
Abstract
Estrogens play central roles in sexual development, reproduction, and hepatocyte proliferation. The ovaries are one of the main organs for estradiol (E2) production. Ovariectomies (OVXs) were performed on the female mice, and hepatocyte proliferation was analyzed. The ovariectomized mice exhibited delayed hepatocyte proliferation after partial hepatectomy (PH) and also exhibited delayed and reduced E2 induction. Both E2 administration and PH induced the gene expression of estrogen receptor α (ERα). The transcripts of ERα were detected specifically in periportal hepatocytes after E2 administration and PH. Moreover, the E2 concentrations and hepatocyte proliferation rates were highest in the proestrus period of the estrous cycle. Taken together, these findings indicate that E2 accelerated ERα expression in periportal hepatocytes and hepatocyte proliferation in the female mice.
Introduction
Estrogens have a number of functions in development, growth, and sex differentiation, and they play important roles in female reproduction and in nonreproductive tract tissues, including the gonads, brain, bone, adipose tissue, and cardiovascular system.Citation1,Citation2 Estrogens contain mainly three compounds; Estron (E1), Estradiol (E2), and Estriol (E3). E2 is most active hormone among these Estrogens. The target factors of estrogens are thought to consist of three proteins, the estrogen receptor α (ERα), estrogen receptor β (ERβ), and the G protein-coupled receptor 30 (GPR30).Citation3–Citation6
Several studies have implicated the liver as an estrogenic organ, and an ER activityCitation7 (later called genomic reaction) has been detected in this organ before the first ER (later named ERα) was cloned from MCF7 (Michigan Cancer Foundation 7), human mammary cell line.Citation7–Citation13
The liver plays a pivotal role in mammalian homeostasis and has the ability to complement its original mass in response to several types of stress, including partial hepatectomy (PH).Citation14,Citation15 After PH, hepatocytes proliferate intensely for a few days, and the liver regenerates in 2 weeks.Citation14,Citation15
Estrogens induce hepatocyte proliferation in vitro in neonate and in vivo after PH.Citation8,Citation11,Citation12 These multiple effects could be mediated by ERα rather than ERβ and GPR30 because ERα has been shown to be expressed in the mouse liver.Citation16–Citation18 Moreover, estradiol (E2) signaling in the liver is a genomic reaction (see above), which indicates that ERα and/or ERβ are targets for E2 signaling in the liver.Citation17,Citation18 The expression of ERβ has not been detected in the liver, but ERα transcripts were detected.Citation19 Taken together, the target factor of E2 in the liver may be ERα.Citation20
To elucidate estrogen signaling in the liver of the female mice, ovariectomies (OVXs) were first performed to decrease E2 production, because the ovary is the tissue that produces the most E2.Citation21,Citation22 Next, the female mice were treated with E2. We showed that circulating E2 is significantly increased after PH in both non-OVX and OVX female mice and demonstrated that ERα, the expression of which is enhanced by E2 administration, plays a crucial role in E2-induced hepatocyte proliferation and liver regeneration.
Materials and methods
Materials
β-Estradiol (052-04041; Wako Pure Chemical Industries, Ltd, Osaka, Japan), 4-hydroxytamoxifen (H7904; Sigma-Aldrich, St Louis, MO, USA), and ICI182780 (ICI, I4409; Sigma-Aldrich) were purchased. The E2 enzyme immunoassay kit (No 582251) was purchased from Funakoshi (Tokyo, Japan). EDTA (15111-45), NaCl (31320-34), and Tris (35406-91) were purchased from Nacalai Tesque, Inc. (Kyoto, Japan). ERα-specific agonist 4,4′, 4′-(4-propyl-[1H]-pyrazole-1,3,5-triyl)trisphenol (PPT, CAS No: 263717-53-9) was purchased from Sigma-Aldrich (H6036). ERβ-specific agonist 2,3-bis(4-hydroxyphenyl)propionitrile (DPN) was purchased from Cayman Chemical, Ann Arbor, MI, USA/Funakoshi 10008842. The Hep G2 cell line from human hepatocellular carcinoma cell line was purchased from the American Type Culture Collection (HB-8065; ATCC, Manassas, VA, USA).
