
Abstract
Globally, breast cancer is the most common cancer type and is one of the most significant causes of deaths in women. To date, multiple clinical interventions have been applied, including surgical resection, radiotherapy, endocrine therapy, targeted therapy and chemotherapy. However, 1) the lack of therapeutic options for metastatic breast cancer, 2) resistance to drug therapy and 3) the lack of more selective therapy for triple-negative breast cancer are some of the major challenges in tackling breast cancer. Given the safe nature of natural products, numerous studies have focused on their anti-cancer potentials. Mangifera indica, commonly known as mango, represents one of the most extensively investigated natural sources. In this review, we provide a comprehensive overview of M. indica extracts (bark, kernel, leaves, peel and pulp) and phytochemicals (mangiferin, norathyriol, gallotannins, gallic acid, pyrogallol, methyl gallate and quercetin) reported for in vitro and in vivo anti-breast cancer activities and their underlying mechanisms based on relevant literature from several scientific databases, including PubMed, Scopus and Google Scholar till date. Overall, the in vitro findings suggest that M. indica extracts and/or phytochemicals inhibit breast cancer cell growth, proliferation, migration and invasion as well as trigger apoptosis and cell cycle arrest. In vivo results demonstrated that there was a reduction in breast tumor xenograft growth. Several potential mechanisms underlying the anti-breast cancer activities have been reported, which include modulation of oxidative status, receptors, signalling pathways, miRNA expression, enzymes and cell cycle regulators. To further explore this medicinal plant against breast cancer, future research directions are addressed. The outcomes of the review revealed that M. indica extracts and their phytochemicals may have potential benefits in the management of breast cancer in women. However, to validate its utility in the creation of innovative and potent therapeutic agents to treat breast cancer, more dedicated research, especially clinical studies are needed to explore the anti-breast cancer potentials of M. indica extracts and their phytochemicals.
Introduction
Globally, breast cancer remains one of the most common cancers and causes of deaths in females.Citation1 In 2020 alone, there were 2,261,419 new cases and 684,996 deaths globally.Citation2 In fact, most breast cancer cases are diagnosed at early stages when the disease is deemed as relatively curable.Citation3 However, approximately 20–30% of these patients suffer from distant relapse with cancer cells spreading from the primary site to distant body parts (eg, bones, brain, distant nodal, liver, lungs and pleural/peritoneal metastases).Citation3
The development of breast cancer is thought to be related to several important risk factors; with aging playing the major role, followed by family history (eg, BRCA1/2 mutations), reproductive factors (eg, early menarche and late menopause), hormonal imbalances (eg, elevated estrogen levels) and lifestyle (eg, high fat intake and alcohol consumption).Citation4 In fact, only 5–10% of breast cancer cases are thought to have a genetic background, with BRCA1/2 germline mutations contributing to approximately 50% of these hereditary cases.Citation5,Citation6
Breast cancer is a heterogenous disease owing to the existence of molecular and phenotypic variations within a patient’s tumor (intra-tumoral heterogeneity) and between different patients’ tumors (inter-tumoral heterogeneity).Citation7 Based on immunohistochemistry and gene expression profiling, breast cancer can be classified into different biological subtypes.Citation8 The main subtype classification system is gene expression profile-based by which tumors are classified into 1) the estrogen receptor (ER)-positive (ie, luminal A and B) group (70%), 2) the human epidermal growth factor receptor 2 (HER2)-enriched group (15–20%) or 3) the basal-like [or triple-negative; ER-, progesterone receptor (PR)- and HER2-negative] group (15%).Citation9 Such subtype classification is clinically important in informing treatment options since these subtypes demonstrate variations in their prognosis and treatment responses ().Citation9,Citation10 Although endocrine therapy is usually effective in most patients with ER-positive tumors, 15–20% and 30–40% of all ER-positive tumors exhibit intrinsic and acquired resistance to the therapy, respectively.Citation11 Triple-negative breast cancer, the subtype known to possess a more aggressive nature, particularly lacks targeted therapy and is reliant on chemotherapy that is non-specific,Citation12 thus creating the need for more selective and effective agents for breast cancer treatment.
Table 1 Typical Systemic Therapeutic Options for the Three Major Molecular Subtypes of Breast Cancer
Natural products are an important source for the discovery of new anti-cancer agents that may provide long-term cancer control with minimal side effects.Citation13 Isolation of paclitaxel from the bark of Pacific yew trees (Taxus brevifolia) or vinblastine and vincristine from the leaves of Madagascar periwinkle plants (Catharanthus rosea) are some of the many examples.Citation14 Fish, green tea, medicinal plants and fruits have been explored for their potentials in breast cancer treatment, with Mangifera indica (M. indica) being one example.Citation15
M. indica, commonly known as mango, belongs to the Anacardiaceae flowering plant family.Citation16 Although initially thought to have originated from India, M. indica has been widely cultivated in Southeast Asia.Citation16 Different M. indica plant parts contain varying types and quantities of phytochemicalsCitation17 (), and they have traditionally been exploited for the treatment of several medical conditions, including gastrointestinal, genitourinary, ophthalmic and respiratory conditions.Citation18,Citation19 Preclinical studies on the extracts prepared from various plant parts have demonstrated anti-cancer, anti-inflammatory, antimicrobial, antioxidant and immunomodulatory activities.Citation16,Citation20–Citation23 Specifically, various studies have reported on the anti-cancer activities of M. indica pulp extracts in breast cancer,Citation24–Citation26 thus suggesting the consumption of M. indica fruits may potentially be beneficial in breast cancer management. Further, the phytochemical profiling of M. indica extracts has suggested that their anti-cancer activities are attributed mainly to the polyphenolic contents.Citation27 Therefore, the phytochemicals have the potentials to be developed as anti-cancer drugs.
Table 2 Type of Compounds Identified from Different Parts of M. indica
This review described preclinical studies conducted on various M. indica extracts and phytochemicals () against breast cancer. The proposed mechanisms for their anti-breast cancer activity were then discussed in detail. Although M. indica or its derived products have not been evaluated in clinical studies for breast cancer treatment, they have been clinically evaluated for central nervous system activities,Citation28 chronic constipation,Citation29 gastrointestinal and upper respiratory tract infectionsCitation30 and rheumatoid arthritis,Citation31 etc. Hence, future research directions are addressed in order to further explore this medicinal plant and its phytochemicals against breast cancer.
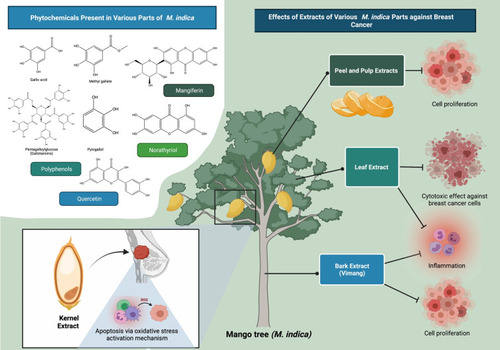
Figure 1 An overview of effects of M. indica extracts and their phytochemicals against breast cancer. Extracts of various M. indica parts (bark, kernel, leaves, peel and pulp) have been used to treat a variety of medical problems including breast cancer; as these plant parts contain varying types and amounts of polyphenols, some of which possess anti-proliferative and pro-apoptotic activities. According to phytochemical profile analysis, the anti-cancer activity of M. indica extracts is mostly attributable to the presence of polyphenolic compounds such as mangiferin, gallotannins, gallic acid, pyrogallol, methyl gallate and quercetin.
Methods
Relevant literature was collected from several scientific databases, including PubMed, Scopus and Google Scholar. The literature search was achieved using the following keywords: “Mango OR Mangifera indica OR Mangiferin AND Breast Cancer”. Papers not written in the English language or without abstracts were excluded from initial screening. Any duplicate was also excluded. Following complete screening and selection of appropriate articles, the yielded information was summarised for 1) the anti-cancer activities of M. indica extracts or their phytochemicals in in vitro and in vivo breast cancer models and 2) the potential mechanisms underlying the bioactivities. All key findings from the selected articles were summarised in and detailed in the following sections.
Table 3 A Summary of in vitro and in vivo Studies of M. indica Extracts and Their Phytochemicals Evaluated for Anti-Breast Cancer Potentials
Effect of Extracts of Various M. indica Parts Against Breast Cancer
Bark Extract of M. indica
Vimang is an aqueous M. indica bark extract consisting of a mixture of polyphenols, fatty acids (FAs), microelements, steroids and terpenoids.Citation32,Citation33 It is a registered anti-inflammatory phytomedicine used either as cream, tablets or syrup in Cuba, where its industrial-scale production has been carried out.Citation34,Citation35 Vimang has shown various pharmacological activities such as analgesic, anti-cancer, anti-inflammatory, antioxidant and immunomodulatory activities.Citation23,Citation36,Citation37 One of its popular uses in Cuba is to improve the quality of life of cancer patients, thus suggesting its potential use in cancer treatment.Citation38 A study showed that Vimang can significantly inhibit the proliferation of MDA-MB-231 (triple-negative) cells in a dose-dependent manner, with half-maximal inhibitory concentration (IC50) at 259 µg/mL.Citation34
Kernel Extract of M. indica
A relevant study reported the ability of M. indica kernel extract to induce a dose-dependent reduction in the viability of MCF-7 (luminal A; ER-positive, PR-positive, HER2-negative), MDA-MB-231 and MCF-10A (normal human mammary epithelial cell line) cells.Citation39 The IC50 values of kernel extract were significantly lower for the cancer cells (15 µg/mL for MCF-7 and 30 µg/mL for MDA-MB-231 cells) than the normal cells (149 µg/mL), suggesting that the kernel extract is cancer cell-selective.Citation39 Subsequent phytochemical profiling of the extract detected 12 major compounds,Citation39 some of which have demonstrated anti-breast cancer potentials in previous studies.Citation40–Citation47
Leaf Extract of M. indica
An emerging body of evidence has suggested the association between chronic inflammation and cancer risk.Citation48M. indica leaf tea has traditionally been used to ameliorate inflammation.Citation27 The polyphenols believed to be the major contributors to M. indica’s anti-cancer activity are present mainly in the bark and leaves.Citation49 Nevertheless, to date, cancer-related studies have generally been focussed on M. indica bark, peel and pulp extracts. A study revealed the cytotoxic effect of ethanolic M. indica leaf extract (200 µg/mL) on BT-474 (luminal B; ER-/PR-/HER2-positive) cells.Citation50 In another study, it was observed that while PEW (50% ethanol:50% water) and FEW (50% ethanol:50% water fraction previously extracted with supercritical carbon dioxide) extracts (0.01–10 µg/mL) with a higher methyl gallate and homomangiferin content are more cytotoxic to MDA-MB-231 cells, PET (pure ethanol) and CEW (50% carbon dioxide:25% ethanol:25% water) extracts (0.01–0.1 µg/mL) with a higher gallotannin content are more cytotoxic to MCF-7 cells.Citation27 This observation suggests that phytochemicals can influence the selectivity of extracts towards different subtypes of breast cancer cells.Citation27 Additionally, PET, CEW, PEW and FEW extracts caused only slight reduction in the viability of MCF-10 cells, suggesting that the leaf extracts are cancer cell-selective.Citation27
Peel and Pulp Extracts of M. indica
The anti-breast cancer activities of M. indica peel and pulp extracts have been more frequently reported compared to the other plant parts. García-Solís et alCitation25 reported the ability of aqueous M. indica pulp extract (4% v/v) to induce a significant reduction in MCF-7 cell proliferation. In other studies, Wilkinson et alCitation26 and Shaban et alCitation51 observed a significant reduction in MCF-7 cell viability following treatment with ethyl acetate fractions prepared from M. indica peels and pulps as well as ethanolic M. indica peel extract, respectively. On the other hand, Banerjee et alCitation24 reported the ability of M. indica pulp extract (2.5–20.0 mg GAE/L) to dose-dependently inhibit BT-474 cell proliferation. In the same study, M. indica pulp extract (0.8 mg GAE/day; 35 days) also significantly reduced tumor volume in nude mice bearing BT-474 xenografts.
As different M. indica varieties have different phytochemical profiles and bioactivities, two studies have evaluated the anti-breast cancer activities of methanolic peel and pulp extracts prepared from three M. indica varieties (Irwin, Kensington Pride and Nam Doc Mai).Citation52,Citation53 Both studies consistently reported Nam Doc Mai peel extract as the only extract with significant cytotoxicity against MCF-7 cells. Additionally, only Nam Doc Mai peel extract was observed to possess significant cell-death inducing activity in MDA-MB-231 cells.Citation52 Previous phytochemical profiling of M. indica peel and pulp extracts revealed that Nam Doc Mai peel extract uniquely contained gallic acid and galloylated derivatives as well as a higher level of methyl gallate,Citation54 which may explain its higher bioactivities.
Effect of Phytochemicals from M. indica Against Breast Cancer
Polyphenols
The anti-cancer activity of M. indica has frequently been attributed to its polyphenolic content. Noratto et alCitation55 reported that the polyphenolic extracts prepared from different M. indica varieties suppressed MDA-MB-231 cell growth in a dose-dependent manner. In another study conducted by Nemec et al,Citation56 a low molecular weight fraction of M. indica polyphenols (1 mg/L) was found to significantly reduce the proliferation of MCF10DCIS [a ductal carcinoma in situ (DCIS) model cell line] cells. In the same study, combined treatment with M. indica polyphenols and a conventional anti-cancer agent [5-fluorouracil (5-FU)] demonstrated a greater anti-proliferative effect on MCF10DCIS cells than either treatment alone, which is suggestive of a synergistic effect. The same research group did a follow-up study on the in vivo anti-cancer effects of M. indica polyphenols (0.8 mg GAE/day; 4 weeks) in nude mice carrying MCF10DCIS xenografts, whereby a reduction in tumor volume (>70%) was observed following treatment.Citation57
Various studies have investigated into the anti-breast cancer activities of individual M. indica polyphenols, instead of M. indica polyphenolic mixtures, as described below.
Mangiferin
Mangiferin (1,3,6,7-tetrahydroxyxanthone-C2-β-D-glucoside), a plant natural xanthonoid, is the predominant phytochemical found in various M. indica parts, including bark, fruits, leaves and roots.Citation58,Citation59 Mangiferin has been reported to exhibit various pharmacological activities such as analgesic, anti-bacterial, anti-cancer, anti-diabetic, anti-inflammatory, antioxidant and immunomodulatory activities.Citation59–Citation64 Importantly, mangiferin has shown protective effects against various types of cancers, including breast cancer.Citation65
As reported by Li et al,Citation66 mangiferin (75–300 µM) caused a dose-dependent reduction in the proliferation of ER-positive (MCF-7 and T47D) and ER-negative (MDA-MB-231 and BT-549) breast cancer cells, which may be attributed to an increase in apoptosis. In the same study, mangiferin (12.5–50.0 µM) also demonstrated dose-dependent anti-migratory and anti-invasive effects on highly metastatic MDA-MB-231 and BT-549 cells. When the in vivo anticancer activity of mangiferin (100 mg/kg/day; 5 weeks) was evaluated in severe-combined immunodeficiency (SCID) mice bearing MDA-MB-231 xenografts, significant reduction in both tumor volume and tumor weight was observed.Citation66
The cytotoxic, anti-migratory and anti-invasive effects of mangiferin (10–50 µM) have similarly been reported in another study by Deng et al,Citation67 whereby a dose-dependent reduction in the viability of MCF-7 and MDA-MB-231 cells as well as a significant reduction in the migration and invasion of MDA-MB-231 cells were observed. Notably, mangiferin has also been reported by Cuccioloni et alCitation68 to be cancer cell-selective, as evidenced by the observations of greater decreases in cell viability, proliferating cell nuclear antigen (PCNA; a marker of cell proliferation) level and anti-apoptotic protein [B-cell lymphoma 2 (Bcl-2)] level as well as greater increases in pro-apoptotic protein [Bcl-2-associated X protein (Bax), caspase 9 and p27] levels in MCF-7 cells than MCF-10A cells following treatment. The same study also showed that mangiferin could suppress plasmin-induced increase in MCF-7 and MCF-10A cell migration.
Doxorubicin, an anthracycline drug, has been used routinely for the treatment of various cancers, including breast cancer.Citation69 However, its use has been limited mainly due to cardiotoxicity and resistance.Citation70 One of the causes of doxorubicin resistance is the increased expression of drug efflux pumps such as P-glycoprotein (Pgp), multidrug resistance–associated protein-1 (MRP-1) and breast cancer resistance protein (BCRP).Citation71–Citation73 Louisa et alCitation74 found that mangiferin can increase the sensitivity of MCF-7 cells (pre-treated with doxorubicin) towards doxorubicin treatment, as evidenced by a significant reduction in the viability of MCF-7 cells treated with mangiferin (10–50 µM) in combination with doxorubicin. Subsequent evaluation of Pgp, MRP1 and BCRP gene expressions showed that while high dose (50 µM) mangiferin could significantly downregulate Pgp, there were no significant changes in the expressions of MRP1 and BCRP at all doses.Citation74 These findings thus suggest that high-dose mangiferin can potentially serve as a chemosensitizer for doxorubicin by downregulating Pgp expression.Citation74
In contrast to the above-mentioned studies, a number of studies have failed to observe the anti-breast cancer activity of mangiferin,Citation34,Citation75 thus warranting further investigation. In one of the studies, Wilkinson et alCitation75 showed that mangiferin is capable of activating ERα but not ERβ, and suggested that the observed lack of anti-proliferative activity of mangiferin may be linked to the differential activation or heterodimer formation of ERα and ERβ.
