Abstract
Historically, the name of natural killer (NK) cells came from their natural ability to kill tumor cells in vitro. From the 1970s to date, accumulating data highlighted the importance of NK cells in host immune response against cancer and in therapy-induced antitumor response. The recognition and the lysis of tumor cells by NK cells are regulated by a complex balance of inhibitory and activating signals. This review summarizes NK cell mechanisms to kill cancer cells, their role in host immune responses against tumor growth or metastasis, and their implications in antitumor immunotherapies via cytokines, antibodies, or in combination with other therapies. The regulatory role of NK cells in autoimmunity is also discussed.
Introduction to natural killer cells in antitumor immune response
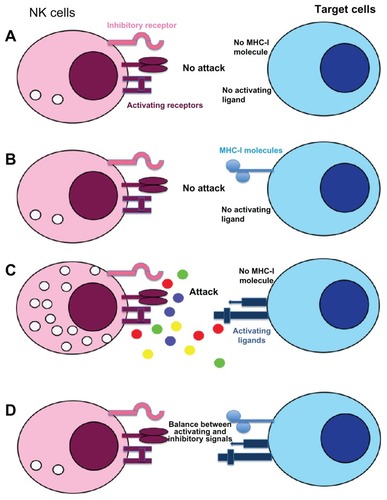
Natural killer (NK) cells were discovered in humans and mice in 1975 due to specific functional criteria corresponding to their ability to lyse certain tumor cells in the absence of prior stimulation.Citation1 They possess a morphology of large granular lymphocytes () and their receptor genes involved in the recognition of pathogens remain in germinal configuration unlike T and B cells.Citation2 Thus, they differ from the T and B lymphocytes by the permanent presence of a significant fraction of educated and primed cells.Citation3 The formulation of the hypothesis of “missing self ” by Klas Kärre, based on the fact that NK cells are able to detect and lyse cells with a deficient expression of major histocompatibility complex class I (MHC-I) molecules, allowed a better understanding of the function and the role of NK cells in the immune response.Citation4 In the 1990s, several studies highlighted the presence of inhibiting and activating receptors expressed by NK cellsCitation5 that led to the identification of a new recognition model called “induced-self.”Citation6 This new model complements the hypothesis of “missing-self ” by explaining why NK cells kill tumor cells expressing MHC-I molecules or save autologous cells with absent MHC-I expression (). Indeed, NK cell triggering is the result of a complex balance between inhibitory and activating signals and require not only a deficient MHC-I expression on target cells but also the expression of inducible ligands of activating NK cell receptors.Citation7 Consequently, these cells have the ability to recognize and destroy a wide range of abnormal cells (including tumor cells, virus-infected cells, cells bound by an antibody, allogeneic cells), as well as stressed cells, without damaging the healthy and normal “self ” cells.Citation8 Therefore, NK cells have several important effector functions such as the initiation and amplification of the inflammatory response, the production of chemokines and cytokines, and the lysis of sensitive target cells.Citation9,Citation10
Figure 1 Electron micrographics of natural killer (A) and NK-92 (B) cells showing large lymphocyte-containing granules (arrows).
Abbreviation: N, nucleus.
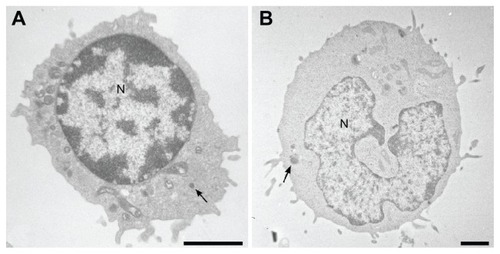
Figure 2 Recognition mechanisms of target cells by NK cells: “missing and induced self” theory. NK cell response is not initiated if neither ligands for NK-activating receptors nor MHC-I are expressed on target cells (A). If inhibitory receptors interact with MHC-I molecules without ligands for activating receptors no cytotoxicity is observed (B), whereas engagement of these receptors in absence of MHC-I molecule induced a strong NK cell response (C). In most cases, NK cell response depends on a balance between inhibitory and activating receptor signaling (D). Normal cells are protected against NK cell cytotoxicity because they usually express MHC-I molecules and no or low level of activating receptor ligands.
Abbreviations: MHC-I: major histocompatibility complex class I; NK, natural killer.
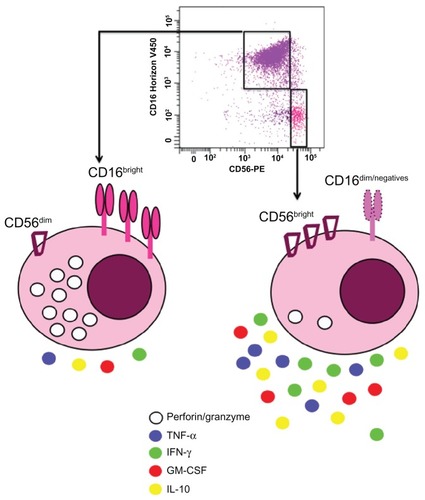
NK cells represent 5% to 20% of peripheral blood mononuclear cells, usually defined as CD16+ CD56+ CD3− cells and are also found in many tissues such as liver, peritoneal cavity, placenta, or the uterine mucosa.Citation11–Citation14 Human NK cells can be divided into two subpopulations according to the density of CD16 and CD56 expression on their surface (). The majority of NK cells in blood (90%–95%) or at inflammation sites have a moderate expression of CD56 (CD56dim) and a strong expression of CD16 (). These cells possess a high cytotoxic potential.Citation15 The CD56bright subpopulation predominates in lymph nodes, expresses no or low levels of CD16, displays little cytotoxicity, and mainly produces cytokines upon activation ().Citation16 CD56 is not expressed on mouse NK cells, but recently Hayakawa and SmythCitation17 categorized these cells depending on their CD27 expression. Mouse CD27bright NK cells share several characteristics with human CD56bright NK cells. Accordingly, they predominate in lymph nodes and produce large amounts of cytokines. Recently, the discovery of a new NK marker, NKp46, allowed to define human and mice NK cells because, in contrast to others markers (CD16 and CD56 for humans or DX5 or NK1.1 for mice), NKp46 is exclusively expressed by all NK cells in both species.Citation18 Several NK cell lines have been established from lymphoma or peripheral blood mononuclear cells such as NK-92Citation19 () or NKG,Citation20 respectively.
