
Abstract
Background
Disc degeneration (DD) is one of the common diseases worldwide, which deeply influences normal life and leads to excruciating pain. However, an effective treatment for DD is still not identified.
Method
The present study systemically examined the effect of melatonin on annulus fibrosus (AF) cells of patients with DD.
Results
Melatonin had the effect of promoting proliferation, inducing autophagy, and suppressing apoptosis on AF cells of patients with DD. Moreover, melatonin contributed to the translation and transcription of autophagy-related protein ATG7 and inhibited the function of miR-106a-5p in AF cells. In addition, the results suggested that miR-106a-5p mediated the expression of ATG7 by directly binding to its 3′UTR in AF cells.
Conclusion
This research not only gained a deep insight of melatonin mode of action, but also indicated its potential target signaling pathway in AF cells.
Introduction
Disc degeneration (DD) is a complicated chronic process, which leads to the dysfunction of intervertebral discs (IVD).Citation1 Nowadays, degenerative disc disease is emerging as a common disorder worldwide. However, the major treatment for this disease is still relying on surgical intervention, which increases the risk for adjacent DD.Citation2 Therefore, an effective agent in the treatment for DD is urgently needed.
IVD are not only essential for the stability and flexibility of the spine, but also critical for resisting tension and bearing weight.Citation3 Both the nucleus pulposus (NP) and annulus fibrosus (AF) regions are essential for maintaining the function of IVD.Citation4
A previous report has indicated that the degeneration of the AF often leads to the dysfunction of IVD, which subsequently develops into disc herniation.Citation5 Therefore, the stability of AF is critical for maintaining the health of IVD.
It has been confirmed that autophagy regulates the cell death pathway, which plays a key role in the degradation of cellular constituents.Citation6 Moreover, a previous report has demonstrated that autophagy inhibits the apoptosis of AF cells and prevents the degeneration of IVD.Citation7 Therefore, the autophagy process is essential for maintaining the function of AF cells. However, the precise mechanism still needs to be further explored.
Autophagy is highly mediated by autophagy-related genes (ATGs), such as ATG5, ATG7, and ATG12. Growing evidence has indicated that MicroRNAs (miRs) play a key role in the autophagy pathway. miRs are a group of small noncoding RNAs, which have served as a valuable post-transcriptional mediator by binding to the 3′ untranslated regions (UTRs) of the targeting mRNAs.Citation8,Citation9 miR-375 and miR-20a-5p have been identified to inhibit autophagy and reduce cell viability via regulating ATG7.Citation10,Citation11 Moreover, miR-200b has suppressed autophagy through inhibiting ATG12.Citation12 However, the correlation between ATGs and miRs in AF cells is still not identified fully.
Melatonin has been reported as an endogenous neuro-hormone, which promotes proliferation and differentiation of neural stem cells.Citation13 Moreover, it has been confirmed that melatonin inhibits the expression of miR-23a in hepatic metabolic diseases.Citation14 In addition, melatonin suppressed the function of miR-34a in the regulation of neonatal brain inflammation.Citation15 A previous report has elaborated that melatonin promotes type I collagen synthesis in human bone cells, which is the major fibrillary collagen of AF.Citation16 Moreover, melatonin inhibits the apoptosis of neuronal cells.Citation17,Citation18 Additionally, melatonin regulates traumatic optic neuropathy via upregulating autophagy.Citation19 Therefore, targeting autophagy is a promising approach for mitigating DD.Citation20 However, the biological effect of melatonin on DDCitation16,Citation21,Citation22 is still not clear.
In this research, we systematically analyzed the effect of melatonin on AF cells of patients with DD. Our results not only gained a deep insight into the biological function of melatonin but also provided evidences to indicate its possible signaling pathway in AF cells of patients with DD.
