
Abstract
Evidence indicates that for most human cancers the problem is not that gene mutations occur but is more dependent upon how the body deals with damaged cells. It has been estimated that only about 1% of human cancers can be accounted for by unmistakable hereditary cancer syndromes, only up to 5% can be accounted for due to high-penetrance, single-gene mutations, and in total only 5%–15% of all cancers may have a major genetic component. The predominant contribution to the causation of most sporadic cancers is considered to be environmental factors contributing between 58% and 82% toward different cancers. A nutritionally poor lifestyle is associated with increased risk of many cancers, including those of the breast. As nutrition, energy balance, macronutrient composition of the diet, and physical activity levels are major determinants of insulin-like growth factor (IGF-I) bioactivity, it has been proposed that, at least in part, these increases in cancer risk and progression may be mediated by alterations in the IGF axis, related to nutritional lifestyle. Localized breast cancer is a manageable disease, and death from breast cancer predominantly occurs due to the development of metastatic disease as treatment becomes more complicated with poorer outcomes. In recent years, epithelial-to-mesenchymal transition has emerged as an important contributor to breast cancer progression and malignant transformation resulting in tumor cells with increased potential for migration and invasion. Furthermore, accumulating evidence suggests a strong link between components of the IGF pathway, epithelial-to-mesenchymal transition, and breast cancer mortality. Here, we highlight some recent studies highlighting the relationship between IGFs, IGF-binding protein 3, and epithelial-to-mesenchymal transition.
Keywords:
Insulin-like growth factor axis
The activity of insulin-like growth factors (IGFs) within any tissue is due to a combination of some locally expressed components together with IGFs and IGF-binding proteins (IGFBPs) delivered to the tissue from the circulation where high levels are maintained. This provides a mechanism for integrating systemic and local regulation systems. The circulating IGF system is under the influence of growth hormones, insulin, nutrition and systemic disease status, such as cancer, whereas the locally expressed components are controlled by factors specific to each individual tissue.
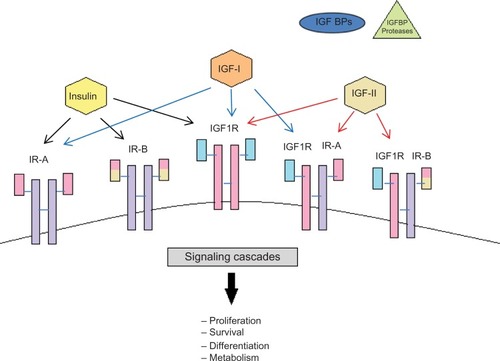
The cellular effects of the IGFs are mediated by a number of cell surface receptors including the type I and type II receptors, insulin receptor (IR), and IR–IGF-I receptor hybrids. Both IGF-I and IGF-II can mediate their actions on cell growth and survival via the IGF-IR, which is a transmembrane, tyrosine kinase that is structurally and functionally homologous to the IR. IGF-I can act via the IR but only at supraphysiological doses.Citation1 IGF-II can bind with high affinity to the IGF-II receptor/mannose-6-phosphate receptor (IGF-IIR), a non-tyrosine kinase receptor considered to play an important role in the clearance and degradation of IGF-II. Additionally, the IGF-IIR is critically involved in the cellular trafficking of lysosomal enzymes and also acts as a high-affinity binding site for latent transforming growth factor-beta (TGFβ)-I, II, and III and retinoids.Citation2 It was originally believed that IGF-II did not bind to the IR with any meaningful affinity until mice knockout studies revealed that IGF-II could act via this receptor during development.Citation3 There are two alternatively spliced isoforms of the IR, A and B, and it has further been demonstrated that IGF-II binds with high affinity to the IRA but not the IRB isoform.Citation4 The IR and IGF-IR can dimerize forming hybrid receptorsCitation5 and each IR isoform is equally able to form hybrids with the IGF-IR. Hybrid IGF-IR/IRB receptors have a high affinity for IGF-I and a lower affinity for insulin, whereas hybrid IGF-IR/IRA receptors not only have an even higher affinity for IGF-I but also bind IGF-II and insulinCitation6 (). Activation of the IGF-IR by IGFs results in oligomerization, autophosphorylation, and activation of the intrinsic tyrosine kinase.Citation1 The IGF-IR tyrosine kinase directly phosphorylates a number of intracellular substrates including IR substrates (IRS)-1, 2, and 4,Citation7–Citation9 src homology domain containing (Shc),Citation10,Citation11 phosphatidylinositol-3 kinase (PI3-K),Citation12 growth factor receptor–bound protein 10,Citation13 focal adhesion kinase,Citation14 and C-Src kinase.Citation15
Figure 1 Schematic representation of the insulin-like growth factor system.
Abbreviations: IGF, insulin-like growth factor; IGFBP, insulin-like growth factor–binding protein; IR, insulin receptor.
