Abstract
We have previously demonstrated that the small leucine-rich proteoglycan decorin may play a role in adipose tissue homeostasis and the pathophysiology of obesity. Biglycan is highly similar in structure to decorin, therefore we hypothesized it would have a similar expression profile and role to decorin in adipose tissue. Real time polymerase chain reaction was used to measure biglycan mRNA levels in adipose tissue from normal glucose tolerant and impaired glucose tolerant and type 2 diabetic (T2D) Psammomys obesus. Biglycan mRNA was found to be highly expressed in adipose tissue, and gene expression was significantly higher in visceral compared to subcutaneous adipose tissue, with elevated levels in obese, T2D compared to lean normal glucose tolerant P. obesus (P < 0.04). Biglycan mRNA was predominantly expressed by stromal/vascular cells of fractionated adipose tissue (P = 0.023). Biglycan expression in adipose tissue, particularly in the obese state, was markedly upregulated. Collectively, our data suggest that the small leucine-rich proteoglycan family proteins biglycan and decorin may play a role in the development of obesity and T2D, possibly by facilitating expansion of adipose tissue mass.
Introduction
We previously identified the small leucine-rich proteoglycan (SLRP) decorin as a secreted protein not previously associated with the development of obesity, type 2 diabetes (T2D), and the metabolic syndrome.Citation1 Decorin was highly expressed in adipose tissue, with significantly higher gene expression in visceral compared to subcutaneous adipose tissue depots in both Psammomys obesus and human subjects (P = 0.002 and P = 0.001, respectively).Citation1 Furthermore, decorin was shown to be expressed adjacent to blood vessels in the adipose tissue.Citation1 These findings suggest that this SLRP may play a role in adipose tissue homeostasis and in the pathophysiology of obesity.
Biglycan is a SLRP closely related to decorin, with sequence and structure analysis showing the genes are 55% homologous at the amino acid level and are most likely a product of gene duplication.Citation2 Like decorin, biglycan binds transforming growth factor βCitation3 and interacts with collagen.Citation4 The similarity in structure and overlapping expression profiles in skeletal muscle and connective tissues suggest that it is likely that these two proteins function redundantly and synergistically.Citation5–Citation7 The targeted disruption of decorin in mice results in mice with fragile skin, reduced tensile strength, and abnormal collagen morphology,Citation8 whereas biglycan knockout mice have an osteoporosis-like phenotype.Citation9 The biglycan knockout mice are born with no apparent defects however after 3 months they have a reduced growth rate and decreased bone mass suggesting biglycan is a postnatal regulator of skeletal mass.Citation9 Previous studies investigating the biglycan,Citation9 decorin,Citation8 and biglycan/decorinCitation10 deficient mouse models have suggested that these two genes may have redundant functions, and furthermore, may compensate for each other’s functions when one of them is deficient.Citation6,Citation11 Mice deficient in decorin and biglycan exhibit a more severe phenotype than mice deficient in either gene alone.Citation10 Biglycan and decorin therefore may partly rescue or compensate for each other’s absence in the singly deficient mice.Citation7,Citation11–Citation13
Given the expression and functional similarity between decorin and biglycan, the aim of this study was to characterize biglycan in relation to obesity and T2D in P. obesus.
Materials and methods
Experimental animals
A colony of P. obesus were maintained at Deakin University as previously described.Citation1 At 16 weeks of age, animals were classified into three groups according to their blood glucose and plasma insulin concentrations.Citation1,Citation14 The groups were as follows: (1) lean and normal glucose tolerant (NGT), (2) overweight and impaired glucose tolerant (IGT), and (3) obese, T2D. Phenotypic characteristics of each group are previously published.Citation1 At 18 weeks of age, animals were killed by anesthetic overdose (pentobarbitone, 120 mg/kg; Sigma, St Louis, MO) and tissues were rapidly excised, snap frozen in liquid nitrogen, and stored at −80°C for subsequent RNA extraction. All experiments were conducted according to strict National Health and Medical Research Council guidelines and approved by the Deakin University Animal Welfare Committee.