Materials and methods
Cell culture
Annular tissues of patients with DD and healthy volunteers were washed three times with sterile saline solution under aseptic conditions. Then, all samples were transferred to a sterile centrifuge tube and cut into pieces smaller than 1 mm3. Next, type II collagenase (0.2%) and trypsin solution (0.25%) were used to digest samples followed by adding DMEM (Trueline, Nashville, TN, USA) that contained 10% fetal bovine serum (FBS; Thermo Fisher Scientific). Cells were dispersed by mechanical shaking. Then, cells were cultured in T25 cell culture media (replaced every 3 days) at 37°C, in a 5% CO2 incubator after counting. Subcultures were performed when the primary culture of cells were 90% confluent. After that, cells were washed twice using sterile PBS solution and digested by trypsin (0.25%) at 37°C for 2 minutes. The passage 2 cells were treated with melatonin (Aladdin, Shanghai, People’s Republic of China) with different concentrations, including 10, 25, 50, 100, and 200 µmol/L, and used for subsequent analysis.
All patients were informed and gave written consent. This study was in accordance with the Declaration of Helsinki. Moreover, our research was approved by the independent ethics committee of Peking University Third Hospital.
Immunofluorescence detection
Immunofluorescent detection of light chain 3 (LC3) associations with autophagosomes was carried out as follows. In brief, cells were fixed using paraformaldehyde (4%) for 30 minutes after incubation with nigericin. Then, samples were washed using PBS (0.02 mol) for 3 minutes three times at 25°C. After that, cells were blocked using BSA (1%; Solarbio, Beijing, People’s Republic of China) for 1 hour at room temperature. Subsequently, cells were incubated with the rabbit anti-LC3 antibody (abcam, UK) in PBS overnight at 4°C followed by goat anti-rabbit IgG (H+L) (Beyotime, Haimen, People’s Republic of China) for 1 hour at room temperature. Images were acquired by an ECLIPSE Ni microscope and a digital image analyzer (NIKON, Tokyo, Japan). Three replicates were needed for each analysis.
RNA extraction and real-time PCR
Total RNA from different samples were extracted by using TRIzol Reagent (Thermo Fisher Scientific, Waltham, MA, USA). Then, the cDNA synthesis kit (Thermo Fisher Scientific) was used to reverse transcribe RNA into complementary DNA (cDNA) according to the instructions of the manufacturer. GAPDH was used to normalize the gene expression and measured using the 2−ΔΔCt method. Three replicates were needed for each analysis. Details of the primers used in this study are provided in the Supplementary materials, primer sequence information.
Cell transfection
The miR-106a-5p mimics, inhibitor, and negative control (NC) were obtained from GenePharma (Shanghai, People’s Republic of China). Cells were grown to 80% confluence prior for transfection. Lipofectamine 2000 (Thermo Fisher Scientific) was used to transfect the miR-106a-5p mimics or inhibitor into AF cells according to the protocol of the manufacturer. Then 48 hours post-transfection, the efficacy of transfection was measured by qRT-PCR. The sequence information is provided in Table S1. Three replicates were needed for each analysis.
Western blot
Total protein lysates were extracted by using RIPA lysis buffer (JRDUN, Shanghai, People’s Republic of China) with EDTA-free Protease inhibitor Cocktail (Roche, Germany). The protein concentration was quantified by an Enhanced BCA Protein Assay kit (Thermo Fisher Scientific). Equal amounts of protein (25 µg) were fractionated on 10% SDS-PAGE and transferred to a nitrocellulose membrane (Millipore, Billerica, MA, USA) overnight. After being blocked with 5% non-fat dry milk for 1 hour at 25°C, the membranes were probed at 4°C overnight with the primary antibodies, followed by secondary antibody anti-mouse IgG (1:1,000; Beyotime) for 1 hour at 37 °C. An enhanced chemiluminescence system (Tanon, Shanghai, People’s Republic of China) was used to detect protein content. The detailed information of primary antibodies is provided in Table S2. Three replicates were needed for each analysis.
Cell proliferation
CCK-8 assay kits (SAB, College Park, MD, USA) were uti-lized to determine cell proliferation according to the protocol of the manufacturer. Briefly, cells transfected as indicated were seeded in 96-well plates and cultured for 0, 24, 48, and 72 hours, CCK-8 solution (1:10) was mixed in each well and incubated for 1 hour. A microplate reader (Pulangxin, Beijing, People’s Republic of China) was used to measure the OD value at 450 nm. Triplicates were performed at each time point.