Figure 3 Human natural killer cell subsets based on CD56 and CD16 expressions: Around 90% of natural killer cells isolated from the blood display dim level of CD56 and high density of CD16 (CD56dimCD16bright).
Abbreviations: GM-CSF, granulocyte–macrophage colony-stimulating factor; IFN-γ, interferon γ; IL-10, interleukin 10; TNF-α, tumor necrosis factor α.
From their discovery, NK cells were described as white blood cells able to lyse tumor cells such as K562, a tumor cell line generated from a patient with chronic leukemia.Citation21 Shortly thereafter, NK cells were shown to eliminate circulating tumor cells in mice,Citation22 but also to kill spontaneously cells deficient for MHC-I.Citation4 The generation of mice genetically deficient for NK cells or depleted of these cells by antibodies, highlighted that NK cells have a role in immunosurveillance of cancer and the ability to prevent the tumor growth.Citation23–Citation25 Indeed, the transplantation of lymphoma cells in mice conduct to a more aggressive tumor growth in NK cell deficient mice compared to wild type mice.Citation26
Review of the activation and mechanism of action of NK cells
Unlike T and B lymphocytes, NK cells will not rearrange their genes encoding for receptor antigen recognition, but they have the ability to recognize target cells directly through inhibitory or activating receptors expressed on the cell surface. The balance between stimulatory and inhibitory signals will determine the activation status of NK cells since lysis of the target cell will only happen when the activating signals outweigh the inhibitory signals. The first checkpoint is the expression of MHC-I molecules.Citation4,Citation27 In fact, downregulation of MHC-I is observed during tumor transformationCitation28 or viral infectionCitation29 and prevents the binding of inhibitory receptors of NK cells to the target cell. Simultaneously, ligands for activating receptor of NK cells must be expressed on the target cell to trigger NK cell cytotoxicity. These ligands are absent or expressed in low amounts on normal cells, although they are highly expressed on potentially harmful cells as a consequence of cellular stress, viral infection, or tumor transformation.
Different mechanisms are known to be involved in the destruction of tumor cells by NK cells:
Perforin/granzyme-mediated cytotoxicity: The release of cytotoxic granules composed of perforin and granzymesCitation30 is the fastest and also the most powerful way to lyse tumor cells. By creating a synapse with the target cell, NK cells will drop, at this junction, perforin and granzyme molecules inducing the lysis of the target cell.Citation31 Mice deficient for perforin are less efficient (about 10 to 100 times) to suppress syngeneic MHC-I deficient tumor cells, suggesting that this molecule is required for NK cell cytotoxicity.Citation32 Moreover, many studies have shown that perforin is important for the immunosurveillance of several spontaneous malignanciesCitation33 whereas the role of granzymes is less characterized.Citation34
Death receptor mediated apoptosis: The death of the target cell, induced by apoptosis via tumor necrosis factor (TNF) family ligands, Fas ligand (CD178), TNF, and TRAIL (tumor-necrosis factor-related apoptosis-inducing ligand),Citation35 is an alternative to the release of granules. This second mechanism, which is slower (several hours) and often less efficient than the previous one, requires the presence of the TNF family ligand expression on the surface of NK cells. These ligands will bind to a receptor Fas on the surface of the target cell. The effectiveness of this pathway is controlled by various factors such as expression of the receptor for FasL or TRAIL by the cancer cells or intracellular mechanisms protecting against apoptosis. For example, a murine subset of NK cells in liver expressing TRAIL has been shown to kill cancer cells and clear tumors from the liver.Citation36
Interferon-γ effector functions: After activation, NK cells secrete various cytokines such as interferon γ (IFN-γ), TNF-α, granulocyte–macrophage colony-stimulating factor (GM-CSF), interleukin (IL)-10, or IL-13 and their antitumor activities can be mediated by IFN-γ.Citation37 Indeed, IFN-γ produced by NK cells contributes to eliminate tumor metastases and sarcoma induced by methylcholanthrene in a murine model.Citation38 This cytokine inhibits proliferation of tumor cells in vitro and indirectly the tumor growth in vivo by inducing the antiangiogenic factors, IP-10.Citation39 IFN-γ has been also described to enhance NK cell cytotoxicity by overexpressing adhesion molecules or by increasing the sensitivity of tumor cells to cytotoxicity mediated by granule release or death receptor engagement.Citation40 TRAIL expression on murine liver NK cells is dependent on IFN-γ expression and contributes to the natural antimetastatic role of these NK cells.Citation36 IFN-γ induced TRAIL expression is also implicated in the IL-12 mediated anti-metastatic effect.Citation24
Additionally, IFN-γ plays a role in the stimulation of dendritic cells (DC). In combination with CD40 engagement, IFN-γ induces IL-12 production by DCs.Citation41 In this way, NK cells contribute indirectly to tumor control by helping the initiation and maintenance of an efficient T cell-mediated antitumor response via a crosstalk with DC.Citation42,Citation43
Overview on the NK cells receptor types, history, and discovery
NK cell inhibitory receptors
As previously mentioned, it was originally proposed by Kärre et alCitation4 that NK cells discriminate target cells from normal cells by the level of MHC-I expression on the cell surface. NK cells preferentially lyse cells expressing few or no MHC-I.Citation44 The sensitivity of cells with low MHC-I expression to NK cell lysis may be prevented by the re-expression of these molecules.Citation45,Citation46 Three families of inhibitory receptors recognizing MHC-I molecules were described (): KIR (killer cell immunoglobulin-like receptor, humans),Citation47,Citation48 Ly49 (mice),Citation49,Citation50 and CD94/NKG2A (human and mice)Citation51–Citation53 receptor family.
