
Abstract
Purpose
Cutaneous squamous cell carcinoma (cSCC) is a skin malignant tumor account for approximately one-third of all nonmelanoma skin cancers. Studies have shown that TEA domain transcription factor 1 (TEAD1) is discovered to be involved in the pathogenesis of some human cancers, but to our knowledge its role in cSCC has not been reported.
Patients and Methods
Samples from 16 cSCC patients and 27 healthy individuals were obtained for immunohistochemical staining of TEAD1. The expressions of TEAD1 in SCL-1, HSC-1 cells compared with the primary neonatal human epithelial keratinocytes were detected by Western blot and RT-qPCR. Proliferation and cell cycle of TEAD1 knockdown in cSCC cell lines were examined by MTT and flow cytometry analysis. Annexin V/PI and JC-1 staining were used to determine the cell apoptosis.
Results
The expression of TEAD1 decreased significantly in cSCC compared to its expression in normal skin tissues and cell lines. Down-regulation of TEAD1 in cSCC cell lines promoted cell growth via regulation of the G2/M progression. Additionally, silence of TEAD1 also protected cells against 5-Fluorouracil-induced apoptosis and decreased the expression of apoptosis-related protein (p53).
Conclusion
Our results suggested that TEAD1 expression is down-regulated and functioned as a tumor suppressor in cSCC and that it may serve as a biomarker or therapeutic target of cSCC.
Introduction
Cutaneous squamous cell carcinomas (cSCC) are originated from the keratinocytes located in the epidermis or adnexal structures. They account for approximately one-third of all nonmelanoma skin cancers (NMSCs), with its incidence rising steeply for decades.Citation1 About 2% of cSCCs metastasize and their 5-year disease-specific survival was 79.1%.Citation2 More than two-thirds of patients with cSCC die from local invasion or nodal metastases, rather than distant organ metastases.Citation3
In unresectable and metastatic cases, the treatment remains challenging. Targeted therapy with anti-EGFR (epidermal growth factor receptor) antibodies and PD-1 immune checkpoint inhibitors have been used in advanced cSCC.Citation4 However, each of these methods has its own limitations. EGFR inhibitors have activity only in a subset of patients. Primary or secondary resistance is a problem that cannot be ignored for PD-1 immune checkpoint inhibitors.Citation5 Further studies are needed to search for more predictive biomarkers and better therapeutic targets.
Hippo pathway is an important regulator of tumorigenesis.Citation6 TEAD1 (TEA domain transcription factor 1) proteins belong to a family of 4 transcription factors (TEAD1–4), which are major downstream nuclear effectors of the Hippo signaling pathway.Citation7 Previous research reported TEAD1 enhanced cell proliferation in colorectal cancerCitation8 and mediated oncogenic activities in osteosarcoma.Citation9 Nevertheless, to our knowledge, the role of TEAD family in cSCC has not been investigated. In this study, we examined TEAD1 expression in cSCC, as well as the biological function of TEAD1 by a knockdown strategy.
Materials and Methods
Patient Samples
Patient samples were acquired from the tissue bank of the Department of Dermatology at the Second Affiliated Hospital of Xi’an Jiaotong University. There were a total of 43 samples collected, including 16 cSCCs (9 males and 7 females, 49 to 83 years) and 27 normal skin tissues (17 males and 10 females, 44 to 80 years) obtained from cosmetic surgery. All biopsies were done under local anesthesia. Prior to the study, written informed consent for organizational procurement was obtained from all patients, and ethics approval was received from the Institutional Ethics Committee of Xi’an Jiaotong University. The studies were conducted in accordance with Declaration of Helsinki guidelines.
IHC
Immunohistochemical staining was performed according to standard protocol.Citation10 Based on the following scoring system, two independent pathologists assessed and quantified the staining results under microscopes. Firstly, a positive rate score was estimated by the positive cell percentage (5% ¼ 0, 5%~25% ¼ 1, 25% ~50% ¼ 2, 50%~75% ¼ 3, >75% ¼ 4). Next, a score of staining intensity was given by objective colorless (colorless ¼ 0, yellow ¼ 1, yellowish brown ¼ 2, chocolate brown ¼ 3). Finally, the total score is added by aforementioned two scores. The average score of five fields was taken as the final score of TEAD1 expression for each slide.