Norathyriol
Norathyriol (1,3,6,7-tetrahydroxyxanthone), an aglycone derivative of mangiferin, is structurally similar to mangiferin except for a C-glucosyl bond.Citation76 Ample of evidence has shown that orally consumed mangiferin is firstly subjected to de-glycosylation by intestinal bacterium to form norathyriol prior to being absorbed by the colon, suggesting norathyriol is likely to contribute to the observed bioactivities of mangiferin.Citation77,Citation78 Studies focussing on norathyriol have reported a range of pharmacological activities.Citation79–Citation83 For example, a study showed that norathyriol (100 µM) significantly reduced the viability of MCF-7 cells.Citation75
Gallotannins, Gallic Acid, Pyrogallol and Methyl Gallate
M. indica is a rich source of gallotannins and lower molecular weight gallates such as gallic acid.Citation56 Although orally consumed gallotannins are un-absorbable owing to their high molecular weight, they can be hydrolysed in the gastrointestinal tract by tannase to release free gallic acid, which can subsequently undergo decarboxylation to pyrogallol via the activity of gallic acid decarboxylase in intestinal microbiota.Citation84,Citation85 Gallotannins have been reported to exhibit a wide range of pharmacological activities, including anti-cancer,Citation86–Citation88 anti-diabetic,Citation89 anti-inflammatory,Citation90 antimicrobial,Citation91 and antioxidantCitation92 activities. Pentagalloylglucose (1,2,3,4,6-penta-O-galloyl-β-D-glucose; 5GG), a gallotannin that can be found in M. indica, has been investigated for its effects on breast cancer cell lines.Citation93 For instance, treatment with 5GG (0.5–80 µM) was reported to exhibit growth-inhibitory effect on MCF-7 cells in both time- and dose-dependent manner.Citation94 Another study similarly reported the dose-dependent growth-inhibitory effect of 5GG (1–100 µM) on MCF-7 cells.Citation95 Further cell cycle analysis revealed a dramatic increase in G1-phase cell number, suggesting that 5GG can induce a G1-phase arrest in MCF-7 cells.Citation95
Gallic acid (3,4,5-trihydroxybenzoic acid), one of the most abundantly distributed plant phenolic compounds that can be found in certain fruits and medicinal plants, has been used extensively in both food and pharmaceutical industries.Citation96 Gallic acid has demonstrated various pharmacological activities.Citation97–Citation101 Its anti-cancer activity, in particular, has been extensively studied in various cancer types.Citation102–Citation110 In a study, MDA-MB-231 cells treated with gallic acid showed a significant dose-dependent reduction in cell viability, with IC50 at 10 µg/mL.Citation34 Another study reported the ability of gallic acid (1–10 or 2–12 µg/mL) to induce a reduction in cell proliferation and an increase in S-phase and G2/M-phase ratios in MCF-7 cells in a dose-dependent manner.Citation111 Similarly, the anti-proliferative activity of gallic acid (1mg/L) has also been demonstrated in MCF10DCIS cells.Citation56
Of note, Ke Wang et alCitation112 observed a dose-dependent reduction in MCF-7 cell growth following gallic acid treatment (IC50: 80.5 µM) and attributed this effect to apoptosis induction. Subsequent analyses revealed increased Fas cell surface death receptor (Fas) and Fas ligand (FasL) protein levels and caspase 8 activity, suggesting the induction of death receptor apoptotic pathway.Citation112 Reduced mitochondrial membrane potential (ΔΨm) as well as increased Bax/Bcl-2 ratio, cytosolic cytochrome c level and caspase 9 activity were also observed, which was suggestive of the induction of mitochondrial apoptotic pathway.Citation112 Moreover, caspase 8-mediated Bid cleavage, which is known to be followed by cytochrome c release and caspase 9 activation, was observed in gallic acid-treated MCF-7 cells.Citation112 Collectively, these findings suggest that gallic acid-induced apoptosis of MCF-7 cells involved both mitochondrial and death receptor pathways as well as a cross-link between the two pathways.Citation112
Pyrogallol (1,2,3-trihydroxybenzene), a microbial metabolite of M. indica gallotannins, can also be found naturally in many other fruits and vegetables.Citation113 Pyrogallol is used commercially mainly in the production of pharmaceuticals and pesticides.Citation114 Similar to gallotannins and gallic acid, pyrogallol has been found to possess multiple pharmacological activitiesCitation113,Citation115,Citation116 in addition to anti-carcinogenic properties in various cancer types.Citation117–Citation120 For instance, pyrogallol (1 mg/L) has been reported to exert anti-proliferative effect on MCF10DCIS cells.Citation56 However, pyrogallol at a higher dose (10 mg/L) was observed to have insignificant cytotoxicity against normal human breast epithelial cells MCF-12F, suggesting its selectivity of action towards cancer cells.Citation56 The same research group then studied the in vivo effect of pyrogallol, and found that treatment with pyrogallol (0.2 mg/day; 4 weeks) significantly reduced tumor volume (>70%) in mice bearing MCF10DCIS xenografts.Citation57
Methyl gallate (methyl-3,4,5-trihydroxybenzoate), a methyl ester of gallic acid, is naturally found in M. indica and various other plants.Citation121–Citation124 Methyl gallate possesses a wide range of medicinal properties.Citation122,Citation125–Citation131 For instance, methyl gallate (1 mg/L) was observed to induce a >40% reduction in MCF10DCIS cell proliferation.Citation56
Quercetin
Quercetin (3,5,7,3′,4′-pentahydroxyflavone) is a natural flavonoid that can be found abundantly in various fruits and vegetables.Citation132 For instance, there have been reports of the presence of quercetin in extracts prepared from M. indica leaves, peels and pulps.Citation27,Citation51,Citation54 Quercetin possesses a range of bioactive effects that can potentially be beneficial to human health,Citation133–Citation139 including its anti-cancer activities in a broad range of cancer types.Citation140–Citation146
In a study, Wilkinson et alCitation75 found that quercetin (100 µM) can significantly reduce MCF-7 cell viability. Another study by Zhang et alCitation147 reported that quercetin (25–100 µM) can inhibit MCF-7 cell growth in both dose- and time-dependent manner via the induction of apoptosis and S-phase arrest. Chou et alCitation148 similarly reported the ability of quercetin (10–175 µM) to dose- and time-dependently reduce MCF-7 cell viability via the induction of apoptosis and S-phase arrest. Subsequent analyses revealed increased protein levels of cyclic AMP-dependent transcription factor 6-alpha (ATF6), 78-kDa glucose-regulated protein (GRP78), protein kinase RNA-like endoplasmic reticulum kinase (PERK) and growth arrest and DNA damage 153 (GADD153) in quercetin-treated MCF-7 cells, all of which were suggested to be correlated with endoplasmic reticulum (ER) stress.Citation148 Furthermore, ΔΨm loss, increased protein levels of apoptosis-inducing factor (AIF), Bid, caspases 6, 8 and 9, Fas and tumor necrosis factor-related apoptosis inducing ligand (TRAIL), decreased protein levels of Bcl-2, poly-ADP ribose polymerase (PARP) and X-linked inhibitor of apoptosis protein (XIAP), increased activities of caspases 6, 8 and 9 as well as increased nuclear translocation of AIF and GADD153 were also observed, indicating the induction of apoptosis possibly in response to ER stress.Citation148 Overall, the findings also suggest that quercetin-induced apoptosis in MCF-7 cells is likely to be mediated through both mitochondrial and death receptor pathways.Citation148
In another breast cancer cell line (MDA-MB-231 cells), Chien et alCitation149 also observed a dose- and time-dependent reduction in cell viability following quercetin treatment (IC50: 278 µM). The observed reduction in cell viability can likely be explained by quercetin-induced apoptosis and G2/M-phase arrest,Citation149 which contrasted the findings by Zhang et alCitation147 and Chou et al Citation148 (ie, S-phase arrest in MCF-7 cells). Findings from the subsequent protein expression analysis of various ER stress and apoptotic markers in quercetin-treated MDA-MB-231 cells were similarly suggestive of the occurrence of ER stress-induced apoptosis and the involvement of two caspase-dependent apoptotic pathways (mitochondrial- and caspase 3-dependent pathways).Citation149 Another study also linked the anti-proliferative effect of quercetin (15 µM) on MDA-MB-231 cells to the induction of apoptosis and G2/M-phase arrest.Citation150 The same study also showed that quercetin exhibited anti-migratory activity against MDA-MB-231 cells. The researchers then evaluated the in vivo anti-cancer activity of quercetin (15 or 45 mg/kg body weight; thrice weekly for 13 weeks) in SCID mice bearing MDA-MB-231 xenografts, whereby both treatment groups demonstrated significantly reduced (~70%) tumor growth.Citation150
Overall Anti-Breast Cancer Mechanisms of Action of M. indica Extracts and Their Phytochemicals
Activation of Oxidative Stress-Induced Cell Death
Reactive oxygen species (ROS) are by-products of normal cellular metabolism.Citation151 They have physiological roles in signalling pathway (eg, apoptosis, carcinogenesis and proliferation) stimulation but are very transient due to their high reactivity.Citation152,Citation153 This maintenance of cellular redox balance is critical and is achieved via an antioxidant system.Citation154 Oxidative stress occurs when ROS production exceeds and depletes the antioxidant defences.Citation155 The balance of oxidative stress plays an important role in cancer. While moderately elevated ROS levels can contribute to carcinogenesis, cancer cell survival and metastasis, elevated ROS levels above the toxic threshold can cause cancer cell death usually via apoptosis induction.Citation156–Citation158 In recent years, oxidative stress-induced apoptosis has attracted attentions as a potential anti-cancer mechanism.Citation159
In a study conducted by Abdullah et alCitation160 ethanolic M. indica kernel extract was reported to induce both mitochondrial and death receptor pathways of apoptosis in MCF-7 cells via the induction of oxidative stress and the consequent upregulation of p53, as evidenced by the observations of dose- and time-dependent increases in ROS generation and malondialdehyde (MDA; a lipid peroxidation product) level as well as decrease in glutathione (GSH; an antioxidant) level in MCF-7 cells treated with the extracts (5–50 µg/mL). A similar study conducted by the same research group also linked the pro-apoptotic effect of ethanolic M. indica kernel extracts on MDA-MB-231 cells to oxidative stress induction and p53 upregulation.Citation161
As reported by Zhang et al,Citation147 the exposure of MCF-7 cells to quercetin (25–100 µM) dose-dependently caused the rightward-shift of dihydroethidium (DHE) signals and increased dichlorofluorescin diacetate (DCFH-DA) signals, which were suggestive of the stimulation of superoxide anion (O2−) release and the increased generation of hydrogen peroxide (H2O2), respectively. These findings indicate that quercetin is likely to induce apoptosis in MCF-7 cells by increasing oxidative stress.Citation147
Antioxidant Activity
Moderately elevated ROS level is considered to be pro-tumorigenic, as ROS function as signalling molecules in various pathways (eg, cell differentiation, proliferation, survival and migration) that can ultimately lead to the promotion of cancer initiation and progression.Citation162,Citation163 Depleting tumorous cells from these ROS-sensitive signalling pathways via the use of natural antioxidants can therefore potentially be a useful anti-cancer mechanism.Citation164
The leaves of M. indica are known to contain various families of phenolic compounds that possess potent antioxidant activity.Citation165 There have been reports that indicate the subcritical water, water, 50% ethanol:50% water, ethanol and 50% carbon dioxide:25% ethanol:25% water extracts of M. indica leaves exhibit a more potent antioxidant activity than (+)-α-tocopherol in the 2,2-diphenyl-1-picrylhydrazyl radical (DPPH) assay.Citation165,Citation166 A study also reported on the antioxidant activities of PET, CEW, PEW and FEW M. indica leaf extracts (0.01–100 µg/mL) in non-tumorous (MCF-10A) and tumorous (MCF-7 and MDA-MB-231) cells by detecting changes in intracellular ROS levels, subsequent to the observation of their cytotoxic effects on breast cancer cells.Citation27 However, the antioxidant activity was only observed at lower extract concentrations.Citation27 At the highest extract concentration (100 µg/mL), all extracts appeared to be pro-oxidant.Citation27 The cytotoxic effect observed at this high concentration was thus likely the outcome of oxidative stress-induced cell death.Citation27
Studies have also reported on antioxidant activities of M. indica peel and pulp extracts. For instance, aqueous M. indica pulp extract consistently demonstrated a high antioxidant activity in both DPPH and ferric reducing antioxidant power (FRAP) assays.Citation25 Another study confirmed the antioxidant activity of ethanolic M. indica peel extract in terms of its anti-lipid peroxidation, DPPH radical scavenging, ferric reducing and nitric oxide (NO) radical scavenging activities via the respective use of thiobarbituric acid reaction, DPPH assay, FRAP assay and Griess’ reagent.Citation51
Besides, the polyphenolic extracts (10 mg GAE/L) of different M. indica varieties have also demonstrated antioxidant potentials in the oxygen radical absorbance capacity (ORAC) assay, and a strong correlation between antioxidant activity and total phenolic content has been noted.Citation55 summarises how M. indica extracts and phytochemicals exert their anti-cancer effects by activating oxidative stress-induced cell death or acting as antioxidants.
Figure 2 Activation of oxidative stress-induced cell death and antioxidant activity as the anti-cancer mechanisms of M. indica extracts and phytochemicals.

Modulation of Peroxisome Proliferator-Activated Receptors
Peroxisome proliferator-activated receptors (PPARs), comprising three subtypes (PPARα, PPARβ/δ and PPARγ), are ligand-activated transcription factors belonging to the nuclear hormone receptor superfamily.Citation167 PPAR activation occurs in response to the binding of endogenous ligands, including Fas, prostacyclins, prostaglandins and triglycerides.Citation168 By binding the peroxisome proliferator responsive elements (PPREs) upstream of their target genes as a PPAR-retinoid X receptor (RXR) heterodimer, PPARs are able to activate the expressions of genes that are mostly involved in energy homeostasis and metabolic functions.Citation169
PPARs have also been implicated in carcinogenesis. Although PPARα has been linked to hepatocarcinogenesis in rodents, several epidemiological studies suggest that this is unlikely to be observed in humans.Citation170–Citation173 On the other hand, while some studies suggest that PPARδ may contribute to cancer progression,Citation174–Citation176 other studies suggest that PPARδ may contribute to cancer suppression.Citation177,Citation178 Similarly, although PPARγ activation has often been linked to proliferation inhibition and apoptosis induction in cancer cells,Citation179–Citation181 various studies have yielded contradictory results that suggest PPARγ activation may promote cancer development.Citation182,Citation183
In a study, Wilkinson et alCitation26 reported the ability of M. indica peel and pulp ethyl acetate fractions to reduce PPAR activity in Cos-7 cells. However, given the controversy surrounding the consequences of PPARγ activation in cancer, whether the PPARγ-inhibitory activity of M. indica peel and pulp fractions will be beneficial in breast cancer management requires further evaluation.Citation26
Suppression of Phosphoinositide 3-Kinase/Protein Kinase B Pathway
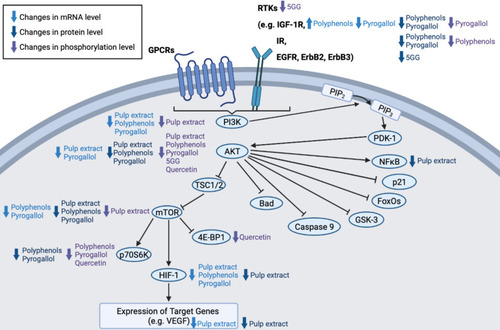
The phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) pathway can be stimulated in response to the activation of G-protein-coupled receptors (GPCRs) and receptor tyrosine kinases (RTKs) by cytokines, hormones and growth factors.Citation184 Upon GPCR or RTK activation, PI3K phosphorylates phosphatidylinositol 4, 5-bisphosphate (PIP2) to form phosphatidylinositol 3, 4, 4-triphosphate (PIP3) that can, in turn, recruit AKT and phosphoinositide-dependent kinase-1 (PDK-1) to the plasma membrane.Citation185 Following phosphorylation and activation by PDK-1, the activated AKT can then phosphorylate its downstream substrates, including Bcl-2-associated agonist of cell death (Bad), caspase 9, glycogen synthase kinase-3 (GSK-3), mammalian forkhead transcription factors of the O class (FoxOs), mTOR, NFκB and p21, which have an impact on cell cycle, growth, proliferation and glycometabolism.Citation184 Mutations and the consequent inappropriate activation of the PI3K/AKT pathway have been observed in many cancer types, contributing to genomic instability, metabolic reprogramming, uncontrolled cell growth and proliferation as well as multidrug resistance.Citation186–Citation188 In breast cancer, resistance to endocrine therapy, HER2-targeted therapy and cytotoxic chemotherapy has been reported.Citation189,Citation190 The downregulation of PI3K/AKT signalling may therefore offer benefits in breast cancer management.
