
Abstract
Purpose
This study seeks to investigate the effect of evodiamine on psoriasis and psoriatic pruritus.
Methods
Imiquimod-induced psoriasiform dermatitis in mice was used as a model, and evodiamine was topically applied for seven days. The mice were observed daily for skin damage on the back, clinical score and their scratching behavior was recorded. Blood samples were collected on the final day of the experiment, and the serum levels of pruritus-associated inflammatory cytokines tumor necrosis factor (TNF) -α, interleukin (IL) −23, and IL-17A were measured using enzyme-linked immunosorbent assay. Histopathological changes were observed in Hematoxylin and Eosin-stained skin specimens. The expression levels of transient receptor potential vanilloid (TRPV) 1, TRPV3, TRPV4, and the pruritus-related mediators Substance P (SP), nerve growth factor (NGF), and calcitonin gene-related peptide (CGRP) in the skin lesions were analyzed using Western blot and qRT-PCR. The effect of evodiamine on the exploratory behavior, motor, and coordination abilities of mice was assessed using open field, suspension, and Rota-Rod experiments. Molecular docking was utilized to verify the binding of evodiamine to the residues of TRPV1, TRPV3, and TRPV4.
Results
Evodiamine reduced pruritus and inhibited inflammation by decreasing the levels of inflammatory mediators TNF-α, IL-23, and IL-17A in the serum of Imiquimod-induced mice and attenuated the mRNA and protein expression levels of SP, NGF, CGRP, TRPV1, TRPV3, and TRPV4 in the skin.
Conclusion
Evodiamine is an effective treatment for psoriasis and pruritus, due to its ability to inhibit immune inflammation and pruritic mediators.
Keywords:
Introduction
Psoriasis is a common, chronic skin disorder caused by a T cell-mediated immune response.Citation1 The prevalence of psoriasis demonstrates significant global variation. The prevalence of psoriasis in children is 0% in Taiwan China, and approximately 2.1% in Italy. In contrast, adult prevalence rates vary, ranging from a low of 0.4% in Asian countries to a peak of 8.5% in Norway.Citation2
It is characterized by red patches and silvery-white scales on the skin and often involves recurrent attacks and prolonged discomfort. Up to 80% of psoriasis patients also experience itching, an unpleasant sensation that can lead to further skin damage if scratched.Citation3 Studies have also demonstrated a strong correlation between emotional distress and itching intensity in severe cases, highlighting the substantial negative impact of itching on patients.Citation4 Despite the high prevalence of psoriatic pruritus, its pathogenesis remains unclear. Various theories attempt to elucidate the origin of psoriatic pruritus. One theory posits that the mechanism is related to neurogenic inflammation of the skin. Neuropeptides and cytokines such as substance P (SP), neurokinin A (NKA), nerve growth factor (NGF), opioids, and interleukin (IL) −2 have been implicated in psoriatic pruritus. Another hypothesis is the cutaneous overinnervation theory. The expression of NGF and the NGF receptor, TrkA, is increased in psoriatic lesions. This upregulation leads to hypersensitivity of peripheral sensory nerves to pruritic stimuli. Another well-known pathway mediating itch is the opioid pathway. Activation of κ receptors (OPRK) attenuates itch, whereas activation of OPRM enhances itch.Citation5 Owing to the complex pathogenesis and chronic nature of psoriasis, its treatment is challenging and prolonged. In the past, systemic medications such as methotrexate, cyclosporine, and acitretin have been the mainstay of treatment for moderate to severe psoriasis. However, the treatment of itching associated with psoriasis has not received adequate attention. Antihistamines have been employed in the treatment of psoriatic itch, yet they have yielded very poor results. Antihistamines have been used to treat psoriatic itch, but with very poor results. The recent approval of novel biologic therapies targeting tumor necrosis factor (TNF) -α, IL-23, and IL-17A has led to a major shift in the treatment of psoriasis. One of these therapies, a monoclonal antibody targeting IL-17, has shown over 70% improvement in psoriasis-induced pruritus. However, there is no clear evidence indicating a direct involvement of IL-17 in itch. Another effective strategy is to target more downstream pathways of the itch receptors. An example is the use of JAK1 inhibitors in cases where IL-31 receptor signaling causes itch in psoriasis patients. However, this approach may increase the potential for adverse side effects due to the involvement of many upstream receptors.Citation6
Fructus Evodiae, the dried and nearly mature fruit of the Rutaceae plant (), is a traditional Chinese medicinal herb documented in the “Shennong Materia Medica Classic”. It possesses a pungent and bitter taste, a warming nature, and slight toxicity, and is associated with the liver, spleen, stomach, and kidney meridians. Pharmacological studies have revealed that Fructus Evodiae possesses a wide range of effects, including anti-inflammatory, antibacterial, and antipruritic properties.Citation7 Evodiamine (), a major component of Fructus Evodiae, is a tryptophan-derived indole alkaloid with a molecular formula of C19H17N3O and a molecular weight of 303.36. Evodiamine is soluble in acetone, soluble in DMSO (up to 5 mg/mL) slightly soluble in ethanol, ether, chloroform, almost insoluble in water, petroleum ether, benzene. It is also hepatotoxic and cardiotoxic. This substance has shown various medicinal effects, including antitumor, anti-inflammatory, analgesic, antibacterial, endocrine regulatory, hypoglycemic, and smooth muscle regulatory properties.Citation8 However, the efficacy of evodiamine in reducing psoriatic itch has not yet been established. This study was designed to determine whether evodiamine can alleviate the inflammatory and itching in psoriasis-like mice and to explore its mechanism of action.