Cell Culture
The human cSCC cell line SCL-1Citation11 and HSC-1Citation12 cells were from Beijing Beinachuanglian Biotechnology Research Institute (Beijing, China). The cells were maintained in Dulbecco’s Modified Eagle’s Medium supplemented with 10% fetal bovine serum (FBS), penicillin (100 units/mL), and streptomycin (100 mg/mL). The cells were conventionally cultured at 37°C and 5% CO2 in a humidified incubator. Lysates of primary neonatal human epithelial keratinocytes (HEKn cells) were kindly provided by Dr. Hao Wang at Xi’an Jiaotong University.
Transient Transfection of TEAD1 siRNA
The following siRNA oligonucleotides were purchased from GenePharma (Shanghai, China): TEAD1-1 siRNA (5′-CCACUGCCAUUCAUAACAATT-3′), TEAD1-2 siRNA (5′-CUGCAGAUAAGCCAAUUGATT-3′). Cells were transfected with siRNA according to the procedure recommended by IMAX transfection reagent (Invitrogen, Carlsbad, CA). 100,000/well cSCC cells were plated into 6 well plate 24 hours before transfection.
MTT Assay
Cell proliferation was detected by MTT assay. Cells were seeded in 96-well plates with a density of 4*103 cells/well and transfected with siRNA 24 hours later. Then, relative cell numbers were measured every 24 hours by incubating cells with 0.4 mg/mL MTT followed by optical density (OD) reading. Average readings from three independent experiments were plotted at each time point. Six technical replicates were performed for each data point.
Cell Cycle Analysis
Cells were harvested and fixed with 75% ethanol at 4°C overnight. For the analysis, PI staining solution (50 mg/mL PI and 100 mg/mL ribonuclease A) was added to the cells and incubated for 30 minutes in darkness at 37°C. The cells were analyzed using NovoCyte flow cytometry (ACEA Biosciences). Three independent experiments were conducted.
Annexin V/PI Staining
The siRNA-transfected cells were treated with 5 ug/mL of 5-Fu for 48 hours before harvest. For the analysis, cells were stained with the annexin V-fluorescein isothiocyanate/PI Apoptosis Detection Kit (BD Pharmingen), following the manufacturer’s instructions. Stained cells were analyzed by NovoCyte flow cytometry (ACEA Biosciences). Three independent experiments were performed.
JC-1 Staining
The loss of MMP (ΔΨm), a hallmark of apoptosis, was measured using the Mitochondrial membrane potential assay kit with JC-1 from Beyotime (Shanghai, China) according to the manufacturer’s instructions. After 5 ug/mL of 5-Fu treatment of 48 hours, cells were treated with JC-1 probe (2 μg/mL) for 30 min at 37°C and measured at an excitation wavelength of 490 nm and an emission wavelength of 530/590 nm by a spectrophotometer (FLUOstar Omega, Labtech). Then, the ΔΨm was displayed by a change in the ratio of red (aggregated JC-1) and green (monomeric JC-1) fluorescence. Three independent experiments were performed.
Western Blot Analysis
Protein expression levels were analyzed by standard Western blotting protocols. The following antibodies were used in this study: TEAD1 (ab133533) was purchased from Abcam (Cambridge, UK). Cyclin B1 (sc-595) was purchased from Santa Cruz Biotechnology (Dallas, TX, USA). P53 (10442-1-AP) was from Proteintech (Chicago, IL, USA). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (AC001) was from Abclonal (China).
RT-qPCR
Total RNA was extracted from the cells using the Trizol reagent (Invitrogen) and then reverse transcribed into complementary DNA. The primer oligonucleotide sequences were as follows: human TEAD1: forward 5′-TTCGCCACGTGTGTTTGTTC-3′ and reverse 5′- ACGCTTACATCACAAGCCCA-3′, human GAPDH: forward 5′-CTCCTCCACCTTTGACGCTG-3′ and reverse 5′-TCCTCTTGTGCTCTTGCTGG −3′. GAPDH was used as an internal control, and data were analyzed by the 2-ΔΔCT method.
Statistical Analysis
All data were expressed by the standard error of the mean (SEM). SPSS statistical software package 26 (SPSS, Chicago, IL, USA) was used for data analysis. The groups were compared by chi-square tests (χ2) or independent-samples t-tests (t). P < 0.05 was considered statistically significant.