There are six high-affinity IGFBPs 1–6 that all have greater affinity for binding to the IGFs, than the IGF-IR, and can modulate IGF actions in many cell types. The IGFBPs slow the clearance of the IGFs, particularly IGFBP-3 and IGFBP-5, which also bind to a glycoprotein called the acid labile subunit forming ternary complexes, which have very long half-lives in the circulation. It is these complexes that maintain very high concentrations of IGFs in the body. Accumulating evidence indicates that most of the IGFBPs can also act in an intrinsic manner, independent of IGF binding, affecting various aspects of cell function including growth, apoptosis, migration, and attachment.Citation16–Citation18 The nature and complexity of the IGF axis during development and in disease states have been defined through the use of advanced molecular techniques, transgenic and knockout mouse models, which are beyond the scope of this review but have been elegantly summarized elsewhere.Citation19–Citation22
Metabolic disturbance and breast cancer
In all Western societies, women who present with breast cancer are increasingly likely to also suffer from comorbid conditions such as diabetes and obesity due to the very high prevalence of these conditions in the general population. In a study of over 1000 women treated for breast cancer at the MD Anderson Cancer Center in Houston, 30% were found to be obese and a further 32% were overweight.Citation23 In addition, the prevalence of metabolic syndrome in patients with breast cancer has been reported to be up to 50%.Citation24,Citation25
Evidence derived from epidemiology, in vitro and in vivo studies suggest that metabolic disturbances adversely affect breast cancer cell survival and progression. In women with breast cancer, having metabolic syndrome also was associated with more aggressive tumor characteristicsCitation24 and being obese was associated with worse overall survival.Citation23 In an orthotopic syngeneic mouse model of triple-negative breast cancer, a high-energy diet promoted tumor growth and increased the number of metastases to the lungs.Citation26 In vitro, it has been shown that breast cancer cells exposed to high levels of glucose, such as those associated with type 2 diabetes causes resistance to chemotherapy.Citation27 As IGF is metabolically regulated and mediates the effects of nutrition on cell growth,Citation28 it is not surprising that evidence indicates that IGF-I plays a critical role in also affecting the impact of altered metabolism on tumor growth and survival. Calorie restriction has long been known to reduce obesity, improve metabolic status, and reduce the propensity for cancer development.Citation29 Calorie restriction studies using the 4T1 mammary tumor model showed a reduction in tumor growth and metastasis that was associated with decreased circulating IGF-I levels.Citation30 Furthermore, using orthotopically transplanted mammary tumors in control and calorie-restricted mice, it was concluded that a reduction in mammary tumor growth could be accounted for in part by reduced levels of IGF-I.Citation31
Most cancer cells have an increased ability to enhance their glucose uptake and rely on aerobic glycolysis, a phenomenon termed “the Warburg effect” in order to generate energy, in contrast to normal differentiated cells, which use mitochondrial oxidative phosphorylation to generate the energy needed for cellular processes. Such strategies employed by cancer cells to ensure progression also have an impact on other aspects of tumorigenesis: artificially stimulating the Warburg effect in animal models and in cell culture has been found to induce epithelial-to-mesenchymal transition (EMT). For example, genetic silencing of the respiratory enzyme citrate synthase, resulted in greatly increased glycolytic metabolism that was reported to induce EMT and increase metastases in a mouse xenograft model.Citation32 In addition, reversal of the Warburg effect by silencing of expression of pyruvate dehydrogenase kinase 1 in breast cancer cells restored their sensitivity to anoikis when detached and reduced lung metastasis when they were inoculated in a mouse model.Citation33 It has been reported that developmental EMT can be induced by exposure to hyperglycemia, and it has recently been demonstrated that exposure to hyperglycemia can similarly induce EMT of human lung adenocarcinoma cells.Citation34
EMT in breast cancer progression and metastasis
Metastasis is the spread of cancer cells to distant sites in the body. It is the ultimate cause of death in more than 90% of breast cancer patients and therefore represents one of the biggest challenges in cancer research. In about 6%–10% of breast cancer diagnoses, cancer has already metastasized to the other parts of the body and approximately 30% of patients with early-stage breast cancer have a recurrent or metastatic disease.Citation35
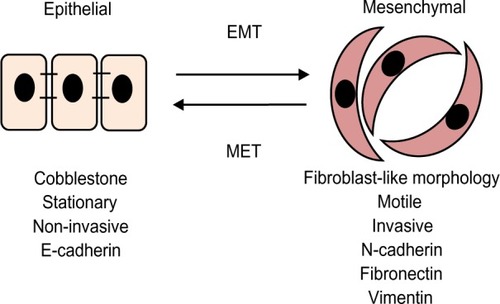
The seeding and growth of breast cancer cells at sites distinct from the primary tumor is a complex and multistage process that involves the ability to detach and invade through the basement membrane, intravasation into blood vessels or lymphatics, ability to transit through the vasculature, and extravasation from the circulation to establish new secondary tumors typically in organs such as bone, lung, liver, or brain (). Emerging evidence suggests that in the initial stage, in the so-called invasion–metastasis cascade, cancer cells undergo a dramatic phenotypic change from an epithelial-to-mesenchymal phenotype via a process referred to as EMT. The plasticity of the epithelial phenotype is illustrated by the occurrence of the reverse process, mesenchymal-to-epithelial transition, during which cancer cells regain the epithelial properties of the primary tumor to promote outgrowth at metastatic sites. The EMT process was originally identified as being essential for embryonic development, mesoderm and neural crest formation, and also during wound healing.Citation36 It is, however, now increasingly clear that inappropriate activation of EMT is a critical component of the progression of many cancers including breast.Citation37 Cellular switching from an epithelial into mesenchymal cell requires a large number of complex modifications that include loss of cell adhesion, phenotypic change from typical cuboidal to an elongated spindle shape, rearrangement in the cytoskeletal architecture, and adoption of migratory and invasive phenotype (). Epithelial cells are typically arranged as sheets of cells exhibiting apical–basal polarity. These cells are held together through a variety of structures including adherent junctions, tight junctions, and desmosomes and are separated from the underlying tissues by a thin layer of specialized extracellular matrix called the basement membrane, all of which help the epithelium to fulfill its function as a protective barrier. Mesenchymal cells on the other hand form disorganized cellular layers that are neither polarized nor tightly adherent, which allows for increased migratory capacity. They provide support and structure to the epithelial cells through the production of extracellular matrix components. A defining feature of EMT is a loss of epithelial cell adhesion molecule E-cadherin and a gain of mesenchymal cell markers such as vimentin, N-cadherin, and fibronectin.Citation38
Figure 2 Schematic outline of breast cancer progression.
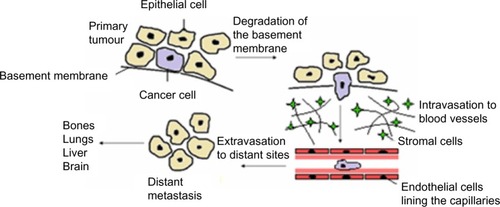
Figure 3 Epithelial-to-mesenchymal transitions.
Abbreviations: EMT, epithelial-to-mesenchymal transition; MET, mesenchymal-to-epithelial transition.
A number of studies have shown a strong correlation between the EMT and high invasive and metastatic behavior of breast cancer. Particularly interesting is the recent finding that the EMT process appears to be associated with a basal-like breast cancer phenotype, which tends to be more aggressive and carry a poor prognosis.Citation39 In the clinical setting, studies indicate that circulating tumor cells (CTCs) in patients with metastatic breast cancer frequently express mesenchymal markers, whereas mesenchymal markers are only found in rare cells within the corresponding primary tumours.Citation40 In breast cancer, CD24−CD44+ cells or cells with high aldehyde dehydrogenase activityCitation41 have been shown to be enriched in breast cancer stem cells. Intriguingly, it has also been shown that a major proportion of CTCs found in the blood samples of breast cancer patients also express the stem cell marker aldehyde dehydrogenase 1Citation42. A small subset of cells have been prospectively identified and characterized in the primary tumor that exhibited the exclusive capacity to form colonies, using a colony formation assay, and to form tumors in mice; these cells exhibited a stem cell phenotype.Citation43 Abraham identified that breast cancer stem cells only accounted for 10%–20% of the cell population in a primary tumourCitation44 that supports the concept of an exclusive subset of cells in the primary tumor with stem cell properties that have the propensity to metastasize. These findings support the notion that EMT is associated with “stemness” properties and that activation of EMT is critical for cancer cells to disseminate.