Table 1 Main receptors on NK cells involved in anti-tumor immune response
Immunoreceptor tyrosine-based inhibitory motif (ITIM), present in all cytoplasmic domains of NK cell inhibitory receptors, is able to recruit intracellular tyrosine phosphatases SHP-1 and SHP-2, which inhibit cytotoxicity and cytokine production.Citation54
KIR receptors are encoded by about twelve polymorphic genes and are expressed on NK cells and memory T lymphocytes. The binding to one type of KIR receptor is sufficient to prevent activation of NK cells, whereas it usually takes several different activating signals to induce degranulation and death of the target cell. The CD94/NKG2A receptor, conserved in rodents and primates, is expressed on most NK cells and a subset of CD8+ memory T cells.Citation55 CD94 has no cytoplasmic signalization domain while the receptor NKG2A contains two ITIM.Citation56 KIR, but also murine Ly49 receptors, bind directly to MHC-I molecules, whereas the CD94/NKG2A receptor binds to a peptide derived from the signal sequence of MHC-I.Citation50,Citation54 All inhibitory receptors are found by overlapping NK cell subpopulations since each cell expresses only a few types of inhibitory receptors. Consequently, NK cells have many complex combinations of MHC-I repertoire randomly distributed on the cells. The only rule that seems to be established is that all NK cells have at least one inhibitory receptor specific for MHC-I molecule to avoid autoreactivity.
NK cell-activating receptors
In addition to inhibitory receptors, NK cells express a wide range of activating receptors. Their biological role is not yet fully known because all their ligands have not been identified. The main activating receptors involved in tumor lysis are CD16, NKG2D receptor, and the natural cytotoxicity receptors (NCR)Citation57–Citation60 ().
The activating receptors have no ITIM in their cytoplasmic domains. Instead, they have charged residues in their transmembrane domains, which are necessary for the association with adapter proteins. These proteins have short extracellular domains and, therefore, do not participate in ligand binding. The intracellular domains of these adapter proteins have docking sites for signaling molecules that play a role downstream of the stimulation. Most of the adapter proteins (FcɛRγI, CD3ζ, DAP12, and DAP10) contain immunoreceptor tyrosine-based activation motif in their cytoplasmic domains, which enable them to associate with ZAP70 and/or proteins of syk kinases family.Citation61,Citation62
CD16 is a low-affinity receptor for the Fc portion of immunoglobulin (FcγRIII). This receptor with a transmembrane domain (FcγRIIIa) is found on the surface of NK cells but also on some DC, T lymphocytes, monocytes, and macrophages, and a glycosylphosphatidylinositol-linked receptor (FcγRIIIb) is expressed on neutrophils.Citation63 When IgG molecules recognize specific antigens on a tumor cell, NK cells, via CD16, are able to bind tumor cell coated with antibodies and induce tumor cell death. This reaction called antibody-dependent cell-mediated cytotoxicity is a dominant component of antibody based immunotherapy against tumors.Citation64 Recently, CD16 has been implicated in the recognition of human papillomavirus by NK cells in uterine preneoplastic lesions.Citation65
NKG2D, unlike the other NKG2 receptors, does not have an ITIM sequence and is not associated with CD94.Citation66 It is expressed on most NK cells but also on γδ and CD8+ T cells. It is associated with the adapter molecule DAP10 in humans, and DAP10 and DAP12 in mice.Citation67,Citation68 Binding of NKG2D ligands, such as MICA or MICB, leads to an increase in proliferation, cytotoxicity, and production of cytokines and chemokines (IFN-γ, GM-CSF, TNF-α).Citation69 Expression of NKG2D ligands is observed on many tumor cell lines and tumor tissues.Citation70,Citation71
More specifically expressed on NK cells, the principal NCR are NKp46, NKp44, and NKp30.Citation59,Citation72,Citation73 Molecular cloning of NCR confirmed that they were structurally distinct although they belong to the same immunoglobulin superfamily.Citation74 NKp46 (human and mice) and NKp30 (human) are expressed on both resting and activated NK cells, while the NKp44 receptor is only present on IL-2-activated human NK cells and a minor subset of γδ T cells.Citation72 The density of NCR on the cell surface varies with individuals and there is a direct correlation between NCR expression on human NK cells and their capacity to kill tumor cells.Citation75 NKp46, the first identified NCR, is a 46 kDa glycoprotein with a transmembrane domain that interacts with the adapter molecule CD3ζ.Citation59,Citation76 The activation of NKp46 leads to mobilization of calcium in the development of cytolytic activity and cytokine production.Citation59 Monoclonal antibodies against NKp46 block the lysis of a wide range of tumor cells showing that this receptor plays a major role in NK cell cytotoxicity.Citation77 This function of NKp46 is negatively regulated by the interaction between inhibitory receptors and MHC-I. NKp44 is a 44 kDa glycoprotein whose expression is inducible by IL-2, suggesting that it could contribute to the increased efficiency of activated NK cells to lyse tumor cells.Citation72 NKp44 acts in association with the immunoreceptor tyrosine-based activation motif on the adapter molecule DAP12. The blocking of NKp44 by monoclonal antibodies induces a partial inhibition of the cytolytic activity against some tumor cells and this inhibition is strongly enhanced by the addition of antibodies blocking NKp46.Citation78 NKp30 is a glycoprotein of 30 kDa that associates with the adapter molecules CD3ζ and FcɛRγI.Citation73 Its expression at the cell surface is correlated with NKp46. In addition, NKp30 cooperates with NKp46 and NKp44 to induce NK cell cytotoxicity against a variety of target cells.Citation73 NKp46, NKp44, and NKp30 bind to heparan sulfates on the surface of tumor cells.Citation79,Citation80 In addition, NKp30 is involved in the lysis of tumor cells by binding to factor BAT3 (HLA-B associated transcript (3) or to B7-H6 present on their surface.Citation81,Citation82
Role of NK cells in immune response against tumor growth and metastasis
In vivo and in vitro studies have shown that NK cells can eliminate tumor cells.Citation83 In mice, tumor rejection is dependent upon the presence or absence of NK cell receptor ligands on the tumor. Especially the lack of MHC-I expression, overexpression of NKG2D ligands (H60, Rae1β, Rae1δ, Rae1γ, Mult-1), or costimulatory signals makes the tumor more susceptible to lysis by NK cells.Citation84 Moreover, engagement of NCR complements NKG2D pathway in the killing of tumor cells by NK cells.Citation85 Little is known about the mechanism of NK cell migration in tumor, but selectins seem to play a role in this recruitment.Citation86 However, Smyth et alCitation25,Citation87 showed that mice deficient in NK cells are more susceptible to methylcholanthrene-induced sarcomas, demonstrating that NK cells play a role in tumor immunosurveillance. NK cells also protect against the growth of B cell lymphomas in mice lacking perforin and β2 microglobulin.Citation88 Moreover, NK cells participate to immune response against metastasis; in an immunotherapy protocol using synthetic oligodeoxynucleotides containing CpG motifs, NK cells prevent pulmonary metastasis and peritoneal dissemination. Citation89 In a mouse model, NK cells inhibit pulmonary metastasis formation after IFN-γ treatment.Citation90
Also, in humans, there is evidence that NK cells play a role in the tumor immunosurveillance. An 11-year follow-up survey has shown that a low NK cell activity is associated with an increased cancer risk.Citation91 This was confirmed in several human malignancies. For example, a decrease of NK cell activity is observed in patients with hereditary colorectal adenocarcinomaCitation92,Citation93 and melanoma patients with metastatic disease have an impaired perforin-dependent NK cell cytotoxic mechanism.Citation94 Tumor growth can disturb the functional maturation of NK cells by interrupting the IL-15 signaling pathway,Citation95 but further studies are necessary to better understand the immunologic basis of NK cell defects in tumor.