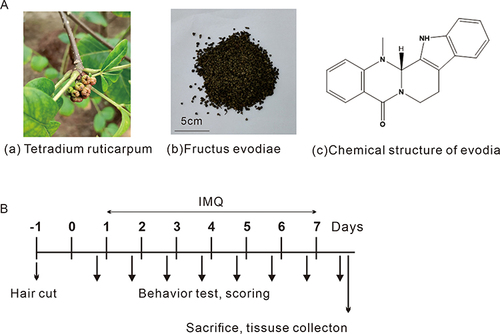
Figure 1 Materials and experimental schedule. (Aa) Tetradium ruticarpum. (Ab) Fructus Evodiae. (Ac) chemical structure of evodiamine. (B) Schematic diagram of the experimental procedure.
Materials and Methods
Mice
Forty male C57BL/6J Mice, aged 6–8 weeks and weighing 20–25 g, were obtained from Hunan Sleek Jingda Experimental Animal Co., Ltd. (License number SCXK (Xiang) 2019–0004). Mice were housed in a laboratory animal room at a temperature of 24 ±1°C, with a relative humidity of 40–70%, and a 12:12 h light-dark cycle. The animals were provided with a standard diet and water ad libitum. Animal experiments were conducted in strict accordance with the guidelines for the Care and Use of Laboratory Animals. All experiments received approval from the Institutional Animal Care and Use Committee of Guangxi University of Chinese Medicine (Ethics license DW20181220-153).
Preparing Evodiamine Solutions
The evodiamine, batch number MUST-21050405, was purchased from Chengdu Wande Biotechnology Co., Ltd., Sichuan, China. Solutions with concentrations of 0.75, 1.50, and 3.00 mg/mL were prepared by dissolving evodiamine in a mixture of 1% dimethyl sulfoxide (DMSO), 0.5% Tween-80, and 98.5% normal saline. The evodiamine solution was stored at 4°C until further use.
Experimental Group of Mice
Mice were randomly divided into five groups: control, model (Imiquimod, IMQ), and three evodiamine groups (evo-30 (IMQ + 30g/kg evodiamine), evo-15 (IMQ + 15g/kg evodiamine), and evo-7.5 (IMQ + 7.5g/kg evodiamine). Two days before treatment, the backs of the mice (4.0 cm × 2.0 cm) were shaved and hair removal cream was applied for further hair removal. In the model and evodiamine groups, mice received a daily topical dose of 62.5 mg of 5% IMQ cream (Mingxin Pharmaceutical Co., Ltd., Sichuan, China) to induce psoriasis-like skin inflammation on their shaved backs. The evodiamine groups were treated with IMQ for 4 hours daily and given different concentrations of evodiamine by gavage for 7 consecutive days. Mice in the model group were administered an equal volume of normal saline. An observer took photos and weighed the mice daily. Three evaluators rated the erythema, desquamation, and hypertrophy of the mice’s back skin on a scale of 0 to 4. At the experiment’s conclusion, all mice were euthanized, and their skin specimens were preserved for measurement and further analysis. Dermoscopic observations were made on day 7 of the experiment on the back of the mice centred on the central area of the skin lesions. The experimental plan is illustrated in .
Histopathological Analysis
On the seventh day, the back skin samples were fixed in 10% formalin. Following slicing and paraffin embedding, the 4 μm thick sections were stained with Hematoxylin and Eosin. The sections were examined using an OLYMPUS DP80 microscope and photographed at 10× magnification (n=3 for each group). The thickness of 10 epidermal sites on each image was randomly measured using ImageJ software.
Scratching Behaviour
During the experiment from day 0 to 7, the behavior of the mice was monitored twice daily for an hour each time, following a 30-minute adaptation period to the testing environment. The mice were placed in individual acrylic boxes containing an adequate amount of corn kernels. The number of scratches, defined as quick touches of the front or hind paw at the affected area ending with licking, biting the toes, or placing the hind paw on the ground, was counted from offline videos. All experiments were conducted under randomized, controlled, and observer-blind conditions.
Elisa
The levels of TNF-α, IL-17A, and IL-23 in mouse serum were quantified using commercial enzyme-linked immunosorbent assay (ELISA) kits (BioTek, Epoch), employing standard curves derived from various dilutions of purified recombinant TNF-α, IL-17A, TGF-1β, and IL-23. Absorbance was measured at 450 nm following the manufacturer’s protocol.
qRT-PCR
Total RNA was extracted from skin lesions using the TRIzol method (Ambion, USA). Subsequently, cDNA was synthesized using reverse transcriptase (Vazyme, China). Real-time quantitative PCR was performed using SYBR Green Master Mix (Vazyme, China) on an EDC-810 real-time fluorescence PCR system (Eastwin, China). The β-actin gene served as a reference for normalizing experimental data and calculating the relative expression levels of target genes. The ΔCt method was employed to analyze the relative quantitative levels of each sample gene, and the primers used are listed in .