M. indica pulp extract has demonstrated inhibitory effect on the PI3K/AKT pathway in both BT-474 cells and mice bearing BT-474 xenografts.Citation24 In BT-474 cells, M. indica pulp extract (2.5–10 mg GAE/L) lowered the messenger ribonucleic acid (mRNA) levels of PI3K, AKT, hypoxia-inducible factor-1 alpha (HIF-1α) and vascular endothelial growth factor (VEGF) as well as the protein levels of phosphorylated PI3K (pPI3K), AKT, phosphorylated AKT (pAKT), NFκB-p65 and VEGF.Citation24 In BT-474 xenografts, M. indica pulp extract (0.8 mg GAE/day; 35 days) downregulated the protein levels of pPI3K, pAKT, NFκB-p65, mTOR, phosphorylated mTOR (pmTOR), HIF-1α and VEGF.Citation24
The observed anti-proliferative activities of M. indica polyphenols (eg, pyrogallol) in breast cancer cells have also been linked to the downregulation of PI3K/AKT pathway.Citation56 Nemec et al Citation56 analysed the mRNA levels of various PI3K/AKT pathway components in MCF10DCIS cells treated with M. indica polyphenols or pyrogallol (10 mg/L). Both M. indica polyphenols and pyrogallol lowered the mRNA levels of PI3K, mTOR and HIF-1α.Citation56 However, while M. indica polyphenols increased the mRNA level of insulin-like growth factor-1 receptor (IGF-1R) and induced no significant change in the mRNA level of AKT, pyrogallol lowered the mRNA levels of both IGF-1R and AKT.Citation56 As measuring changes in the mRNA levels of pathway components provided limited indication on their respective protein activities, the authors also conducted protein expression analysis.Citation56 A reduction in the total protein levels of IGF-1R, insulin receptor (IR), AKT, mTOR and P70 S6 kinase (p70S6K; a kinase regulated by mTOR) as well as the phosphorylated protein levels of IR, AKT and p70S6K were observed in MCF10DCIS cells following M. indica polyphenol treatment.Citation56 On the other hand, pyrogallol reduced the total protein levels of IGF-1R, IR, insulin receptor substrate-1 (IRS-1), AKT, mTOR and p70S6K as well as the phosphorylated protein levels of IGF-1R, AKT and p70S6K.Citation56 The same research group similarly reported a suppression of the PI3K/AKT pathway by M. indica polyphenols (0.8 mg GAE/day; 4 weeks) and pyrogallol (0.2 mg/day; 4 weeks) in mice bearing MCF10DCIS xenografts, based on findings from mRNA and protein expression analyses.Citation57
A study by Hua et alCitation94 linked the growth-inhibitory effect of 5GG on MCF-7 cells to a suppression of the PI3K/AKT pathway. In the study, it was observed that while 5GG (10–40 µM) had insignificant impact on total AKT protein level, it dose- and time-dependently reduced the phosphorylation of AKT at serine 473 and directly inhibited the activity of AKT. Moreover, 5GG (20 µM) also inhibited epidermal growth factor- and/or estradiol-induced phosphorylation of RTKs upstream of the PI3K/AKT pathway and downregulated the protein levels of several RTKs (EGFR, ErbB2 (or HER2) and ErbB3).Citation94 In particular, 5GG-induced ErbB2 depletion was blocked by pre-treatment with chloroquine (CQ; a lysosomal inhibitor) but not carbobenzoxy-L-leucyl-L-leucyl-L-leucinal (MG132; a proteasome inhibitor), suggesting 5GG was likely to deplete ErbB2 by promoting its lysosomal degradation.Citation94 Overall, 5GG can suppress PI3K/AKT signalling by inhibiting AKT and RTK phosphorylation as well as depleting RTKs.Citation94
According to Rivera Rivera et al,Citation150 quercetin (1–15 µM) also exhibits inhibitory activity against the PI3K-AKT pathway, as it is capable of inhibiting the phosphorylation of AKT at serine 473 and the phosphorylation of the downstream effectors of mTOR [p70S6K and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1)] in MDA-MB-231 cells. summarises changes in the mRNA, total protein and total phosphorylated protein levels of various PI3K/AKT pathway components induced by M. indica extracts and phytochemicals.
Figure 3 Suppression of PI3K/AKT/mTOR pathway by M. indica extracts and phytochemicals via reduction of pathway components’ mRNA, total protein and phosphorylated protein levels.
Induction of MicroRNA-126 (miR-126) Expression
MicroRNAs (miRNAs) are short non-coding RNAs implicated in the post-transcriptional regulation of eukaryotic gene expression.Citation191 miRNAs bind to the 3’-untranslated regions of their target mRNAs and, depending on the degree of miRNA-mRNA complementarity, can either result in mRNA degradation or translation inhibition.Citation192 miRNAs are involved in various cellular processes such as cell development, differentiation, proliferation and apoptosis.Citation192,Citation193 A reduction in miR-126 expression has frequently been observed in breast cancer, and various studies have reported that a restoration of miR-126 expression can suppress breast cancer cell growth, metastasis and invasion; suggesting miR-126 functions as a tumor suppressor in breast cancer.Citation194,Citation195 Therefore, the induction of miR-126 expression can be beneficial in treating breast cancer.
A study showed that M. indica pulp extract can induce miR-126 expression in BT-474 cells (2.5–10.0 mg GAE/L) and BT-474 xenografts in mice (0.8 mg GAE/day; 35 days).Citation24 In the same study, the miRNA expression profile screening of extract-treated xenografts further revealed significant changes in the expression of several miRNAs that have a major role in the regulation of tumor cell growth and proliferation. These changes are potentially linked to the observed anti-proliferative effect of M. indica pulp extract.Citation24
Inhibition of Aromatase Enzymatic Activity and Expression
Approximately 70% of all breast cancer cases are ER-positive, where ER activation by estrogen plays a key role in the stimulation of breast cancer growth and progression.Citation196 The use of endocrine therapy, either selective estrogen-receptor modulators (SERMs) or aromatase inhibitors (AIs), as an adjuvant treatment has demonstrated improvement in the disease-free survival of ER-positive breast cancer patients.Citation197 However, the risk of side effects associated with long-term AI use has triggered research into the discovery of a natural product-based new generation of AIs.Citation51
Aromatase is a member of the cytochrome P450 superfamily of enzymes that catalyses the conversion of testosterone to estrogen in the final step of estrogen biosynthesis.Citation198 Aromatase expression is tissue-specific and dependent upon 10 alternative untranslated exons I.Citation199 Aromatase expression in normal breast adipose tissues is maintained at a low level, driven primarily by promoter I.4 and minimally by promoters I.3 and PII.Citation200 In estrogen-dependent breast cancer, however, elevated aromatase expression has frequently been observed to be driven by mainly promoters I.3, I.4, I.7 and PII.Citation200,Citation201
In a study, ethanolic M. indica peel extract was reported to dose-dependently lower the activity of human placental aromatase, with IC50 at 86.09 µg/mL.Citation51 The same study also reported that ethanolic M. indica peel extract (33 or 66 µg/mL) can significantly reduce aromatase mRNA level in MCF-7 cells. The extract at 33 µg/mL caused a significant reduction in I.3-containing mRNA level (78%), a significant increase in I.6-containing mRNA level (1478%), a non-significant increase in I.7- and I.f-containing mRNA levels (294% and 212%) and non-significant change in PII-containing mRNA level.Citation51 The extract at 66 µg/mL, on the other hand, caused a significant reduction in I.3-, I.7- and PII-containing mRNA levels (94%, 89.7% and 82%), a significant increase in I.6-containing mRNA level (500%) and a non-significant increase in I.f-containing mRNA level (100%).Citation51 These observations collectively suggest the potential of M. indica peel extract to act as a tissue-specific AI in ER-positive breast cancer.Citation51
Modulation of Intracellular Calcium Signalling
The calcium (Ca2+) signalling is implicated in multiple cellular processes such as autophagy,Citation202 cell cycle,Citation203 death,Citation204 differentiation,Citation205 division,Citation206 invasion,Citation207 metabolism,Citation208 migrationCitation209 and transcription.Citation210 The key to Ca2+ signalling is the differential distribution of Ca2+ concentrations across cell membranes, between extracellular space and cytoplasm as well as between cytoplasm and organelles such as ER and Golgi apparatus.Citation211 Movement of Ca2+ ions down their electrochemical gradients produces Ca2+ signals.Citation211 This can occur with electrical, hormonal or mechanical stimulation of cells, which can activate either GPCRs that typically trigger Ca2+ mobilisation from intracellular stores via lipid signal transduction; or ion channels that allow Ca2+ movement through central pore opening.Citation212 The resulting increase in cytoplasmic Ca2+ concentration and Ca2+-binding of calcium-binding proteins (CBPs) ultimately leads to the initiation of cellular functions.Citation213
The dysregulated expression of a number of Ca2+ channels and pumps has been reported in either breast cancer tissues or breast cancer cell lines, whereby they result in the remodelling of Ca2+ signalling.Citation214 There have been evidences linking altered Ca2+ signalling to various malignant functions such as cell proliferation, migration, invasion and metastasis in breast cancer.Citation211 Ca2+ channels and pumps, therefore, may be potentially useful anti-cancer targets in breast cancer treatment.
Taing et alCitation53 evaluated the effects of methanolic M. indica peel and pulp extracts (30 or 60 µg/mL) on Ca2+ signalling in MCF-7 cells. In untreated MCF-7 cells, adenosine triphosphate (ATP) stimulation resulted in a rapid increase in intracellular Ca2+ concentration [Ca2+]I, which was followed by a recovery to basal [Ca2+]I.Citation53 In extract-treated MCF-7 cells, however, significant reduction in peak relative [Ca2+]I and delay in [Ca2+]I recovery were noted.Citation53 Further studies are thus required to ascertain the target(s) upon which the extracts are acting to cause the observed changes in Ca2+ signalling, and to assess if the extracts have an influence on any Ca2+-dependent signalling pathways.Citation53
Studies by Chien et alCitation149 and Chou et al)Citation148 also showed that quercetin (250 or 150 µM) can elevate cytosolic Ca2+ level in MDA-MB-231 and MCF-7 cells, respectively. These findings suggest that further studies would be required to evaluate the influence of quercetin on Ca2+-dependent signalling pathways and the link between this influence and quercetin’s anti-breast cancer activity.Citation148,Citation149
Inhibition of Nuclear Factor Kappa B Signalling Pathway
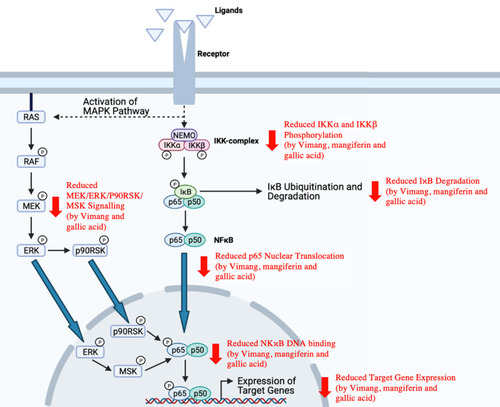
Nuclear factor kappa B (NFκB), a pro-inflammatory transcription factor, is expressed in virtually all cell types.Citation215 The NFκB family of transcription factors comprises five members, including c-Rel, NFκB1 (p50), NFκB2 (p52), RelA (p65) and RelB.Citation216 Different combinations of these members make up homodimeric or heterodimeric NFκB, with the most common combination being a p65/p50 heterodimer.Citation216,Citation217 NFκB dimers are usually complexed with the inhibitor of NFκB (IκB), which retains NFκB dimers in the cytoplasm and thereby prevents NFκB dimers from binding DNA.Citation218 NFκB activation can occur via two major signalling pathways (ie, classical/canonical and alternative/non-canonical pathways).Citation219 The eventual phosphorylation and ubiquitination of IκB within their signal responsive domain (SRD) results in signal responsive degradation, thereby releasing NFκB from IκB and enabling NFκB to bind to κB sites within the promoters of their target genes.Citation218
Alterations of NFκB pathway are often observed in solid and hematopoietic malignancies.Citation220 For instance, abnormal, constitutive NFκB activation is frequently seen in breast cancer, and it is believed to contribute to breast cancer development and progression by promoting angiogenesis, cell survival, proliferation, metastasis as well as resistance to chemotherapy, endocrine therapy and radiotherapy.Citation216,Citation221–Citation225 NFκB pathway has thus been considered as a potential target for breast cancer therapy.
A study by García-Rivera et alCitation34 reported that Vimang (200 µg/mL), mangiferin (100 µg/mL) and gallic acid (10 µg/mL) can inhibit the classical NFκB activation pathway in MDA-MB-231 cells by inhibiting the phosphorylation of IκB kinase alpha (IKKα; at serines 176 and 180) and IκB kinase beta (IKKβ; at serines 177 and 181), which can consequently result in reduced IκB degradation, p65 nuclear translocation and NFκB DNA binding. The optimal activation of NFκB also involves the phosphorylation of its functional domains by various protein kinases.Citation226,Citation227 In the same study, Vimang and gallic acid were observed to inhibit the MEK/ERK/p90RSK/MSK signalling cascade while mangiferin appeared to have insignificant effect.Citation34 Since gallic acid can inhibit both classical and parallel NFκB activation pathways while mangiferin can inhibit only the classical NFκB activation pathway, gallic acid is likely to be a more potent NFκB inhibitor.Citation34 As a consequence of NFκB pathway inhibition, there was also a significant reduction in the expression of NFκB target genes, including interleukin (IL)-6, IL-8, VEGF, Bcl-2, XIAP, COX-2 and C-X-C chemokine receptor type 4 (CXCR4), in MDA-MB-231 cells treated with Vimang, mangiferin or gallic acid.Citation34
Consistent with the findings of García-Rivera et al,Citation34 a previous study conducted by Sarkar et alCitation228 also reported on the inhibitory effect of mangiferin on NFκB activation. In the study, mangiferin (0.5–10 µg/mL) suppressed TNF-stimulated activation of NFκB in MCF-7 cells. illustrates various processes along the NFκB signalling pathway that have been shown to be suppressed by M. indica extracts and phytochemicals.
Figure 4 Inhibition of classical and parallel NFκB activation pathways by M. indica extracts and phytochemicals.
Inactivation of β-Catenin Pathway
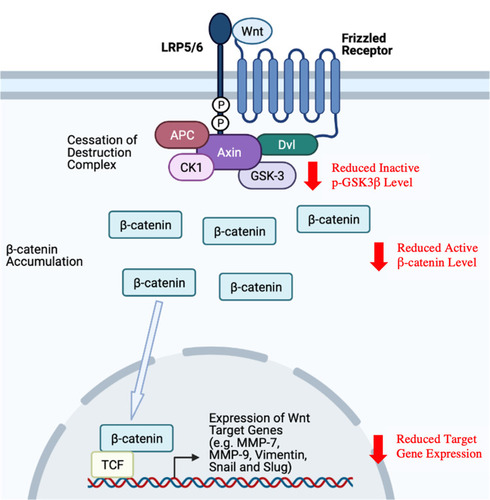
Wnts, a family of 19 secreted glycoproteins, are chief regulators of β-catenin activity.Citation229 In the absence of Wnt signal, β-catenin is constantly maintained at a low cytoplasmic level via the action of a destruction complex consisting of Axin, adenomatous polyposis coli protein (APC), casein kinase 1 (CK1) and GSK-3, where β-catenin phosphorylation by CK1 and GSK-3 eventually leads to its ubiquitination and proteasomal degradation.Citation230 Activation of the Wnt/β-catenin pathway is initiated with the binding of Wnt to the Frizzled (Fz) receptor and its co-receptor [ie, low-density lipoprotein receptor-related protein 5/6 (LRP5/6)].Citation231 The subsequent recruitment of Dishevelled (Dvl) protein, the phosphorylation and activation of LRP5/6 as well as the recruitment of destruction complex eventually ceases β-catenin degradation, thereby leading to β-catenin accumulation and nuclear translocation.Citation231 The expressions of Wnt target genes are usually repressed by T cell-factor/transducin-like enhancer of split (TCF/TLE) proteins and histone deacetylase (HDAC).Citation232 As β-catenin binds to lymphoid enhancer factor (LEF)/TCF family of transcription factors to form a binding complex, it can act as a co-activator for LEF/TCF proteins to activate the expression of Wnt target genes involved in various biological processes.Citation232,Citation233
Dysregulation of the Wnt/β-catenin signalling pathway has been associated with different diseases, including human cancers.Citation234 Various studies have demonstrated that aberrant activation of the Wnt/β-catenin signalling pathway can promote angiogenesis, cell proliferation, migration, invasion and chemoresistance, thereby contributing to cancer initiation and progression.Citation235–Citation237 For example, constitutively active Wnt/β-catenin pathway has been linked to metastasis in breast cancer.Citation234,Citation238
Mangiferin (12.5–50 µM) has been reported to downregulate matrix metalloproteinases (MMP)-7 and −9 and reverse epithelial-mesenchymal transition (EMT) in MDA-MB-231 and BT-549 cells, as evidenced by the observations of morphological changes (from fibroblast-like to cobblestone-like), significantly increased levels of epithelial marker (E-cadherin) and significantly decreased levels of mesenchymal markers (vimentin, snail and slug) following treatment.Citation66 Similar in vivo effects have been observed in MDA-MB-231 xenografts of mice subjected to mangiferin treatment (100 mg/kg/day; 5 weeks).Citation66 The same study also suggested that mangiferin can inhibit the β-catenin pathway by lowering the protein levels of inactive phospho-GSK3β (p-GSK3β) and active β-catenin both in vitro and in vivo. Furthermore, while β-catenin overexpression reversed the anti-proliferative, anti-migratory and anti-invasive effects of mangiferin in MDA-MB-231 and BT-549 cells, β-catenin silencing downregulated MMP-7, MMP-9 and snail, indicating that the inactivation of β-catenin pathway may contribute to the anti-cancer effects of mangiferin by modulating MMP-7, MMP-9 and EMT.Citation66 The overall study findings by Li et alCitation66 on mangiferin-mediated inactivation of β-catenin pathway are as summarised in .
Figure 5 Inactivation of β-catenin pathway by mangiferin.