Cell apoptosis
In brief, an Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit (Beyotime) was used to stain AF cells according to the instructions of the manufacturer after 48 hours of viral infection. Then, a flow cytometer (BD, San Diego, CA, USA) was used to detect the cells. Three replicates were needed for each analysis.
Dual-luciferase reporter assays
The ATG7 3′-UTR, containing the wild type or mutated target sequences of miR-106a-5p was cloned into Dual-Luciferase Expression Vector pGL3-Promoter (Promega, Madison, WI, USA). AF cells were transfected with NC or miR-106a-5p mimic in addition to the wild-type or mutant luciferase reporter, and incubated for 48 hours. Then, cells were harvested after 48 hours of transfection and then detected by using the Dual-GLO Luciferase Assay Kit (Promega). Three replicates were needed for each analysis.
Statistical analysis
GraphPad Prism software Version 7.0 (CA, USA) was uti-lized for the statistical analyses. Data were presented as the mean±SD of at least three samples. Statistical significance was determined by ANOVA for multiple comparisons and was accepted by a P-value<0.05.
Results
Melatonin stimulated proliferation, induced autophagy, and inhibited apoptosis in AF cells of patients with DD
To further examine the correlation between melatonin and DD, AF cells of patients with DD were cultured in the presence of melatonin with different concentrations, including 10, 25, 50, 100, and 200 µmol/L. The untreated cells acted as the blank control. CCK-8 assay was utilized to examine the effect of melatonin in the proliferation on AF cells. As the concentration of melatonin increased, the cell proliferation rate was gradually increased (). These results suggest that melatonin contributed to the proliferation of AF cells of patients with DD.
Figure 1 Overload of melatonin stimulated proliferation, induced autophagy, and apoptosis in AF cells of patients with DD. (A) Cell proliferation was detected 0, 24, 48, and 72 hours after treatment in AF cells of patients with DD treated with different melatonin concentrations. (B) Relative protein level of LC3-II in AF cells of patients with DD treated with 0, 25, 50, and 100 µmol/L of melatonin. (C) Representative images of immunofluorescent detection of LC3-II (green) in normal AF cells and AF cells of patients with DD. (D) The cell apoptosis profile of AF cells. (E and F) stand for the mRNA and protein level of MMP-3, ADAMTS-5, Aggrecan, COL2A1, and melatonin receptor 1A/1B in melatonin cultured cells.
Abbreviations: AF, annulus fibrosus; DD, disc degeneration; LC3, light chain 3; M0, M25, M50, and M100, Melatonin of 0, 25, 50, and 100 µmol/L; MR 1A/1B, melatonin receptor 1A/1B.
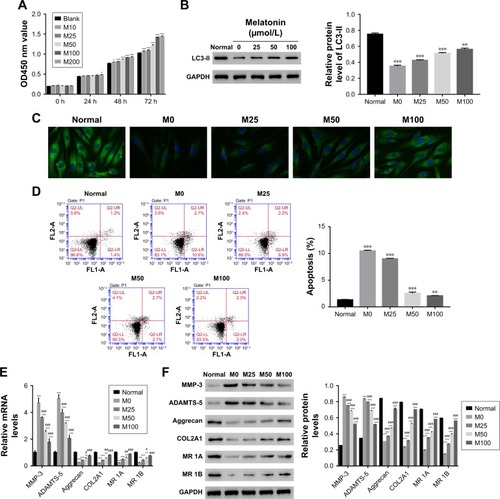
LC3-II has been reported as an autophagy-associated protein.Citation21 In this study, the protein level of LC3-II was significantly decreased in AF cells of patients with DD compared with that of normal AF cells. Moreover, it was easily identified that melatonin promoted the expression of LC3-II in AF cells of patient with DD, which was consistent with the result of LC3-II immunofluorescent detection (). In addition, the cell apoptosis rate of AF cells as indicated was much higher than that of normal AF cells. Interestingly, the cell apoptosis rate was deeply suppressed by melatonin (). All these results indicated that melatonin induced autophagy and inhibited apoptosis in AF cells of patients with DD.