RNA extraction
Total RNA was extracted from P. obesus (n = 5–6 per group) tissues using TRIzol (Invitrogen, Carlsbad, CA) in conjunction with RNeasy columns (Qiagen, Hilden, Germany). The quality and concentration of RNA was determined using the RNA 6000 Nano Assay and an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA). Total RNA was extracted in this study from the following tissues: testes; lung, small and large intestine; adrenal; spleen; kidney; stomach; heart; perirenal, mesenteric, epididymal, intramuscular, and subscapular adipose tissue; soleus, extensor digitorum longus, plantaris, white and red gastrocnemius; liver; brain stem, mid brain, hippocampus, cerebellum, cortex, and hypothalamus. With regards to adipose tissue depots, brown adipose tissue depots were excised and discarded in order to freeze only the white adipose tissue in each depot type. The intramuscular fat originates from a fat depot associated with the gastrocnemius muscle.
Real time polymerase chain reaction
First strand cDNA was generated from RNA using Super-Script First-Strand Synthesis System (Invitrogen) for real time polymerase chain reaction (RT-PCR). Biglycan gene expression levels were quantitated using SYBR Green PCR Master Mix (Applied Biosystems, Worthington, UK) with an ABI PRISM 7700 Sequencing Detector (Perkin Elmer, Waltham, MA). Arbitrary units (AU) were calculated using the delta cycle threshold method. Gene expression was normalized to the group of samples with the lowest mean cycle threshold. RT-PCR primers were as follows: P. obesus biglycan forward 5′-GACAACCGTATCCGCAAAGTG -3′, reverse 5′-GAGCTTCAGGCCATCAAAGG -3′.
Adipose tissue fractionation
P. obesus (n = 5) were sacrificed and the epididymal adipose tissue was removed and digested with type I colleganse digestion as previously described.Citation1 Fractions were frozen in liquid nitrogen for subsequent RNA extraction and RT-PCR as described above.
Statistical analysis
Statistical analysis was performed using SPSS (v 14.0; SPSS Inc, Chicago, IL). Levene’s test for homogeneity of variance was used to determine if variance between two groups was equal. As homogeneity was equal, a one-way analysis of variance with a post hoc least significant difference test was used. Associations between gene expression levels and phenotypic measures were determined using Pearson’s correlation for normally distributed data or Spearman’s correlation for data that were not normally distributed. To compare mean phenotypic values between groups, independent samples t-test was used. Differences and correlations were considered significant at P < 0.05.
Results
Biglycan is highly expressed in adipose tissue from P. obesus
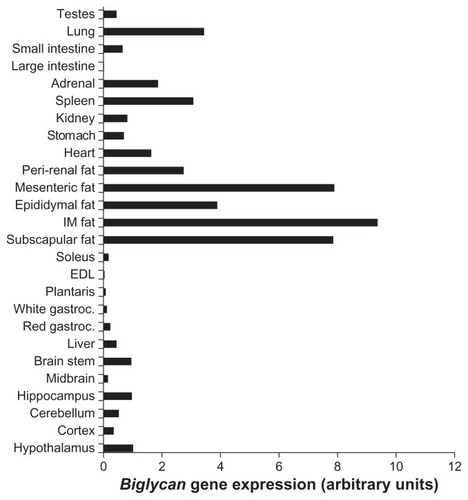
Biglycan gene expression was analyzed in a variety of tissues from P. obesus, an animal model of obesity and T2D. Similar to decorin, biglycan had markedly higher expression within adipose tissue depots compared with all other tissues examined (). Biglycan was also expressed at lower levels in the brain, liver, red gastrocnemius, heart, stomach, kidney, spleen, adrenal, lung, and testes.
Figure 1 Biglycan gene expression in a variety of P. obesus tissues.
Abbreviations: P. obesus, Psammomys obesus; IM, intramuscular; EDL, extensor digitorum longus; gastroc, gastrocnemius; T2D, type 2 diabetes; RT-PCR, real time polymerase chain reaction.