Inhibition of 3-Hydroxy-3-Methyl-Glutaryl-Coenzyme A Reductase, Proteasome and Plasmin Enzymatic Activities
3-Hydroxy-3-methyl-glutaryl-coenzyme A (HMG-CoA) reductase is an enzyme that catalyses the rate-limiting step (ie, conversion of HMG-CoA to mevalonate) of cholesterol biosynthesis.Citation239 Low-density lipoprotein (LDL) cholesterol signalling has been associated with the promotion of cell proliferation, migration and invasion in breast cancer, suggesting the control of cholesterol level may be beneficial.Citation240,Citation241 The statins, a class of cholesterol-lowering drugs, exert their hypocholesterolemic effects by blocking the activity of HMG-CoA reductase.Citation242 There have been evidences suggesting that the inhibition of HMG-CoA reductase by statins can impair breast cancer cell proliferation, making statins potentially useful in breast cancer treatment.Citation243 Cuccioloni et al Citation68 found that mangiferin at low µM concentrations (ie, inhibition constant of 3±0.2 µM) can similarly reduce the activity of microsomal HMG-CoA reductase. As expected, this translated to a significant dose-dependent reduction in the cytoplasmic cholesterol levels of MCF-7 and MCF-10A cells following exposure to mangiferin.Citation68 These effects of mangiferin on cholesterol homeostasis can, thus, potentially explain its observed anti-cancer activity in breast cancer cells.Citation68
The ubiquitin-proteasome pathway (UPP) is the major pathway responsible for degrading more than 80–90% of the intracellular proteins in mammalian cells.Citation244 Importantly, UPP is also known to play an important role in the regulation of cellular processes involved in both tumor cell growth and survival.Citation245 Studies with proteasome inhibitors have demonstrated anti-proliferative and pro-apoptotic activities in tumor cells, thereby suggesting the potential usefulness of proteasome inhibitors as anti-cancer agents.Citation246–Citation248 When Cuccioloni et al Citation68 evaluated the effect of mangiferin (1–20 µM) on various catalytic activities of isolated 20S proteasome, a significant dose-dependent reduction was observed for chymotrypsin-like (ChT-L), trypsin-like (T-L), peptidylglutamyl peptide hydrolase (PGPH) and branched-chain amino acid preferring (BrAAP) activities, with T-L activity being the most sensitive to inhibition by 20 µM mangiferin. When proteasome activities in the lysates of mangiferin-treated MCF-7 and MCF-10A cells were measured, significantly reduced ChT-L, T-L, PGPH and BrAAP activities were similarly observed.Citation68
Plasminogen, a single-chain glycoprotein, is usually found in the blood circulation in its inactive form.Citation249 Upon association with cellular receptors, the resulting conformational changes induced in plasminogen promote the catalysis of its activation to plasmin by either urokinase plasminogen activator (uPA) or tissue plasminogen activator (tPA).Citation250 Plasmin is a serine protease that has a primary role in fibrinolysis.Citation251 However, the ability of plasmin to catalyse the degradation of ECM proteins and cell–cell adhesion proteins has also associated plasmin with cellular migration in both physiological (eg, embryonic angiogenesis and wound healing) and pathological (eg, tumor growth and metastasis) conditions.Citation252–Citation254 This provides a rationale for plasmin inhibition in cancer management. In a study, Cuccioloni et alCitation68 observed that mangiferin (0–200 µM) exhibited a moderate inhibitory potency against isolated plasmin. As expected, mangiferin was also reported to cause a significant dose-dependent reduction in the cleavage of E-cadherin by plasmin in both MCF-7 and MCF-10A cells.Citation68 The plasmin inhibitory effect of mangiferin can thus, at least partly, help to explain its observed anti-migratory activity in breast cancer cells.Citation68
Inhibition of Rac1/WAVE2 Signalling Pathway
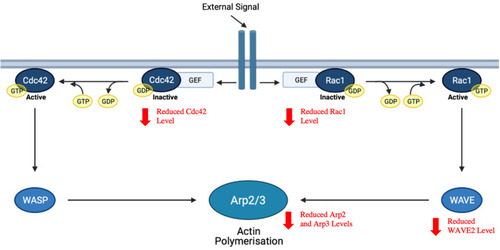
Rho-family of small GTPases play an important role in the regulation of a range of cellular processes, including cell apoptosis, motility, mitosis and proliferation.Citation255 Cell division control protein 42 homolog (Cdc42) and Ras-related C3 botulinum toxin substrate 1 (Rac1), two members of the Rho-family of small GTPases, have specifically been associated with the activation of actin polymerisation and thereby, the modulation of cellular cytoskeletal reorganisation.Citation256 Wiskott–Aldrich Syndrome protein (WASP) and WASP-family verprolin-homologous protein (WAVE), the respective downstream effectors of Cdc42 and Rac1, are capable of activating the actin-related protein 2/3 (Arp2/3) complex that is the major cellular machinery for actin nucleation.Citation257 Activation of the Arp2/3 complex eventually results in the remodelling of actin filament networks as well as the formation of membrane protrusions and ruffles required for cellular motility in tumor cells.Citation256,Citation258 Despite the existence of other members of the WAVE family of proteins, it has been shown that WAVE2 tends to be the most important in mediating breast cancer cell motility.Citation259 As such, inhibiting actin polymerisation via the suppression of Rac1/WAVE2 signalling pathway may be effective in preventing metastasis.
It was reported by Deng et alCitation67 that mangiferin (2.5–10 µM) can induce a significant dose-dependent reduction in the protein levels of Rac1/Cdc42, phospho-Rac1/Cdc42 (pRac1/Cdc42), WAVE2, Arp2 and Arp3 in MDA-MB-231 cells, which is indicative of a suppression of the Rac1-WAVE2 signalling pathway (). The authors linked this suppressive effect of mangiferin to its observed anti-migratory and anti-invasive activities in breast cancer cells.
Figure 6 Inhibition of Rac1/WAVE2 signalling pathway by mangiferin. Arp2/3, activating the actin-related protein 2/3; Cdc42, cell division control protein 42 homolog; GEF, guanine nucleotide exchange factor; Rac1, Ras-related C3 botulinum toxin substrate 1; WASP, Wiskott–Aldrich syndrome protein; WAVE, WASP-family verprolin-homologous protein.
Modulation of Estrogen Receptor Activity
ERs belong to the steroid hormone superfamily of nuclear receptors.Citation260 Upon activation by estrogen binding, the subsequent ER-mediated signalling can occur in either a genomic or non-genomic manner.Citation261 Direct genomic signalling, the classical estrogen signalling mechanism, involves conformational change and dimerization of ER, nuclear translocation of ER complex and their subsequent interaction with chromatin in estrogen response elements (EREs) to directly regulate gene expression.Citation262 In contrast, non-genomic signalling is mediated by membrane-bound ER and can indirectly regulate gene expression via the activation of various intracellular signalling cascades.Citation263
There are two structurally and functionally distinct ER subtypes (ie, ERα and ERβ), both of which are expressed in normal breast tissues.Citation264 In breast cancer, while ERα expression is frequently found to be upregulated and is clearly associated with both prognosis and endocrine treatment response, ERβ expression is usually observed to be downregulated and has no clear association with any clinical parameters.Citation264 ERα and ERβ have been known to play a different role in tumorigenesis, where ERα is proliferative while ERβ appears to be anti-proliferative.Citation265 However, it has been reported that ERβ can oppose the proliferative activity of ERα in co-expression, possibly via the formation of ERα/β heterodimers.Citation266 Moreover, ERα/β heterodimer-selective ligands have demonstrated growth-inhibitory effect on tumor cells and thus can potentially be useful in the treatment of ERα- and ERβ-positive breast cancer.Citation266
To elucidate the mechanisms underlying the observed anti-breast cancer activities of M. indica phytochemicals, a study examined the effects of quercetin, mangiferin and norathyriol (1–1000 µM) on the ERE transactivation activities of ERα and ERβ in Cos-7 cells.Citation75 While it was reported that all three M. indica phytochemicals are capable of activating ERα, only norathyriol is capable of activating ERβ.Citation75 Therefore, further studies are required to investigate the link of these differential ER-modulatory effects to the observed lack of anti-proliferative effect of mangiferin as well as the observed anti-proliferative effects of quercetin and norathyriol on MCF-7 cells; and to evaluate the effects of these M. indica phytochemicals on ERα/β heterodimer activity.Citation75
5GG, another M. indica phytochemical, has also been reported to exert growth-inhibitory effect on MCF-7 by inhibiting ERα function.Citation94 Specifically, the study revealed that 5GG (1–40 µM) can inhibit ERα activity by promoting ERα degradation through the lysosomal pathway or blocking all known pathways of ERα activation – estrogen-mediated activation, growth factor-induced phosphorylation of the amino-terminal activation function-1 (AF-1) and cyclin D1-mediated activation.Citation94
Activation of AMP-Activated Protein Kinase Signalling Pathway
AMP-activated protein kinase (AMPK) is a serine/threonine kinase complex consisting of a catalytic subunit (AMPK-α) and two regulatory subunits (AMPK-β and AMPK-γ).Citation267 It is regarded as a sensor of intracellular adenosine nucleotide levels, where it is activated in response to cellular stresses that can reduce intracellular ATP level.Citation268 AMPK activation involves the phosphorylation of its conserved threonine 172 residue by an upstream kinase such as Ca2+/calmodulin-dependent protein kinase β (CaMKKβ), liver kinase B1 (LKB1) and transforming growth factor β activated kinase 1 (TAK1).Citation269 More recent studies have further suggested AMPK’s role as an oxidative stress sensor in addition to its traditional role as an energy sensor, where it can be activated in response to increased intracellular ROS/reactive nitrogen species (RNS) levels.Citation270–Citation272
AMPK has physiological roles in the regulation of cell growth, metabolism, polarity and autophagy, and AMPK activation has been recognised to exert beneficial effects in various pathological conditions.Citation268 The downregulation of AMPK has been associated with poor outcomes in breast cancer patients.Citation273 However, there have been reports that suggest AMPK activation can exert a growth-inhibitory effect on breast cancer cells, thus making AMPK signalling pathway a promising target in breast cancer treatment.Citation274–Citation276
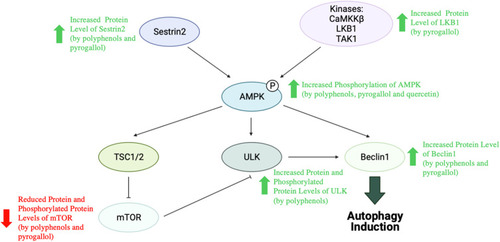
A study proposed that the observed inhibitory effects of M. indica polyphenols (0.8 mg GAE/day; 4 weeks) and pyrogallol (0.2 mg/day; 4 weeks) on MCF10DCIS xenograft tumor growth can be attributed at least partly to AMPK pathway activation and AKT/mTOR pathway suppression.Citation57 In the study, mice of both treatment groups showed elevated levels of Sestrin2, phosphorylated AMPK and Beclin1 as well as reduced levels of mTOR and pmTOR. However, while elevated level of LKB1 was only seen in pyrogallol-treated mice, elevated levels of Unc-51-like kinase (ULK) and phosphorylated ULK were only seen in polyphenol-treated mice.Citation57 Sestrin2, a highly conserved stress-inducible protein, is known to be capable of activating AMPK that can in turn inhibit mTOR.Citation277 In addition, both ULK and Beclin1, which are involved in the induction of autophagic process, are also the downstream targets of AMPK.Citation278 Subsequent in vitro assays performed on MCF10DCIS cells linked AMPK pathway activation induced by M. indica polyphenols and pyrogallol (1–20 mg/L) to their abilities to increase ROS generation.Citation57 Moreover, it was reported that pyrogallol can also activate the AMPK pathway by binding directly to the allosteric site of AMPK or upregulating AMPK expression.Citation57
On the other hand, quercetin’s anti-cancer activity has been linked to an upregulation of AMPK signalling in another study, whereby quercetin (15 µM) was observed to significantly increase AMPK phosphorylation (by twofold) in MDA-MB-231 cells.Citation150 summarises the study observations suggesting an upregulation of the AMPK signalling pathway following treatment with M. indica extracts and their phytochemicals.
Figure 7 Activation of AMPK signalling pathway by M. indica extracts and phytochemicals.
Modulation of Cell Cycle Regulators
Cell cycle is a process comprising four major phases (ie, G1-, S-, G2- and M-phases), during which the doubling of cellular components and their accurate segregation into two daughter cells occur.Citation279 Transition through each phase of the cell cycle is under tight regulation by specific proteins.Citation279 Cyclin-dependent kinases (CDKs), a family of serine/threonine protein kinases, are the key regulatory proteins that control cell cycle progression when complexed with their regulatory subunits (ie, cyclins).Citation280 G1-to-S-phase transition involves the activation of CDK4, 6 activity by D-type cyclin binding.Citation281 One target of cyclin D-CDK4, 6 activity is the retinoblastoma tumor suppressor protein (Rb), which will dissociate from E2F transcription factors following phosphorylation to enable E2F-induced expression of cyclins E, A and thymidylate synthase, etc.Citation282,Citation283 Continued phosphorylation and complete inactivation of Rb in the late G1-phase is achieved by the complexes formed from cyclins E and A as well as CDK1 and 2.Citation284 On the other hand, cyclinB-CDK1 activity is known to drive cellular events associated with mitosis, thereby facilitating G2-to-M-phase transition.Citation285 The activity of cyclin-CDK complexes can be counteracted by two gene families of CDK inhibitors (CKIs), thereby resulting in cell cycle arrest.Citation286 While INK4 gene family members (p16INK4a, p15INK4b, p18INK4c and p19INK4d) are capable of inhibiting CDK4, 6 activity, Cip/Kip gene family members (p21Cip1/Waf1/Sdi1, p27Kip1 and p57Kip2) can interfere with the activities of cyclins D-, E-, and A-CDK complexes.Citation286
Deregulation of cell cycle, which confers cells with the ability to undergo cell division endlessly, is one of the hallmarks of cancer.Citation287 In breast cancer, deregulation of cell cycle often results from an aberrant activation of CDKs and cyclins as well as an inactivation of tumor suppressors.Citation288 As such, several CDK4/6 inhibitors (ie, abemaciclib, palbociclib and ribociclib) have been approved by FDA for the treatment of breast cancer, and a number of cell cycle inhibitors (eg, TTK protein kinase [TTK] and polo-like kinase 4 [PLK4] inhibitors) are currently under development.Citation287
Chen et alCitation95 reported the ability of 5GG (50 µM) to induce a G1-phase arrest in MCF-7 cells and proceeded to investigate the underlying mechanisms of this arrest. Overall study findings suggested that 5GG treatment can increase the protein level of p27Kip, which can in turn bind to and inhibit CDK2 and 4 kinase activities, resulting in a reduced Rb phosphorylation and thus preventing E2F-induced expression of S-phase genes.Citation95 Moreover, it was also reported that 5GG may block the association of p21Cip with cyclin D-CDK4 complexes, which would usually be required to increase the stability and activity of cyclin D-CDK4 complexes implicated in G1-to-S-phase transition.Citation95
Gallic acid (2–12 µg/mL) has also been reported to induce S- and G2/M-phase arrest in MCF-7 cells, which is linked to its ability to dose-dependently lower the protein levels of cyclin A, cyclin B1, cdc2/CDK1 and CDK2.Citation111 Additionally, gallic acid can also dose- and time-dependently downregulate S-phase kinase-associated protein 2 (Skp2) and prevent it from ubiquitinating p21Cip1 and p27Kip1, thereby preventing the proteasomal degradation of p21Cip1 and p27Kip1 and leading to cell cycle arrest.Citation111 However, as the silencing of p27Kip1 but not p21Cip1 can significantly diminish gallic acid-induced increase in G2/M-phase ratio, p27Kip1 may play a more important role than p21Cip1 in the observed cell cycle arrest induced by gallic acid.Citation111
Quercetin can also induce cell cycle arrest in breast cancer cell lines. For instance, Chien et alCitation149 observed a G2/M-phase arrest in MDA-MB-231 cells following quercetin treatment (200 µM), and correlated this with a reduction in the protein levels of cyclin A, cyclin B and thymidylate synthase as well as an increase in the protein levels of p53 and p57. On the other hand, Chou et alCitation148 reported that quercetin (150 µM) can induce a S-phase arrest in MCF-7 cells by downregulating the protein levels of CDK2, cyclin A, cyclin E and thymidylate synthase as well as upregulating the protein levels of p53 and p57.
Conclusion and Future Perspectives
This review provides a comprehensive summary of the studies on the anti-cancer effects of extracts prepared from various parts of M. indica (ie, bark, kernel, leaves, peels and pulps) and its polyphenolic phytochemicals (ie, mangiferin, norathyriol, gallotannins, gallic acid, pyrogallol, methyl gallate and quercetin) in preclinical models of breast cancer. Overall, there was a reduction in cell viability, proliferation, growth, migration and invasion as well as an induction of apoptosis and cell cycle arrest. Reduced tumor weight and volume were also observed. A number of mechanisms have been proposed so far: activation of oxidative stress-induced cell death, antioxidant activity, modulation of PPARs, suppression of P13K/AKT pathway, induction of miR-126 expression, inhibition of aromatase enzymatic activity and expression, modulation of intracellular Ca2+ signalling, inhibition of NFκB signalling pathway, inactivation of β-catenin pathway, inhibition of HMG-CoA reductase, proteasome and plasmin enzymatic activities, inhibition of Rac1/WAVE2 signalling pathway, modulation of ER activity, activation of AMPK signalling pathway as well as modulation of cell cycle regulators.
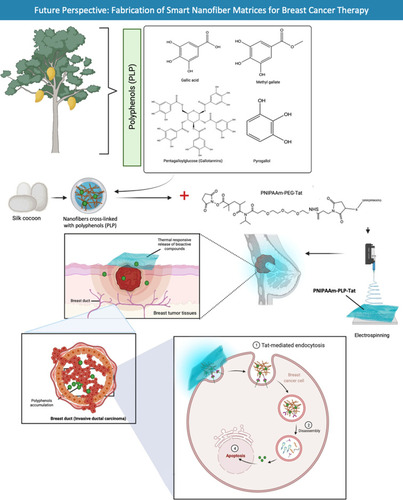
Further selective compound studies should be performed on M. indica extracts that possess anti-breast cancer activities to identify and validate phytochemicals that are responsible for the observed bioactivities. Extracts of different M. indica parts showed different bioactivities, suggesting variabilities in the type and amount of phytochemicals and thus highlighting the importance of targeting certain plant parts. Additionally, variabilities in bioactivities have also been observed for M. indica extracts despite utilisation of the same plant parts, indicating the presence of different phytochemical profiles in these extracts and further emphasizing the importance of selecting the right M. indica varieties, extraction method and extraction solvent. Once identified and validated, the phytochemical(s) of interest should be isolated for subsequent studies on their efficacies, toxicities, mechanisms of action and drug metabolism and pharmacokinetics (DMPK) profiles. It has been supported by various studies that multiple M. indica phytochemicals may act synergistically such that their combined anti-breast cancer efficacy is higher than that of a single phytochemical, suggesting the potential benefits of their combined usage in cancer treatment. Furthermore, while M. indica fruit consumption may be beneficial for women’s health in relation to breast cancer given the reported anti-breast cancer activities of M. indica pulp extracts, further bioavailability studies are required to determine whether the bioactive compounds within the fruits exhibit reasonable bioavailability to exert their anti-cancer effects following fruit consumption. More committed studies, particularly clinical trials, are needed to explore the anti-breast cancer potential of M. indica extracts and their phytochemicals in order to validate their utility in the development of unique and potent therapeutic agents to treat breast cancer. Apart from that, the phytochemicals that are present in M. indica extracts may vary in concentration depending on the food source and season, hence the development of health supplements is a better option for consistent deliveries. Furthermore, chemotherapeutic agents lack tumor selectivity such that they may damage normal cells and cause major side effects. Therefore, targeted delivery of bioactive compounds to tumors is required to minimise toxicity to normal cells. Drug delivery techniques based on nanofibers have recently emerged as a potential platform for delivering bioactive compounds to tumor sites. Nanofibers cross-linked with polyphenols from M. indica can be coupled to PNIPAAm-PEG-Tat to enable smart tumor microenvironment-responsive targeted distribution of M. indica polyphenols. Because of their ability to deliver medicines and genes intracellularly, arginine-rich cell-penetrating peptides (CPPs) like transactivating transcriptional activator (TAT) have attracted a lot of attention (). On the other hand, in-depth study into alternative delivery systems should be examined as a possibility for optimizing localisation at tumor target areas and therefore boosting therapeutic efficacy.