As already mentioned, NK cells collaborate with antigen-presenting cells to amplify the immune response.Citation96 This collaboration can help to induce a T cell-mediated antitumor immunity.Citation97 After the implantation of MHC-I low tumor cells in mice, the release of IFN-γ by NK cells stimulate the maturation of DC to a IL-12-producing DC1 phenotype that promote a strong and protective antitumor CD8+ T cell response.Citation42,Citation98
Implications for tumor management
The observation that IL-2 increased in vitro the cytotoxic activity of NK cells against tumor cellsCitation99 has been conducted to perform clinical trials with adoptive transfer of high doses of this cytokine in patients with metastatic melanoma or renal cell carcinoma.Citation100,Citation101 Due to systemic toxicity of IL-2 and to the fact that IL-2 preferentially drives expansion of regulatory T cells, which can inhibit antitumor immunity, other cytokines, such as IL-15, were tested in nonhuman primates. IL-15 shares similar properties with IL-2 and Berger et alCitation102 showed that intermittent administration of IL-15 should be considered in clinical studies. Also other cytokines, such as IL-12 and IL-18, synergistically enhance NK cytotoxicity against tumor targets and IFN-γ production by NK cells.Citation103,Citation104
Tumor-targeted monoclonal antibodies can induce NK cell antibody-dependent cell-mediated cytotoxicity and the rapid degranulation of NK cells results in tumor cell destruction.Citation105 The clinical efficacy of monoclonal antibodies directed against CD20 (rituximab), Her2/neu (trastuzumab), epidermal growth factor receptor (cetuximab), or disialoganglioside (GD2) is, at least partially, due to NK antibody-dependent cell-mediated cytotoxicity.Citation106–Citation109 Moreover, in a Phase I trial, coadministration of IL-12 with trastuzumab enhances the antitumor response induced by the antibody.Citation110
Besides the development of protocols to stimulate NK cell activity, inhibitory receptors on NK cells could be the target for antitumor therapy. Blocking Ly49 inhibitory receptors enhanced antitumor activity in vitro and in vivoCitation111 and in human, antibodies blocking KIR are currently tested in a Phase II clinical trial.Citation112 NK allogeneic recognition via their KIR repertoire has a major role in reducing the risk of relapse by inducing a graft versus leukemia effect after allogeneic hematopoietic stem cell transplantation.Citation113
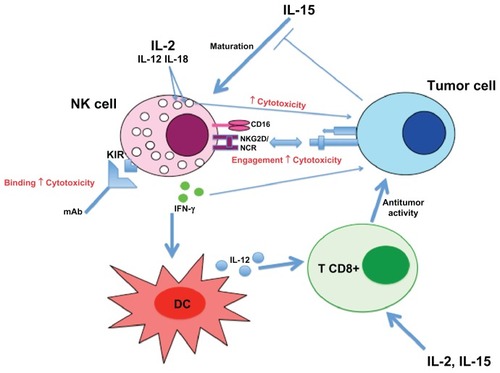
NK cells seem important in tumor vaccination with DC, since clinical responses are correlated with superior levels of activated NK cells in responders.Citation114 In a Phase I clinical trial with melanoma patients, DC derived-exosomes enhanced NKG2D-dependent function of NK cells in half of the patients.Citation115 Indirectly, immunotherapy protocols targeting an oncogenic viral proteinCitation116 or the recruitment of antigen-presenting cellsCitation117 boost NK cell response. Even some chemotherapeutic agents could inhibit secretion of inhibitory soluble NKG2D ligands.Citation118 A NK cell treatment combined with radiation therapy has been proposed, since radiation increases NK-activating ligand (eg, NKG2DL) expression via DNA damage response.Citation119 summarizes the different mechanisms implicated into NK cell response against tumor cells.
Figure 4 Overview of NK cell responses against tumor cell.