Hepatocyte proliferation in the female mice
The hepatocyte proliferation rates of the female mice were analyzed with bromodeoxyuridine (BrdU, M0744; Dako Denmark A/S, Glostrup, Denmark) immunohistochemistry (IHC, SK-4105; Vector Marketing Corporation, Gibbsboro, NJ, USA). The females were injected intraperitoneally with 50 mg/kg of BrdU 2 hours before dissection, and the livers were removed, rinsed, and embedded in the tissue-Tek OCT compound (Sakura Finetek Japan Co, Ltd, Tokyo, Japan). Ten-micrometer cryosections were fixed with 4% paraformaldehyde, incubated with an antibody against BrdU (No 11170376001, Hoffman-La Roche Ltd, Basel, Switzerland) that was diluted 50-fold in 0.1% bovine serum albumin/phosphate-buffered saline (BSA/PBS), revealed by CY3- conjugated donkey anti-rabbit IgG antibody, and mounted with Vectashield medium (Vector Laboratories Ltd, Burlingame, CA, USA). The numbers of BrdU-positive hepatocyte nuclei in at least five low-magnification microscopic fields of each sample (~2000 hepatocytes) were counted.Citation23–Citation25
In situ hybridization
The RNA probes were prepared. The ERα and ERβ cDNAs were cloned into pSG5 vector,Citation26 and the 35S-labeled antisense probes were synthesized by T7 in vitro transcription and translation system (Promega Corporation, Fitchburg, WI, USA). The specimens were prehybridized for 2 hours at 50°C in prehybridization buffer (50% formamide, 0.3 M NaCl, 10 mM Tris–HCl at pH 6.8, 10 mM NaPO4 at pH 6.8, 5 mM Ethylene-diaminetetraacetic aicd (EDTA), 1× Denhart’s, 10 mM DTT, 500 mg/mL yeast RNA, 100 mg/mL salmon sperm DNA, and 500 nmol/mL nonlabeled α-thio-UTP [DuPont, Wilmington, DE, USA]). After the RNase A treatment, the slides were washed for 1 hour in the washing buffer (50% formamide, 0.3 M NaCl, 10 mM Tris–HCl at pH 6.8, 10 mM NaPO4 at pH 6.8, 5 mM EDTA, 1× Denhart’s, and 10 mM DTT). The slides were subsequently washed in 2× Saline-Sodium Citrate buffer (SSC) for 15 minutes at room temperature, in 0.1× SSC for 15 minutes at 50°C, and then in 0.1× SSC for 30 minutes at room temperature. After dehydration of the sections, they were coated with Kodak NTB-2 emulsion, dried, and stored at 4°C. The exposure time ranged from 12 days to 15 days. Kodak D19 developer was used for 2 minutes at room temperature. The sections were then stained in toluidine blue, dehydrated in ethanol, and mounted under coverslips in Eukitt mounting medium.Citation26–Citation29
Reverse transcription polymerase chain reaction
Total liver RNA was extracted by the guanidium thiocyanate-phenol-chloroform method. cDNA was synthesized for 20 minutes at 50°C from 1 µg of RNA with Moloney murine leukemia virus reverse transcriptase. The transcribed cDNA was amplified by 30 cycles of PCR for ERα (5′-CGG CTG CCA CTT ACC TGG GAG CTC TCA GAT-3′ and 5′-GGG GAG CCT GGG AGC TCT CAG AT-3′), ERβ (5′-TCT CTG AGA GCA TCATGT CC-3′ and 5′-CAG CCT GGC CGT CAC TGT GA-3′), and hypoxanthine phosphoribosyltransferase (5′-GTA ATG ATC AGT CAA CGG GGG AC-3′ and 5′-CCA GCA AGC TTG CAA CCT TAA CCA-3′).Citation24,Citation30,Citation31
Surgeries (PHs, orchiectomies, and OVXs)
Liver resection of the left and median lobes was performed following midventral laparotomy between 8 am and 11 am under isoflurane anesthesia as previously described.Citation23,Citation32 The bilateral OVX procedure was performed as follows: the mice were anesthetized with peritoneal injections of pentobarbital, one central lateral incision was made in the skin, and two lateral incisions were made in the muscle layer, and the ovaries were extracted through the incision and excised after ligation.
