretase inhibitor in Alzheimer diseaseArch Neurol20086581031103818695053
- TammamJWareCEffersonCDown-regulation of the Notch pathway mediated by a gamma-secretase inhibitor induces anti-tumour effects in mouse models of T-cell leukaemiaBr J Pharmacol200915851183119519775282
- RealPJToselloVPalomeroTGamma-secretase inhibitors reverse glucocorticoid resistance in T cell acute lymphoblastic leukemiaNat Med2009151505819098907
- RadtkeFFasnachtNMacdonaldHRNotch signaling in the immune systemImmunity2010321142720152168
- DouGRWangYCHuXBRBP-J, the transcription factor downstream of Notch receptors, is essential for the maintenance of vascular homeostasis in adult miceFASEB J20082251606161718096813
- Ya nMCallahanCABeyerJCChronic DLL4 blockade induces vascular neoplasmsNature20104637282E6E720147986
- LiuZTurkozAJacksonENNotch1 loss of heterozygosity causes vascular tumors and lethal hemorrhage in miceJ Clin Invest2011121280080821266774
- DemehriSTurkozAKopanREpidermal Notch1 loss promotes skin tumorigenesis by impacting the stromal microenvironmentCancer Cell2009161556619573812
- NicolasMWolferARajKNotch1 functions as a tumor suppressor in mouse skinNat Genet200333341642112590261
- ExtanceAAlzheimer’s failure raises questions about disease-modifying strategiesNat Rev Drug Discov201091074975120885394
- WangNJSanbornZArnettKLLoss-of-function mutations in Notch receptors in cutaneous and lung squamous cell carcinomaProc Natl Acad Sci U S A201110843177611776622006338
- KondratyevMKresoAHallettRMGamma-secretase inhibitors target tumor-initiating cells in a mouse model of ERBB2 breast cancerOncogene20123119310321666715
- EffersonCLWinkelmannCTWareCDownregulation of Notch pathway by a gamma-secretase inhibitor attenuates AKT/mammalian target of rapamycin signaling and glucose uptake in an ERBB2 transgenic breast cancer modelCancer Res20107062476248420197467
- WattersJWChengCMajumderPKDe novo discovery of a gamma-secretase inhibitor response signature using a novel in vivo breast tumor modelCancer Res200969238949895719903844