Figure 8 Fabrication of smart nanofiber matrices for breast cancer therapy. Polyphenolic compounds (PLP) from M. indica have been studied extensively as anti-inflammatory and anti-cancer agents. In the proposed future study, M. indica PLP will be cross-linked to silk fibroin (cocoon) nanofibers before being conjugated with PNIPAAm-PEG-Tat to yield PNIPAAm-PLP-Tat, which can then be released in a thermal responsive burst at breast tumor tissues. The use of cell-penetrating peptide (Tat) permits targeted intracellular transport of PLP into breast cancer cells.
Consent for Publication
The final version of the manuscript was reviewed by all the authors who consented to its submission.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Acknowledgments
The authors appreciate Universiti Kuala Lumpur Royal College of Medicine Perak, Ipoh, Perak, Malaysia, for providing the facilities and services required to complete the study. Figures were created with the support of https://biorender.com under the paid subscription.
Disclosure
The authors have no conflict of interest associated with the publication. There is also no significant financial support for this work.
References
- SiegelRL, MillerKD, JemalA. Cancer statistics, 2016. CA Cancer J Clin. 2016;66(1):7–30. doi:10.3322/caac.2133226742998
- SungH, FerlayJ, SiegelRL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.2166033538338
- KenneckeH, YerushalmiR, WoodsR, et al. Metastatic behavior of breast cancer subtypes. J Clin Oncol. 2010;28(20):3271–3277. doi:10.1200/JCO.2009.25.982020498394
- SunYS, ZhaoZ, YangZN, et al. Risk factors and preventions of breast cancer. Int J Biol Sci. 2017;13(11):1387–1397. doi:10.7150/ijbs.2163529209143
- Abu-HelalahM, AzabB, MubaidinR, et al. BRCA1 and BRCA2 genes mutations among high risk breast cancer patients in Jordan. Sci Rep. 2020;10(1):17573. doi:10.1038/s41598-020-74250-233067490
- LarsenMJ, ThomassenM, GerdesAM, KruseTA. Hereditary breast cancer: clinical, pathological and molecular characteristics. Breast Cancer (Auckl). 2014;8:145–155.25368521
- KorenS, Bentires-AljM. Breast tumor heterogeneity: source of fitness, hurdle for therapy. Mol Cell. 2015;60(4):537–546. doi:10.1016/j.molcel.2015.10.03126590713
- NielsenTO, HsuFD, JensenK, et al. Immunohistochemical and clinical characterization of the basal-like subtype of invasive breast carcinoma. Clin Cancer Res. 2004;10(16):5367–5374. doi:10.1158/1078-0432.CCR-04-022015328174
- WaksAG, WinerEP. Breast cancer treatment: a review. JAMA. 2019;321(3):288–300. doi:10.1001/jama.2018.1932330667505
- FragomeniSM, SciallisA, JerussJS. Molecular subtypes and local-regional control of breast cancer. Surg Oncol Clin N Am. 2018;27(1):95–120. doi:10.1016/j.soc.2017.08.00529132568
- LeiJT, AnuragM, HaricharanS, GouX, EllisMJ. Endocrine therapy resistance: new insights. Breast. 2019;48(Suppl 1):S26–S30. doi:10.1016/S0960-9776(19)31118-X31839155
- AysolaK, DesaiA, WelchC, et al. Triple negative breast cancer – an overview. Hereditary Genet. 2013;Suppl 2:1.
- MukherjeeAK, BasuS, SarkarN, GhoshAC. Advances in cancer therapy with plant based natural products. Curr Med Chem. 2001;8(12):1467–1486. doi:10.2174/092986701337209411562277
- MondalS, BandyopadhyayS, GhoshMK, MukhopadhyayS, RoyS, MandalC. Natural products: promising resources for cancer drug discovery. Anticancer Agents Med Chem. 2012;12(1):49–75. doi:10.2174/18715201279876469721707502
- BonofiglioD, GiordanoC, De AmicisF, LanzinoM, AndòS. Natural products as promising antitumoral agents in breast cancer: mechanisms of action and molecular targets. Mini Rev Med Chem. 2016;16(8):596–604. doi:10.2174/138955751566615070911095926156544
- KhalidF, NawazH, HanifMA, RehmanR, Al-SadiAM. Mango. Medicinal Plants of South Asia: Novel Sources for Drug Discovery. Amsterdam, Oxford, Cambridge MA: Elsevier; 2020:495–508.
- MasiboM, HeQ. Major mango polyphenols and their potential significance to human health. Compr Rev Food Sci Food Saf. 2008;7(4):309–319. doi:10.1111/j.1541-4337.2008.00047.x33467788
- DereseS, GuantaiEM, SouaibouY, KueteV. Mangifera indica L. (Anacardiaceae). Medicinal Spices and Vegetables from Africa. London, San Diego, Cambridge MA, Oxford: Elsevier; 2017:451–483.
- ShahKA, PatelMB, PatelRJ, ParmarPK. Mangifera indica (Mango). Pharmacogn Rev. 2010;4(7):42–48. doi:10.4103/0973-7847.6532522228940
- El-GiedAAA, JosephMR, MahmoudIM, AbdelkareemAM, Al HakamiAM, HamidME. Antimicrobial activities of seed extracts of mango (Mangifera indica L.). Adv Microbiol. 2012;2(4):571–576. doi:10.4236/aim.2012.24074
- KabukiT, NakajimaH, AraiM, UedaS, KuwabaraY, DosakoS. Characterization of novel antimicrobial compounds from mango (Mangifera indica L.) kernel seeds. Food Chem. 2000;71(1):61–66. doi:10.1016/S0308-8146(00)00126-6
- MakareN, BodhankarS, RangariV. Immunomodulatory activity of alcoholic extract of Mangifera indica L. in mice. J Ethnopharmacol. 2001;78(2–3):133–137. doi:10.1016/S0378-8741(01)00326-911694357
- MartinezG, DelgadoR, PérezG, GarridoG, Núñez-SellésA, LeónOS. Evaluation of the in vitro antioxidant activity of Mangifera indica L. extract (Vimang). Phytother Res. 2000;14(6):424–427. doi:10.1002/1099-1573(200009)14:6<424::AID-PTR643>3.0.CO;2-810960895
- BanerjeeN, KimH, KrenekK, TalcottST, Mertens-TalcottSU. Mango polyphenolics suppressed tumor growth in breast cancer xenografts in mice: role of the PI3K/AKT pathway and associated microRNAs. Nutr Res. 2015;35(8):744–751. doi:10.1016/j.nutres.2015.06.00226194618
- García-SolísP, YahiaEM, Morales-TlalpanV, Díaz-MuñozM. Screening of antiproliferative effect of aqueous extracts of plant foods consumed in Mexico on the breast cancer cell line MCF-7. Int J Food Sci Nutr. 2009;60(Suppl 6):32–46. doi:10.1080/09637480802312922
- WilkinsonAS, FlanaganBM, PiersonJT, et al. Bioactivity of mango flesh and peel extracts on peroxisome proliferator‐activated receptor γ[PPARγ] activation and MCF‐7 cell proliferation: fraction and fruit variability. J Food Sci. 2011;76(1):H11–H18. doi:10.1111/j.1750-3841.2010.01899.x21535682
- Fernández-PonceMT, López-BiedmaA, Sánchez-QuesadaC, et al. Selective antitumoural action of pressurized mango leaf extracts against minimally and highly invasive breast cancer. Food Funct. 2017;8(10):3610–3620. doi:10.1039/C7FO00877E28891568
- López-RíosL, WiebeJC, Vega-MoralesT, GerickeN. Central nervous system activities of extract Mangifera indica L. J Ethnopharmacol. 2020;260:112996. doi:10.1016/j.jep.2020.11299632473365
- VenancioVP, KimH, SirvenMA, et al. Polyphenol-rich mango (Mangifera indica L.) ameliorate functional constipation symptoms in humans beyond equivalent amount of fiber. Mol Nutr Food Res. 2018;62(12):e1701034. doi:10.1002/mnfr.20170103429733520
- Anaya-LoyolaMA, García-MarínG, García-GutiérrezDG, et al. A mango (Mangifera indica L.) juice by-product reduces gastrointestinal and upper respiratory tract infection symptoms in children. Food Res Int. 2020;136:109492. doi:10.1016/j.foodres.2020.10949232846573
- López MantecónAM, GarridoG, Delgado-HernándezR, Garrido-SuárezBB. Combination of Mangifera indica L. extract supplementation plus methotrexate in rheumatoid arthritis patients: a pilot study. Phytother Res. 2014;28(8):1163–1172. doi:10.1002/ptr.510824344049
- RodeiroI, DonatoMT, JiménezN, GarridoG, DelgadoR, Gómez-LechónMJ. Effects of Mangifera indica L. aqueous extract (Vimang) on primary culture of rat hepatocytes. Food Chem Toxicol. 2007;45(12):2506–2512. doi:10.1016/j.fct.2007.05.02717651882
- Núñez SellésAJ, Vélez CastroHT, Agüero-AgüeroJ, et al. Isolation and quantitative analysis of phenolic antioxidants, free sugars, and polyols from mango (Mangifera indica L.) stem bark aqueous decoction used in Cuba as a nutritional supplement. J Agric Food Chem. 2002;50(4):762–766. doi:10.1021/jf011064b11829642
- García-RiveraD, DelgadoR, BougarneN, HaegemanG, BergheWV. Gallic acid indanone and mangiferin xanthone are strong determinants of immunosuppressive anti-tumour effects of Mangifera indica L. bark in MDA-MB231 breast cancer cells. Cancer Lett. 2011;305(1):21–31. doi:10.1016/j.canlet.2011.02.01121420233
- Pardo-AndreuGL, DortaDJ, DelgadoR, et al. Vimang (Mangifera indica L. extract) induces permeability transition in isolated mitochondria, closely reproducing the effect of mangiferin, Vimang’s main component. Chem Biol Interact. 2006;159(2):141–148. doi:10.1016/j.cbi.2005.10.10916352298
- GarridoG, GonzálezD, DelporteC, et al. Analgesic and anti-inflammatory effects of Mangifera indica L. extract (Vimang). Phytother Res. 2001;15(1):18–21. doi:10.1002/1099-1573(200102)15:1<18::AID-PTR676>3.0.CO;2-R11180516
- GarridoG, Blanco-MolinaM, SanchoR, MachoA, DelgadoR, MuñozE. An aqueous stem bark extract of Mangifera indica (Vimang) inhibits T cell proliferation and TNF-induced activation of nuclear transcription factor NF-kappaB. Phytother Res. 2005;19(3):211–215. doi:10.1002/ptr.165615934029
- TamayoD, MariE, GonzalezS, et al. Vimang as natural antioxidant supplementation in patients with malignant tumors. Minerva Med. 2001;92(Suppl 1–3):95–97.
- AbdullahAH, MohammedAS, AbdullahR, MirghaniMES, Al-QubaisiM. Cytotoxic effects of Mangifera indica L. kernel extract on human breast cancer (MCF-7 and MDA-MB-231 cell lines) and bioactive constituents in the crude extract. BMC Complement Altern Med. 2014;14(1):199. doi:10.1186/1472-6882-14-19924962691
- BlackHS, Mathews-RothMM. Cytotoxicity and apoptosis induction by butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). Photochem Photobiol. 1991;53(5):707–716. doi:10.1111/j.1751-1097.1991.tb08501.x1881965
- SaitoM, SakagamiH, FujisawaS. Cytotoxicity and apoptosis induction by butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). Anticancer Res. 2003;23(6C):4693–4701.14981915
- KurodaK, AkaoM. Antitumor and anti-intoxication activities of fumaric acid in cultured cells. Gan. 1981;72(5):777–782.7327376
- KurodaK, TeraoK, AkaoM. Inhibitory effect of fumaric acid on hepatocarcinogenesis by thioacetamide in rats. J Natl Cancer Inst. 1987;79(5):1047–1051.3479633
- OkokonJE, DarA, ChoudharyMI. Immunomodulatory, cytotoxic and antileishmanial activity of phytoconstituents of Croton zambesicus. Phytopharmacol. 2013;4(1):31–40.
- SmiljkovicM, StanisavljevicD, StojkovicD, et al. Apigenin-7-O-glucoside versus apigenin: insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017;16:795–807.28827996
- SrivastavaJ, GuptaS. Anti-proliferative and apoptotic effects of chamomile extract in various human cancer cells. J Agric Food Chem. 2007;55(23):9470–9478. doi:10.1021/jf071953k17939735
- PosnerGH, PloypradithP, HapangamaW, et al. Trioxane dimers have potent antimalarial, antiproliferative and antitumor activities in vitro. Bioorg Med Chem. 1997;5(7):1257–1265. doi:10.1016/S0968-0896(97)00079-59377085
- ShacterE, WeitzmanSA. Chronic inflammation and cancer. Oncology. 2002;16(2):217–226, 229.11866137
- BarretoJC, TrevisanMTS, HullWE, et al. Characterization and quantitation of polyphenolic compounds in bark, kernel, leaves, and peel of mango (Mangifera indica L.). J Agric Food Chem. 2008;56(14):5599–5610. doi:10.1021/jf800738r18558692
- GanogpichayagraiA, PalanuvejC, RuangrungsiN. Antidiabetic and anticancer activities of Mangifera indica cv. Okrong leaves. J Adv Pharm Technol Res. 2017;8(1):19–24. doi:10.4103/2231-4040.19737128217550
- ShabanNZ, HegazyWA, Abdel-RahmanSM, AwedOM, KhalilSA. Potential effect of Olea europaea leaves, Sonchus oleraceus leaves and Mangifera indica peel extracts on aromatase activity in human placental microsomes and CYP19A1 expression in MCF-7 cell line: comparative study. Cell Mol Biol (Noisy-Le-Grand). 2016;62(9):11–19.
- HoangVLT, PiersonJT, CurryMC, et al. Polyphenolic contents and the effects of methanol extracts from mango varieties on breast cancer cells. Food Sci Biotechnol. 2015;24(1):265–271. doi:10.1007/s10068-015-0035-x
- TaingMW, PiersonJT, ShawPN, et al. Mango fruit extracts differentially affect proliferation and intracellular calcium signalling in MCF-7 human breast cancer cells. J Chem. 2015;2015:1–10. doi:10.1155/2015/613268
- PiersonJT, MonteithGR, Roberts-ThomsonSJ, DietzgenRG, GidleyMJ, ShawPN. Phytochemical extraction, characterisation and comparative distribution across four mango (Mangifera indica L.) fruit varieties. Food Chem. 2014;149:253–263. doi:10.1016/j.foodchem.2013.10.10824295704
- NorattoGD, BertoldiMC, KrenekK, TalcottST, StringhetaPC, Mertens-TalcottSU. Anticarcinogenic effects of polyphenolics from mango (Mangifera indica) varieties. J Agric Food Chem. 2010;58(7):4104–4112. doi:10.1021/jf903161g20205391
- NemecMJ, KimH, MarcianteAB, BarnesRC, TalcottST, Mertens-TalcottSU. Pyrogallol, an absorbable microbial gallotannins-metabolite and mango polyphenols (Mangifera Indica L.) suppress breast cancer ductal carcinoma in situ proliferation in vitro. Food Funct. 2016;7(9):3825–3833. doi:10.1039/C6FO00636A27491891
- NemecMJ, KimH, MarcianteAB, et al. Polyphenolics from mango (Mangifera indica L.) suppress breast cancer ductal carcinoma in situ proliferation through activation of AMPK pathway and suppression of mTOR in athymic nude mice. J Nutr Biochem. 2017;41:12–19. doi:10.1016/j.jnutbio.2016.11.00527951515
- BenardO, ChiY. Medicinal properties of mangiferin, structural features, derivative synthesis. Pharmacokinetics Biol Activities Mini Rev Med Chem. 2015;15(7):582–594. doi:10.2174/1389557515666150401111410
- BiswasT, SenA, RoyR, MajiS, MajiHS. Isolation of mangiferin from flowering buds of Mangifera indica L and its evaluation of in vitro antibacterial activity. J Pharm Anal. 2015;4(3):49–56.