EMT is triggered and orchestrated by several growth factors released by the stromal cells including TGFβ, hepatocyte growth factor, platelet-derived growth factor, fibroblast growth factor, and Wnt and Notch ligands. Among these, TGFβ is undoubtedly the most potent driver of EMT that elicits its actions through activation of Smad and non-Smad signaling pathways.Citation45 In fact, simple treatment with exogenous TGFβ can induce EMT in various epithelial cells. TGFβ plays a dual role in cancer: it is tumor suppressive during the early stages of tumor growth, but paradoxically, it becomes pro-oncogenic and promotes EMT and metastasis during late stages of malignancy.Citation46 This switch in TGFβ function during advanced breast cancer is particularly reflected in increased expression of phosphorylated Smad2 in breast tumours.Citation47 TGFβ activates a series of EMT-inducing transcription factors including Snail, Slug, Twist, Zeb1, Zeb2, and FoxC2, which in turn are under the control of multiple micro-RNAs.Citation48–Citation51 Most attention has been directed at the study of Snail. This zinc finger transcription factor was first noted during mesoderm formation in Drosophila melanogaster and was subsequently found to be a potent repressor of E-cadherin and a major EMT inducer.Citation52,Citation53 It is known that Snail binds to E-boxes in the proximal promoter region of E-cadherin and by recruiting histone deacetylases represses its expression.Citation53 The significance of Snail in EMT has been shown in a variety of in vitro and in vivo models. It has been shown that Snail is required for tumor growth and lymph node metastasis of human breast carcinoma MDA–MB-231 cells and that knockdown of Snail increases the chemosensitivity of these cells to gemcitabine and docetaxel.Citation54 Similarly using a mammary-specific, inducible HER2/Neu transgenic mouse model, Moody et al found high expression of Snail in recurrent breast tumours.Citation55 Snail also interacts with other transcription factors as exemplified by a recent study, which shows that Snail cooperates with Twist to induce Zeb1 expression during EMT.Citation56 It has also been reported that Snail co-immunoprecipitates with a Smad3/4 complex to promote TGFβ-mediated repression of E-cadherin, occludin, and coxsackievirus and adenovirus receptor and thereby cause EMT.Citation57
In addition, an increasing number of studies demonstrate that EMT not only facilitates metastasis but also contributes to drug resistance. Using several drug-resistant sublines of MCF-7, Iseri et al reported an increase in the expression of EMT-associated genes (Slug, N-cadherin, and vimentin) together with loss of E-cadherin and the estrogen receptor (ER) α.Citation58
IGF axis, EMT, and breast cancer
The conclusions drawn from a recent case control study nested within The European Prospective Investigation into Cancer and Nutrition cohort confirmed that a wealth of previous epidemiology studies that indicated higher levels of IGF-I within the normal range increase the risk of estrogen receptor–positive (ER+ve) breast cancers in pre- and postmenopausal women but are not related to the risk of developing ER-negative (ER−ve) tumors.Citation59–Citation62 Such a relationship of IGF-I with ER+ve tumors is consistent with the vast amount of literature indicating the synergy that is known to exist between IGF and estrogen-induced signaling pathways in promoting breast cancer progression (see Hawsawi et alCitation63 and Tsonis et alCitation64 for reviews).
For the purposes of this review, we are focusing on the role of IGF-I/II and IGFBP-3 in EMT for which the majority of evidence has been obtained: in terms of circulating IGFBP-3, there has been both positive and negative associations of IGFBP-3 with the risk of breast cancer,Citation65–Citation67 but when RoddamCitation62 pooled 17 prospective trials, he concluded that the risk of breast cancer was not associated with circulating levels of IGFBP-3. These data suggest that for IGFBP-3, either circulating levels do not play a role or they are not reflective of the local situation in the tumor itself.
It is becoming increasingly clear that IGFs are important for inducing EMT by enhancing growth, survival, motility, migration, and metastasis of breast cancer cells. IGF-I and IGF-II are expressed in many tumors and IGF-II in particular has been observed to be overexpressed in a number of different tumors, including the breast,Citation68 and increased expression of either IGF-ICitation69 or IGF-IICitation70–Citation72 has been reported to be associated with more aggressive tumor phenotypes. Similarly increased expression of IGF-II and the IGF-IR has been associated with a more metastatic phenotype.Citation73 Increased expression of the IGF-IR has been reported in many cancer cell lines and in human tumor biopsies.Citation74,Citation75 The IGF-IR appears to play a critical role in malignant transformation and in the maintenance of a transformed cell phenotypeCitation76 due to its ability to maintain cell survival and protect cells from apoptosis via multiple signaling pathways.Citation77 In human mammary epithelial cells, overexpression of a constitutively active IGF-IR caused their transformation and growth in immunocompromised mice that was associated with the initiation of EMT, via upregulation of Snail and downregulation of E-cadherin.Citation78 In keeping with this, overexpression of the IGF-IR has been reported to be associated with an aggressive phenotype of a variety of tumours.Citation79,Citation80 An in vivo experimental brain metastasis model demonstrated that IGF-IR silencing in brain-seeking breast cancer cells reduced the propensity of these cells to develop brain metastases.Citation81
As mentioned previously, EMT has been associated with features of “stemness”, and recently, Chang et alCitation82 reported that expression of activated IGF-IR was greater in cancer stem cells than normal stem cells. They also reported that those cells expressing high levels of activated IGF-IR exhibited factors associated with EMT that were lost upon silencing or inhibiting the IGF-IR. Even in normal breast epithelial cells, MCF-10A, IGF-I was able to induce EMT when Akt1 was downregulated but expression of Akt2 was maintained.Citation83 To become motile, cells have to depolarize and work has indicated that IGF-I depolarizes breast cancer cells in a phosphatidylinositol-3 kinase–dependent manner.Citation84 MEMO is a protein recognized as being a critical mediator of motility in response to receptor tyrosine kinase activation and can specifically affect IGF-IR–dependent signaling in breast cancer cell lines, via binding to IR substrate-1 and activating phosphatidylinositol-3 kinase/Akt signaling, which leads to upregulation of the transcription factor, Snail, that is key to effecting EMT.Citation85 IGF-I has also been shown to upregulate Zeb1, another key transcription factor involved in EMT, to induce invasion of MDA–MB-231 cells.Citation86 The ability of IGF-I to drive EMT in another hormone-responsive cancer, the prostate, is also dependent upon its ability to upregulate Zeb-1.Citation87 In order for breast cancer cells to metastasize, they have to travel in the circulation to distant sites, and it has recently been shown that the IGF-IR is present on CTCs of breast cancer patients.Citation88 High circulating IGF-I has been correlated with breast cancer mortalityCitation89 and lymph node metastasis in patients with ER+ve disease.Citation90 These data suggest that one way in which serum IGF-I