Animal study compliance
All experiments were performed in accordance with the ethical guidelines for animal care of the National Center for Geriatrics and Gerontology (NCGG). The experimental protocols were approved by the Animal Care Committee of the NCGG. All the surgeries were performed under sodium pentobarbital anesthesia, and all efforts were made to minimize the suffering of the animals. E2 injections were decreased from five repetitions to a single injection to decrease the suffering of the sacrificed and operated mice.
Hep G2 cell culture
Hep G2 cells from a human liver carcinoma were maintained in α-minimal essential medium (11900-073; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal calf serum (FCS, CC3008-504, Cell Culture Technology, Tokyo, Japan). The cells were plated at a density of 1.5–3×106 cells/60-mm dish, and after 48 hours, the culture medium was exchanged for α-minimal essential medium without phenol red supplemented with 0.5% charcoal-treated FCS. After 24 hours, the cells were treated with β-estradiol or an equal volume (0.01 [v/v]%) of vehicle (EtOH, 14712-34; Nacalai Tesque, Inc.).Citation20,Citation25 Phenol red is a known phytoestrogen. The FCS contains higher endogenous estrogen concentrations, and the charcoal treatment reduces this concentration in the FCS.
Statistical analysis
The values are reported as the mean ± SEM. Statistical significance (ANOVA and Student’s t-test) is indicated as follows: *P<0.05; **P<0.005; ***P<0.0001. Nonsignificant differences (P>0.05) are indicated as NS.Citation23–Citation25,Citation28
Results
PH induces E2 concentration in orchiectomized and OVX mice
Previous studies indicate that PH induces E2 concentration in human beings, rats, and male mice ().Citation9,Citation11,Citation12,Citation20 E2 is converted from testosterone by aromatase, and the highest expression of aromatase is observed in the ovary and the testis; however, other tissues (eg, the gonads, placenta, adipose tissue, etc) also weakly produce aromatase.Citation21,Citation22 These observations suggested that no PH-induced E2 production was observed in orchiectomized (ORC) mice. So, the male mice were operated with ORC, and then, these ORC mice were operated with PH. After PH, plasmas were collected, and circulating E2 concentrations were measured (). Plasma E2 was strongly elevated after 6–48 hours in the control mice after PH (open squares) and was also induced after 24–48 hours in the ORC mice (filled triangles with dotted lines), which indicates that the E2 induction following PH was delayed and reduced in the ORC mice. PH-induced E2 concentrations were mainly from testes (of control mice) and majorly from nontestes organs (of ORC mice, eg, gonads, adipose tissue; ).
Figure 1 Delayed elevations of circulating E2 and ERα expression after PH were observed in the ORC and OVX mice.
Abbreviations: E2, estradiol; ERα, estrogen receptor α; PH, partial hepatectomy; ORC, orchiectomy; OVX, ovariectomy; SEM, standard error of the mean.
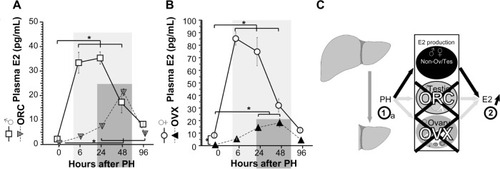
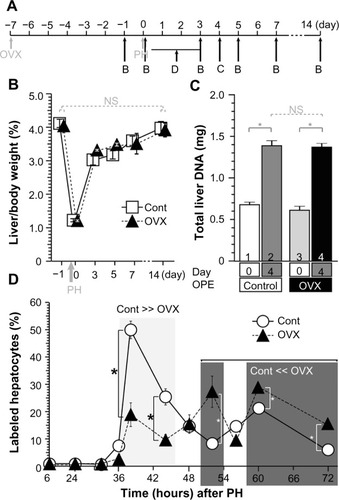
Female mice have endogenous higher E2 concentration than male mice; so, OVXs were performed () to analyze the effects of E2 concentration after PH. E2 concentrations were significantly decreased in the OVX mice (). After PH, E2 concentrations were induced in both OVX and control females. The E2 peaks in control females were ~80 pg/mL after 6–48 hours, and those of OVX mice were ~30 pg/mL after 24–48 hours, indicating delayed and small E2 induction by OVX. Taken together, PH induced E2 concentration mainly from testes and ovaries, and partly and delayed from other organs (unknown) in . Taken together, this is the first observation where PH triggered E2 concentration in both ORC and OVX animals.