Results
TEAD1 Expression is Down-Regulated in cSCC
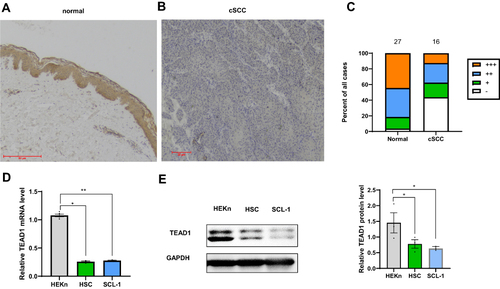
Immunohistochemistry was used to investigate TEAD1 protein expression in tissue samples from patients, including normal skin tissues (n = 27), cSCC (n = 16). The results showed cytoplasmic staining of TEAD1 (). The positive TEAD1 staining was significantly lower in the cSCC group than the control group (χ2 = 8.159, P ≤ 0.01) (, ). In addition, in contrast to the primary neonatal human epithelial keratinocytes (), TEAD1 RNA and protein expression were down-regulated in two cSCC cell lines, HSC-1, and SCL-1.
Table 1 Positive Expression Rates of TEAD1 (%)
Figure 1 Analysis of TEAD1 expression in different tissues and cells. (A) Normal skin (n=27). Bar length =50μm. (B) cSCC (n =16). Bar length =20μm. (C) Semiquantitative analysis of TEAD1 staining. (D) qPCR analysis of TEAD1 RNA expression in HEKn and two cSCC cell lines: HSC-1 and SCL-1 (E) Western blot analysis of TEAD1 protein expression in HEKn and two cSCC cell lines: HSC-1 and SCL-1. All the quantitative data are presented as mean standard error of the mean (SEM). *P < 0.05, **P < 0.01.
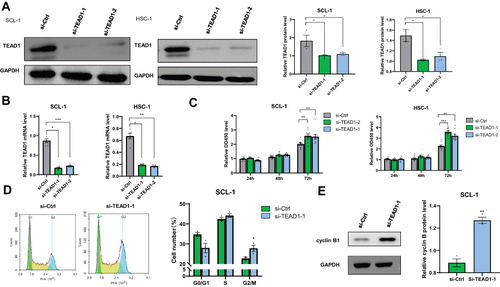
Si-TEAD1 Promotes cSCC Cell Growth via Regulation of the G2/M Progression
As for decreased expression of TEAD1 in cSCC skin lesions, we speculated that knockdown of TEAD1 may promote development of cSCC. Small-interfering RNAs (siRNAs) were used to knockdown TEAD1 in two cSCC cell lines, SCL-1 and HSC-1 (). Next, we determined cell proliferation and cell cycle by MTT and propidium iodide (PI) staining, respectively. The results showed that depletion of TEAD1 promoted cell growth in both cell lines (). In addition, TEAD1 knockdown prompted cell growth via the G2/M stage transition (), with the significant elevation of cell cycle regulator cyclin B1 ().
Figure 2 Effect of down-regulation TEAD1 on cell proliferation and cell cycle. (A and B) TEAD1 RNA and protein expression (48 hours after transfection) was measured after TEAD1-siRNA interference. (C) Cells were transfected with TEAD1 siRNA, and MTT assay was performed after 24, 48, and 72 hours. (D) The cell cycle profiles were analyzed 48 hours after siRNA by flow cytometry. (E) Cell cycle regulators were analyzed by Western blot analysis after TEAD1 knockdown, with GAPDH being an internal control. All the quantitative data are presented as mean standard error of the mean (SEM). *P < 0.05, **P < 0.01, ***P < 0.001.
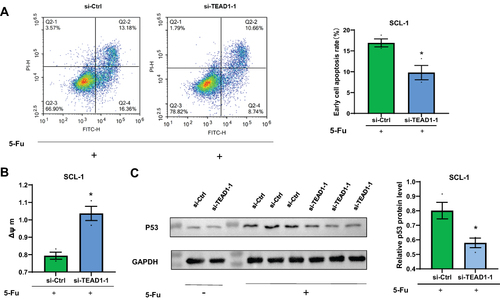
Si-TEAD1 Protects cSCC Cells Against Apoptosis
We explored the influence of TEAD1 on regulation of apoptosis, via annexin V/PI and JC-1 staining. Considering the low basal apoptosis levels, we firstly used 5-fluorouracil (5-Fu) to induce cell apoptosis. As expected, 5-Fu treatment increased apoptosis rate of SCL-1 cells. Then, we found that, compared to the control group, knockdown of TEAD1 reduced the apoptosis rate, which was promoted by 5-Fu () and inhibited the activation of p53 () in SCL-1 cells. JC-1 dye was adopted to detect the depolarization of the inner mitochondrial membrane, which was a signal of apoptosis initiation. Similarly, si-TEAD1 increased mitochondrial membrane potential against 5-Fu induced apoptosis compared with the control group (). Therefore, si-TEAD1 protected cSCC cells against 5-Fu induced apoptosis.