may influence metastasis may be by activating the IGF-IR present on CTCs and maintaining survival of these detached cells in the circulation. Although not in breast cancer cells, a complementary DNA microarray was employed to assess gene expression patterns specifically regulated by IGF-I, differential over those regulated by insulin and found that a key transcription factor upregulated in EMT, Twist, was one of the most upregulated genes and that Twist was key in mediating the antiapoptotic effects of IGF-I in NWTb3 fibroblast cells.Citation91,Citation92 In addition to the IGF-IR, the IR, particularly the IRA isoform, that has a higher affinity for IGF-II similar to that for insulin, together with the hybrid IR/IGF-IR receptors may also be present and play a role in mediating the actions of the IGFs in certain tumors.Citation93,Citation94 IR isoform assessment from formalin-fixed, paraffin-embedded sections, showed that the IRA was more abundant than the IRB isoform in breast tumours.Citation95 In contrast to the IGF-I receptor, the IGF-IIR serves to limit IGF-II actions and hence reduce its growth-promoting and cell survival potential. A number of genetic disruptions resulting in loss of IGF-IIR have been described in various tumor types including missense mutations, loss of heterozygosity, and microsatellite instability.Citation96,Citation97 Loss of the IGF-IIR has been associated with increased tumor growth potential,Citation98 decreasing IGF-IIR expression has a similar effect,Citation99 whereas introduction of the IGF-IIR into cancer cells reduces growth and increases apoptosis.Citation100
The IGFBPs have the potential to either inhibit or enhance IGF actions in many cell types. Most evidence suggests that IGFBPs generally restrict tumor growth and progression by limiting IGF-mitogenic and cell survival actions. The actions of many antiproliferative agents appear to operate, at least in part, via upregulation of endogenous IGFBPs produced by the tumor cells, including TGFβ,Citation101 retinoids,Citation102 vitamin D,Citation103 tamoxifen,Citation104 and butyrate.Citation105
In vitro studies suggest that the more aggressive ER−ve, compared with ER+ve, cell lines secrete higher levels of IGFBP-3.Citation106 These reports are consistent with the observation that higher concentrations of IGFBP-3 in breast tissue are associated with increased risk of mortality andCitation107 are correlated with tumor size and poor prognostic characteristics.Citation108,Citation109 Accumulating evidence indicates that most of the IGFBPs can also act in an intrinsic manner, independent of IGF binding, affecting various aspects of cell function. Growth inhibition and modulation of apoptosis have been described in a variety of cancer cell lines.Citation17,Citation18,Citation110–Citation112
Studies conducted in our laboratory and others have demonstrated that rhIGFBP-3 enhances the cell cycle arrest and apoptotic effects of paclitaxel and radiation in human breast cancer cells in vitro.Citation110,Citation113 In addition, our studies suggest that when normal breast epithelial cells are challenged with exogenous ceramide, rhIGFBP-3 has the opposite effect and acts as a survival factor.Citation114 In preliminary in vivo studies, subcutaneous administration of rhIGFBP-3 to mice bearing MCF-7 breast tumor xenografts enhanced the tumor growth inhibition effect of paclitaxel from 33% to 61%.Citation115 However, in vitro studies clearly indicate that the ability of IGFBP-3 to intrinsically affect breast cancer cell growth, survival, and adhesion is dependent upon the cellular environment. In conditions that reflect a more advanced tumor environment, such as increased fibronectin, IGFBP-3 acts in an opposite manner and promotes tumor progression by acting as a survival factor and growth promoter, whereas in early stages of the disease, IGFBP-3 enhances triggers of apoptosis and inhibits cell growth thus acting as a tumor suppressor.Citation16,Citation116,Citation117 The association of high local expression of IGFBP-3 with poor prognosis could be explained by the above observations that the actions of IGFBP-3 on cancer cells can be switched when the cells are exposed to a more advanced tumor environment such as increased fibronectin.Citation118 There are a number of theories as to the mechanisms underlying the intrinsic actions of IGFBP-3 and these have been extensively reviewed recently (refer BaxterCitation119 and Johnson and FirthCitation120).
TGFβ clearly plays a significant role in breast cancer progression, and it has been shown that IGF-I can activate latent TGFβ in breast cancer cells.Citation121 Like IGFBP-3, TGFβ acts as a tumor suppressor in the early stages, but in later stages of the disease, it switches function and acts to promote tumorigenesis. TGFβ can differentially regulate IGFBP-3 depending on the cell type. With nonmalignant breast epithelial cells, where IGFBP-3 promotes growth and TGFβ is a growth inhibitor, TGFβ was found to downregulate IGFBP-3. Whereas in breast cancer cells, where TGFβ and IGFBP-3 both act as growth inhibitors, TGFβ was found to upregulate IGFBP-3.Citation114 It is well documented that in the later stages of disease, TGFβ enhances the production of extracellular matrix production, such as fibronectinCitation122 as part of its role in inducing EMT. Having shown that IGFBP-3 can act as a tumor promoter in the presence of fibronectin, it would be interesting to see whether TGFβ-induced IGFBP-3 in this context could mediate the pro-tumorigenic actions of TGFβ to induce EMT.
Breast cancer treatment: is the IGF-IR a viable target?
Breast cancer can be treated with different combinations of therapies: surgery, radiation therapy, chemotherapy, and hormone therapy. This depends upon the classification of the tumor and whether it remains localized in the breast or has spread to other sites in the body. Tumors can be classified histologically, of which 21 distinct histological types have been identified.Citation123 Through the use of molecular profiling, breast cancers are now also categorized into four main subtypes: basal-like, HER2+, Luminal A, or Luminal B. Other less common molecular subtypes have also been described including normal breast-like, apocrine molecular type, and claudin-low type. Breast cancers that do not fall into any of these subtypes are often listed as unclassified.Citation124
Rigorous laboratory and preclinical research identified that the IGF-IR played a key role in the progression of a number of different cancers, including the breast and as a result, a number of agents targeting the IGF-IR were developed and taken forward into clinical trials: these included antibodies against the receptor itself, against the receptor ligands, IGF-I and II, in addition to IGF-IR tyrosine kinase inhibitors (see Refs Citation125–Citation127 for reviews). Despite encouraging preliminary data, the advanced clinical trials failed to show clinical benefit, which culminated in most drug companies terminating current programs targeting the IGF-IR. These trials were undertaken on unselected patients and no other targeted therapy has yet been found to work generally in cohorts of unselected patients. The results of the trials so far suggest that perhaps this receptor will only be an effective target in a subset of patients and to identify them will require a greater understanding of the context in terms of the complement of receptors present: levels of IRA and hybrid receptors and also the cooperative signaling events initiated by receptor signaling.