E2 administration and the estrous cycle induced E2 concentration and hepatocyte proliferation
PH induces E2 concentrationCitation9,Citation11,Citation12,Citation20 () as well as hepatocyte proliferation ().Citation14,Citation15 E2 was administered to the OVX mice, and hepatocyte proliferation was analyzed (). E2 administration induced hepatocyte proliferation in the OVX mice in a dose-dependent manner ().Citation32 The target factors of E2 are ERα, ERβ, and GPR30,Citation3–Citation6 and the ICI182780 (ICI) is one of the selective ER modulators (SERMs). ICI is an antagonist for ERα and ERβ, but an agonist for GPR30.Citation5,Citation6,Citation33,Citation34 ICI reduced hepatocyte proliferation (), which indicates that the target(s) of E2 action is related to hepatocyte proliferation that was not GPR30 but could have been ERα and/or ERβ. Moreover, no ERβ expression was observed in the male livers.Citation19,Citation20
Figure 2 The estrous cycle regulated E2 concentration and hepatocyte proliferation.
Abbreviations: E2, estradiol; BrdU, bromodeoxyuridine; SEM, standard error of the mean.
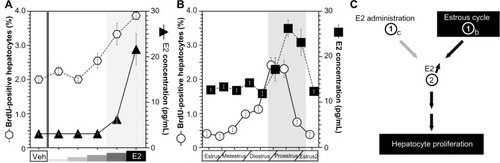
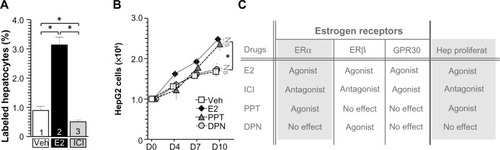
Figure 3 SERM administration regulated hepatocyte proliferation.
Abbreviations: SERM, selective estrogen receptor modulator; E2, estradiol; ICI, ICI182780; BrdU, bromodeoxyuridine; GPR, G protein-coupled receptor; ERα, estrogen receptor α; ERβ, estrogen receptor β; SEM, standard error of the mean; DPN, 2,3-bis(4-hydroxyphenyl)propionitrile; PPT, 4,4′,4′-(4-propyl-[1H]-pyrazole-1,3,5-triyl)trisphenol.
E2 is primarily produced by the ovaries in female mice.Citation21,Citation22 Generally, E2 production is dependent on the estrous cycle of nonpregnant mice, and E2 production is elevated during the proestrus period and reduced in the estrous cycle.Citation35,Citation36 The estrous cycles, E2 concentrations, and hepatocyte proliferations of nonpregnant female mice were analyzed (). The peak E2 concentrations and hepatocyte proliferations occurred during the proestrus phase/period (). E2 was administered to the OVX mice, and E2 concentrations and hepatocyte proliferations were analyzed (). Both the E2 concentrations and hepatocyte proliferations were similarly elevated in an E2-dependent manner (). Taken together, both E2 administration and the estrous cycle induced E2 concentration and induced E2 concentration lead hepatocyte proliferation ().
E2 and SERM administration regulated hepatocyte proliferation
To access the target factor for hepatocyte proliferation, several SERMs were administrated to hepatocytes (). At first, E2 (filled bar in ) and ICI (ERα and β antagonist, GPR30 agonist, gray bar in ) were administrated to B6 WT mice (). E2-stimulated hepatocyte proliferation and ICI-inhibited hepatocyte proliferation indicated that hepatocyte proliferation was due to ERα and/or ERβ, not due to GPR30 (). Other SERM, such as PPT (ERα-specific agonist, triangles, CAS No: 263717-53-9)Citation37 induced HepG2 cell proliferation significantly, but no induction was observed with ERβ-specific agonist DPN (circles, CAS No: 1428-67-7), indicating that ERα is the E2 target factor for E2-induced hepatocyte proliferation (). Using ERα and ERβ knock out (KO) (and their control) mice,Citation31 we confirmed that E2-induced hepatocyte proliferation was via ERα, not ERβ (data not shown and manuscript in preparation). Moreover, hepatocyte-specific ERα KO miceCitation23,Citation27 were established and analyzed, resulting in similar results (data not shown and manuscript in preparation). Taken together, E2-induced hepatocyte proliferation might be via hepatocyte-ERα with ligand and receptor analyses.