Table 1 Sequence of Primers
Western Blot
Skin samples from the dorsal lesions of mice were homogenized in RIPA lysate (Beyotime, China), and the supernatant was utilized for total protein extraction. The total protein concentration was determined using a BCA protein concentration assay kit (Beyotime, Shanghai, China). Equal amounts of protein were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membranes (0.45 μm, Millipore, United States). The membrane was incubated with rabbit anti-transient receptor potential vanilloid (TRPV) 1, TRPV3, TRPV4, SP, NGF, and calcitonin gene-related peptide (CGRP) antibodies (Affinity), and mouse anti-β-actin antibody (Proteintech Group, Inc)., followed by HRP-conjugated goat anti-rabbit and goat anti-mouse secondary antibodies (Wuhan Boster Biological Technology, Ltd). Target proteins were visualized using ECL reagents, and protein expression levels were quantified using Image Pro Plus (IPP). β-Actin served as an internal reference protein.
Open-Field Experiments
Following 1 hour and 24 hours of pretreatment, the mice were positioned in the center of a rectangular box (62 cm × 62 cm × 30.5 cm), constructed of polymethyl methacrylate and divided into 16 equal squares (15.5 cm × 15.5 cm each). The mice were allowed to freely explore the area for 5 minutes, with the experiment being recorded by a high-definition camera. Adhering to the double-blind principle, the total number of squares crossed and the total number of front paws raised were recorded to assess the excitability, exploratory behavior, and hind limb motor skills of each experimental group. After each exploration session, the platform was cleansed with 75% ethanol to eliminate odors before introducing the next mouse.Citation9
Suspension Experiment
Mice were positioned between two sides of a horizontal metal wire (1000 mm × 2 mm), set 50 cm above the padding. Each mouse underwent three consecutive experimental rounds, with a 120-second interval between each. A score of 2 points was assigned if the mouse hung on the wire for more than 120 seconds or ran to the other end. A score of 1 point was recorded if the mouse remained on the wire for more than 60 seconds but less than 120 seconds (including 120 seconds). A score of 0 was given if the mouse fell from the wire within 60 seconds (including 60 seconds). The average score from the three trials was recorded. One hour prior to the suspension experiment, 0.2 mL of oral distilled water and high, medium, and low doses of evodiamine were administered to the respective groups for pre-treatment. Two suspension trials were performed on the mice at 1 hour and 24 hours post-pre-treatment, and the experimenter recorded data to investigate the effects of different evodiamine concentrations on motor skills.Citation10
Rota-Rod Experiment
To assess the impact of evodiamine on the mice’s motor coordination, the mice were placed on a rotarod apparatus. The apparatus was programmed to gradually increase its speed from 4 r/min to 40 r/min over a span of 5 minutes. Prior to the experiment, the mice underwent training three times daily for three days. Following 1 hour and 24 hours of pre-treatment, two separate rotarod trials were performed on the mice, with each mouse undergoing three consecutive test rounds and a 5-minute interval between each. The time until each mouse fell was recorded, and the average value was calculated. The data were analyzed to evaluate the motor coordination abilities of the mice in each experimental group.
Molecular Docking
The initial step involved obtaining the three-dimensional structural files of target proteins TRPV1 (ID:6LP3), TRPV3 (ID:6HA6), and TRPV4 (ID:4DX1) from the Protein Data Bank (PDB) database. Conversely, the structure of evodiamine was sourced from the PubChem database and converted to PDB format using PyMOL software. Subsequently, AutoDock Tools software was used to identify the protein receptors, add non-polar hydrogen atoms, and calculate the active site values. Molecular docking was then executed using Perl scripts, revealing the binding energy between the protein receptors and small molecule ligands. PyMOL software was also employed for visualizing the molecular docking structures.
Statistical Analysis
All experimental data were statistically analyzed using GraphPad Prism version 8.0.2 (GraphPad Software Corporation, La Jolla, CA, USA). Data are presented as mean ± standard error of the mean (SEM). Analysis of variance (ANOVA) was utilized to evaluate differences in means between groups, and the Wilcoxon rank-sum test was applied to samples with non-normal distributions. A p-value less than 0.05 was considered indicative of statistical significance.
Results
Evodiamine Alleviates IMQ-Induced Psoriasis-Like Skin Inflammation and Damage
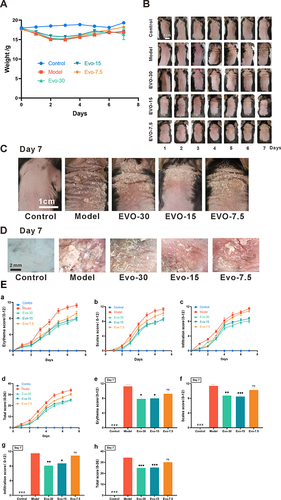
The Results of the experiment () demonstrated that the skin condition scores of the mice gradually stabilized by the 5th day. Compared to the control group, in the IMQ-induced psoriasis mouse model, there were signs of erythema, scales, and infiltration in the skin lesions, evidencing a marked difference in scores between the two groups by day 7 ( and ). Following IMQ treatment, evodiamine administration resulted in a reduction of the skin inflammation caused by IMQ, including erythema, scales, infiltration, and overall score (). Furthermore, mouse weights were monitored throughout the experiment, revealing no significant differences in the effects of IMQ and evodiamine on their weights (). The results indicate that evodiamine effectively alleviates IMQ-induced psoriasis-like skin inflammation without significantly affecting the weight of the mice.