- DarA, FaiziS, NaqviS, et al. Analgesic and antioxidant activity of mangiferin and its derivatives: the structure activity relationship. Biol Pharm Bull. 2005;28(4):596–600. doi:10.1248/bpb.28.59615802793
- GongX, ZhangL, JiangR, YeM, YinX, WanJ. Anti-inflammatory effects of mangiferin on sepsis-induced lung injury in mice via up-regulation of heme oxygenase-1. J Nutr Biochem. 2013;24(6):1173–1181. doi:10.1016/j.jnutbio.2012.09.00323266284
- MiuraT, IchikiH, HashimotoI, et al. Antidiabetic activity of a xanthone compound, mangiferin. Phytomedicine. 2001;8(2):85–87. doi:10.1078/0944-7113-0000911315760
- Núñez SellesAJ, DagliaM, RastrelliL. The potential role of mangiferin in cancer treatment through its immunomodulatory, anti-angiogenic, apoptopic, and gene regulatory effects. Biofactors. 2016;42(5):475–491. doi:10.1002/biof.129927219221
- RajendranP, JayakumarT, NishigakiI, et al. Immunomodulatory effect of mangiferin in experimental animals with benzo(a)pyrene-induced lung carcinogenesis. Int J Biomed Sci. 2013;9(2):68–74.23847456
- ImranM, ArshadMS, ButtMS, KwonJH, ArshadMU, SultanMT. Mangiferin: a natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017;16(1):84. doi:10.1186/s12944-017-0449-y28464819
- LiH, HuangJ, YangB, et al. Mangiferin exerts antitumor activity in breast cancer cells by regulating matrix metalloproteinases, epithelial to mesenchymal transition, and β-catenin signaling pathway. Toxicol Appl Pharmacol. 2013;272(1):180–190. doi:10.1016/j.taap.2013.05.01123707762
- DengQ, TianYX, LiangJ. Mangiferin inhibits cell migration and invasion through Rac1/WAVE2 signalling in breast cancer. Cytotechnology. 2018;70(2):593–601. doi:10.1007/s10616-017-0140-129455393
- CuccioloniM, BonfiliL, MozzicafreddoM, et al. Mangiferin blocks proliferation and induces apoptosis of breast cancer cells via suppression of mevalonate pathway and by proteasome inhibition. Food Funct. 2016;7(10):4299–4309. doi:10.1039/C6FO01037G27722367
- ThornCF, OshiroC, MarshS, et al. Doxorubicin pathways: pharmacodynamics and adverse effects. Pharmacogenet Genomics. 2011;21(7):440–446. doi:10.1097/FPC.0b013e32833ffb5621048526
- Al-MalkyHS, Al HarthiSE, OsmanAMM. Major obstacles to doxorubicin therapy: cardiotoxicity and drug resistance. J Oncol Pharm Pract. 2020;26(2):434–444. doi:10.1177/107815521987793131594518
- MechetnerE, KyshtoobayevaA, ZonisS, et al. Levels of multidrug resistance (MDR1) P-glycoprotein expression by human breast cancer correlate with in vitro resistance to taxol and doxorubicin. Clin Cancer Res. 1998;4(2):389–398.9516927
- SargentJM, WilliamsonCJ, MaliepaardM, ElgieAW, ScheperRJ, TaylorCG. Breast cancer resistance protein expression and resistance to daunorubicin in blast cells from patients with acute myeloid leukaemia. Br J Haematol. 2001;115(2):257–262. doi:10.1046/j.1365-2141.2001.03122.x11703319
- ZhangYK, WangYJ, GuptaP, ChenZS. Multidrug resistance proteins (MRPs) and cancer therapy. AAPS J. 2015;17(4):802–812. doi:10.1208/s12248-015-9757-125840885
- LouisaM, SoediroTM, SuyatnaFD. In vitro modulation of P-glycoprotein, MRP-1 and BCRP expression by mangiferin in doxorubicin-treated MCF-7 cells. Asian Pac J Cancer Prev. 2014;15(4):1639–1642. doi:10.7314/APJCP.2014.15.4.163924641381
- WilkinsonAS, TaingMW, PiersonJT, et al. Estrogen modulation properties of mangiferin and quercetin and the mangiferin metabolite norathyriol. Food Funct. 2015;6(6):1847–1854. doi:10.1039/C5FO00133A25940566
- WangF, YanJ, NiuY, et al. Mangiferin and its aglycone, norathyriol, improve glucose metabolism by activation of AMP-activated protein kinase. Pharm Biol. 2014;52(1):68–73. doi:10.3109/13880209.2013.81469124033319
- HanB, YangB, YangX, et al. Host-guest inclusion system of norathyriol with β-cyclodextrin and its derivatives: preparation, characterization, and anticancer activity. J Biosci Bioeng. 2014;117(6):775–779. doi:10.1016/j.jbiosc.2013.12.00124508024
- SanugulK, AkaoT, LiY, KakiuchiN, NakamuraN, HattoriM. Isolation of a human intestinal bacterium that transforms mangiferin to norathyriol and inducibility of the enzyme that cleaves a C-glucosyl bond. Biol Pharm Bull. 2005;28(9):1672–1678. doi:10.1248/bpb.28.167216141538
- ShiZL, LiuYD, YuanYY, et al. In vitro and in vivo effects of norathyriol and mangiferin on α-glucosidase. Biochem Res Int. 2017;2017:1206015. doi:10.1155/2017/120601528168055
- WangJP, HoTF, LinCN, TengCM. Effect of norathyriol, isolated from Tripterospermum lanceolatum, on A23187-induced pleurisy and analgesia in mice. Naunyn Schmiedebergs Arch Pharmacol. 1994;350(1):90–95. doi:10.1007/BF001800167935860
- LiJ, MalakhovaM, MottamalM, et al. Norathyriol suppresses solar UV-induced skin cancer by targeting ERKs. Cancer Res. 2012;72(1):260–270. doi:10.1158/0008-5472.CAN-11-259622084399
- TengCM, KoFN, WangJP, et al. Antihaemostatic and antithrombotic effect of some antiplatelet agents isolated from Chinese herbs. J Pharm Pharmacol. 1991;43(9):667–669. doi:10.1111/j.2042-7158.1991.tb03561.x1685529
- KoFN, LinCN, LiouSS, HuangTF, TengCM. Vasorelaxation of rat thoracic aorta caused by norathyriol isolated from Gentianaceae. Eur J Pharmacol. 1991;192(1):133–139. doi:10.1016/0014-2999(91)90079-61645671
- BarnesRC, KimH, FangC, et al. Body mass index as a determinant of systemic exposure to gallotannin metabolites during 6‐week consumption of mango (Mangifera indica L.) and modulation of intestinal microbiota in lean and obese individuals. Mol Nutr Food Res. 2019;63(2):e1800512. doi:10.1002/mnfr.20180051230427574
- FarràsM, Martinez-GiliL, PortuneK, et al. Modulation of the gut microbiota by olive oil phenolic compounds: implications for lipid metabolism, immune system, and obesity. Nutrients. 2020;12(8):2200. doi:10.3390/nu12082200
- GaliHU, PerchelletEM, GaoXM, BottariV, PerchelletJP. Antitumor-promoting effects of gallotannins extracted from various sources in mouse skin in vivo. Anticancer Res. 1993;13(4):915–922.7688939
- HuhJE, LeeEO, KimMS, et al. Penta-O-galloyl-beta-D-glucose suppresses tumor growth via inhibition of angiogenesis and stimulation of apoptosis: roles of cyclooxygenase-2 and mitogen-activated protein kinase pathways. Carcinogenesis. 2005;26(8):1436–1445. doi:10.1093/carcin/bgi09715845650
- MizushinaY, ZhangJ, PuglieseA, KimSH, LüJ. Anti-cancer gallotannin penta-O-galloyl-beta-D-glucose is a nanomolar inhibitor of select mammalian DNA polymerases. Biochem Pharmacol. 2010;80(8):1125–1132. doi:10.1016/j.bcp.2010.06.03120599777
- MohanCG, ViswanathaGL, SavinayG, RajendraCE, HalemaniPD. 1,2,3,4,6 Penta-O-galloyl-β-d-glucose, a bioactivity guided isolated compound from Mangifera indica inhibits 11β-HSD-1 and ameliorates high fat diet-induced diabetes in C57BL/6 mice. Phytomedicine. 2013;20(5):417–426. doi:10.1016/j.phymed.2012.12.02023353053
- ErdèlyiK, KissA, BakondiE, et al. Gallotannin inhibits the expression of chemokines and inflammatory cytokines in A549 cells. Mol Pharmacol. 2005;68(3):895–904. doi:10.1124/mol.105.01251815976037
- EngelsC, KnödlerM, ZhaoYY, CarleR, GänzleMG, SchieberA. Antimicrobial activity of gallotannins isolated from mango (Mangifera indica L.) kernels. J Agric Food Chem. 2009;57(17):7712–7718. doi:10.1021/jf901621m19655802
- CaiY, LuoQ, SunM, CorkeH. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004;74(17):2157–2184. doi:10.1016/j.lfs.2003.09.04714969719
- LimKJA, CabajarAA, LobarbioCFY, TaboadaEB, LacksDJ. Extraction of bioactive compounds from mango (Mangifera indica L. var. Carabao) seed kernel with ethanol-water binary solvent systems. J Food Sci Technol. 2019;56(5):2536–2544. doi:10.1007/s13197-019-03732-731168135
- HuaKT, WayTD, LinJK. Pentagalloylglucose inhibits estrogen receptor alpha by lysosome-dependent depletion and modulates ErbB/PI3K/Akt pathway in human breast cancer MCF-7 cells. Mol Carcinog. 2006;45(8):551–560. doi:10.1002/mc.2022616637063
- ChenWJ, ChangCY, LinJK. Induction of G1 phase arrest in MCF human breast cancer cells by pentagalloylglucose through the down-regulation of CDK4 and CDK2 activities and up-regulation of the CDK inhibitors p27(Kip) and p21(Cip). Biochem Pharmacol. 2003;65(11):1777–1785. doi:10.1016/S0006-2952(03)00156-412781329
- KahkeshaniN, FarzaeiF, FotouhiM, et al. Pharmacological effects of gallic acid in health and diseases: a mechanistic review. Iran J Basic Med Sci. 2019;22(3):225–237.31156781
- KimSH, JunCD, SukK, et al. Gallic acid inhibits histamine release and pro-inflammatory cytokine production in mast cells. Toxicol Sci. 2006;91(1):123–131. doi:10.1093/toxsci/kfj06316322071
- KroesBH. Anti-inflammatory activity of gallic acid. Planta Med. 1992;58(6):499–504. doi:10.1055/s-2006-9615351336604
- SorrentinoE, SucciM, TipaldiL, et al. Antimicrobial activity of gallic acid against food-related Pseudomonas strains and its use as biocontrol tool to improve the shelf life of fresh black truffles. Int J Food Microbiol. 2018;266:183–189. doi:10.1016/j.ijfoodmicro.2017.11.02629227905
- GichnerT, PospísilF, VelemínskýJ, VolkeováV, VolkeJ. Two types of antimutagenic effects of gallic and tannic acids towards N-nitroso-compounds-induced mutagenicity in the Ames Salmonella assay. Folia Microbiol (Praha). 1987;32(1):55–62. doi:10.1007/BF028772593546027
- Velderrain-RodríguezGR, Torres-MorenoH, Villegas-OchoaMA, et al. Gallic acid content and an antioxidant mechanism are responsible for the antiproliferative activity of ‘Ataulfo’ mango peel on LS180 cells. Molecules. 2018;23(3):695. doi:10.3390/molecules23030695
- ZhaoB, HuM. Gallic acid reduces cell viability, proliferation, invasion and angiogenesis in human cervical cancer cells. Oncol Lett. 2013;6(6):1749–1755. doi:10.3892/ol.2013.163224843386
- SubramanianAP, JaganathanSK, MandalM, SupriyantoE, MuhamadII. Gallic acid induced apoptotic events in HCT-15 colon cancer cells. World J Gastroenterol. 2016;22(15):3952–3961. doi:10.3748/wjg.v22.i15.395227099438
- FariedA, KurniaD, FariedLS, et al. Anticancer effects of gallic acid isolated from Indonesian herbal medicine, Phaleria macrocarpa (Scheff.) Boerl, on human cancer cell lines. Int J Oncol. 2007;30(3):605–613.17273761
- TsaiCL, ChiuYM, HoTY, et al. Gallic acid induces apoptosis in human gastric adenocarcinoma cells. Anticancer Res. 2018;38(4):2057–2067.29599323
- LuY, JiangF, JiangH, et al. Gallic acid suppresses cell viability, proliferation, invasion and angiogenesis in human glioma cells. Eur J Pharmacol. 2010;641(2–3):102–107. doi:10.1016/j.ejphar.2010.05.04320553913
- GuR, ZhangM, MengH, XuD, XieY. Gallic acid targets acute myeloid leukemia via Akt/mTOR-dependent mitochondrial respiration inhibition. Biomed Pharmacother. 2018;105:491–497. doi:10.1016/j.biopha.2018.05.15829883944
- SunG, ZhangS, XieY, ZhangZ, ZhaoW. Gallic acid as a selective anticancer agent that induces apoptosis in SMMC-7721 human hepatocellular carcinoma cells. Oncol Lett. 2016;11(1):150–158. doi:10.3892/ol.2015.384526870182
- ZhangT, MaL, WuP, et al. Gallic acid has anticancer activity and enhances the anticancer effects of cisplatin in non‑small cell lung cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol Rep. 2019;41(3):1779–1788.30747218
- JangYG, KoEB, ChoiKC. Gallic acid, a phenolic acid, hinders the progression of prostate cancer by inhibition of histone deacetylase 1 and 2 expression. J Nutr Biochem. 2020;84:108444. doi:10.1016/j.jnutbio.2020.10844432615369
- HsuJD, KaoSH, OuTT, ChenYJ, LiYJ, WangCJ. Gallic acid induces G2/M phase arrest of breast cancer cell MCF-7 through stabilization of p27(Kip1) attributed to disruption of p27(Kip1)/Skp2 complex. J Agric Food Chem. 2011;59(5):1996–2003. doi:10.1021/jf103656v21299246
- WangK, ZhuX, ZhangK, ZhuL, ZhouF. Investigation of gallic acid induced anticancer effect in human breast carcinoma MCF-7 cells. J Biochem Mol Toxicol. 2014;28(9):387–393. doi:10.1002/jbt.2157524864015
- Ozturk SarikayaSB. Acetylcholinesterase inhibitory potential and antioxidant properties of pyrogallol. J Enzyme Inhib Med Chem. 2015;30(5):761–766. doi:10.3109/14756366.2014.96570025297710
- National Toxicology Program. Toxicology and carcinogenesis studies of pyrogallol (CAS No. 87-66-1) in F344/N rats and B6C3F1/N mice (dermal studies). Natl Toxicol Program Tech Rep Ser. 2013;574:1–167.
- NicolisE, LamprontiI, DechecchiMC, et al. Pyrogallol, an active compound from the medicinal plant Emblica officinalis, regulates expression of pro-inflammatory genes in bronchial epithelial cells. Int Immunopharmacol. 2008;8(12):1672–1680. doi:10.1016/j.intimp.2008.08.00118760383
- KocaçalişkanI, TalanI, TerziI. Antimicrobial activity of catechol and pyrogallol as allelochemicals. Z Naturforsch C J Biosci. 2006;61(9–10):639–642. doi:10.1515/znc-2006-9-100417137106
- AhnH, ImE, LeeDY, LeeHJ, JungJH, KimSH. Antitumor effect of pyrogallol via miR-134 mediated S phase arrest and inhibition of PI3K/AKT/Skp2/cMyc signaling in hepatocellular carcinoma. Int J Mol Sci. 2019;20(16):3985. doi:10.3390/ijms20163985
- ParkWH, ParkMN, HanYH, KimSW. Pyrogallol inhibits the growth of gastric cancer SNU-484 cells via induction of apoptosis. Int J Mol Med. 2008;22(2):263–268.18636183
- RevathiS, HakkimFL, Ramesh KumarN, et al. In vivo anti cancer potential of pyrogallol in murine model of colon cancer. Asian Pac J Cancer Prev. 2019;20(9):2645–2651. doi:10.31557/APJCP.2019.20.9.264531554359
- YangCJ, WangCS, HungJY, et al. Pyrogallol induces G2-M arrest in human lung cancer cells and inhibits tumor growth in an animal model. Lung Cancer. 2009;66(2):162–168. doi:10.1016/j.lungcan.2009.01.01619233505
- Abou-ZaidMM, LombardoDA, NozzolilloC. Methyl gallate is a natural constituent of maple (Genus Acer) leaves. Nat Prod Res. 2009;23(15):1373–1377. doi:10.1080/1478641080242045719809908
- ChaudhuriD, GhateNB, SinghSS, MandalN. Methyl gallate isolated from Spondias pinnata exhibits anticancer activity against human glioblastoma by induction of apoptosis and sustained extracellular signal-regulated kinase 1/2 activation. Pharmacogn Mag. 2015;11(42):269–276. doi:10.4103/0973-1296.15307825829764
- MishraRK, RamakrishnaM, MishraV, et al. Pharmaco-phylogenetic investigation of methyl gallate isolated from Acacia nilotica (L.) Delile and Its cytotoxic effect on NIH3T3 mouse fibroblast. Curr Pharm Biotechnol. 2016;17(6):540–548. doi:10.2174/138920101766616012711075926813302
- WangCR, ZhouR, NgTB, WongJH, QiaoWT, LiuF. First report on isolation of methyl gallate with antioxidant, anti-HIV-1 and HIV-1 enzyme inhibitory activities from a mushroom (Pholiota adiposa). Environ Toxicol Pharmacol. 2014;37(2):626–637. doi:10.1016/j.etap.2014.01.02324572641
- RahmanN, JeonM, KimYS. Methyl gallate, a potent antioxidant inhibits mouse and human adipocyte differentiation and oxidative stress in adipocytes through impairment of mitotic clonal expansion. Biofactors. 2016;42(6):716–726. doi:10.1002/biof.131027412172
- KamathamS, KumarN, GudipalliP. Isolation and characterization of gallic acid and methyl gallate from the seed coats of Givotia rottleriformis Griff. and their anti-proliferative effect on human epidermoid carcinoma A431 cells. Toxicol Rep. 2015;2:520–529. doi:10.1016/j.toxrep.2015.03.00128962387
- LeeH, LeeH, KwonY, et al. Methyl gallate exhibits potent antitumor activities by inhibiting tumor infiltration of CD4+CD25+ regulatory T cells. J Immunol. 2010;185(11):6698–6705. doi:10.4049/jimmunol.100137321048105
- ChoiJG, KangOH, LeeYS, et al. In vitro activity of methyl gallate isolated from galla rhois alone and in combination with ciprofloxacin against clinical isolates of salmonella. J Microbiol Biotechnol. 2008;18(11):1848–1852.19047831
- KaneCJ, MennaJH, SungCC, YehYC. Methyl gallate, methyl-3,4,5-trihydroxybenzoate, is a potent and highly specific inhibitor of herpes simplex virus in vitro. II. Antiviral activity of methyl gallate and its derivatives. Biosci Rep. 1988;8(1):95–102. doi:10.1007/BF011289762840133
- CorreaLB, SeitoLN, ManchopeMF, et al. Methyl gallate attenuates inflammation induced by Toll-like receptor ligands by inhibiting MAPK and NF-Κb signaling pathways. Inflamm Res. 2020;69(12):1257–1270. doi:10.1007/s00011-020-01407-033037469
- AsnaashariM, FarhooshR, SharifA. Antioxidant activity of gallic acid and methyl gallate in triacylglycerols of Kilka fish oil and its oil-in-water emulsion. Food Chem. 2014;159:439–444. doi:10.1016/j.foodchem.2014.03.03824767079
- LiY, YaoJ, HanC, et al. Quercetin, Inflammation and Immunity. Nutrients. 2016;8(3):167. doi:10.3390/nu803016726999194
- JiJJ, LinY, HuangSS, ZhangHL, DiaoYP, LiK. Quercetin: a potential natural drug for adjuvant treatment of rheumatoid arthritis. Afr J Tradit Complement Altern Med. 2013;10(3):418–421.24146468
- BuleM, AbdurahmanA, NikfarS, AbdollahiM, AminiM. Antidiabetic effect of quercetin: a systematic review and meta-analysis of animal studies. Food Chem Toxicol. 2019;125:494–502. doi:10.1016/j.fct.2019.01.03730735748
- WangS, YaoJ, ZhouB, et al. Bacteriostatic effect of quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. J Food Prot. 2018;81(1):68–78. doi:10.4315/0362-028X.JFP-17-21429271686
- ComaladaM, CamuescoD, SierraS, et al. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-kappaB pathway. Eur J Immunol. 2005;35(2):584–592. doi:10.1002/eji.20042577815668926
- SongX, WangY, GaoL. Mechanism of antioxidant properties of quercetin and quercetin-DNA complex. J Mol Model. 2020;26(6):133. doi:10.1007/s00894-020-04356-x32399900
- AlexanderSP. Flavonoids as antagonists at A1 adenosine receptors. Phytother Res. 2006;20(11):1009–1012. doi:10.1002/ptr.197517006974
- YinG, WangZ, WangZ, WangX. Topical application of quercetin improves wound healing in pressure ulcer lesions. Exp Dermatol. 2018;27(7):779–786. doi:10.1111/exd.1367929733461
- ZhangXA, ZhangS, YinQ, ZhangJ. Quercetin induces human colon cancer cells apoptosis by inhibiting the nuclear factor-kappa B pathway. Pharmacogn Mag. 2015;11(42):404–409. doi:10.4103/0973-1296.15309625829782
- HanC, GaoH, ZhangX. Quercetin anti-cancer effect in renal cancer through regulating survivin expression and caspase 3 activity. Med One. 2016;1(1):3.