There have also been suggestions that IGFBP-3 could be developed as a cancer therapeutic due to its ability to inhibit IGF-actions and its IGF-independent pro-apoptotic actions. Although preclinical studies have provided some promising evidence,Citation128 these have not progressed to clinical trials. This is partly due to not only pharmaco-formulation issues but also the increasingly conflicting clinical observations of both positive and negative effects of IGFBP-3, which have added caution to its development as a clinical intervention.
Summary
EMT is clearly important in effecting cancer progression; a better characterization of the role of the IGF axis in this process may lead to a greater understanding of its activity and co-operativity with other signaling pathways that are involved. This may lead to more effective interventions for metastatic disease and help to determine what causes the emergence of resistance to current therapeutic agents and provide potential critical biomarkers of response.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
We thank the “Aiming to Beat Cancer Fund”, Bristol Oncology Centre, for supporting our work.
References
- LeRoithDWernerHBeitner-JohnsonDRobertsCTJrMolecular and cellular aspects of the insulin-like growth factor I receptorEndocr Rev1995161431637540132
- BraulkeTType-2 IGF receptor: a multi-ligand binding proteinHorm Metab Res19993124224610226808
- RotherKIAcciliDRole of insulin receptors and IGF receptors in growth and developmentPediatr Nephrol200014755856110912518
- FrascaFPandiniGScaliaPInsulin receptor isoform A, a newly recognized, high-affinity insulin-like growth factor II receptor in fetal and cancer cellsMol Cell Biol1999193278328810207053
- SeelyBLReichartDRTakataYYipCOlefskyJMA functional assessment of insulin/insulin-like growth factor-I hybrid receptorsEndocrinology1995136163516417895674
- PandiniGFrascaFMineoRSciaccaLVigneriRBelfioreAInsulin/insulin-like growth factor I hybrid receptors have different biological characteristics depending on the insulin receptor isoform involvedJ Biol Chem2002277396843969512138094
- MyersMGJrGrammerTCWangLMInsulin receptor substrate-1 mediates phosphatidylinositol 3′-kinase and p70S6k signaling during insulin, insulin-like growth factor-1, and interleukin-4 stimulationJ Biol Chem199426928783287897961833
- HeWCraparoAZhuYInteraction of insulin receptor substrate-2 (IRS-2) with the insulin and insulin-like growth factor I receptors. Evidence for two distinct phosphotyrosine-dependent interaction domains within IRS-2J Biol Chem199627111641116458662806
- FantinVRSparlingJDSlotJWKellerSRLienhardGELavanBECharacterization of insulin receptor substrate 4 in human embryonic kidney 293 cellsJ Biol Chem199827310726107329553137
- PelicciGLanfranconeLGrignaniFA novel transforming protein (SHC) with an SH2 domain is implicated in mitogenic signal transductionCell199270931041623525
- GiorgettiSPelicciPGPelicciGVan ObberghenEInvolvement of Src-homology/collagen (SHC) proteins in signaling through the insulin receptor and the insulin-like-growth-factor-I-receptorEur J Biochem19942231952028033892
- LamotheBBucchiniDJamiJJoshiRLInteraction of p85 subunit of PI 3-kinase with insulin and IGF-1 receptors analysed by using the two-hybrid systemFEBS Lett199537351557589433
- MorrioneAValentinisBLiSOoiJYMargolisBBasergaRGrb10: A new substrate of the insulin-like growth factor I receptorCancer Res199656316531678764099
- BaronVCallejaVFerrariPAlengrinFVan ObberghenEp125Fak focal adhesion kinase is a substrate for the insulin and insulin-like growth factor-I tyrosine kinase receptorsJ Biol Chem1998273716271689507031
- Arbet-EngelsCTartare-DeckertSEckhartWC-terminal Src kinase associates with ligand-stimulated insulin-like growth factor-I receptorJ Biol Chem19992745422542810026153
- McCaigCPerksCMHollyJMIntrinsic actions of IGFBP-3 and IGFBP-5 on Hs578T breast cancer epithelial cells: inhibition or accentuation of attachment and survival is dependent upon the presence of fibronectinJ Cell Sci20021154293430312376561
- PerksCMBowenSGillZPNewcombPVHollyJMDifferential IGF-independent effects of insulin-like growth factor binding proteins (1–6) on apoptosis of breast epithelial cellsJ Cell Biochem19997565266410572248
- OhYMüllerHLLamsonGRosenfeldRGInsulin-like growth factor (IGF)-independent action of IGF-binding protein-3 in Hs578T human breast cancer cells. Cell surface binding and growth inhibitionJ Biol Chem199326814964149717686909
- DupontJHolzenbergerMBiology of insulin-like growth factors in developmentBirth Defects Res C Embryo Today20036925727114745968
- YakarSSunHZhaoHMetabolic effects of IGF-I deficiency: lessons from mouse modelsPediatr Endocrinol Rev20053111916369209
- SilhaJVMurphyLJInsights from insulin-like growth factor binding protein transgenic miceEndocrinology20021433711371412239079
- HollyJMPerksCMInsulin-like growth factor physiology: what we have learned from human studiesEndocrinol Metab Clin North Am201241249263v22682629
- LittonJKGonzalez-AnguloAMWarnekeCLRelationship between obesity and pathologic response to neoadjuvant chemotherapy among women with operable breast cancerJ Clin Oncol2008264072407718757321
- HealyLARyanAMCarrollPMetabolic syndrome, central obesity and insulin resistance are associated with adverse pathological features in postmenopausal breast cancerClin Oncol (R Coll Radiol)20102228128820189371
- StebbingJSharmaANorthBA metabolic phenotyping approach to understanding relationships between metabolic syndrome and breast tumour responses to chemotherapyAnn Oncol20122386086621821546
- PhoenixKNVumbacaFFoxMMEvansRClaffeyKPDietary energy availability affects primary and metastatic breast cancer and metformin efficacyBreast Cancer Res Treat201012333334420204498
- ZengLBiernackaKMHollyJMHyperglycaemia confers resistance to chemotherapy on breast cancer cells: the role of fatty acid synthaseEndocr Relat Cancer20101753955120356977
- NarasimhanSDYenKTissenbaumHAConverging pathways in lifespan regulationCurr Biol200919R657R66619674551