In addition, the matrix metabolism is critical for the development of nucleus pulposus. The dysfunction of matrix metabolism leads to intervertebral disc degeneration.Citation22,Citation23 A previous report has indicated that melatonin inhibits the remodeling of the extracellular matrix (ECM) in nucleus pulposus (NP).Citation24
In this study, we also examined the effect of melatonin on matrix biomarkers in AF cells of patients with DD, including catabolic factors (MMP-3, ADAMTS-5) and anabolic genes (Aggrecan and COL2A1). As shown in , both the mRNA and protein of MMP-3 and ADAMTS-5 were deeply suppressed by melatonin. Meanwhile, the level of anabolic genes, Aggrecan, and COL2A1, were promoted by melatonin in AF cells as indicated. Taken together, all these results demonstrated that melatonin inhibited the degradation of ECM in AF cells as indicated. In addition, the level of melatonin receptors 1A and 1B was also significantly upregulated in AF cells of patients with DD.
Melatonin promoted the translation of autophagy-related proteins in AF cells of patients with DD
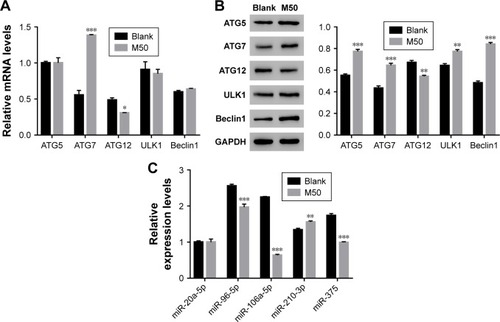
Next, quantitative real-time polymerase chain reaction (qRT-PCR) and Western blot were performed to quantify the mRNA and protein content of autophagy-related proteins, including ATG5, ATG7, ATG12, ULK1, and Beclin. The mRNA levels of ATG5, ULK1, and Beclin showed no significant difference between control and melatonin-treated AF cells (). However, the expression of ATG7 was significantly promoted by melatonin in AF cells of patients with DD. Interestingly, the expression of ATG12 was slightly inhibited by melatonin in AF cells. Moreover, nearly all indicated proteins (except for ATG12) were significantly upregulated in melatonin cultured cells (). Taken together, all these results indicated that melatonin affected the especially ATG7.
Figure 2 High level of melatonin affected the level of autophagy-related proteins and microRNAs in AF cells of patients with DD. (A and B) Show the mRNA and protein levels of ATG5, ATG7, ATG12, ULK1, and Beclin 1 in AF cells of patients with DD and treated with melatonin of 50 µmol/L. (C) The mRNA levels of miR-20a-5p, miR-96-5p, miR-106a-5p, miR-210-3p, and miR-375 in AF cells of patients with DD and treated with melatonin of 50 µmol/L.
Abbreviations: AF, annulus fibrosus; DD, disc degeneration; M50, melatonin of 50 µmol/L.
miR-106a-5p regulated ATG7 expression by directly binding to its 3′UTRin AF cells of patients with DD
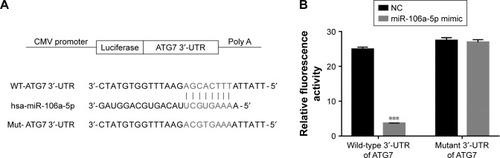
Some highly conserved regions in 3′UTR of ATG7 were investigated by using the Targetscan database (http://www.targetscan.org), which might have served as the binding sites for miRs. Then, we examined the level of five miRs that might target ATG7 in AF cells, including miR-20a-5p, miR-96-5p, miR-106a-5p, miR-210-3p, and miR-375. As shown in , most of them were downregulated in melatonin-treated AF cells, especially miR-106a-5p.