- DongY, YangJ, YangL, LiP. Quercetin inhibits the proliferation and metastasis of human non-small cell lung cancer cell line: the key role of src-mediated fibroblast growth factor-inducible 14 (Fn14)/nuclear factor kappa B (NF-κB) pathway. Med Sci Monit. 2020;26:e920537. doi:10.12659/MSM.92053732225128
- OngCS, TranE, NguyenTT, et al. Quercetin-induced growth inhibition and cell death in nasopharyngeal carcinoma cells are associated with increase in Bad and hypophosphorylated retinoblastoma expressions. Oncol Rep. 2004;11(3):727–733.14767529
- TeekaramanD, ElayapillaiSP, ViswanathanMP, JagadeesanA. Quercetin inhibits human metastatic ovarian cancer cell growth and modulates components of the intrinsic apoptotic pathway in PA-1 cell line. Chem Biol Interact. 2019;300:91–100. doi:10.1016/j.cbi.2019.01.00830639267
- AngstE, ParkJL, MoroA, et al. The flavonoid quercetin inhibits pancreatic cancer growth in vitro and in vivo. Pancreas. 2013;42(2):223–229. doi:10.1097/MPA.0b013e318264ccae23000892
- WardAB, MirH, KapurN, GalesDN, CarrierePP, SinghS. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J Surg Oncol. 2018;16(1):108. doi:10.1186/s12957-018-1400-z29898731
- ZhangH, ZhangM, YuL, ZhaoY, HeN, YangX. Antitumor activities of quercetin and quercetin-5’,8-disulfonate in human colon and breast cancer cell lines. Food Chem Toxicol. 2012;50(5):1589–1599. doi:10.1016/j.fct.2012.01.02522310237
- ChouCC, YangJS, LuHF, et al. Quercetin-mediated cell cycle arrest and apoptosis involving activation of a caspase cascade through the mitochondrial pathway in human breast cancer MCF-7 cells. Arch Pharm Res. 2010;33(8):1181–1191. doi:10.1007/s12272-010-0808-y20803121
- ChienSY, WuYC, ChungJG, et al. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum Exp Toxicol. 2009;28(8):493–503. doi:10.1177/096032710910700219755441
- Rivera RiveraA, Castillo-PichardoL, GerenaY, DharmawardhaneS. Anti-breast cancer potential of quercetin via the Akt/AMPK/mammalian target of rapamycin (mTOR) signaling cascade. PLoS One. 2016;11(6):e0157251. doi:10.1371/journal.pone.015725127285995
- SchieberM, ChandelNS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014;24(10):453–462. doi:10.1016/j.cub.2014.03.034
- MatésJM, SeguraJA, AlonsoFJ, MárquezJ. Intracellular redox status and oxidative stress: implications for cell proliferation, apoptosis, and carcinogenesis. Arch Toxicol. 2008;82(5):273–299. doi:10.1007/s00204-008-0304-z18443763
- NogueiraV, HayN. Molecular pathways: reactive oxygen species homeostasis in cancer cells and implications for cancer therapy. Clin Cancer Res. 2013;19(16):4309–4314. doi:10.1158/1078-0432.CCR-12-142423719265
- NimseSB, PalD. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015;5(35):27986–28006. doi:10.1039/C4RA13315C
- BurtonGJ, JauniauxE. Oxidative stress. Best Pract Res Clin Obstet Gynaecol. 2011;25(3):287–299. doi:10.1016/j.bpobgyn.2010.10.01621130690
- AggarwalV, TuliHS, VarolA, et al. Role of reactive oxygen species in cancer progression: molecular mechanisms and recent advancements. Biomolecules. 2019;9(11):735. doi:10.3390/biom9110735
- MatésJM, SeguraJA, AlonsoFJ, MárquezJ. Oxidative stress in apoptosis and cancer: an update. Arch Toxicol. 2012;86(11):1649–1665.22811024
- StorzP. Oxidative stress and cancer. In: JakobU, ReichmannD, editors. Oxidative Stress and Redox Regulation. Berlin: Springer; 2004:427–447.
- GreenDR. Means to an End: Apoptosis and Other Cell Death Mechanisms. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 2011.
- AbdullahAH, MohammedAS, RasedeeA, MirghaniMES. Oxidative stress-mediated apoptosis induced by ethanolic mango seed extract in cultured estrogen receptor positive breast cancer MCF-7 cells. Int J Mol Sci. 2015a;16(2):3528–3536. doi:10.3390/ijms1602352825664859
- AbdullahAH, MohammedAS, RasedeeA, MirghaniMES, Al-QubaisiMS. Induction of apoptosis and oxidative stress in estrogen receptor-negative breast cancer, MDA-MB231 cells, by ethanolic mango seed extract. BMC Compl Alternative Med. 2015b;15:45. doi:10.1186/s12906-015-0575-x
- SchumackerPT. Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell. 2006;10(3):175–176. doi:10.1016/j.ccr.2006.08.01516959608
- StorzP. Reactive oxygen species in tumor progression. Front Biosci. 2005;10:1881–1896. doi:10.2741/166715769673
- LiouGY, StorzP. Reactive oxygen species in cancer. Free Radic Res. 2010;44(5):479–496. doi:10.3109/1071576100366755420370557
- Fernández-PonceMT, CasasL, MantellC. Use of high pressure techniques to produce Mangifera indica L. leaf extracts enriched in potent antioxidant phenolic compounds. Innov Food Sci Emerg Tech. 2015;29:94–106. doi:10.1016/j.ifset.2015.04.006
- Fernández PonceMT, CasasL, MantellC, RodríguezM. Extraction of antioxidant compounds from different varieties of Mangifera indica leaves using green technologies. J Supercrit Fluids. 2012;72:168–175. doi:10.1016/j.supflu.2012.07.016
- TyagiS, GuptaP, SainiAS, KaushalC, SharmaS. The peroxisome proliferator-activated receptor: a family of nuclear receptors role in various diseases. J Adv Pharm Technol Res. 2011;2(4):236–240. doi:10.4103/2231-4040.9087922247890
- WagnerN, WagnerKD. PPAR Beta/Delta and the Hallmarks of Cancer. Cells. 2020;9(5):1133. doi:10.3390/cells9051133
- TachibanaK, YamasakiD, IshimotoK, DoiT. The role of PPARs in cancer. PPAR Res. 2008;2008:102737. doi:10.1155/2008/10273718584037
- AshbyJ, BradyA, ElcombeCR, et al. Mechanistically-based human hazard assessment of peroxisome proliferator-induced hepatocarcinogenesis. Hum Exp Toxicol. 1994;13(Suppl 2):S1–S117. doi:10.1177/096032719401300201
- IssemannI, GreenS. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature. 1990;347:645–650. doi:10.1038/347645a02129546
- ReddyJK, AzarnoffDL, HigniteCE. Hypolipidaemic hepatic peroxisome proliferators form a novel class of chemical carcinogens. Nature. 1980;283:397–398. doi:10.1038/283397a06766207
- SahaSA, KizhakepunnurLG, BahekarA, AroraRR. The role of fibrates in the prevention of cardiovascular disease—a pooled meta-analysis of long-term randomized placebo-controlled clinical trials. Am Heart J. 2007;154(5):943–953. doi:10.1016/j.ahj.2007.07.01117967602
- ParkBH, VogelsteinB, KinzlerKW. Genetic disruption of PPARδ decreases the tumorigenicity of human colon cancer cells. Proc Natl Acad Sci U S A. 2001;98(5):2598–2603. doi:10.1073/pnas.05163099811226285
- WangD, WangH, GuoY, et al. Crosstalk between peroxisome proliferator-activated receptor δ and VEGF stimulates cancer progression. Proc Natl Acad Sci U S A. 2006;103(50):19069–19074. doi:10.1073/pnas.060794810317148604
- ZuoX, PengZ, MoussalliMJ, et al. Targeted genetic disruption of peroxisome proliferator–activated receptor-δ and colonic tumorigenesis. J Natl Cancer Inst. 2009;101(10):762–767. doi:10.1093/jnci/djp07819436036
- HarmanFS, NicolCJ, MarinHE, WardJM, GonzalezFJ, PetersJM. Peroxisome proliferator-activated receptor-delta attenuates colon carcinogenesis. Nat Med. 2004;10(5):481–483. doi:10.1038/nm102615048110
- MarinHE, PerazaMA, BillinAN, et al. Ligand activation of peroxisome proliferator-activated receptor beta inhibits colon carcinogenesis. Cancer Res. 2006;66(8):4394–4401. doi:10.1158/0008-5472.CAN-05-427716618765
- DivyaGS, MansoorKP, RasheedSP, KumarA. PPAR gamma agonists: an effective strategy for cancer treatment. J Pharm Sci Innov. 2013;2(5):1–3. doi:10.7897/2277-4572.02575
- KitamuraS, MiyazakiY, HiraokaS, et al. PPARgamma agonists inhibit cell growth and suppress the expression of cyclin D1 and EGF-like growth factors in ras-transformed rat intestinal epithelial cells. Int J Cancer. 2001;94(3):335–342. doi:10.1002/ijc.147011745411
- YangWL, FruchtH. Activation of the PPAR pathway induces apoptosis and COX-2 inhibition in HT-29 human colon cancer cells. Carcinogenesis. 2001;22(9):1379–1383. doi:10.1093/carcin/22.9.137911532858
- LefebvreAM, ChenI, DesreumauxP, et al. Activation of the peroxisome proliferator-activated receptor γ promotes the development of colon tumors in C57BL/6J-APCMin/+ mice. Nature Med. 1998;4(9):1053–1057.9734399
- SaezE, TontonozP, NelsonMC, et al. Activators of the nuclear receptor PPARgamma enhance colon polyp formation. Nat Med. 1998;4(9):1058–1061. doi:10.1038/20429734400
- XuF, NaL, LiY, ChenL. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci. 2020;10(1):54. doi:10.1186/s13578-020-00416-032266056
- HemmingsBA, RestucciaDF. PI3K-PKB/Akt Pathway. Cold Spring Harb Perspect Biol. 2012;4(9):a011189. doi:10.1101/cshperspect.a01118922952397
- OrtegaMA, Fraile-MartínezO, AsúnsoloA, BujánJ, García-HonduvillaN, CocaS. Signal transduction pathways in breast cancer: the important role of PI3K/Akt/mTOR. J Oncol. 2020;2020:9258396. doi:10.1155/2020/925839632211045
- PaplomataE, O’ReganR. The PI3K/AKT/mTOR pathway in breast cancer: targets, trials and biomarkers. Ther Adv Med Oncol. 2014;6(4):154–166. doi:10.1177/175883401453002325057302
- LiuR, ChenY, LiuG, et al. PI3K/AKT pathway as a key link modulates the multidrug resistance of cancers. Cell Death Dis. 2020;11(9):797. doi:10.1038/s41419-020-02998-632973135
- BrownKK, TokerA. The phosphoinositide 3-kinase pathway and therapy resistance in cancer. F1000Prime Rep. 2015;7:13. doi:10.12703/P7-1325750731
- WilksST. Potential of overcoming resistance to HER2-targeted therapies through the PI3K/Akt/mTOR pathway. Breast. 2015;24(5):548–555. doi:10.1016/j.breast.2015.06.00226187798
- LiS, PatelDJ. Drosha and dicer: slicers cut from the same cloth. Cell Res. 2016;26(5):511–512. doi:10.1038/cr.2016.1927125999
- DongY, FuC, GuanH, ZhangZ, ZhouT, LiB. Prognostic significance of miR-126 in various cancers: a meta-analysis. Onco Targets Ther. 2016;9:2547–2555. doi:10.2147/OTT.S10348127217773
- MeisterJ, SchmidtMHH. miR-126 and miR-126*: new Players in Cancer. ScientificWorldJournal. 2010;10:2090–2100. doi:10.1100/tsw.2010.19820953557
- TavazoieSF, AlarcónC, OskarssonT, et al. Endogenous human microRNAs that suppress breast cancer metastasis. Nature. 2008;451(7175):147–152. doi:10.1038/nature0648718185580
- WangCZ, YuanP, LiY. MiR-126 regulated breast cancer cell invasion by targeting ADAM9. Int J Clin Exp Pathol. 2015;8(6):6547–6553.26261534
- ChumsriS, HowesT, BaoT, SabnisG, BrodieA. Aromatase, aromatase inhibitors, and breast cancer. J Steroid Biochem Mol Biol. 2011;125(1–2):13–22. doi:10.1016/j.jsbmb.2011.02.00121335088
- Saha RoyS, VadlamudiRK. Role of estrogen receptor signaling in breast cancer metastasis. Int J Breast Cancer. 2012;2012:654698. doi:10.1155/2012/65469822295247
- ChumsriS. Clinical utilities of aromatase inhibitors in breast cancer. Int J Womens Health. 2015;7:493–499. doi:10.2147/IJWH.S6990726005359
- ZhaoH, InnesJ, BrooksDC, et al. A novel promoter controls Cyp19a1 gene expression in mouse adipose tissue. Reprod Biol Endocrinol. 2009;7(1):37. doi:10.1186/1477-7827-7-3719393092
- ZhaoH, ZhouL, ShangguanAJ, BulunSE. Aromatase expression and regulation in breast and endometrial cancer. J Mol Endocrinol. 2016;57(1):R19–R33. doi:10.1530/JME-15-031027067638
- ChenS, ZhouD, OkuboT, et al. Prevention and treatment of breast cancer by suppressing aromatase activity and expression. Ann N Y Acad Sci. 2002;963(1):229–238. doi:10.1111/j.1749-6632.2002.tb04115.x12095949
- SunF, XuX, WangX, ZhangB. Regulation of autophagy by Ca2+ . Tumour Biol. 2016;37(12):15467–15476. doi:10.1007/s13277-016-5353-y
- MachacaK. Ca2+ signaling, genes and the cell cycle. Cell Calcium. 2010;48(5):243–250. doi:10.1016/j.ceca.2010.10.00321084120
- PintonP, GiorgiC, SivieroR, ZecchiniE, RizzutoR. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene. 2008;27(50):6407–6418. doi:10.1038/onc.2008.30818955969
- TonelliFMP, SantosAK, GomesDA, et al. Stem cells and calcium signaling. Adv Exp Med Biol. 2012;740:891–916.22453975
- BaranI. Calcium and cell cycle progression: possible effects of external perturbations on cell proliferation. Biophys J. 1996;70(3):1198–1213. doi:10.1016/S0006-3495(96)79679-08785278
- IamshanovaO, Fiorio PlaA, PrevarskayaN. Molecular mechanisms of tumour invasion: regulation by calcium signals. J Physiol. 2017;595(10):3063–3075. doi:10.1113/JP27284428304082
- RimessiA, PedrialiG, VezzaniB, et al. Interorganellar calcium signaling in the regulation of cell metabolism: a cancer perspective. Semin Cell Dev Biol. 2020;98:167–180.31108186
- TsaiFC, KuoGH, ChangSW, TsaiPJ. Ca2+ signaling in cytoskeletal reorganization, cell migration, and cancer metastasis. Biomed Res Int. 2015;2015:409245. doi:10.1155/2015/40924525977921
- VilborgA, PassarelliMC, SteitzJA. Calcium signaling and transcription: elongation, DoGs, and eRNAs. Receptors Clin Investig. 2016;3(1):e1169.