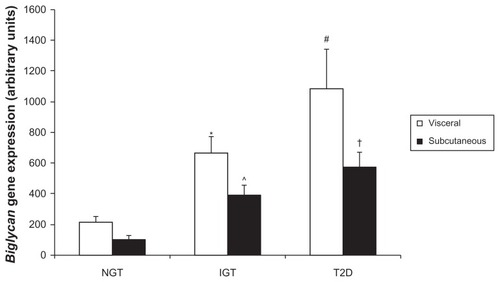
Due to the remarkable expression within adipose tissue depots, biglycan gene expression was next analyzed in visceral (mesenteric) and subcutaneous (subscapular) adipose tissue of P. obesus by RT-PCR. Biglycan expression was higher in visceral compared to subcutaneous adipose tissue overall (P = 0.038, gene expression (mean AU ± standard error of mean): visceral 655 ± 123, subcutaneous 357 ± 61). In P. obesus with impaired glucose tolerance, biglycan gene expression was significantly higher in visceral compared with subcutaneous adipose tissue (P = 0.049, ). Separately, within subcutaneous tissue, biglycan gene expression was significantly elevated in IGT and T2D P. obesus compared with healthy (NGT) littermates (P = 0.009 and P < 0.001, respectively) and significantly correlated with body weight (r2 = 0.38, P = 0.012), plasma glucose, and insulin concentration (r2 = 0.38, P = 0.011 and r2 = 0.29, P = 0.031, respectively), and percentage body fat (r2 = 0.50, P = 0.003). Biglycan gene expression was significantly elevated in visceral T2D compared to NGT (P = 0.002) and was significantly correlated with body weight (r2 = 0.31, P = 0.024), plasma glucose concentration (r2 = 0.46, P = 0.004), and percentage body fat (r2 = 0.38, P = 0.014) in visceral adipose tissue. This expression profile was similar to that of decorin.
Figure 2A Analysis of biglycan gene expression in P. obesus visceral and subcutaneous adipose tissue.
Abbreviations: P. obesus, Psammomys obesus; NGT, normal glucose tolerant; IGT, impaired glucose tolerant; T2D, type 2 diabetes; RT-PCR, real time polymerase chain reaction; SEM, standard error of mean.
Biglycan gene expression was measured in the same NGT, IGT, and T2D mesenteric and subscapular adipose tissue samples as decorin. This allowed us to analyze a potential association between the two genes during the development of obesity and T2D. Bivariate analysis revealed a strong correlation between decorin and biglycan gene expression (r2 = 0.86, P < 0.001) during the development of obesity and T2D in P. obesus mesenteric and subscapular adipose tissue.
Biglycan is predominantly expressed within stromal/vascular cells in fractionated adipose tissue
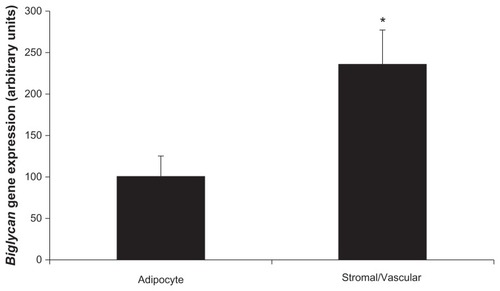
To determine which cells in adipose tissue express biglycan, gene expression was measured in fractionated epididymal adipose tissue from IGT P. obesus. There were two cellular populations – adipocytes and stromal/vascular cells. Comparable to decorin which had almost exclusive expression within stromal/vascular cells, biglycan was predominantly expressed in stromal/vascular cells compared to adipocytes (P = 0.023, ). Successful fractionation was confirmed using adipocyte-specific (leptin) and stromal/vascular-specific (CD68, Pref-1) markers. As expected, leptin gene expression was significantly higher in the adipocyte compared to the stromal/vascular fraction (P < 0.001), and CD68/Pref-1 gene expression was significantly higher in the stromal/vascular compared to adipocyte fraction (P < 0.001; data not shown).
Figure 2B Biglycan expression in fractionated P. obesus epididymal adipose tissue.
Abbreviations: P. obesus, Psammomys obesus; RT-PCR, real time polymerase chain reaction; SEM, standard error of mean.
Discussion
We have previously shown the SLRP decorin to be highly expressed in adipose tissue, and upregulated in obesity suggesting a potential role in adipose tissue homeostasis or in the pathophysiology associated with obesity such as extracellular matrix (ECM) remodeling, inflammation, and angiogenesis within expanding adipose tissue. Here we extend these findings to include the closely related SLRP, biglycan, which also exhibits high expression in adipose tissue (and particularly visceral adipose tissue) compared to a wide variety of tissues examined, and increased expression in obesity.