To further investigate the connection between miR-106a-5p and ATG7, luciferase reporters that contained the wild type (WT) or mutant 3′UTR of ATG7 were constructed (). As shown in , the luciferase activity of the wild type reporter was deeply abolished by the miR-106a-5p mimic. However, the luciferase activity of the mutant reporter showed no significant difference between miRs NC or miR-106a-5p mimic-transfected cells. Therefore, miR-106a-5p regulated the expression of ATG7 by directly binding to its 3′UTR in AF cells of patients with DD.
Figure 3 miR-106a-5p directly binds to the 3′-UTR region of ATG7. (A) Schematic of the luciferase construct with the ATG7 3′-UTR containing an miR-106a-5p binding sequence. (B) Dual-luciferase assays of miR-106a-5p and ATG7 3′-UTR. Wild-type or mutant 3′-UTR of ATG7 was cloned into luciferase reporter vectors. AF cells were transfected with NC or miR-106a-5p mimic in addition to the wild-type or mutant luciferase reporter vector and incubated for 48 hours.
Abbreviations: AF, annulus fibrosus; NC, negative control; UTR, untranslated region.
Melatonin inhibited the function of miR- 106a-5p in AF cells of patients with DD
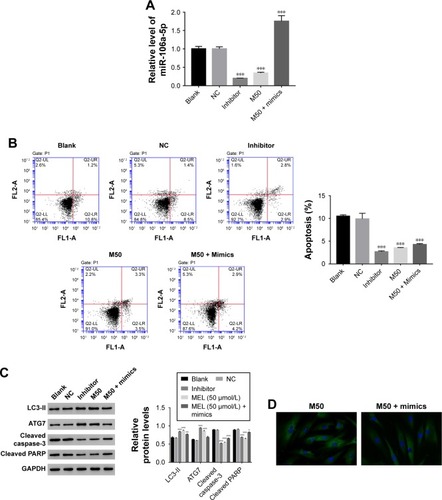
Then, we utilized gain or loss of function assay by transiently transfecting the miR-106a-5p mimic, inhibitor, and NC into AF cells. As shown in , all the transfected sequences functioned well in AF cells as expected. Moreover, our result suggested that miR-106a-5p was a pro-apoptotic element. However, melatonin deeply suppressed the function of miR-106a-5p in the cell apoptosis process (). Moreover, Western blot was performed to determine the protein levels of LC3-II, ATG7, cleaved caspase-3, and cleaved poly-ADP-ribose polymerases-1 (PARP). As shown in , the protein content of LC3-II was significantly upregulated in miR-106a-5p inhibitor transfected cells. Moreover, the protein content of LC3-II showed no significant difference between melatonin and miR-106a-5p mimics transfected cells. These results indicated that melatonin relieved the suppression of miR-106a-5p on ATG7. Meanwhile, cleaved caspase-3 and cleaved PARP-1 were positively correlated with miR-106a-5p. Moreover, the analysis of LC3-II immunofluorescent detection also obtained similar results (). Taken together, all these results demonstrated that melatonin suppressed the function of miR-106a-5p in AF cells of patients with DD.
Figure 4 Melatonin inhibited the effect of miR-106a-5p in AF cells of patients with DD. (A) The expression level of miR-106a-5p was examined in AF cells that transfected with negative control, inhibitor, and treated with M50 or M50 + mimics, respectively. (B) The cell apoptosis profile of AF cells of patients with DD. (C) The relative protein level of LC3-II, ATG7, cleaved caspase-3, and cleaved PARP in AF cells of patients with DD. (D) Representative images of immunofluorescent detection of LC3-II (green) in AF cells of patient with DD.
Abbreviations: AF, annulus fibrosus; DD, disc degeneration; LC3, light chain 3; M50, melatonin of 50 µmol/L; PARP, poly-ADP-ribose polymerases-1.
Discussion
DD is a major risk for back pain in advanced age, which has caused huge socioeconomic implications.Citation25 With the rising of the aging population, it is imperative to develop novel therapeutic approaches for the treatment of DD. The present study systematically analyzed the effect of melatonin on AF cells of patients with DD. Although the phenotype of the AF cells would change under monolayer culture conditions, and was different from the in-vivo condition of the IVD, our results gained a deep understanding into the effect of melatonin on AF cells.