Figure 3 Effect of down-regulation of TEAD1 on apoptosis. (A) Cells were transfected with si-TEAD1 for 24 hours before the addition of 5 ug/mL of 5-fluorouracil. After another 48 hours, cell apoptosis was analyzed by annexin V/propidium iodide staining and flow cytometry analysis. (B) Cell apoptosis of (A) was analyzed by JC-1 (Δψ m). (C) Western blot analysis on indicated proteins. GAPDH was used as an internal control. All the quantitative data were presented as the standard error of the mean (SEM). *P < 0.05, **P < 0.01.
Discussion
TEAD1 is discovered to be involved in the pathogenesis of several kinds of human cancers.Citation8,Citation9,Citation13,Citation14 But its role in cSCC is still unclear. Here, we investigated the functional role of TEAD1 in cSCC. As a result, the TEAD1 was proved to be down-regulated in cSCC patient samples and cell lines. Knockdown of TEAD1 cSCC promotes cell survival and resistance to 5-Fu.
Although the central components of Hippo pathway are well defined, factors modulating Hippo transcriptional outputs remain elusive. Hippo effectors YAP and TAZ were co-activators for the TEAD family of transcription factors (TEAD1-4). YAP and TAZ can act either as tumor suppressors in the cytoplasm or tumor enhancer in the nucleus.Citation15–19 Based on opposite pro- or anti-cancer YAP activity, cancers can be classified into YAPon or YAPoff.Citation20 However, the expression patterns of YAP and TAZ were not always the same, for example, seven cell lines of small cell lung cancer were YAPoff but one of them expressed TAZ.Citation20
Previous study has manifested that Hippo pathway is essential for skin homeostasis and epithelial stem cell maintenance. TEAD transcriptional-network, downstream of Hippo pathway, was reported to be down-regulated during keratinocyte differentiation.Citation21,Citation22 In primary human keratinocytes, TEAD1 and TEAD3 work crucially in the regulation of human epidermal proliferation and compensate for each other.Citation23
The expression of TEAD1 decreased significantly in cSCC and was mainly localized in the cytoplasm according to our study. Significantly, these results are consistent with the known tumor suppressor function of its subcellular localization. Recent studies indicated that YAP1 and TAZ both contribute to skin squamous (SCC) and basal (BCC) cell carcinoma formation.Citation24,Citation25 YAP1 has been identified as an oncogene in cSCC, which is expressed mainly in the nucleus.Citation26 It is possible that the function of TEAD1 may be independent of YAP1 because of different subcellular localizations. A similar example is that TEAD suppresses IL6 through the mechanism, which is independent of functional TEAD-YAP activity.Citation27 Therefore, there is abundant space for further exploration in investigating whether TEAD1 proteins make cytoplasmic translocationCitation28 in cSCC.
Our data also discovered that si-TEAD1 promoted cSCC cell growth by regulating the G2/M transition, and TEAD1 depletion increased the expression of cyclin B1, the key regulator of the G2/M cell cycle transition.Citation29 Similar regulation of cell proliferation by YAP/TEAD activity has been observed in other studies. For example, expressing YAP/TEAD inhibited 2D or 3D growth in retinoblastoma and reduced tumor load in small-cell lung cancer.Citation20
Mitochondrial membrane potential (Δψ m) is an indicator of mitochondrial state. Loss of Δψ is an early event in apoptosis.Citation30 Our results of JC-1 and Annexin V/PI staining support the anti-apoptotic role of si-TEAD1 against 5-Fu. 5-Fu, a widely used chemotherapy drug, can induce p53 activation and p53-dependent apoptosis.Citation31–33 Thus, we further detected p53 expression in TEAD1 knockdown cells with 5-Fu treatment. As expected, Western blot analysis indicates that si-TEAD1 suppress p53 activation increased by 5-Fu. Interestingly, in addition to inducing apoptosis by TEAD1 in cardiomyocytes,Citation33 YAP/TAZ-TEAD expression is inversely proportional to apoptosis in existing research.Citation26 Its exact mechanism of action needs to be investigated in further studies.