- HurstingSDSmithSMLashingerLMHarveyAEPerkinsSNCalories and carcinogenesis: lessons learned from 30 years of calorie restriction researchCarcinogenesis201031838919969554
- De LorenzoMSBaljinnyamEVatnerDEAbarzuaPVatnerSFRabsonABCaloric restriction reduces growth of mammary tumors and metastasesCarcinogenesis2011321381138721665891
- NogueiraLMLavigneJAChandramouliGVLuiHBarrettJCHurstingSDDose-dependent effects of calorie restriction on gene expression, metabolism, and tumor progression are partially mediated by insulin-like growth factor-1Cancer Med2012127528823342276
- LinCCChengTLTsaiWHLoss of the respiratory enzyme citrate synthase directly links the Warburg effect to tumor malignancySci Rep2012278523139858
- KamarajugaddaSStemboroskiLCaiQGlucose oxidation modulates anoikis and tumor metastasisMol Cell Biol2012321893190722431524
- Alisson-SilvaFFreire-de-LimaLDonadioJLIncrease of O-glycosylated oncofetal fibronectin in high glucose-induced epithelial-mesenchymal transition of cultured human epithelial cellsPLoS One20138e6047123593224
- O’ShaughnessyJExtending survival with chemotherapy in metastatic breast cancerOncologist200510Suppl 3202916368868
- KalluriRWeinbergRAThe basics of epithelial-mesenchymal transitionJ Clin Invest20091191420142819487818
- ThieryJPEpithelial-mesenchymal transitions in tumour progressionNat Rev Cancer2002244245412189386
- ZeisbergMNeilsonEGBiomarkers for epithelial-mesenchymal transitionsJ Clin Invest20091191429143719487819
- SarrioDRodriguez-PinillaSMHardissonDCanoAMoreno-BuenoGPalaciosJEpithelial-mesenchymal transition in breast cancer relates to the basal-like phenotypeCancer Res20086898999718281472
- YuMBardiaAWittnerBSCirculating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal compositionScience201333958058423372014
- AktasBTewesMFehmTHauchSKimmigRKasimir-BauerSStem cell and epithelial-mesenchymal transition markers are frequently overexpressed in circulating tumor cells of metastatic breast cancer patientsBreast Cancer Res200911R4619589136
- ManiSAGuoWLiaoMJThe epithelial-mesenchymal transition generates cells with properties of stem cellsCell200813370471518485877
- Al-HajjMWichaMSBenito-HernandezAMorrisonSJClarkeMFProspective identification of tumorigenic breast cancer cellsProc Natl Acad Sci U S A20031003983398812629218
- AbrahamBKFritzPMcClellanMHauptvogelPAthelogouMBrauchHPrevalence of CD44+/CD24−/low cells in breast cancer may not be associated with clinical outcome but may favor distant metastasisClin Cancer Res2005111154115915709183
- MiyazonoKTransforming growth factor-beta signaling in epithelial-mesenchymal transition and progression of cancerProc Jpn Acad Ser B Phys Biol Sci200985314323
- BachmanKEParkBHDuel nature of TGF-beta signaling: tumor suppressor vs tumor promoterCurr Opin Oncol200517495415608513
- XieWMertensJCReissDJAlterations of Smad signaling in human breast carcinoma are associated with poor outcome: a tissue microarray studyCancer Res20026249750511809701
- FendrichVWaldmannJFeldmannGUnique expression pattern of the EMT markers Snail, Twist and E-cadherin in benign and malignant parathyroid neoplasiaEur J Endocrinol200916069570319176646
- FangXCaiYLiuJTwist2 contributes to breast cancer progression by promoting an epithelial-mesenchymal transition and cancer stem-like cell self-renewalOncogene2011304707472021602879
- ManiSAYangJBrooksMMesenchyme Forkhead 1 (FOXC2) plays a key role in metastasis and is associated with aggressive basal-like breast cancersProc Natl Acad Sci U S A2007104100691007417537911
- GuoFParker KerriganBCYangDPost-transcriptional regulatory network of epithelial-to-mesenchymal and mesenchymal-to-epithelial transitionsJ Hematol Oncol201471924598126
- AlbergaABoulayJLKempeEDennefeldCHaenlinMThe snail gene required for mesoderm formation in Drosophila is expressed dynamically in derivatives of all three germ layersDevelopment19911119839921879366
- PeinadoHBallestarEEstellerMCanoASnail mediates E-cadherin repression by the recruitment of the Sin3A/histone deacetylase 1 (HDAC1)/HDAC2 complexMol Cell Biol20042430631914673164
- OlmedaDMoreno-BuenoGFloresJMFabraAPortilloFCanoASNAI1 is required for tumor growth and lymph node metastasis of human breast carcinoma MDA-MB-231 cellsCancer Res200767117211173118089802
- MoodySEPerezDPanTCThe transcriptional repressor Snail promotes mammary tumor recurrenceCancer Cell2005819720916169465
- DaveNGuaita-EsteruelasSGutarraSFunctional cooperation between Snail1 and twist in the regulation of ZEB1 expression during epithelial to mesenchymal transitionJ Biol Chem2011286120241203221317430
- VincentTNeveEPJohnsonJRA SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transitionNat Cell Biol20091194395019597490
- IseriODKarsMDArpaciFAtalayCPakIGunduzUDrug resistant MCF-7 cells exhibit epithelial-mesenchymal transition gene expression patternBiomed Pharmacother201165404521177063
- KaaksRJohnsonTTikkKInsulin-like growth factor I and risk of breast cancer by age and hormone receptor status-A prospective study within the EPIC cohortInt J Cancer20141342683269024248481
- SchernhammerESHollyJMHunterDJPollakMNHankinsonSEInsulin-like growth factor-I, its binding proteins (IGFBP-1 and IGFBP-3), and growth hormone and breast cancer risk in The Nurses Health Study IIEndocr Relat Cancer20061358359216728584
- BagliettoLEnglishDRHopperJLMorrisHATilleyWDGilesGGCirculating insulin-like growth factor-I and binding protein-3 and the risk of breast cancerCancer Epidemiol Biomarkers Prev20071676376817416768
- Endogenous Hormones and Breast Cancer Collaborative GroupKeyTJApplebyPNReevesGKRoddamAWInsulin-like growth factor 1 (IGF1), IGF binding protein 3 (IGFBP3), and breast cancer risk: pooled individual data analysis of 17 prospective studiesLancet Oncol20101153054220472501
- HawsawiYEl-GendyRTwelvesCSpeirsVBeattieJInsulin-like growth factor – oestradiol crosstalk and mammary gland tumourigenesisBiochim Biophys Acta2013183634535324189571
- TsonisAIAfratisNGialeliCEvaluation of the coordinated actions of estrogen receptors with epidermal growth factor receptor and insulin-like growth factor receptor in the expression of cell surface heparan sulfate proteoglycans and cell motility in breast cancer cellsFEBS J20132802248225923374155