Autophagy is an essential process for lysosomal degradation, which plays a key role in cell development.Citation26 Moreover, autophagy has been reported as a protective mechanism against apoptosis in AF cells and IVD degeneration.Citation7 In the present analysis, our result indicated that melatonin promoted the autophagy and inhibited the apoptosis of AF cells of patients with DD. Although the precise mechanism of disc degeneration is still not fully understood, our analysis indicated that melatonin might be a promising agent in the prevention of IVD.
It has been reported that ATG7 is essential for maintaining the homeostasis of axons and preventing their degeneration.Citation27 Previous report has indicated that ATG7 is negatively correlated with the activity of caspase-3.Citation28 In this study, we also obtained similar results. Therefore, melatonin might suppress the apoptosis of AF cells through inhibiting the activity of caspase-3 and PARP.
Moreover, PARP-1 was cleaved by activated caspases. Cleaved PARP-1 is reported as an apoptotic marker, which is responsible for DNA repair and cell viability.Citation29 Besides that, PARP is an important regulator in the beginning of autophagy.Citation30 Our results also indicated a negative correlation between ATG7 and cleaved PARP. Therefore, ATG7 was also involved in the regulation of the cell apoptosis process. Further analyses are needed to investigate the detailed correlation among ATG7, caspase-3, and PARP in AF cells.
It has been identified that miR-106a-5p suppresses cell metastases and promotes apoptosis.Citation31–Citation33 In this study, melatonin deeply inhibited the function of miR-106a-5p in AF cells of patients with DD. Moreover, our results demonstrated that miR-106a-5p suppressed the expression of ATG7 by directly binding to its 3′UTR in AF cells of patients with DD. However, this effect was deeply released by melatonin. Therefore, our results indicated miR-106a-5p might be an upstream component in the ATG7 signaling pathway. Melatonin might benefit DD through suppressing the miR-106a-5p/ATG7 signaling pathway.
Conclusion
The present study investigated the effect of melatonin on AF cells of patients with DD. Our results not only demonstrated that melatonin was a promising agent in the treatment for DD, but also indicated the possible signaling pathway it targets in AF cells.
Supplementary materials
Primer sequence information
Homo sapiens autophagy related 5 (ATG5), transcript variant 2, mRNA
NM_001286106.1
Primer F 5′ GGCTGAGTGAACATCTGAG 3′
Primer R 5′ CCCAGTTGCCTTATCTGAC 3′
Pos: 976–1208
Amplified product: Size: 233 bps
Homo sapiens autophagy related 7 (ATG7), transcript variant 2, mRNA
NM_001136031.2
Primer F 5′ ACCCAGTGACGCCAGATTTC 3′
Primer R 5′ AGGCAGGCACAGATGCTATG 3′
Pos: 2704–2808
Amplified product: Size: 105 bps
Homo sapiens autophagy related 12 (ATG12), transcript variant 5, mRNA
NM_001277783.1
Primer F 5′ AGCGTTTCGGTCTTGTTG 3′
Primer R 5′ TGAGGTCAGTCAGGAGTTTG 3′
Pos: 2277–2495
Amplified product: Size: 219 bps
Homo sapiens unc-51 like autophagy activating kinase 1 (ULK1), mRNA
NM_003565.2
Primer F 5′ GTCACACGCCACATAACAG 3′
Primer R 5′ TTCCCAGGACTCAGGATTC 3′
Pos: 4782–4981
Amplified product: Size: 200 bps
Homo sapiens beclin 1 (beclin 1), transcript variant 2, mRNA
NM_001313998.1
Primer F 5′ AGGGATGGAAGGGTCTAAG 3′
Primer R 5′ GGGCTGTGGTAAGTAATGG 3′
Pos: 161–315
Amplified product: Size: 155 bps
Homo sapiens glyceraldehyde-3-phosphatedehydrogenase (GAPDH), transcript variant 2, mRNA
NM_001256799.2
Primer F 5′AATCCCATCACCATCTTC 3′
Primer R 5′AGGCTGTTGTCATACTTC 3′