Conclusion
In conclusion, our study indicated that TEAD1 plays a crucial role in human cSCC. Restoration of protein expression of TEAD1 may reduce tumor cell survival and drug resistance of 5-Fu in cSCC patients. In other words, it sheds light on another carcinogenic mechanism in the development of cSCC as well as novel target for treatment and prognostic purpose.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
We thank Dr. Hao Wang at Xi’an Jiaotong University for providing the primary neonatal human epithelial keratinocyte lysates. This study was supported by grants from the National Natural Science Foundation of China (81573055, 81773328).
References
- Chang MS, Azin M, Demehri S. Cutaneous squamous cell carcinoma: the frontier of cancer immunoprevention. Annu Rev Pathol. 2022;17:101–119. doi:10.1146/annurev-pathol-042320-120056
- Venables ZC, Autier P, Nijsten T, et al. Nationwide incidence of metastatic cutaneous squamous cell carcinoma in England. JAMA Dermatol. 2019;155(3):298–306. doi:10.1001/jamadermatol.2018.4219
- Eigentler TK, Leiter U, Häfner H-M, Garbe C, Röcken M, Breuninger H. Survival of patients with cutaneous squamous cell carcinoma: results of a prospective cohort study. J Invest Dermatol. 2017;137(11):2309–2315. doi:10.1016/j.jid.2017.06.025
- Peris K, Piccerillo A, Del Regno L, Di Stefani A. Treatment approaches of advanced cutaneous squamous cell carcinoma. J Eur Acad Dermatol Venereol. 2022;36(Suppl 1):19–22. doi:10.1111/jdv.17400
- Vesely MD, Zhang T, Chen L. Resistance Mechanisms to Anti-PD Cancer Immunotherapy. Annu Rev Immunol. 2022;40:45–74. doi:10.1146/annurev-immunol-070621-030155
- Sanchez-Vega F, Mina M, Armenia J, et al. Oncogenic signaling pathways in the cancer genome atlas. Cell. 2018;173(2):321–337.
- Osman I, He X, Liu J, et al. TEAD1 (TEA Domain Transcription Factor 1) promotes smooth muscle cell proliferation through upregulating SLC1A5 (Solute Carrier Family 1 Member 5)-mediated glutamine uptake. Circ Res. 2019;124(9):1309–1322. doi:10.1161/CIRCRESAHA.118.314187
- Yu M-H, Zhang W. TEAD1 enhances proliferation via activating SP1 in colorectal cancer. Biomed Pharmacother. 2016;83:496–501. doi:10.1016/j.biopha.2016.06.058
- Chai J, Xu S, Guo F. TEAD1 mediates the oncogenic activities of Hippo-YAP1 signaling in osteosarcoma. Biochem Biophys Res Commun. 2017;488(2):297–302. doi:10.1016/j.bbrc.2017.05.032
- Pinheiro C, Longatto-Filho A, Scapulatempo C, et al. Increased expression of monocarboxylate transporters 1, 2, and 4 in colorectal carcinomas. Virchows Arch. 2008;452(2):139–146. doi:10.1007/s00428-007-0558-5
- Boukamp P, Tilgen W, Dzarlieva RT, et al. Phenotypic and genotypic characteristics of a cell line from a squamous cell carcinoma of human skin. J Natl Cancer Inst. 1982;68(3):415–427.