Role of NK cells in autoimmunity
NK cells are viewed as effector cells whose rapid killing of transformed or infected cells provides a first-line of defense prior to the initiation of an adaptive immune response against tumor or infection. However, studies on NK cells suggest a broad role in immunity including the potential to function as regulatory cells. While NK cells can assist in DC maturation and T cell polarization, increasing evidence indicates that NK cells can also prevent and limit adaptive autoimmune responses.Citation120 Autoimmune diseases are a multistep process caused by inappropriate activation of cells of the adaptive immune system (T and B cells) which results in cell-specific, organ-specific, or systemic tissue damage.Citation121 Autoimmunity has been linked to cancer, for example, patients with scleroderma have an increased risk to develop a tumor.Citation122 Moreover autoimmunity is associated with immunotherapy of cancer.Citation123
Several studies suggested that NK cells play a role at the different stages of the autoimmune response.Citation124 NK cells can either augment or ameliorate autoimmune diseases.Citation125,Citation126 A reduction of circulating NK cells has been shown in diverse autoimmune diseases, which contributed to an impaired NK activity.Citation127,Citation128 The modulation in the number of circulating NK cells seems to be a primary event instead of an active inflammation/drug administration consequence during autoimmune processes. However, some biotherapies are linked with changes of circulating NK cells compartment. Daclizumab (anti-IL-2Rα) treatment in multiple sclerosisCitation129 or in uveitisCitation130 pathogenesis is associated with an increase in the number of CD56bright NK cells. This augmentation of CD56bright NK cells is correlated with the suppression of the disease activity.Citation129 NK cells can also promote autoimmune diseases, ie, autoimmune diabetes is prevented in nonobese-diabetic mice by blocking the activating NK cell receptor NKG2D.Citation131 In humans, the predisposition to rheumatoid arthritis,Citation132 psoriatic arthritis,Citation133 scleroderma,Citation134 and psoriasis vulgarisCitation135 is linked to the expression of certain KIR and HLA alleles, but the precise role of NK cells in these diseases is still under investigation.
Conclusion
NK cells could be very efficient killers of tumor cells and could help to induce an optimal adaptive immune response against cancer. A better knowledge in the basic biology of NK cells is a key to develop strategies to manipulate NK cells for therapeutic purposes.
Acknowledgments
IL and VMR are supported by a Télévie grant from the Belgian National Fund for Scientific Research (FNRS) and NJ is a research associate of the FNRS.
References
- KiesslingRKleinEWigzellH“Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotypeEur J Immunol1975521121171234049
- BironCAvan den ElsenPTuttMMMedveczkyPKumarVTerhorstCMurine natural killer cells stimulated in vivo do not express the T cell receptor alpha, beta, gamma, T3 delta, or T3 epsilon genesJ Immunol19871395170417103497976
- Di SantoJPNatural killer cell developmental pathways: a question of balanceAnnu Rev Immunol20062425728616551250
- KärreKLjunggrenHGPiontekGKiesslingRSelective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategyNature198631960556756783951539
- LanierLLNatural killer cells: from no receptors to too manyImmunity1997643713789133416
- RauletDHVanceREMcMahonCWRegulation of the natural killer cell receptor repertoireAnnu Rev Immunol20011929133011244039
- LanierLLMissing self, NK cells, and the white albumJ Immunol200517411656515905491
- CaligiuriMAHuman natural killer cellsBlood2008112346146918650461
- VivierETomaselloEBaratinMWalzerTUgoliniSFunctions of natural killer cellsNat Immunol20089550351018425107
- WuJLanierLLNatural killer cells and cancerAdv Cancer Res20039012715614710949
- KingABurrowsTLokeYWHuman uterine natural killer cellsNat Immun199615141529032767
- LuoDVermijlenDVanderkerkenKInvolvement of LFA-1 in hepatic NK cell (pit cell)-mediated cytolysis and apoptosis of colon carcinoma cellsJ Hepatol199931111011610424290
- RobertsonMJRitzJBiology and clinical relevance of human natural killer cellsBlood19907612242124382265240
- TrinchieriGBiology of natural killer cellsAdv Immunol1989471873762683611
- NaglerALanierLLCwirlaSPhillipsJHComparative studies of human FcRIII-positive and negative natural killer cellsJ Immunol198914310318331912530273
- CooperMAFehnigerTATurnerSCHuman natural killer cells: a unique innate immunoregulatory role for the CD56(bright) subsetBlood200197103146315111342442
- HayakawaYSmythMJCD27 dissects mature NK cells into two subsets with distinct responsiveness and migratory capacityJ Immunol200617631517152416424180
- WalzerTBleryMChaixJIdentification, activation, and selective in vivo ablation of mouse NK cells via NKp46Proc Natl Acad Sci U S A200710493384338917360655
- GongJHMakiGKlingemannHGCharacterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cellsLeukemia1994846526588152260
- ChengMMaJChenYEstablishment, characterization and successful adaptive therapy against human tumors of NKG Cell, a new human NK cell lineCell TransplantJune 72011 [Epub ahead of print.]
- OrtaldoJROldhamRKCannonGCHerbermanRBSpecificity of natural cytotoxic reactivity of normal human lymphocytes against a myeloid leukemia cell lineJ Natl Cancer Inst1977591778269036
- RiccardiCSantoniABarlozzariTPuccettiPHerbermanRBIn vivo natural reactivity of mice against tumor cellsInt J Cancer19802544754866154658
- KimSIizukaKAguilaHLWeissmanILYokoyamaWMIn vivo natural killer cell activities revealed by natural killer cell-deficient miceProc Natl Acad Sci U S A20009762731273610694580
- SmythMJCretneyETakedaKTumor necrosis factor-related apoptosis-inducing ligand (TRAIL) contributes to interferon gamma-dependent natural killer cell protection from tumor metastasisJ Exp Med2001193666167011257133