Collectively, these data raise the possibility that increased expression of SLRPs in adipose tissue is a key feature during the development of obesity, T2D, and the metabolic syndrome. SLRPs are constituents of the ECM which undergoes constant degradation and renewal,Citation15 allows the diffusion of nutrients, metabolites, and hormones between blood and cells within the tissueCitation16 and functions not only as a scaffold for cells within tissues, but also as a reservoir of growth factors and cytokines and modulates their activities.Citation17 SLRPs bind and interact with numerous proteins involved in matrix assembly, cell proliferation, and tissue morphogenesis. Citation18 With regards to adipocyte differentiation, remarkable changes in cell morphology, cytoskeletal components, and the amount and type of ECM components secreted occurs.Citation19 Furthermore, the development of obesity requires extensive reorganization of adipose tissue and the expression of genes involved in matrix remodeling was found to be increased in obese db/db mice and resulted in the net accumulation of ECM components in parallel to increased adipocyte size.Citation20 It has also been proposed that complications of diabetes are closely associated with changes in the ECMCitation21,Citation22 as macroangiopathic and microangiopathic complications resultant from the presence of diabetes have involved abnormalities within the extracellular intima.Citation21
Given the above background information regarding SLRPs, it is therefore tempting to speculate that SLRPs may play a role in angiogenesis and ECM remodeling within expanding adipose tissue during the development of obesity and T2D. Recent studies have demonstrated biglycan to be upregulated in a murine model of diet-induced obesity with atherosclerosis,Citation23 to be involved in vascular and proatherogenic remodeling in a rat model of metabolic syndrome,Citation24 to be proangiogenic,Citation25 and interestingly, to have upregulated expression in parallel with matrix degradation genes in white adipose tissue from db/db mice fed a high fat diet.Citation20 Biglycan has also been demonstrated to be upregulated in human omental compared to lean adipose tissue.Citation26 This evidence builds upon the hypothesized role for biglycan in adipose tissue homeostasis, particularly in states of obesity where ECM remodeling is required for adipose tissue expansion.
Biglycan has been shown to be proinflammatory and acts through toll-like receptors in macrophages.Citation27–Citation29 Recent studies have supported a strong association of biglycan with inflammation, demonstrating increased levels of tumor necrosis factor α (a major inflammatory cytokine implicated in T2D) and biglycan in human omental adipose tissue, suggesting they may be coregulated.Citation26 This potentially associates biglycan to obesity which is associated with a chronic low grade inflammatory state in adipose tissue (particularly visceral adipose tissue),Citation30,Citation31 and the subsequent development of insulin resistance and T2D.Citation30 However, these are postulated theories, therefore additional experiments to explore a role in inflammation and substantiate these theories are now required.
In summary, our data suggest that the family of SLRPs may play a role in the development of obesity and T2D, with a hypothesized role of facilitating the expansion of adipose tissue mass within an animal model. Further work is now required to corroborate these results within humans.
Acknowledgments
This work was funded by Verva Pharmaceuticals Ltd. This manuscript contains original research and has not been previously published in another journal.
Disclosure
The authors of this manuscript have no conflicts of interest to disclose and have approved the final article submitted to Biologics: Target and Therapeutics. Author contribution as follows: KB project design, data collection and analysis, preparation of manuscript; DS/KW: project design and editorial assistance with manuscript. All authors have read and approved this manuscript.