E2 administration and PH induced ERα expression in periportal hepatocytes
The mRNA expressions of ERα and ERβ in the liver were analyzed using reverse transcription polymerase chain reaction (RT-PCR) () and in situ hybridization (). First, ERβ mRNA was detected in the positive control of ovary, and not amplified from liver samples ().Citation19 PH and E2 administration induced the transcripts of ERα (). Note that both E2 injection and PH induced ERα expression synergetically. Delayed induced expression of ERα was observed in OVX mice at 24 hours after PH, similar to delayed E2 induction in OVX mice (). These observations suggested that PH induced delayed E2 concentration and ERα expression.
Figure 4 PH induced ERα expression in the hepatocyte of periportal area.
Abbreviations: PH, partial hepatectomy; ERα, estrogen receptor α; RT-PCR, reverse transcription polymerase chain reaction; ISH, in situ hybridization; OVX, ovariectomy; E2, estradiol; HPRT, hypoxanthine-guanine phosphoribosyltransferase.
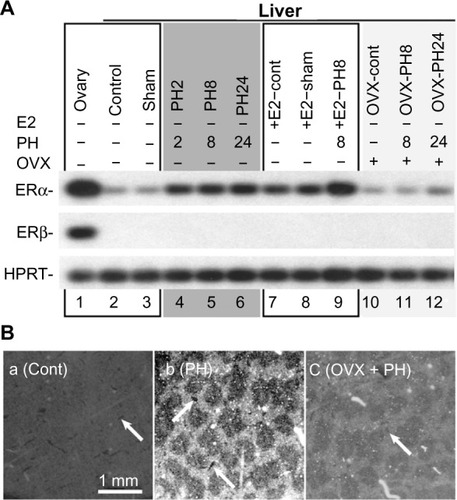
Moreover, E2 and PH induced ERα expression mainly in the hepatocytes located in the periportal area (), and these hepatocytes are known to actively proliferate after PH.Citation38
Delayed hepatocyte proliferation after PH in OVX mice
No significant differences in liver weight were observed between the OVX and control mice before or after PH (). Hepatocytes are the main parenchymal cells of the liver, and DNA from the recovered livers was extracted and analyzed. The liver DNA quantities were restored by 4 days after PH in the OVX and control mice, and no significant differences were observed between the OVX and control mice (); these findings indicate that OVX had no significant effect on the liver regeneration. Moreover, hepatocyte proliferation following PH was analyzed based on BrdU incorporation. The rate of hepatocyte proliferation in the control mice was significantly elevated 36–44 hours after PH, but that of the OVX mice was significantly higher at 52 hours, and 60–72 hours after PH (). These data indicate that the OVX mice displayed slightly delayed hepatocyte proliferation () but no significant change in liver weight/DNA amount recovery ().
Figure 5 OVX did not affect liver weight recovery after PH, and the hepatocyte proliferations were delayed.
Abbreviations: OVX, ovariectomy; PH, partial hepatectomy; BrdU-IHC, bromodeoxyuridine immunohistochemistry; SEM, standard error of the mean; NS, nonsignificant; OPE, operation.
Discussion
PH induced increases in E2 concentrations in ORC and OVX living organisms
This is the first study that PH induced E2 production in ORC and OVX animals. In the papers in the 1970s and 1980s, and in our latest studies, no observation of E2 induction in ORC/OVX rodents or human beings was reported.
E2 administration and PH induced ERα expression in the livers of the female mice
The first ERα was cloned by Professor Pierre Chambon in 1986. The first Francavilla’s study was published in 1984; they did not know the ERα itself. Recently ERα expression is induced in male mice, indicating that this is the first study to report ERα expression in females.
Hepatocyte proliferation was also observed in the proestrus period of the estrous cycle
There are no studies about hepatocyte proliferation in the estrous cycle, and this is the first case of hepatocyte proliferation that was observed in sex cycles of females.