- SmythMJCroweNYGodfreyDINK cells and NKT cells collaborate in host protection from methylcholanthrene-induced fibrosarcomaInt Immunol200113445946311282985
- HalfteckGGElboimMGurCAchdoutHGhadiallyHMandelboimOEnhanced in vivo growth of lymphoma tumors in the absence of the NK-activating receptor NKp46/NCR1J Immunol200918242221223019201876
- LodoenMBLanierLLViral modulation of NK cell immunityNat Rev Microbiol200531596915608700
- GarridoFAlgarraIMHC antigens and tumor escape from immune surveillanceAdv Cancer Res20018311715811665717
- HewittEWThe MHC class I antigen presentation pathway: strategies for viral immune evasionImmunology2003110216316914511229
- ClementMVHaddadPSoulieAInvolvement of granzyme B and perforin gene expression in the lytic potential of human natural killer cellsRes Immunol199014164774892284495
- BrycesonYTMarchMELjunggrenHGLongEOActivation, coactivation, and costimulation of resting human natural killer cellsImmunol Rev2006214739117100877
- KagiDLedermannBBurkiKCytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient miceNature1994369647531378164737
- SmythMJThiaKYStreetSEMacGregorDGodfreyDITrapaniJAPerforin-mediated cytotoxicity is critical for surveillance of spontaneous lymphomaJ Exp Med2000192575576010974040
- CaoXCaiSFFehnigerTAGranzyme B and perforin are important for regulatory T cell-mediated suppression of tumor clearanceImmunity200727463564617919943
- ZamaiLAhmadMBennettIMAzzoniLAlnemriESPerussiaBNatural killer (NK) cell-mediated cytotoxicity: differential use of TRAIL and Fas ligand by immature and mature primary human NK cellsJ Exp Med199818812237523809858524
- TakedaKHayakawaYSmythMJInvolvement of tumor necrosis factor-related apoptosis-inducing ligand in surveillance of tumor metastasis by liver natural killer cellsNat Med2001719410011135622
- TrinchieriGNatural killer cells wear different hats: effector cells of innate resistance and regulatory cells of adaptive immunity and of hematopoiesisSemin Immunol19957283887579198
- StreetSECretneyESmythMJPerforin and interferon-gamma activities independently control tumor initiation, growth, and metastasisBlood200197119219711133760
- AngiolilloALSgadariCTaubDDHuman interferon-inducible protein 10 is a potent inhibitor of angiogenesis in vivoJ Exp Med199518211551627540647
- BironCANguyenKBPienGCCousensLPSalazar-MatherTPNatural killer cells in antiviral defense: function and regulation by innate cytokinesAnnu Rev Immunol19991718922010358757
- SnijdersAKalinskiPHilkensCMKapsenbergMLHigh-level IL-12 production by human dendritic cells requires two signalsInt Immunol19981011159315989846688
- AdamCKingSAllgeierTDC-NK cell cross talk as a novel CD4+ T-cell-independent pathway for antitumor CTL inductionBlood2005106133834415769894
- JacobsNBoniverJHubertPDelvennePDendritic cells: more than just adaptive immunity inducers?Curr Immunol Rev200731722
- LjunggrenHGKärreKHost resistance directed selectively against H-2-deficient lymphoma variants. Analysis of the mechanismJ Exp Med19851626174517593877776
- FrankssonLGeorgeEPowisSButcherGHowardJKärreKTumorigenicity conferred to lymphoma mutant by major histocompatibility complex-encoded transporter geneJ Exp Med199317712012058418201
- LjunggrenHGSturmhofelKWolpertEHammerlingGJKärreKTransfection of beta 2-microglobulin restores IFN-mediated protection from natural killer cell lysis in YAC-1 lymphoma variantsJ Immunol199014513803862113557
- WagtmannNRajagopalanSWinterCCPeruzziMLongEOKiller cell inhibitory receptors specific for HLA-C and HLA-B identified by direct binding and by functional transferImmunity1995368018098777725
- JamilKMKhakooSIKIR/HLA interactions and pathogen immunityJ Biomed Biotechnol2011201129834821629750
- YokoyamaWMJacobsLBKanagawaOShevachEMCohenDIA murine T lymphocyte antigen belongs to a supergene family of type II integral membrane proteinsJ Immunol19891434137913862787364
- OrrMTLanierLLInhibitory Ly49 receptors on mouse natural killer cellsCurr Top Microbiol Immunol2011350678720680808
- CarreteroMCantoniCBellonTThe CD94 and NKG2-A C-type lectins covalently assemble to form a natural killer cell inhibitory receptor for HLA class I moleculesEur J Immunol19972725635679045931
- HouchinsJPYabeTMcSherryCBachFHDNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on human natural killer cellsJ Exp Med19911734101710202007850
- PetrieEJClementsCSLinJCD94-NKG2A recognition of human leukocyte antigen (HLA)-E bound to an HLA class I leader sequenceJ Exp Med2008205372573518332182
- Le DreanEVelyFOlceseLInhibition of antigen-induced T cell response and antibody-induced NK cell cytotoxicity by NKG2 A: association of NKG2 A with SHP-1 and SHP-2 protein-tyrosine phosphatasesEur J Immunol19982812642769485206
- MasilamaniMNguyenCKabatJBorregoFColiganJCD94/NKG2A inhibits NK cell activation by disrupting the actin network at the immunological synapseJ Immunol200617763590359616951318
- CarreteroMPalmieriGLlanoMSpecific engagement of the CD94/NKG2-A killer inhibitory receptor by the HLA-E class Ib molecule induces SHP-1 phosphatase recruitment to tyrosine-phosphorylated NKG2-A: evidence for receptor function in heterologous transfectantsEur J Immunol1998284128012919565368
- BauerSGrohVWuJActivation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICAScience1999285542872772910426993
- BiassoniRNatural killer cell receptorsAdv Exp Med Biol2008640355219065782
- SivoriSVitaleMMorelliLp46, a novel natural killer cell-specific surface molecule that mediates cell activationJ Exp Med19971867112911369314561
- BrycesonYTMarchMEBarberDFLjunggrenHGLongEOCytolytic granule polarization and degranulation controlled by different receptors in resting NK cellsJ Exp Med200520271001101216203869
- LanierLLCorlissBCWuJLeongCPhillipsJHImmunoreceptor DAP12 bearing a tyrosine-based activation motif is involved in activating NK cellsNature199839166687037079490415
- OlceseLCambiaggiASemenzatoGBottinoCMorettaAVivierEHuman killer cell activatory receptors for MHC class I molecules are included in a multimeric complex expressed by natural killer cellsJ Immunol199715811508350869164921