Figure 2 Evodiamine improves skin damage in a mouse model of psoriasis. (A) Body weight monitoring of mice throughout the experimental period. (B) Seven-day observations of skin changes in mice treated with either IMQ alone or a combination of IMQ and EVO (30, 15, and 7.5 mg/kg). (C) Day 7: observation of mouse back condition (Scale bar = 1 cm). (D) Day 7: microscopic observation of skin from each mouse group (Scale bar = 2 cm). (E) Dermatitis in all rats was scored (0–4) by 3 randomized individuals and the 3 scores were summed to obtain a total score (0–36), (a-d) consecutive days 0–7: erythema (a), scaling (b), hypertrophy (c), total score (d). (e-h) Day 7 of the experiment: erythema (e), scaling (f), hypertrophy (g), total score (h). Data expressed as mean ± SEM (n=8 per group).###p<0.001 indicates control group vs. model group. *p<0.05, **p<0.01, and ***p<0.001 indicates evodiamine groups vs. model group.
Evodiamine Inhibits Epidermal Hyperplasia Induce by IMQ
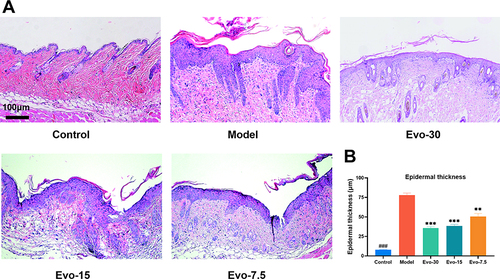
Histopathological analysis of mouse skin lesions using H&E staining () revealed that, compared to the control group, the model group exhibited visible alterations such as increased epidermal thickness, incomplete keratinization, and an abundance of neutrophils. Statistical analysis of the epidermal thickness showed a significant difference between the model and control groups (). Our results indicate that IMQ can effectively induce typical psoriasis-related histopathological changes in mice. Additionally, the epidermal thickness in the evodiamine group was lower than that in the model group, indicating that evodiamine can inhibit epidermal thickening in mice with psoriasis in a dose-dependent manner. Overall, the findings of this study demonstrate that evodiamine is capable of reducing epidermal thickening in psoriasis-affected mice.
Figure 3 Effect of evodiamine on IMQ-induced lesion skin in mice. (A) Representative H&E-stained skin Images. (B) Measurements of epidermal thickness on day 7 (Scale bar = 100 μm). Data were expressed as mean ± SEM (n=3). ###p<0.001 indicates control group vs. model group. **p<0.01, and ***p<0.001 indicates evodiamine groups vs. model group.
Evodiamine Inhibits IMQ-Induced Pruritus
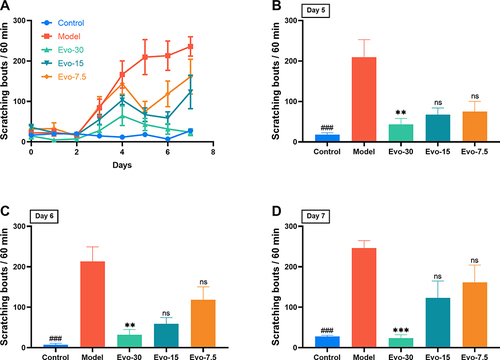
To investigate the effect of evodiamine on inhibiting IMQ-induced itching behavior, various concentrations of evodiamine were administered and their antipruritic effects were observed. As shown in , the number of scratching events in the control group remained below 50 for 7 days, forming a flat curve. The itching behavior in the model group increased significantly from the second day of the experiment, stabilized by the fifth day, and reached its peak on the last day. The itching behavior of the evo-7.5, evo-15, and evo-30 groups also increased on the second day, but the increase was less pronounced than that of the model group. The findings indicated that on days 5–7 of the experiment (), the scratching behavior of the model group was markedly higher than that of the control group. The scratching behavior of the evo-30 group showed a significant reduction compared to the model group. The scratching behavior of the evo-7.5 and evo-15 groups also exhibited a decrease in comparison to the model group, but this was not statistically significant. The results indicate that evodiamine inhibits IMQ-induced itching behavior.
Figure 4 Evodiamine inhibited scratching in IMQ-induced psoriasis in mice. (A) Measurement and comparison of spontaneous scratching among groups (n=8 per group). (B–D) Scratching bouts in mice on days 5, 6, and 7 post-treatment with controls or three doses of evodiamine. ###p<0.001 indicates control group vs. model group. nsp>0.05, **p<0.01, and ***p<0.001 indicates evodiamine groups vs. model group.
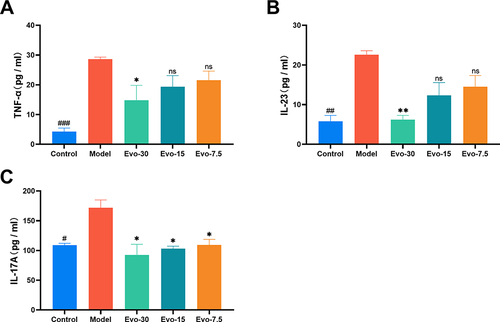
Evodiamine Decrease Serum Inflammatory Cytokines Level
We examined the levels of TNF-α, IL-23, and IL-17A in the serum of each group of mice. In the model group, the levels of the inflammatory cytokine TNF-α, along with IL-23 and IL-17A, were markedly elevated compared to the control group. However, the mice treated with evodiamine exhibited significantly reduced levels of TNF-α (), IL-23 (), and IL-17A () compared to the model group. This result suggests that evodiamine may alleviate IMQ-induced skin inflammation.