- PrattSJP, Hernández-OchoaE, MartinSS. Calcium signaling: breast cancer’s approach to manipulation of cellular circuitry. Biophys Rev. 2020;12(6):1343–1359. doi:10.1007/s12551-020-00771-933569087
- BootmanMD. Calcium signaling. Cold Spring Harb Perspect Biol. 2012;4(7):a011171. doi:10.1101/cshperspect.a01117122751152
- YáñezM, Gil-LongoJ, Campos-ToimilM. Calcium binding proteins. Adv Exp Med Biol. 2012;740:461–482.22453954
- SoCL, SaunusJM, Roberts-ThomsonSJ, MonteithGR. Calcium signalling and breast cancer. Semin Cell Dev Biol. 2019;94:74–83. doi:10.1016/j.semcdb.2018.11.00130439562
- GhoshS, MayMJ, KoppEB. NF-kappa B and Rel proteins: evolutionarily conserved mediators of immune responses. Annu Rev Immunol. 1998;16(1):225–260. doi:10.1146/annurev.immunol.16.1.2259597130
- WangW, NagSA, ZhangR. Targeting the NFκB signaling pathways for breast cancer prevention and therapy. Curr Med Chem. 2015;22(2):264–289. doi:10.2174/092986732166614110612431525386819
- GhoshG, WangVY, HuangDB, FuscoA. NF-κB regulation: lessons from structures. Immunol Rev. 2012;246(1):36–58. doi:10.1111/j.1600-065X.2012.01097.x22435546
- O’DeaE, HoffmannA. NF-κB signaling. Wiley Interdiscip Rev Syst Biol Med. 2009;1(1):107. doi:10.1002/wsbm.3020151024
- LiuT, ZhangL, JooD, SunSC. NF-κB signaling in inflammation Signal Transduct Target Ther. Signal Transduct Targeted Therapy. 2017;2:17023.
- ParkYH. The nuclear factor-kappa B pathway and response to treatment in breast cancer. Pharmacogenomics. 2017;18(18):1697–1709. doi:10.2217/pgs-2017-004429182047
- HuberMA, AzoiteiN, BaumannB, et al. NF-kappaB is essential for epithelial-mesenchymal transition and metastasis in a model of breast cancer progression. J Clin Invest. 2004;114(4):569–581. doi:10.1172/JCI20042135815314694
- KhongthongP, RoseweirAK, EdwardsJ. The NF-KB pathway and endocrine therapy resistance in breast cancer. Endocr Relat Cancer. 2019;26(6):R369–R380. doi:10.1530/ERC-19-008732013374
- LiF, SethiG. Targeting transcription factor NF-kappaB to overcome chemoresistance and radioresistance in cancer therapy. Biochim Biophys Acta. 2010;1805(2):167–180.20079806
- ShibataA, NagayaT, ImaiT, FunahashiH, NakaoA, SeoH. Inhibition of NF-κB activity decreases the VEGF mRNA expression in MDA-MB-231 breast cancer cells. Breast Cancer Res Treat. 2002;73(3):237–243. doi:10.1023/A:101587253167512160329
- XiaY, ShenS, VermaIM. NF-κB, an active player in human cancers. Cancer Immunol Res. 2014;2(9):823–830. doi:10.1158/2326-6066.CIR-14-011225187272
- BaudV, KarinM. Is NF-κB a good target for cancer therapy? Hopes and pitfalls. Nat Rev Drug Discov. 2009;8(1):33–40. doi:10.1038/nrd278119116625
- ViatourP, MervilleMP, BoursV, ChariotA. Phosphorylation of NF-kB and IkB proteins: implications in cancer and inflammation. Trends Biochem Sci. 2005;30(1):43–52. doi:10.1016/j.tibs.2004.11.00915653325
- SarkarA, SreenivasanY, RameshGT, MannaSK. beta-D-Glucoside suppresses tumor necrosis factor-induced activation of nuclear transcription factor kappaB but potentiates apoptosis. J Biol Chem. 2004;279(32):33768–33781. doi:10.1074/jbc.M40342420015161907
- KomiyaY, HabasR. Wnt signal transduction pathways. Organogenesis. 2008;4(2):68–75. doi:10.4161/org.4.2.585119279717
- HeX, SemenovM, TamaiK, ZengX. LDL receptor-related proteins 5 and 6 in Wnt/β-catenin signaling: arrows point the way. Development. 2004;131(8):1663–1677. doi:10.1242/dev.0111715084453
- MacDonaldBT, TamaiK, HeX. Wnt/β-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17(1):9–26. doi:10.1016/j.devcel.2009.06.01619619488
- KhalafAM, FuentesD, MorshidAI, et al. Role of Wnt/β-catenin signaling in hepatocellular carcinoma, pathogenesis, and clinical significance. J Hepatocell Carcinoma. 2018;5:61–73. doi:10.2147/JHC.S15670129984212
- SethiJK, Vidal-PuigA. Wnt signalling and the control of cellular metabolism. Biochem J. 2010;427(1):1–17. doi:10.1042/BJ2009186620226003
- RenL, ChenH, SongJ, et al. MiR-454-3p-mediated Wnt/β-catenin signaling antagonists suppression promotes breast cancer metastasis. Theranostics. 2019;9(2):449–465. doi:10.7150/thno.2905530809286
- ArendRC, Londoño-JoshiAI, StraughnJMJ, BuchsbaumDJ. The Wnt/β-catenin pathway in ovarian cancer: a review. Gynecol Oncol. 2013;131(3):772–779. doi:10.1016/j.ygyno.2013.09.03424125749
- DeP, CarlsonJH, WuH, MarcusA, Leyland-JonesB, DeyN. Wnt-beta-catenin pathway signals metastasis-associated tumor cell phenotypes in triple negative breast cancers. Oncotarget. 2016;7(28):43124–43149. doi:10.18632/oncotarget.898827281609
- QuB, LiuBR, DuYJ, et al. Wnt/β-catenin signaling pathway may regulate the expression of angiogenic growth factors in hepatocellular carcinoma. Oncol Lett. 2014;7(4):1175–1178. doi:10.3892/ol.2014.182824944688
- JangGB, KimJY, ChoSD, et al. Blockade of Wnt/β-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci Rep. 2015;5(1):12465. doi:10.1038/srep1246526202299
- DeBose-BoydRA. Feedback regulation of cholesterol synthesis: sterol-accelerated ubiquitination and degradation of HMG CoA reductase. Cell Res. 2008;18(6):609–621. doi:10.1038/cr.2008.6118504457
- DaniloC, FrankPG. Cholesterol and breast cancer development. Curr Opin Pharmacol. 2012;12(6):677–682. doi:10.1016/j.coph.2012.07.00922867847
- SantosCR, DominguesG, MatiasI, et al. LDL-cholesterol signaling induces breast cancer proliferation and invasion. Lipids Health Dis. 2014;13(1):16. doi:10.1186/1476-511X-13-1624428917
- StancuC, SimaA. Statins: mechanism of action and effects. J Cell Mol Med. 2001;5(4):378–387. doi:10.1111/j.1582-4934.2001.tb00172.x12067471
- BorgquistS, BjarnadottirO, KimbungS, AhernTP. Statins: a role in breast cancer therapy?J Intern Med. 2018;284(4):346–357. doi:10.1111/joim.1280629923256
- HuangQ, Figueiredo-PereiraME. Ubiquitin/proteasome pathway impairment in neurodegeneration: therapeutic implications. Apoptosis. 2010;15(11):1292–1311.20131003
- CrawfordLJ, WalkerB, IrvineAE. Proteasome inhibitors in cancer therapy. J Cell Commun Signal. 2011;5(2):101–110. doi:10.1007/s12079-011-0121-721484190
- AgyinJK, SanthammaB, NairHB, RoySS, TekmalRR. BU-32: a novel proteasome inhibitor for breast cancer. Breast Cancer Res. 2009;11(5):R74. doi:10.1186/bcr241119821999
- JonesMD, LiuJC, BarthelTK, et al. A proteasome inhibitor, bortezomib, inhibits breast cancer growth and reduces osteolysis by downregulating metastatic genes. Clin Cancer Res. 2010;16(20):4978–4989. doi:10.1158/1078-0432.CCR-09-329320843837
- ShinoharaK, TomiokaM, NakanoH, TonéS, ItoH, KawashimaS. Apoptosis induction resulting from proteasome inhibition. Biochem J. 1996;317(Pt 2):385–388. doi:10.1042/bj31703858713062
- van der VormLN, RemijnJA, de LaatB, HuskensD. Effects of plasmin on von Willebrand factor and platelets: a narrative review. TH Open. 2018;2(2):e218–e228.31249945
- DidiasovaM, WujakL, WygreckaM, ZakrzewiczD. From plasminogen to plasmin: role of plasminogen receptors in human cancer. Int J Mol Sci. 2014;15(11):21229–21252. doi:10.3390/ijms15112122925407528
- SwedbergJE, HarrisJM. Natural and engineered plasmin inhibitors: applications and design strategies. Chembiochem. 2012;13(3):336–348. doi:10.1002/cbic.20110067322238174
- CastellinoFJ, PloplisVA. Structure and function of the plasminogen/plasmin system. Thromb Haemost. 2005;93(4):647–654. doi:10.1160/TH04-12-084215841308
- DeryuginaEI, QuigleyJP. Cell surface remodeling by plasmin: a new function for an old enzyme. J Biomed Biotechnol. 2012;2012:564259. doi:10.1155/2012/56425923097597
- HayashidoY, HamanaT, YoshiokaY, KitanoH, KoizumiK, OkamotoT. Plasminogen activator/plasmin system suppresses cell-cell adhesion of oral squamous cell carcinoma cells via proteolysis of E-cadherin. Int J Oncol. 2005;27(3):693–698.16077918
- SpieringD, HodgsonL. Dynamics of the Rho-family small GTPases in actin regulation and motility. Cell Adh Migr. 2011;5(2):170–180. doi:10.4161/cam.5.2.1440321178402
- AlgayadhIG, DronamrajuV, SylvesterPW. Role of Rac1/WAVE2 signaling in mediating the inhibitory effects of γ-tocotrienol on mammary cancer cell migration and invasion. Biol Pharm Bull. 2016;39(12):1974–1982. doi:10.1248/bpb.b16-0046127904039
- BompardG, CaronE. Regulation of WASP/WAVE proteins: making a long story short. J Cell Biol. 2004;166(7):957–962. doi:10.1083/jcb.20040312715452139
- KurisuS, TakenawaT. WASP and WAVE family proteins: friends or foes in cancer invasion?Cancer Sci. 2010;101(10):2093–2104. doi:10.1111/j.1349-7006.2010.01654.x20707804
- FernandoHS, DaviesSR, ChhabraA, et al. Expression of the WASP verprolin-homologues (WAVE members) in human breast cancer. Oncology. 2007;73(5–6):376–383. doi:10.1159/00013615718509249
- PorterBA, OrtizMA, BratslavskyG, KotulaL. Structure and function of the nuclear receptor superfamily and current targeted therapies of prostate cancer. Cancers. 2019;11(12):1852. doi:10.3390/cancers11121852
- BjörnströmL, SjöbergM. Mechanisms of estrogen receptor signaling: convergence of genomic and nongenomic actions on target genes. Mol Endocrinol. 2005;19(4):833–842. doi:10.1210/me.2004-048615695368
- AcconciaF, KumarR. Signaling regulation of genomic and nongenomic functions of estrogen receptors. Cancer Lett. 2006;238(1):1–14. doi:10.1016/j.canlet.2005.06.01816084012
- VrtačnikP, OstanekB, Mencej-BedračS, MarcJ. The many faces of estrogen signaling. Biochem Med (Zagreb). 2014;24(3):329–342. doi:10.11613/BM.2014.03525351351
- WilliamsC, LinCY. Oestrogen receptors in breast cancer: basic mechanisms and clinical implications. Ecancermedicalscience. 2013;7(1):370.24222786
- HartmanJ, GustafssonJA. Estrogen receptor beta in breast cancer–diagnostic and therapeutic implications. Steroids. 2009;74(8):635–641. doi:10.1016/j.steroids.2009.02.00519463683
- PowellE, ShanleE, BrinkmanA, et al. Identification of estrogen receptor dimer selective ligands reveals growth-inhibitory effects on cells that co-express ERα and ERβ. PLoS One. 2012;7(2):e30993. doi:10.1371/journal.pone.003099322347418
- SanzP. AMP-activated protein kinase: structure and regulation. Curr Protein Pept Sci. 2008;9(5):478–492. doi:10.2174/13892030878591525418855699
- MihaylovaMM, ShawRJ. The AMP-activated protein kinase (AMPK) signaling pathway coordinates cell growth, autophagy, & metabolism. Nat Cell Biol. 2011;13(9):1016–1023. doi:10.1038/ncb232921892142
- XuQ, SiLY. Protective effects of AMP-activated protein kinase in the cardiovascular system. J Cell Mol Med. 2010;14(11):2604–2613. doi:10.1111/j.1582-4934.2010.01179.x20874722
- HinchyEC, GruszczykAV, WillowsR, et al. Mitochondria-derived ROS activate AMP-activated protein kinase (AMPK) indirectly. J Biol Chem. 2018;293(44):17208–17217. doi:10.1074/jbc.RA118.00257930232152
- RabinovitchRC, SamborskaB, FaubertB, et al. AMPK maintains cellular metabolic homeostasis through regulation of mitochondrial reactive oxygen species. Cell Rep. 2017;21(1):1–9. doi:10.1016/j.celrep.2017.09.02628978464
- WangS, SongP, ZouMH. AMP-activated protein kinase, stress responses and cardiovascular diseases. Clin Sci (Lond). 2012;122(12):555–573. doi:10.1042/CS2011062522390198
- YiY, ChenD, AoJ, et al. Transcriptional suppression of AMPKα1 promotes breast cancer metastasis upon oncogene activation. Proc Natl Acad Sci U S A. 2020;117(14):8013–8021. doi:10.1073/pnas.191478611732193335
- FoxMM, PhoenixKN, KopsiaftisSG, ClaffeyKP. AMP-activated protein kinase α 2 isoform suppression in primary breast cancer alters AMPK growth control and apoptotic signaling. Genes Cancer. 2013;4(1–2):3–14. doi:10.1177/194760191348634623946867
- JhaveriTZ, WooJ, ShangX, ParkBH, GabrielsonE. AMP-activated kinase (AMPK) regulates activity of HER2 and EGFR in breast cancer. Oncotarget. 2015;6(17):14754–14765. doi:10.18632/oncotarget.447426143491
- ZouYF, XieCW, YangSX, XiongJP. AMPK activators suppress breast cancer cell growth by inhibiting DVL3-facilitated Wnt/β-catenin signaling pathway activity. Mol Med Rep. 2017;15(2):899–907. doi:10.3892/mmr.2016.609428035400
- Ben-SahraI, DiratB, LaurentK, et al. Sestrin2 integrates Akt and mTOR signaling to protect cells against energetic stress-induced death. Cell Death Differ. 2013;20(4):611–619. doi:10.1038/cdd.2012.15723238567
- KimSH, ParkS, YuHS, KoKH, ParkHG, KimYS. The antipsychotic agent clozapine induces autophagy via the AMPK-ULK1-Beclin1 signaling pathway in the rat frontal cortex. Prog Neuropsychopharmacol Biol Psychiatry. 2018;81:96–104. doi:10.1016/j.pnpbp.2017.10.01229079139
- VermeulenK, Van BockstaeleDR, BernemanZN. The cell cycle: a review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003;36(3):131–149. doi:10.1046/j.1365-2184.2003.00266.x12814430
- CasimiroMC, CrosariolM, LoroE, LiZ, PestellRG. Cyclins and cell cycle control in cancer and disease. Genes Cancer. 2012;3(11–12):649–657. doi:10.1177/194760191347902223634253
- TopacioBR, ZatulovskiyE, CristeaS, et al. Cyclin D-Cdk4,6 drives cell-cycle progression via the retinoblastoma protein’s c-terminal helix. Mol Cell. 2019;74(4):758–770. doi:10.1016/j.molcel.2019.03.02030982746
- BertoliC, SkotheimJM, de BruinRA. Control of cell cycle transcription during G1 and S phases. Nat Rev Mol Cell Biol. 2013;14(8):518–528. doi:10.1038/nrm362923877564
- VoellerD, RahmanL, Zajac-KayeM. Elevated levels of thymidylate synthase linked to neoplastic transformation of mammalian cells. Cell Cycle. 2004;3(8):1005–1007. doi:10.4161/cc.3.8.106415280655
- NarasimhaAM, KaulichM, ShapiroGS, ChoiYJ, SicinskiP, DowdySF. Cyclin D activates the Rb tumor suppressor by mono-phosphorylation. eLife. 2014;3:e02872. doi:10.7554/eLife.02872
- Lara-GonzalezP, MoyleMW, BudrewiczJ, Mendoza-LopezJ, OegemaK, DesaiA. The G2-to-M transition is ensured by a dual mechanism that protects cyclin B from degradation by Cdc20-activated APC/C. Dev Cell. 2019;51(3):313–325. doi:10.1016/j.devcel.2019.09.00531588029
- TavakolianS, GoudarziH, FaghihlooE. Cyclin-dependent kinases and CDK inhibitors in virus-associated cancers. Infect Agent Cancer. 2020;15:27. doi:10.1186/s13027-020-00295-732377232
- ThuKL, Soria-BretonesI, MakTW, CesconDW. Targeting the cell cycle in breast cancer: towards the next phase. Cell Cycle. 2018;17(15):1871–1885. doi:10.1080/15384101.2018.150256730078354
- The Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumors. Nature. 2012;490(7418):61–70. doi:10.1038/nature1141223000897