- Kondo S, Aso K. Establishment of a cell line of human skin squamous cell carcinoma in vitro. Br J Dermatol. 1981;105(2):125–132. doi:10.1111/j.1365-2133.1981.tb01196.x
- Zhou Y, Huang T, Zhang J, et al. TEAD1/4 exerts oncogenic role and is negatively regulated by miR-4269 in gastric tumorigenesis. Oncogene. 2017;36(47):6518–6530. doi:10.1038/onc.2017.257
- Holden JK, Cunningham CN. Targeting the hippo pathway and cancer through the TEAD family of transcription factors. Cancers. 2018;10(3). doi:10.3390/cancers10030081
- Sebio A, Lenz H-J. Molecular pathways: hippo signaling, a critical tumor suppressor. Clin Cancer Res. 2015;21(22):5002–5007. doi:10.1158/1078-0432.CCR-15-0411
- Yuan M, Tomlinson V, Lara R, et al. Yes-associated protein (YAP) functions as a tumor suppressor in breast. Cell Death Differ. 2008;15(11):1752–1759. doi:10.1038/cdd.2008.108
- Huang H, Zhang W, Pan Y, et al. YAP suppresses lung squamous cell carcinoma progression via deregulation of the DNp63-GPX2 axis and ROS accumulation. Cancer Res. 2017;77(21):5769–5781. doi:10.1158/0008-5472.CAN-17-0449
- Jho E. Dual role of YAP: oncoprotein and tumor suppressor. J Thorac Dis. 2018;10(Suppl 33):S3895–S3898. doi:10.21037/jtd.2018.10.70
- Lin Z, Guo H, Cao Y, et al. Acetylation of VGLL4 regulates Hippo-YAP signaling and postnatal cardiac growth. Dev Cell. 2016;39(4):466–479. doi:10.1016/j.devcel.2016.09.005
- Pearson JD, Huang K, Pacal M, et al. Binary pan-cancer classes with distinct vulnerabilities defined by pro- or anti-cancer YAP/TEAD activity. Cancer Cell. 2021;39(8):1115–1134.e12. doi:10.1016/j.ccell.2021.06.016
- Yuan Y, Park J, Feng A, et al. YAP1/TAZ-TEAD transcriptional networks maintain skin homeostasis by regulating cell proliferation and limiting KLF4 activity. Nat Commun. 2020;11(1):1472. doi:10.1038/s41467-020-15301-0
- Li L, Wang Y, Torkelson JL, et al. TFAP2C- and p63-dependent networks sequentially rearrange chromatin landscapes to drive human epidermal lineage commitment. Cell Stem Cell. 2019;24(2):271–284.e8. doi:10.1016/j.stem.2018.12.012
- Li J, Tiwari M, Xu X, Chen Y, Tamayo P, Sen GL. TEAD1 and TEAD3 play redundant roles in the regulation of human epidermal proliferation. J Invest Dermatol. 2020;140(10):S105. doi:10.1016/j.jid.2020.03.820
- Debaugnies M, Sánchez-Danés A, Rorive S, et al. YAP and TAZ are essential for basal and squamous cell carcinoma initiation. EMBO Rep. 2018;19(7). doi:10.15252/embr.201845809
- Miranda MM, Lowry WE. Hip to the Game: YAP/TAZ is required for nonmelanoma skin cancers. EMBO J. 2018;37(17). doi:10.15252/embj.201899921
- Jia J, Li C, Luo S, et al. Yes-associated protein contributes to the development of human cutaneous squamous cell carcinoma via activation of RAS. J Invest Dermatol. 2016;136(6):1267–1277. doi:10.1016/j.jid.2016.02.005
- Kim T, Yang S-J, Hwang D, et al. A basal-like breast cancer-specific role for SRF-IL6 in YAP-induced cancer stemness. Nat Commun. 2015;6:10186. doi:10.1038/ncomms10186
- Lin KC, Moroishi T, Meng Z, et al. Regulation of Hippo pathway transcription factor TEAD by p38 MAPK-induced cytoplasmic translocation. Nat Cell Biol. 2017;19(8):996–1002. doi:10.1038/ncb3581
- Wei D, Parsels LA, Karnak D, et al. Inhibition of protein phosphatase 2A radiosensitizes pancreatic cancers by modulating CDC25C/CDK1 and homologous recombination repair. Clin Cancer Res. 2013;19(16):4422–4432. doi:10.1158/1078-0432.CCR-13-0788
- Troiano L, Ferraresi R, Lugli E, et al. Multiparametric analysis of cells with different mitochondrial membrane potential during apoptosis by polychromatic flow cytometry. Nat Protoc. 2007;2(11):2719–2727. doi:10.1038/nprot.2007.405
- Shin Y, Lim H, Choi B-S, et al. Highly activated p53 contributes to selectively increased apoptosis of latently HIV-1 infected cells upon treatment of anticancer drugs. Virol J. 2016;13(1):141. doi:10.1186/s12985-016-0595-2
- Vemula S, Ramdas B, Hanneman P, Martin J, Beggs HE, Kapur R. Essential role for focal adhesion kinase in regulating stress hematopoiesis. Blood. 2010;116(20):4103–4115. doi:10.1182/blood-2010-01-262790
- Hsu TH, Jiang SY, Chang WL, et al. Involvement of RARRES3 in the regulation of Wnt proteins acylation and signaling activities in human breast cancer cells. Cell Death Differ. 2015;22(5):801–814. doi:10.1038/cdd.2014.175