Figure 5 Effect of evodiamine on serum inflammatory cytokine levels in IMQ-induced psoriasis mice. (A-C) Serum Levels of TNF-α (A), IL-23 (B), and IL-17A (C) decreased following evodiamine administration in IMQ-Induced mice. ###p<0.001 indicates control group vs. model group. *p<0.05 and **p<0.01 indicates evodiamine groups vs. model group.
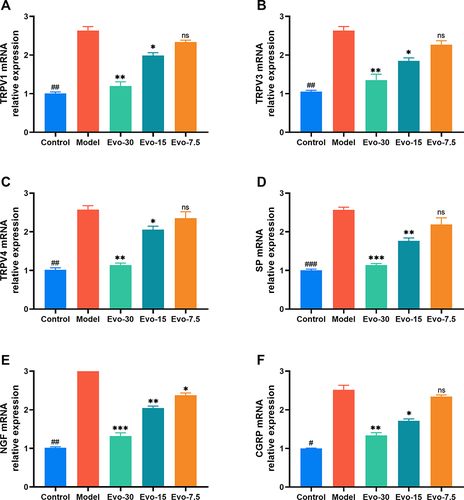
Evodiamine Reduces the mRNA Levels of TRP Channel and Pruritus Mediator
qRT-PCR analysis was used to measure the mRNA levels of TRPV1, TRPV3, TRPV4, SP, NGF, and CGRP in mouse back lesions. The results demonstrated that IMQ enhanced the mRNA expression of TRPV1 (), TRPV3 (), TRPV4 (), SP (), NGF (), and CGRP () in mouse skin tissue compared to the control group, while evodiamine treatment resulted in a significant decrease in the expression of these channels and mediators ().
Figure 6 Effect of evodiamine on pruritic channel and mediators mRNA levels in IMQ-induced psoriasis mice. (A–F) mRNA Levels of TRPV1 (A), TRPV3 (B), TRPV4 (C), SP (D), NGF (E), and CGRP (F) Determined using qRT-PCR. #p<0.05, ##p<0.01, and ###p<0.001 indicates control group vs. model group. nsp>0.05, *p<0.05, **p<0.01, and ***p<0.001 indicates evodiamine groups vs. model group.
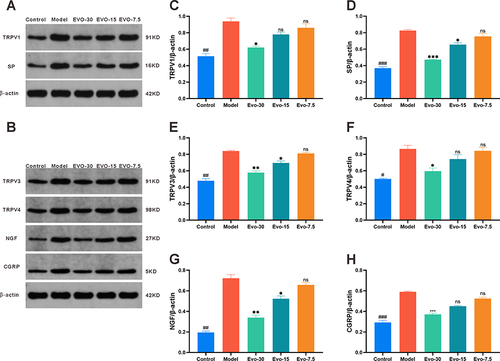
Evodiamine Reduces the Expression of TRP Channel and Pruritus Mediator
Western blotting analysis was employed to evaluate the expression levels of TRPV1, TRPV3, TRPV4, SP, NGF, and CGRP proteins in mouse skin lesion samples for investigating the effects of evodiamine on these proteins. The results of the Western blotting analysis bands are shown in ( and ). The results indicated that the expression levels of TRPV1 (), SP (), TRPV3 (), TRPV4 (), NGF (), and CGRP () in the model group were markedly elevated compared to those in the control group. In contrast, the expression levels of these proteins in the evodiamine group exhibited a significant reduction compared to those in the model group, suggesting that evodiamine may reduce the itching behavior of psoriasis mice by downregulating the expression of TRPV1, TRPV3, TRPV4, SP, NGF, and CGRP.
Figure 7 Effect of evodiamine on pruritic channel and peptides expression levels in IMQ-induced psoriasis mice. (A and B) Expression Levels of TRPV1, TRPV3, TRPV4, SP, NGF, and CGRP (G) Analyzed via Western blotting. (C–H) Expression levels of TRPV1 (C), SP (D), TRPV3 (E), TRPV4 (F), NGF (G), and CGRP (H) Assessed using Image J. #p<0.05, ##p<0.01, and ###p<0.001 indicates control group vs. model group. nsp>0.05, *p<0.05, **p<0.01, and ***p<0.001 indicates evodiamine groups vs. model group.
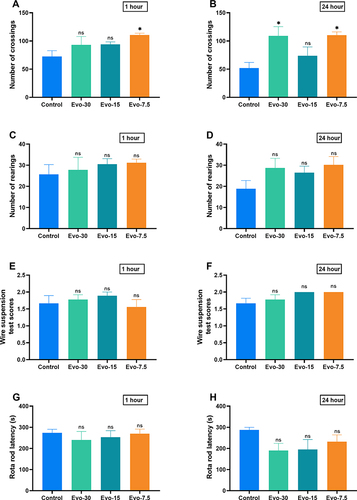
Evodiamine Has No Effect on the Exploration Ability, Motor Ability and Motor Coordination Ability of Mice
To verify the sedative and motor coordination effects of evodiamine, we performed suspension, open-field, and Rota-Rod experiments. The open-field experiment revealed no significant differences in the number of spanning grids ( and ) and standing positions ( and ) between the control group and the evodiamine-treated mice. Similarly, the suspension and Rota-Rod experiments demonstrated no significant differences in scores ( and ) and drop times ( and ) between the control group and the evodiamine-treated mice, respectively. This result indicates that evodiamine does not affect the motor, exploratory, and motor coordination abilities of mice.