- Keinan-BokerLBueno De MesquitaHBKaaksRCirculating levels of insulin-like growth factor I, its binding proteins-1, -2, -3, C-peptide and risk of postmenopausal breast cancerInt J Cancer2003106909512794762
- GronbaekHFlyvbjergAMellemkjaerLSerum insulin-like growth factors, insulin-like growth factor binding proteins, and breast cancer risk in postmenopausal womenCancer Epidemiol Biomarkers Prev2004131759176415533904
- RenehanAGZwahlenMMinderCO’DwyerSTShaletSMEggerMInsulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysisLancet20043631346135315110491
- LivingstoneCIGF2 and cancerEndocr Relat Cancer201320R321R33924080445
- HiranoHLopesMBLawsERJrInsulin-like growth factor-1 content and pattern of expression correlates with histopathologic grade in diffusely infiltrating astrocytomasNeuro-Oncology1999110911911550306
- SohdaTOkaYIwataKCo-localisation of insulin-like growth factor II and the proliferation marker MIB1 in hepatocellular carcinoma cellsJ Clin Pathol1997501351379155694
- KawamotoKOnoderaHKondoSExpression of insulin-like growth factor-2 can predict the prognosis of human colorectal cancer patients: correlation with tumor progression, proliferative activity and survivalOncology1998552422489560057
- RoglerCEYangDRossettiLAltered body composition and increased frequency of diverse malignancies in insulin-like growth factor-II transgenic miceJ Biol Chem199426913779137847514593
- GuerraFKEijanAMPuricelliLVarying patterns of expression of insulin-like growth factors I and II and their receptors in murine mammary adenocarcinomas of different metastasizing abilityInt J Cancer1996658128208631597
- PapaVGliozzoBClarkGMInsulin-like growth factor-I receptors are overexpressed and predict a low risk in human breast cancerCancer Res199353373637408339284
- JammesHPeyratJPBanEInsulin-like growth factor 1 receptors in human breast tumour: localisation and quantification by histo-autoradiographic analysisBr J Cancer1992662482531323990
- ValentinisBBasergaRIGF-I receptor signalling in transformation and differentiationMol Pathol20015413313711376123
- PeruzziFPriscoMDewsMMultiple signaling pathways of the insulin-like growth factor 1 receptor in protection from apoptosisMol Cell Biol1999197203721510490655
- KimHJLitzenburgerBCCuiXConstitutively active type I insulin-like growth factor receptor causes transformation and xenograft growth of immortalized mammary epithelial cells and is accompanied by an epithelial-to-mesenchymal transition mediated by NF-kappaB and snailMol Cell Biol2007273165317517296734
- XieYSkyttingBNilssonGBrodinBLarssonOExpression of insulin-like growth factor-1 receptor in synovial sarcoma: association with an aggressive phenotypeCancer Res1999593588359110446966
- LongLRubinRBrodtPEnhanced invasion and liver colonization by lung carcinoma cells overexpressing the type 1 insulin-like growth factor receptorExp Cell Res19982381161219457063
- SaldanaSMLeeHHLoweryFJInhibition of type I insulin-like growth factor receptor signaling attenuates the development of breast cancer brain metastasisPLoS One20138e7340624039934
- ChangLGrahamPHHaoJAcquisition of epithelial-mesenchymal transition and cancer stem cell phenotypes is associated with activation of the PI3K/Akt/mTOR pathway in prostate cancer radioresistanceCell Death Dis20134e87524157869
- IrieHYPearlineRVGruenebergDDistinct roles of Akt1 and Akt2 in regulating cell migration and epithelial-mesenchymal transitionJ Cell Biol20051711023103416365168
- GuvakovaMASurmaczEThe activated insulin-like growth factor I receptor induces depolarization in breast epithelial cells characterized by actin filament disassembly and tyrosine dephosphorylation of FAK, Cas, and paxillinExp Cell Res199925124425510438590
- SorokinAVChenJMEMO1, a new IRS1-interacting protein, induces epithelial-mesenchymal transition in mammary epithelial cellsOncogene2013323130313822824790
- LorenzattiGHuangWPalACabanillasAMKleerCGCCN6 (WISP3) decreases ZEB1-mediated EMT and invasion by attenuation of IGF-1 receptor signaling in breast cancerJ Cell Sci20111241752175821525039
- GrahamTRZhauHEOdero-MarahVAInsulin-like growth factor-I-dependent up-regulation of ZEB1 drives epithelial-to-mesenchymal transition in human prostate cancer cellsCancer Res2008682479248818381457
- PizonMZimonDCarlSPachmannUPachmannKCamaraOHeterogeneity of circulating epithelial tumour cells from individual patients with respect to expression profiles and clonal growth (sphere formation) in breast cancerEcancermedicalscience2013734323983815
- DugganCWangCYNeuhouserMLAssociations of insulin-like growth factor and insulin-like growth factor binding protein-3 with mortality in women with breast cancerInt J Cancer20131321191120022847383
- MorgilloFDe VitaFAntoniolGSerum insulin-like growth factor 1 correlates with the risk of nodal metastasis in endocrine-positive breast cancerCurr Oncol201320e283e28823904766
- DupontJKhanJQuBHMetzlerPHelmanLLeRoithDInsulin and IGF-1 induce different patterns of gene expression in mouse fibroblast NIH-3T3 cells: identification by cDNA microarray analysisEndocrinology20011424969497511606465
- DupontJFernandezAMGlackinCAHelmanLLeRoithDInsulin-like growth factor 1 (IGF-1)-induced twist expression is involved in the anti-apoptotic effects of the IGF-1 receptorJ Biol Chem2001276266992670711323435
- SciaccaLCostantinoAPandiniGInsulin receptor activation by IGF-II in breast cancers: evidence for a new autocrine/paracrine mechanismOncogene1999182471247910229198
- PandiniGVigneriRCostantinoAInsulin and insulin-like growth factor-I (IGF-I) receptor overexpression in breast cancers leads to insulin/IGF-I hybrid receptor overexpression: evidence for a second mechanism of IGF-I signalingClin Cancer Res199951935194410430101
- HarringtonSCWerohaSJReynoldsCSumanVJLingleWLHaluskaPQuantifying insulin receptor isoform expression in FFPE breast tumorsGrowth Horm IGF Res20122210811522551578
- OatesAJSchumakerLMJenkinsSBThe mannose 6-phosphate/insulin-like growth factor 2 receptor (M6P/IGF2R), a putative breast tumor suppressor geneBreast Cancer Res Treat1998472692819516081