Conclusion
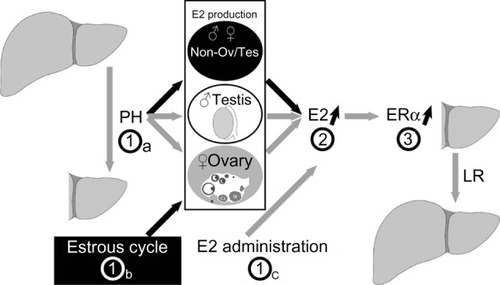
The possible mechanisms of PH- and E2-induced hepatocyte proliferation were identified (). PH (step 1a), the estrous cycle (step 1b), and E2 injection (step 1c) stimulated increases in E2 concentrations (step 2), ERα expression in the periportal hepatocytes (step 3), and hepatocyte proliferation.
Figure 6 Model of hepatocyte proliferation in female mice.
Abbreviations: PH, partial hepatectomy; E2, estradiol; ERα, estrogen receptor α; LR, liver regeneration.
Future perspective
Genetical study
Our previous and latest studies showed that PH induced ERα expression in rodents of both sexes. Using genetically modified mice disrupting ER genes, the hepatic ER function in liver regeneration was contributed (manuscript in preparation).
During pregnancy
We demonstrated that induced E2 production in the estrus period of the estrous cycle triggered hepatocyte proliferation. In general, when are the females exposed highest E2 concentration? It is during pregnancy. The livers during pregnancy were analyzed, and hepatocyte proliferation was observed as expected (manuscript in preparation).
Acknowledgments
We are grateful to our department members in the NCGG for helpful discussions. This work was supported by a grant-in-aid from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) and a grant from the Japan Science and Technology Agency (JST) to TI. The funders had no role in study design, data collection and analysis, the decision to publish, or manuscript preparation.
Disclosure
The authors report no conflicts of interest in this work.
References
- CioccaDRRoigLMEstrogen receptors in human nontarget tissues: biological and clinical implicationsEndocr Rev199516135627758432
- CouseJFKorachKSEstrogen receptor null mice: what have we learned and where will they lead us?Endocr Rev199920335841710368776
- AntalMCPetit-DemoulièreBMezianeHChambonPKrustAEstrogen dependent activation function of ERβ is essential for the sexual behavior of mouse femalesProc Natl Acad Sci U S A201210948198221982723150547
- AdlanmeriniMSolinhacRAbotAMutation of the palmitoylation site of estrogen receptor α in vivo reveals tissue-specific roles for membrane versus nuclear actionsProc Natl Acad Sci U S A20141112E283E29024371309
- ChimentoASirianniRCasaburiIPezziVGPER signaling in spermatogenesis and testicular tumorsFront Endocrinol (Lausanne)2014305
- VaucherLFunaroMGMehtaAActivation of GPER-1 estradiol receptor downregulates production of testosterone in isolated rat Leydig cells and adult human testisPLoS One20149e9242524736568
- JensenEVJordanVCThe estrogen receptor: a model for molecular medicineClin Cancer Res2003961980198912796359
- FisherBGunduzNSafferEAZhengSRelation of estrogen and its receptor to rat liver growth and regenerationCancer Res1984446241024156722782
- FrancavillaAEagonPKDiLeoASex hormone-related functions in regenerating male rat liverGastroenterology1986915126312703758617
- KahnDGavalerJSMakowkaLDoes hyperprolactinemia affect hepatic regeneration independent of sex steroids?J Lab Clin Med198811256446513183497
- FrancavillaAPolimenoLDiLeoAThe effect of estrogen and tamoxifen on hepatocyte proliferation in vivo and in vitroHepatology1984946146202784403
- FrancavillaAGavalerJSMakowkaLEstradiol and testosterone levels in patients undergoing partial hepatectomy. A possible signal for hepatic regeneration?Dig Dis Sci19893468188222721316
- KahnDEagonPKPorterLEEffect of tamoxifen on hepatic regeneration in male ratsDig Dis Sci198934127322910679
- SteerCJLiver regenerationFASEB J1995914139614007589980
- MichalpoulosGKDeFrancesMCLiver regenerationScience1997276530960669082986
- KuiperGGCarlssonBGrandienKComparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and betaEndocrinology199713838639709048584