Figure 8 Effect of evodiamine on locomotion, exploration, and the coordination of motor in IMQ-induced psoriasis mice. Frequency of mice crossing grids and rearing post-evodiamine treatment at 1 hour (A and C), and 24 Hours (B and D) in open field test; Scores from wire suspension experiment post-evodiamine treatment at 1 hour (E), and 24 hours (F); Duration before falling in Rota-Rod experiment post-evodiamine treatment at 1 hour (G), and 24 hours (H). Data were expressed as mean ± SEM (n=6). nsp>0.05 and *p<0.05 indicates evodiamine groups vs. the control group.
Molecular Docking
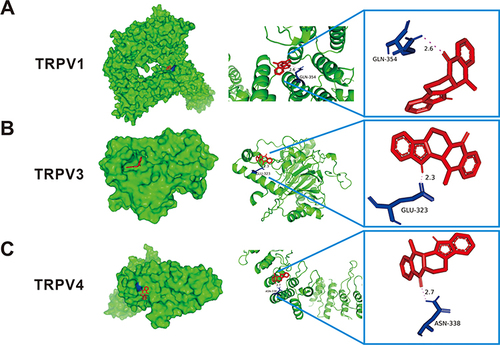
The molecular docking results () indicate that the binding energies of evodiamine with the key target proteins TRPV1, TRPV3, and TRPV4 are −8.9 kcal/mol, −7.9 kcal/mol, and −6.9 kcal/mol, respectively, all of which are lower than −5 kcal/mol, suggesting a secure ligand-receptor binding activity. depicts a protein docking pattern between evodiamine and the protein, showing that the residue GLN-354 of evodiamine and TRPV1 () can form hydrogen bonds with a bond length of 2.6; GLU-323, a residue of TRPV3 (), can form hydrogen bonds with a bond length of 2.3; and the residue ASN-338 of TRPV4 () can form hydrogen bonds with a bond length of 2.7. In conclusion, these findings suggest that evodiamine may exert its pharmacological effects through interaction with TRPV1, TRPV3, and TRPV4.
Table 2 The Docking of EVO and the Key Target Proteins Has Produced Results
Figure 9 Diagram of the molecular docking pattern. The three-dimensional structure of the target protein TRPV1 (A), TRPV3 (B), and TRPV4 (C) are depicted in green, the drug EVO’s structure in red, and the Interacting residues on the target protein with EVO in blue.
Discussion
Evodiamine, extracted as a monomer from the traditional Chinese medicine Fructus Evodiae, is reported to exhibit a wide range of pharmacological effects, including anti-inflammatory, anti-tumor, anti-obesity, anti-bacterial, and anti-aging properties. However, research on its effects on psoriasis and pruritus remains limited.Citation11 This study employed IMQ to induce a psoriasis itching model in mice, and determined that evodiamine exerted a significant inhibitory effect on inflammation and itching. The skin inflammation symptoms, such as erythema, scales, and thickening of the spinous cell layer, showed improvement, and itching behavior was likewise suppressed. These results demonstrate that evodiamine exerts an inhibitory effect on inflammation and itching in a mouse psoriasis model. However, not all types of psoriasis have abundant scales, and it is not yet known whether evodiamine can work in psoriasis models without abundant scales.
The pathophysiology of psoriasis is characterized by a complex network of immune cells and molecules, with the IL-23/IL-17 axis playing a pivotal role in the inflammatory process.Citation12 Immune cells produce TNF, which in turn activates myeloid dendritic cells, resulting in the secretion of IL-12 and IL-23, ultimately leading to the differentiation of Th17 and Th1 cells. Following treatment with evodiamine, there was a decrease in the expression of IL-17A and TNF in the serum of a psoriasis mouse model, indicating that evodiamine might influence the IL-23/IL-17 axis to mitigate skin inflammation in psoriasis mice.
Sensory nerves in the dermis of psoriasis patients are known to release SP and CGRP, which are recognized as potential itch mediators along with NGF.Citation10 These substances prompt mast cells to produce vasoactive amines and draw neutrophils and T cells to the area.Citation13 The expression of NGF in psoriasis patients experiencing itching is higher than in those without itching.Citation14 qRT-PCR analysis was used to detect the mRNA expression of genes related to itching, such as SP, CGRP, and NGF, in the skin, qRT-PCR analysis was used. Compared to the control group, the model group exhibited a significant increase in SP, CGRP, and NGF, aligning with the characteristics of patients with pruritus psoriasis.Citation14 Compared to the model group, there was a decrease in the expression of SP, CGRP, and NGF mRNA in the medium and high dose treatment groups, with a notable reduction in NGF mRNA expression in the low dose treatment group. Western blotting analysis also showed a marked decrease in the expression levels of SP, CGRP, and NGF in the medium and high dose groups compared to the model group. However, no significant difference was observed in the expression levels of SP, CGRP, and NGF between the low-dose treatment group and the model group.