- De SouzaATHankinsGRWashingtonMKFineRLOrtonTCJirtleRLFrequent loss of heterozygosity on 6q at the mannose 6-phosphate/insulin-like growth factor II receptor locus in human hepatocellular tumorsOncogene199510172517297753549
- ByrdJCDeviGRde SouzaATJirtleRLMacDonaldRGDisruption of ligand binding to the insulin-like growth factor II/mannose 6-phosphate receptor by cancer-associated missense mutationsJ Biol Chem1999274244082441610446221
- O’GormanDBCostelloMWeissJFirthSMScottCDDecreased insulin-like growth factor-II/mannose 6-phosphate receptor expression enhances tumorigenicity in JEG-3 cellsCancer Res1999595692569410582686
- SouzaRFWangSThakarMExpression of the wild-type insulin-like growth factor II receptor gene suppresses growth and causes death in colorectal carcinoma cellsOncogene1999184063406810435587
- OhYMullerHLNgLRosenfeldRGTransforming growth factor-beta-induced cell growth inhibition in human breast cancer cells is mediated through insulin-like growth factor-binding protein-3 actionJ Biol Chem199527013589135927539790
- ShemerJYaronAWernerHRegulation of insulin-like growth factor (IGF) binding protein-5 in the T47D human breast carcinoma cell line by IGF-I and retinoic acidJ Clin Endocrinol Metab199377124612507521344
- ColstonKWPerksCMXieSPHollyJMGrowth inhibition of both MCF-7 and Hs578T human breast cancer cell lines by vitamin D analogues is associated with increased expression of insulin-like growth factor binding protein-3J Mol Endocrinol1998201571629513092
- KarasMKleinmanDDanilenkoMComponents of the IGF system mediate the opposing effects of tamoxifen on endometrial and breast cancer cell growthProg Growth Factor Res199565135208817696
- NishimuraAFujimotoMOguchiSFusunyanRDMacDermottRPSandersonIRShort-chain fatty acids regulate IGF-binding protein secretion by intestinal epithelial cellsAm J Physiol1998275E55E639688874
- ClemmonsDRCamacho-HubnerCCoronadoEOsborneCKInsulin-like growth factor binding protein secretion by breast carcinoma cell lines: correlation with estrogen receptor statusEndocrinology1990127267926861701124
- YuHLevesqueMAKhosraviMJPapanastasiou-DiamandiAClarkGMDiamandisEPInsulin-like growth factor-binding protein-3 and breast cancer survivalInt J Cancer1998796246289842972
- RochaRLHilsenbeckSGJacksonJGInsulin-like growth factor binding protein-3 and insulin receptor substrate-1 in breast cancer: correlation with clinical parameters and disease-free survivalClin Cancer Res199731031099815544
- VesteySPerksCSenCCalderCHollyJWintersZImmunohistochemical expression of insulin-like growth factor binding protein-3 in invasive breast cancers and ductal carcinoma in situ: implications for clinicopathology and patient outcomeBreast Cancer Res20057R119R12915642160
- HollowoodADLaiTPerksCMNewcombPVAldersonDHollyJMIGFBP-3 prolongs the p53 response and enhances apoptosis following UV irradiationInt J Cancer20008833634111054660
- GillZPPerksCMNewcombPVHollyJMInsulin-like growth factor-binding protein (IGFBP-3) predisposes breast cancer cells to programmed cell death in a non-IGF-dependent mannerJ Biol Chem199727225602256079325280
- WilliamsACCollardTJPerksCMIncreased p53-dependent apoptosis by the insulin-like growth factor binding protein IGFBP-3 in human colonic adenoma-derived cellsCancer Res200060222710646845
- FowlerCAPerksCMNewcombPVSavagePBFarndonJRHollyJMInsulin-like growth factor binding protein-3 (IGFBP-3) potentiates paclitaxel-induced apoptosis in human breast cancer cellsInt J Cancer20008844845311054675
- McCaigCFowlerCALaurenceNJDifferential interactions between IGFBP-3 and transforming growth factor-beta (TGF-beta) in normal vs cancerous breast epithelial cellsBr J Cancer2002861963196912085194
- YuQBanerjeeKPatersonJAlamiNShiryLLeyland-JonesBIGFBP-3: Single Agent and Synergistic Effects with Chemotherapeutic Drugs on Solid Tumor Models, AACRProceedings of the AACR, 172: Abstract 755Washington DC2003
- McIntoshJDennisonGHollyJMIGFBP-3 can either inhibit or enhance EGF-mediated growth of breast epithelial cells dependent upon the presence of fibronectinJ Biol Chem2010285387883880020851879
- BurrowsCHollyJMLaurenceNJInsulin-like growth factor binding protein 3 has opposing actions on malignant and nonmalignant breast epithelial cells that are each reversible and dependent upon cholesterol-stabilized integrin receptor complexesEndocrinology20061473484350016614079
- IoachimECharchantiABriasoulisEImmunohistochemical expression of extracellular matrix components tenascin, fibronectin, collagen type IV and laminin in breast cancer: their prognostic value and role in tumour invasion and progressionEur J Cancer2002382362237012460779
- BaxterRCIGF binding proteins in cancer: mechanistic and clinical insightsNat Rev Cancer20141432934124722429
- JohnsonMAFirthSMIGFBP-3: a cell fate pivot in cancer and diseaseGrowth Horm IGF Res201424516417324953254
- WalshLADamjanovskiSIGF-1 increases invasive potential of MCF 7 breast cancer cells and induces activation of latent TGF-β1 resulting in epithelial to mesenchymal transitionCell Commun Signal201191021535875
- ReissMBarcellos-HoffMHTransforming growth factor-beta in breast cancer: a working hypothesisBreast Cancer Res Treat19974581959285120
- DieciMVOrvietoEDominiciMContePGuarneriVRare breast cancer subtypes: histological, molecular, and clinical peculiaritiesOncologist20141980581324969162
- KittanehMMonteroAJGluckSMolecular profiling for breast cancer: a comprehensive reviewBiomarkers Cancer201356170
- BooneDNLeeAVTargeting the insulin-like growth factor receptor: developing biomarkers from gene expression profilingCrit Rev Oncog20121716117322471706
- LeeAVYeeDTargeting IGF-1R: at a crossroadOncology (Williston Park)201125535536 discussion 55121717908
- ChenHXSharonEIGF-1R as an anti-cancer target – trials and tribulationsChin J Cancer20133224225223601239
- AlamiNPageVYuQRecombinant human insulin-like growth factor-binding protein 3 inhibits tumor growth and targets the Akt pathway in lung and colon cancer modelsGrowth Horm IGF Res20081848749618502161