- MoldovanIGalonJMaridonneau-PariniIRegulation of production of soluble Fc gamma receptors type III in normal and pathological conditionsImmunol Lett199968112513410397167
- NimmerjahnFRavetchJVAntibodies, Fc receptors and cancerCurr Opin Immunol200719223924517291742
- RenouxVBisigBLangersIHuman papillomavirus entry into NK cells requires CD16 expression and triggers cytotoxic activity and cytokine secretionEur J Immunol201144113240325221830210
- HoELHeuselJWBrownMGMatsumotoKScalzoAAYokoyamaWMMurine Nkg2d and Cd94 are clustered within the natural killer complex and are expressed independently in natural killer cellsProc Natl Acad Sci U S A19989511632063259600963
- ChampsaurMLanierLLEffect of NKG2D ligand expression on host immune responsesImmunol Rev2010235126728520536569
- RosenDBArakiMHamermanJAChenTYamamuraTLanierLLA Structural basis for the association of DAP12 with mouse, but not human, NKG2DJ Immunol200417342470247815294961
- SutherlandCLChalupnyNJSchooleyKVandenBosTKubinMCosmanDUL16-binding proteins, novel MHC class I-related proteins, bind to NKG2D and activate multiple signaling pathways in primary NK cellsJ Immunol2002168267167911777960
- GrohVBahramSBauerSHermanABeauchampMSpiesTCell stress-regulated human major histocompatibility complex class I gene expressed in gastrointestinal epitheliumProc Natl Acad Sci U S A1996932212445124508901601
- CerwenkaANew twist on the regulation of NKG2D ligand expressionJ Exp Med2009206226526819204110
- CantoniCBottinoCVitaleMNKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamilyJ Exp Med1999189578779610049942
- PendeDParoliniSPessinoAIdentification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cellsJ Exp Med1999190101505151610562324
- BottinoCBiassoniRMilloRMorettaLMorettaAThe human natural cytotoxicity receptors (NCR) that induce HLA class I-independent NK cell triggeringHum Immunol20006111610658972
- CostelloRTSivoriSMarcenaroEDefective expression and function of natural killer cell-triggering receptors in patients with acute myeloid leukemiaBlood200299103661366711986221
- PessinoASivoriSBottinoCMolecular cloning of NKp46: a novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicityJ Exp Med199818859539609730896
- SivoriSPendeDBottinoCNKp46 is the major triggering receptor involved in the natural cytotoxicity of fresh or cultured human NK cells. Correlation between surface density of NKp46 and natural cytotoxicity against autologous, allogeneic or xenogeneic target cellsEur J Immunol19992951656166610359120
- VitaleMBottinoCSivoriSNKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysisJ Exp Med199818712206520729625766
- BloushtainNQimronUBar-IlanAMembrane-associated heparan sulfate proteoglycans are involved in the recognition of cellular targets by NKp30 and NKp46J Immunol200417342392240115294952
- HechtMLRosentalBHorlacherTNatural cytotoxicity receptors NKp30, NKp44 and NKp46 bind to different heparan sulfate/heparin sequencesJ Proteome Res20098271272019196184
- Pogge von StrandmannESimhadriVRvon TresckowBHuman leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cellsImmunity200727696597418055229
- BrandtCSBaratinMYiECThe B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humansJ Exp Med200920671495150319528259
- StaggJSmythMJNK cell-based cancer immunotherapyDrug News Perspect200720315516317520092
- DiefenbachAJensenERJamiesonAMRauletDHRae1 and H60 ligands of the NKG2D receptor stimulate tumour immunityNature2001413685216517111557981
- PendeDCantoniCRiveraPRole of NKG2D in tumor cell lysis mediated by human NK cells: cooperation with natural cytotoxicity receptors and capability of recognizing tumors of nonepithelial originEur J Immunol20013141076108611298332
- SobolevOSternPLacy-HulbertAHynesRONatural killer cells require selectins for suppression of subcutaneous tumorsCancer Res20096962531253919258505
- SmythMJSwannJCretneyEZerafaNYokoyamaWMHayakawaYNKG2D function protects the host from tumor initiationJ Exp Med2005202558358816129707
- StreetSEHayakawaYZhanYInnate immune surveillance of spontaneous B cell lymphomas by natural killer cells and gammadelta T cellsJ Exp Med2004199687988415007091
- ZhouSKawakamiSHiguchiYYamashitaFHashidaMThe involvement of NK cell activation following intranasal administration of CpG DNA lipoplex in the prevention of pulmonary metastasis and peritoneal dissemination in miceClin Exp Metastasis2012291637022052391
- KsienzykANeumannBNandakumarRIRF-1 expression is essential for natural killer cells to suppress metastasisCancer Res201171206410641821900395
- ImaiKMatsuyamaSMiyakeSSugaKNakachiKNatural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general populationLancet200035692441795179911117911
- WarrenRPStembridgeAMGardnerEJDeficient immune function of peripheral blood mononuclear cells from patients with Gardner syndromeClin Exp Immunol19856035255313160513
- MarkowitzJFAigesHWCunningham-RundlesSCancer family syndrome: marker studiesGastroenterology19869135815892942437
- JovićVKonjevićGRadulovićSJelićSSpuzićIImpaired perforin-dependent NK cell cytotoxicity and proliferative activity of peripheral blood T cells is associated with metastatic melanomaTumori200187532432911765182
- RichardsJOChangXBlaserBWCaligiuriMAZhengPLiuYTumor growth impedes natural-killer-cell maturation in the bone marrowBlood2006108124625216556890
- ReschnerAHubertPDelvennePBoniverJJacobsNInnate lymphocyte and dendritic cell cross-talk: a key factor in the regulation of the immune responseClin Exp Immunol2008152221922618336590
- KellyJMDarcyPKMarkbyJLInduction of tumor-specific T cell memory by NK cell-mediated tumor rejectionNat Immunol200231839011743585
- MocikatRBraumullerHGumyANatural killer cells activated by MHC class I(low) targets prime dendritic cells to induce protective CD8 T cell responsesImmunity200319456156914563320