Recent research has demonstrated that TRPV channels are closely associated with the development of psoriasis, contributing significantly to its physiological and pathological processes.Citation15 TRPV1 levels are observed to increase in sensitive skin, and it is detected in various skin cells and structures, including keratinocytes, dermal mast cells, dendritic cells, adipocytes, dermal blood vessels, hair follicles, and sweat glands.Citation7,Citation16 Sensitization of TRPV1, induced by mediators from inflamed skin, can lead to pain and itching, and may additionally amplify the peripheral inflammatory response via the expression of other inflammatory mediators.Citation17 Wilzopolski et alCitation18 demonstrated that both TRPV1 and TRPA1 channels play a role in the transmission of histamine-induced itching. IMQ is capable of activating sensory neurons expressing TRPV1, and in the IMQ-induced psoriasis mouse model, TRPV1 inhibitors have been shown to reduce the occurrence of itching and skin inflammation, as well as the expression of CGRP in DRG.Citation19,Citation20 Additionally, there are indications that the activation of TRPV1 on epidermal keratinocytes and the resulting influx of Ca2+ into cells may inhibit cell growth and induce apoptosis. Furthermore, in cases where the epidermal skin barrier is destroyed in mice, TRPV1 stimulation has been observed to delay the recovery of the barrier, while TRPV1 antagonists have been found to speed up the repair process. Finally, research indicatesCitation21 that capsaicin is able to regulate TRPV1 on HEK cells and inhibit protein synthesis, essential for the formation of skin barriers.
TRPV3, a member of the TRP family, is expressed in numerous tissues and organs, with its most notable expression observed in the skin’s epithelial cells. The activation of TRPV3 in skin keratinocytes is implicated in a variety of skin functions, including hair growth, wound healing, and temperature perception. Additionally, the TRPV3 signaling pathway plays a crucial role in maintaining the homeostasis of the epidermal barrier. Research has further indicated that TRPV3 can amplify itching signals. Studies demonstrate that mutations in the TRPV3 gene can lead to severe itching, hyperkeratosis, and increased IgE levels in both mice and humans. This indicates that TRPV3 is a significant contributor to skin itching.Citation22 Consequently, efforts have been made in research to develop TRPV3 antagonists as potential treatments for inflammatory skin diseases, itching, and pain. Additionally, carvacrol, identified as a natural agonist of TRPA1 and TRPV3, has been observed to induce intense itching in mice. Furthermore, the elimination of TRPV3 expression (knockout) has been demonstrated to reduce itching behavior in mice by up to 64%.Citation23
TRPV4, a non-selective cation channel, is present in dorsal root ganglion, trigeminal ganglion, and skin cells and plays a crucial role in the development and transmission of itching. In conditions such as dry skin, contact dermatitis, or rosacea, activation of TRPV4 in keratinocytes results in the release of 5-HT. This 5-HT binds to receptors in itching fibers and transmits a signal to the central nervous system, causing itching. Observations indicate that either overactivation or overexpression of the TRPV4 channel in mast cells can induce the release of histamine and activate receptors responsible for itching.Citation24 Yan et al’s study demonstrated that the expression level of TRPV4 correlates with the severity of psoriasis and its role in its onset and progression.Citation25 Furthermore, the itching symptom associated with psoriasis relies on TRPV4, upregulated in both the epidermis and dorsal root ganglion. Studies have found cimifugin to exhibit an antipruritic effect on psoriasis through its ability to block the activation of TRPV4 channels.
In this experiment, expression levels of TRPV1, TRPV3, and TRPV4 in the skin tissue of the mouse model group were observed to be markedly higher than those in the control group. Following evodiamine treatment, the expression levels of all three channels significantly decreased. Furthermore, after 7 days of treatment, mice in the high-dose group exhibited reduced itching behaviors and lower skin damage scores, a significant deviation from the model group. This indicates that TRPV1, TRPV3, and TRPV4 might play a role in the underlying mechanism of psoriasis itching and dermatitis. Molecular docking experiments demonstrated that evodiamine can form stable bindings with residues of TRPV1, TRPV3, and TRPV4. This implies that evodiamine could regulate the expression of these three channels and inhibit the influx of Ca2+, potentially reducing the secretion of associated neuropeptides and inflammatory molecules, thereby alleviating skin lesions and itching in psoriasis.
Conclusion
Evodiamine attenuated IMQ-induced inflammation and itching in a mouse model of moderate-to-severe psoriasis by modulating serum levels of TNF-α, IL-23, and IL-17A as well as the expression of SP, NGF, CGRP, TRPV1, TRPV3, and TRPV4 in the dermal lesions.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the Natural Science Foundation of China (NSFC) to Guanyi Wu (82060768); the Natural Science Foundation of Guangxi Zhuang Autonomous Region (2020GXNSFAA297244); Innovation Project of Guangxi Graduate Education (YCSW2023392); Guipai Traditional Chinese Medicine Top-Notch Talent Funding Project from Guangxi University of Chinese Medicine; the Second and Third Batch of the “Qihuang Project” High-Level Talent Team Cultivation Project of Guangxi University of Traditional Chinese Medicine (2021001, 202402).