References
- BoltonKSegalDMcMillanJDecorin is a secreted protein associated with obesity and type 2 diabetesInt J Obes (Lond)20083271113112118414424
- FisherLWTermineJDYoungMFDeduced protein sequence of bone small proteoglycan I (biglycan) shows homology with proteoglycan II (decorin) and several nonconnective tissue proteins in a variety of speciesJ Biol Chem19892648457145762647739
- HildebrandARomarisMRasmussenLMInteraction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor betaBiochem J1994302Pt 25275348093006
- SchonherrEWitsch-PrehmPHarrachBRobenekHRauterbergJKresseHInteraction of biglycan with type I collagenJ Biol Chem19952706277627837852349
- BiYStueltenCHKiltsTExtracellular matrix proteoglycans control the fate of bone marrow stromal cellsJ Biol Chem200528034304813048915964849
- YoungMFBiYAmeyeLChenXDBiglycan knockout mice: new models for musculoskeletal diseasesGlycoconj J2002194–525726212975603
- WadhwaSBiYOrtizATImpaired posterior frontal sutural fusion in the biglycan/decorin double deficient miceBone200740486186617188951
- DanielsonKGBaribaultHHolmesDFGrahamHKadlerKEIozzoRVTargeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragilityJ Cell Biol199713637297439024701
- XuTBiancoPFisherLWTargeted disruption of the biglycan gene leads to an osteoporosis-like phenotype in miceNat Genet199820178829731537
- CorsiAXuTChenXDPhenotypic effects of biglycan deficiency are linked to collagen fibril abnormalities, are synergized by decorin deficiency, and mimic Ehlers-Danlos-like changes in bone and other connective tissuesJ Bone Miner Res20021771180118912102052
- AmeyeLYoungMFMice deficient in small leucine-rich proteoglycans: novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome, muscular dystrophy, and corneal diseasesGlycobiology2002129107R116R
- ChenXDFisherLWRobeyPGYoungMFThe small leucine-rich proteoglycan biglycan modulates BMP-4-induced osteoblast differentiationFaseb J200418994895815173106
- CasarJCMcKechnieBAFallonJRYoungMFBrandanETransient up-regulation of biglycan during skeletal muscle regeneration: delayed fiber growth along with decorin increase in biglycan-deficient miceDev Biol2004268235837115063173
- BarnettMCollierGRCollierFMZimmetPO’DeaKA cross-sectional and short-term longitudinal characterisation of NIDDM in Psammomys obesusDiabetologia19943776716767958537
- RahmaniMWongBWAngLVersican: signaling to transcriptional control pathwaysCan J Physiol Pharmacol2006841779216845893
- AlbertsBBrayDLewisJRaffMRobertsKWatsonJDMolecular Biology of the Cell3rd edGarland Publishing1994
- KresseHSchonherrEProteoglycans of the extracellular matrix and growth controlJ Cell Physiol2001189326627411748584
- IozzoRVMurdochADProteoglycans of the extracellular environment: clues from the gene and protein side offer novel perspectives in molecular diversity and functionFaseb J19961055986148621059
- SmasCMSulHSControl of adipocyte differentiationBiochem J1995309Pt 36977107639681
- HuberJLofflerMBilbanMPrevention of high-fat diet-induced adipose tissue remodeling in obese diabetic mice by n-3 polyunsaturated fatty acidsInt J Obes (Lond)20073161004101317130847
- OlssonUBondjersGCamejoGFatty acids modulate the composition of extracellular matrix in cultured human arterial smooth muscle cells by altering the expression of genes for proteoglycan core proteinsDiabetes199948361662210078565
- LehtiTMSilvennoinenMKivelaRKainulainenHKomulainenJEffects of streptozotocin-induced diabetes and physical training on gene expression of extracellular matrix proteins in mouse skeletal muscleAm J Physiol Endocrinol Metab20062905E90090716352670
- KingVLHatchNWChanHWde BeerMCde BeerFCTannockLRA murine model of obesity with accelerated atherosclerosisObesity (Silver Spring)2010181354119498343
- MangatRSuJScottPGRussellJCVineDFProctorSDChylomicron and apoB48 metabolism in the JCR:LA corpulent rat, a model for the metabolic syndromeBiochem Soc Trans200735Pt 347748117511632
- ZhangWChuangYJJinTAntiangiogenic antithrombin induces global changes in the gene expression profile of endothelial cellsCancer Res200666105047505516707426
- WardMAjuwonKMRegulation of pre-adipocyte proliferation and apoptosis by the small leucine-rich proteoglycans, biglycan and decorinCell Prolif201144434335121702857
- MorethKBrodbeckRBabelovaAThe proteoglycan biglycan regulates expression of the B cell chemoattractant CXCL13 and aggravates murine lupus nephritisJ Clin Invest2010120124251427221084753
- BabelovaAMorethKTsalastra-GreulWBiglycan, a danger signal that activates the NLRP3 inflammasome via toll-like and P2X receptorsJ Biol Chem200928436240352404819605353
- SchaeferLBabelovaAKissEThe matrix component biglycan is proinflammatory and signals through Toll-like receptors 4 and 2 in macrophagesJ Clin Invest200511582223223316025156
- Torres-LealFLFonseca-AlanizMHRogeroMMTirapeguiJThe role of inflamed adipose tissue in the insulin resistanceCell Biochem Funct201028862363121104928
- WellenKEHotamisligilGSInflammation, stress, and diabetesJ Clin Invest200511551111111915864338