- GrimmEAMazumderAZhangHZRosenbergSALymphokine-activated killer cell phenomenon. Lysis of natural killer-resistant fresh solid tumor cells by interleukin 2-activated autologous human peripheral blood lymphocytesJ Exp Med19821556182318416176669
- AtkinsMBLotzeMTDutcherJPHigh-dose recombinant interleukin 2 therapy for patients with metastatic melanoma: analysis of 270 patients treated between 1985 and 1993J Clin Oncol19991772105211610561265
- SosmanJAHankJAMooreKHProlonged interleukin-2 (IL-2) treatment can augment immune activation without enhancing antitumor activity in renal cell carcinomaCancer Invest19919135482012995
- BergerCBergerMHackmanRCSafety and immunologic effects of IL-15 administration in nonhuman primatesBlood2009114122417242619605850
- LauwerysBRGarotNRenauldJCHoussiauFACytokine production and killer activity of NK/T-NK cells derived with IL-2, IL-15, or the combination of IL-12 and IL-18J Immunol200016541847185310925263
- GolabJZagozdzonRKozarKPotentiatied anti-tumor effectiveness of combined therapy with interleukin-12 and mitoxantrone of L1210 leukemia in vivoOncol Rep20007117718110601614
- SondelPMHankJAAntibody-directed, effector cell-mediated tumor destructionHematol Oncol Clin North Am200115470372111676280
- WinterMCHancockBWTen years of rituximab in NHLExpert Opin Drug Saf20098222323519243307
- Garnock-JonesKPKeatingGMScottLJTrastuzumab: a review of its use as adjuvant treatment in human epidermal growth factor receptor 2 (HER2)-positive early breast cancerDrugs201070221523920108993
- Garcia-FoncillasJDiaz-RubioEProgress in metastatic colorectal cancer: growing role of cetuximab to optimize clinical outcomeClin Transl Oncol201012853354220709651
- NavidFSantanaVMBarfieldRCAnti-GD2 antibody therapy for GD2-expressing tumorsCurr Cancer Drug Targets201010220020920201786
- Bekaii-SaabTSRodaJMGuenterbergKDA phase I trial of paclitaxel and trastuzumab in combination with interleukin-12 in patients with HER2/neu-expressing malignanciesMol Cancer Ther20098112983299119887543
- KohCYBlazarBRGeorgeTAugmentation of antitumor effects by NK cell inhibitory receptor blockade in vitro and in vivoBlood200197103132313711342440
- AliciEIPH-2101, a fully human anti-NK-cell inhibitory receptor mAb for the potential treatment of hematological cancersCurr Opin Mol Ther201012672473321154164
- PegramHJRitchieDSSmythMJAlloreactive natural killer cells in hematopoietic stem cell transplantationLeuk Res2011351142120719383
- Van TendelooVFVan de VeldeAVan DriesscheAInduction of complete and molecular remissions in acute myeloid leukemia by Wilms’ tumor 1 antigen-targeted dendritic cell vaccinationProc Natl Acad Sci U S A201010731138241382920631300
- ViaudSTheryCPloixSDendritic cell-derived exosomes for cancer immunotherapy: what’s next?Cancer Res20107041281128520145139
- HallezSSimonPMaudouxFPhase I/II immunogenicity of a Human Papillomavirus (HPV) type 16 E7 protein-based vaccine in women with oncogenic HPV-positive cervical intraepithelial neoplasiaCancer Immunol Immunother200453764265014985860
- HubertPDoyenJCapelleXLocal applications of GM-CSF induce the recruitment of immune cells in cervical low-grade squamous intraepithelial lesionsAm J Reprod Immunol20106412613620367631
- AndresenLSkovbakkeSPerssonG2-Deoxy d-glucose prevents cell surface expression of nkg2d ligands through inhibition of n-linked glycosylationJ Immunol201218841847185522227571
- GasserSOrsulicSBrownEJRauletDHThe DNA damage pathway regulates innate immune system ligands of the NKG2D receptorNature200543670541186119015995699
- LunemannALunemannJDMunzCRegulatory NK-cell functions in inflammation and autoimmunityMol Med2009159–1035235819603102
- ShiFDLjunggrenHGSarvetnickNInnate immunity and autoimmunity: from self-protection to self-destructionTrends Immunol20012229710111286711
- ShahAARosenACancer and systemic sclerosis: novel insights into pathogenesis and clinical implicationsCurr Opin Rheumatol201123653053521825998
- AmosSMDuongCPWestwoodJAAutoimmunity associated with immunotherapy of cancerBlood2011118349950921531979
- ShiFDZhouQNatural killer cells as indispensable players and therapeutic targets in autoimmunityAutoimmunity201144131020701455
- ShiFDWangHBLiHNatural killer cells determine the outcome of B cell-mediated autoimmunityNat Immunol20001324525110973283
- KitaichiNKotakeSMorohashiTOnoeKOhnoSTaylorAWDiminution of experimental autoimmune uveoretinitis (EAU) in mice depleted of NK cellsJ Leukoc Biol20027261117112112488492
- Erkeller-YuselFHulstaartFHannetIIsenbergDLydyardPLymphocyte subsets in a large cohort of patients with systemic lupus erythematosusLupus1993242272318268970
- AramakiTIdaHIzumiYA significantly impaired natural killer cell activity due to a low activity on a per-cell basis in rheumatoid arthritisMod Rheumatol200919324525219283441
- BielekovaBCatalfamoMReichert-ScrivnerSRegulatory CD56(bright) natural killer cells mediate immunomodulatory effects of IL-2Ralpha-targeted therapy (daclizumab) in multiple sclerosisProc Natl Acad Sci U S A2006103155941594616585503
- LiZLimWKMaheshSPLiuBNussenblattRBCutting edge: in vivo blockade of human IL-2 receptor induces expansion of CD56(bright) regulatory NK cells in patients with active uveitisJ Immunol200517495187519115843513
- OgasawaraKHamermanJAHsinHImpairment of NK cell function by NKG2D modulation in NOD miceImmunity2003181415112530974
- NamekawaTSnyderMRYenJHKiller cell activating receptors function as costimulatory molecules on CD4+CD28null T cells clonally expanded in rheumatoid arthritisJ Immunol200016521138114510878393
- MartinMPNelsonGLeeJHCutting edge: susceptibility to psoriatic arthritis: influence of activating killer Ig-like receptor genes in the absence of specific HLA-C allelesJ Immunol200216962818282212218090
- MomotTKochSHunzelmannNAssociation of killer cell immunoglobulin-like receptors with sclerodermaArthritis Rheum20045051561156515146426
- SuzukiYHamamotoYOgasawaraYGenetic polymorphisms of killer cell immunoglobulin-like receptors are associated with susceptibility to psoriasis vulgarisJ Invest Dermatol200412251133113615140215