References
- Nestle FO, Kaplan DH, Barker J. Psoriasis. N Engl J Med. 2009;361(5):496–509. doi:10.1056/NEJMra0804595
- Armstrong AW, Mehta MD, Schupp CW, et al. Psoriasis prevalence in adults in the United States. JAMA dermatol. 2021;157(8):940–946. doi:10.1001/jamadermatol.2021.2007
- Hawro T, Hawro M, Zalewska-Janowska A, et al. Pruritus and sleep disturbances in patients with psoriasis. Archiv Dermatological Res. 2020;312(2):103–111. doi:10.1007/s00403-019-01998-7
- Wu JJ, Feldman SR, Koo J, et al. Epidemiology of mental health comorbidity in psoriasis. J Dermatological Treat. 2018;29(5):487–495. doi:10.1080/09546634.2017.1395800
- Kahremany S, Hofmann L, Harari M, et al. Pruritus in psoriasis and atopic dermatitis: current treatments and new perspectives. Pharmacol Rep. 2021;73(2):443–453. doi:10.1007/s43440-020-00206-y
- Li H-J, Wu N-L, Pu C-M, et al. Chrysin alleviates imiquimod-induced psoriasis-like skin inflammation and reduces the release of CCL20 and antimicrobial peptides. Sci Rep. 2020;10(1):2932. doi:10.1038/s41598-020-60050-1
- Qin J, Liao CN, Chen WW, et al. New limonoids and quinolone alkaloids with cytotoxic and anti-platelet aggregation activities from Evodia rutaecarpa (Juss.) Benth. Fitoterapia. 2021;152:104875. doi:10.1016/j.fitote.2021.104875
- Sun Q, Xie L, Song J, Li X. Evodiamine: a review of its pharmacology, toxicity, pharmacokinetics and preparation researches. J Ethnopharmacol. 2020;262:113164. doi:10.1016/j.jep.2020.113164
- De Prá SDT, Ferro PR, Milioli AM, et al. Antinociceptive activity and mechanism of action of hydroalcoholic extract and dichloromethane fraction of Amphilophium crucigerum seeds in mice. J Ethnopharmacol. 2017;195:283–297. doi:10.1016/j.jep.2016.11.032
- Zhu X, Cheng M, Peng M, et al. Basal behavioral characterization of hsf1 deficient mice and its cellular and behavioral abnormalities underlying chronic unpredictable stressors. Behav. Brain Res. 2008;193(2):225–229. doi:10.1016/j.bbr.2008.05.024
- Gangwar RS, Gudjonsson JE, Ward NL. Mouse models of psoriasis: a comprehensive review. J Invest Dermatol. 2022;142(3):884–897. doi:10.1016/j.jid.2021.06.019
- Moynes DM, Vanner SJ, Lomax AE. Participation of interleukin 17A in neuroimmune interactions. Brain Behav Immun. 2014;41:1–9. doi:10.1016/j.bbi.2014.03.004
- Gouin O, Lebonvallet N, L’Herondelle K, et al. Self-maintenance of neurogenic inflammation contributes to a vicious cycle in skin. Experim Dermatol. 2015;24(10):723–726. doi:10.1111/exd.12798
- Nakamura M, Toyoda M, Morohashi M. Pruritogenic mediators in psoriasis vulgaris: comparative evaluation of itch-associated cutaneous factors. Br J Dermatol. 2003;149(4):718–730. doi:10.1046/j.1365-2133.2003.05586.x
- Oishi N, Iwata H, Kambe N, et al. Expression of precipitating factors of pruritus found in humans in an imiquimod-induced psoriasis mouse model. Heliyon. 2019;5(6):e01981. doi:10.1016/j.heliyon.2019.e01981
- Talagas M, Misery L. Role of keratinocytes in sensitive skin. Front Med. 2019;6:108. doi:10.3389/fmed.2019.00108
- Stander S, Moormann C, Schumacher M, et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Experim Dermatol. 2004;13(3):129–139. doi:10.1111/j.0906-6705.2004.0178.x
- Gouin O, L’Herondelle K, Lebonvallet N, et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: pro-inflammatory response induced by their activation and their sensitization. Protein and Cell. 2017;8(9):644–661. doi:10.1007/s13238-017-0395-5
- Wilzopolski J, Kietzmann M, Mishra SK, et al. TRPV1 and TRPA1 channels are both involved downstream of histamine-induced itch. Biomolecules. 2021;11(8):1166. doi:10.3390/biom11081166
- Kim S-J, Park GH, Kim D, et al. Analysis of cellular and behavioral responses to imiquimod reveals a unique itch pathway in transient receptor potential vanilloid 1 (TRPV1)-expressing neurons. Proc Natl Acad Sci. 2011;108(8):3371–3376. doi:10.1073/pnas.1019755108
- Han P, McDonald HA, Bianchi BR, et al. Capsaicin causes protein synthesis inhibition and microtubule disassembly through TRPV1 activities both on the plasma membrane and intracellular membranes. Biochem Pharmacol. 2007;73(10):1635–1645. doi:10.1016/j.bcp.2006.12.035
- Um JY, Kim HB, Kim JC, et al. TRPV3 and itch: the role of TRPV3 in chronic pruritus according to clinical and experimental evidence. Int J Mol Sci. 2022;23(23):14962. doi:10.3390/ijms232314962
- Cui T, Wang G, Wei N, et al. A pivotal role for the activation of TRPV3 channel in itch sensations induced by the natural skin sensitizer carvacrol. Acta Pharmacol Sin. 2018;39(3):331–335. doi:10.1038/aps.2017.152
- Rosenbaum T, Benítez-Angeles M, Sánchez-Hernández R, et al. TRPV4: a physio and pathophysiologically significant ion channel. Int J Mol Sci. 2020;21(11):3837. doi:10.3390/ijms21113837
- Yan J, Ye F, Ju Y, et al. Cimifugin relieves pruritus in psoriasis by inhibiting TRPV4. Cell Calcium. 2021;97:102429. doi:10.1016/j.ceca.2021.102429