- CianaPDi LuccioGBelcreditoSEngineering of a mouse for the in vivo profiling of estrogen receptor activityMol Endocrinol20011571104111311435611
- CianaPRaviscioniMMussiPIn vivo imaging of transcriptionally active estrogen receptorsNat Med200391828612483206
- VaccaMD’AmoreSGrazianoGClustering nuclear receptors in liver regeneration identifies candidate modulators of hepatocyte proliferation and hepatocarcinomaPLoS One20149e10444925116592
- UebiTUmedaMImaiTEstrogen induces estrogen receptor alpha expression and hepatocyte proliferation in the livers of the male miceGenes Cells201520321722325495062
- CarreauSLambardSDelalandeCDenis-GaleraudIBilinskaBBourguibaSAromatase expression and role of estrogens in male gonad: a reviewReprod Biol Endocrinol200313512747806
- BulunSELinZImirGRegulation of aromatase expression in estrogen-responsive breast and uterine disease: from bench to treatmentPharmacol Rev200557335938316109840
- ImaiTJiangMKastnerPChambonPMetzgerDSelective ablation of retinoid X receptor alpha in hepatocytes impairs their lifespan and regenerative capacityProc Natl Acad Sci U S A20019884581458611287642
- ImaiTTakakuwaRMarchandSPeroxisome proliferators-activated receptor γ is required in mature white and brown adipocytes for their survival in the mouseProc Natl Acad Sci U S A2004101134543454715070754
- UmedaMUebiTMaekawaNHandaHImaiTPGJIFs, new mitochondrial PGJ2 target proteins, regulate cell proliferationJ Biosci Med2013131115
- DolléPDubouleDTwo gene members of the murine HOX-5 complex show regional and cell-type specific expression in developing limbs and gonadsEMBO J198985150715152569970
- ImaiTChambonPMetzgerDInducible site-specific somatic mutagenesis in mouse hepatocytesGenesis200026214714810686613
- ImaiTJiangMChambonPMetzgerDImpaired adipogenesis and lipolysis in the mouse upon Cre-ERT2-mediated selective ablation of RXR alpha in adipocytesProc Natl Acad Sci U S A200198122422811134524
- NiederreitherKFraulobVGarnierJMChambonPDolléPDifferential expression of retinoic acid-synthesizing (RALDH) enzymes during fetal development and organ differentiation in the mouseMech Dev20021101–216517111744377
- ImaiTMatsudaKShimojimaTERC-55, a binding protein for the papiloma virus E6 oncoprotein, specifically interacts with vitamin D receptor among nuclear receptorsBiochem Biophys Res Commun199723337657699168930
- DupontSKrustAGansmullerADierichAChambonPMarkMEffect of single and compound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductive phenotypesDevelopment2000127194277429110976058
- HigginsGMAndersonRMExperimental pathology of the liver I. Restoration of the liver of the white rat following partial surgical removalArch Pathol193112186202
- DauvoisSWhiteRParkerMGThe antiestrogen ICI 182780 disrupts estrogen receptor nucleocytoplasmic shuttlingJ Cell Sci19931064137713888126115
- Movérare-SkrticSBörjessonAEFarmanHHThe estrogen receptor antagonist ICI 182,780 can act both as an agonist and an inverse agonist when estrogen receptor α AF-2 is modifiedProc Natl Acad Sci U S A201411131180118524395795
- BlendingerKPhysiology and pathology of the estrous cycle of the bitchItalian Companion Animal Veterinary AssociationProceedings of the SCIVAC CongressRimini, Italy2007:73–77
- SantmyireBRVenkatVBeinderEBaylisCImpact of the estrus cycle and reduction in estrogen levels with aromatase inhibition, on renal function and nitric oxide activity in female ratsSteroids201075121011101520619284
- KitamuraNArayaRKudohMBeneficial effects of estrogen in a mouse model of cerebrovascular insufficiencyPLoS One200944e515919357782
- FerriDMoroLMastrodonatoMUltrastructural zonal heterogeneity of hepatocytes and mitochondria within the hepatic acinus during liver regeneration after partial hepatectomyBiol Cell200597427728815762849