Pos: 436–653
Amplified product: Size: 218 bps
hsa-miR-106a-5p MIMAT000103
RT-Primer 5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTACCT 3′
PCR Primer:
Primer F 5′ CGCGAAAAGTGCTTACAGTGC 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
hsa-miR-20a-5p MIMAT000075
RT-Primer
5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTACCT 3′
PCR Primer:
Primer F 5′ GCGCGTAAAGTGCTTATAGTGC 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
hsa-miR-210-3p MIMAT000267
RT-Primer
5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAGCC 3′
PCR Primer:
Primer F 5′ CGCTGTGCGTGTGACAGC 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
hsa-miR-375 MI0000783
RT-Primer
5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGCCTCA 3′
PCR Primer: Primer F 5′ GTTCGTTCGGCTCGCG 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
hsa-miR-4802 MI0017450
RT-Primer
5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGCTT 3′
PCR Primer:
Primer F 5′ GGAAACCTTCAAGCAGGCC 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
hsa-miR-96-5p MIMAT000095
RT-Primer
5′ GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCAAA 3′
PCR Primer:
Primer F 5′ GCGTTTGGCACTAGCACATT 3′
Primer R 5′ AGTGCAGGGTCCGAGGTATT 3′
Homo sapiens RNA, U6 small nuclear 1 (RNU6-1), small nuclear RNA
NR_004394.1
Primer F 5′ CTCGCTTCGGCAGCACA 3′
Primer R 5′ AACGCTTCACGAATTTGCGT 3′
Pos: 4–97
Amplified product: Size: 94 bps
Table S1 The sequence information for analysis hsa-miR-106a-5p
Table S2 The primary antibodies information
Acknowledgments
This study was supported by the fund for Forstering Young Scholar of Peking University Health Science Center (NO BMU2017PY017).
Disclosure
The authors report no conflicts of interest in this work.
References
- MichaelAARoughleyPJWhat is intervertebral disc degeneration, and what causes it?Spine200631182151216110.1097/01.brs.0000231761.73859.2c16915105
- EckJCHumphreysSCHodgesSDAdjacent-segment degeneration after lumbar fusion: a review of clinical, biomechanical, and radiologic studiesAm J Orthop19992833634010401898
- ShuCCSmithMMSmithSMDartAJLittleCBMelroseJA histopathological scheme for the quantitative scoring of intervertebral disc degeneration and the therapeutic utility of adult mesenchymal stem cells for intervertebral disc regenerationInt J Mol Sci201718510.3390/ijms18051049
- LittleJPPearcyMJTevelenGEvansJHPettetGAdamCJThe mechanical response of the ovine lumbar anulus fibrosus to uniaxial, biaxial and shear loadsJ Mech Behav Biomed Mater20103214615710.1016/j.jmbbm.2009.09.00220129414
- DalyCGhoshPJenkinGOehmeDGoldschlagerTA review of animal models of intervertebral disc degeneration: pathophysiology, regeneration, and translation to the clinicBiomed Res Int20162016595216510.1155/2016/595216527314030
- KlionskyDJEmrSDAutophagy as a regulated pathway of cellular degradationScience200029054971717172111099404
- ShenCYanJJiangLSDaiLYAutophagy in rat annulus fibrosus cells: evidence and possible implicationsArthritis Res Ther2011134R13210.1186/ar344321846367
- AmbrosVThe functions of animal microRNAsNature200443135035510.1038/nature0287115372042
- BartelDPMicroRNAs: target recognition and regulatory functionsCell2009136221523310.1016/j.cell.2009.01.00219167326
- ChangYYanWHeXmiR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditionsGastroenterology20121431177187.e810.1053/j.gastro.2012.04.00922504094
- YuYZhangJJinYMiR-20a-5p suppresses tumor proliferation by targeting autophagy-related gene 7 in neuroblastomaCancer Cell Int201818510.1186/s12935-017-0499-229311760
- PanBChenLFengBChenYSongHMiR-200b regulates autophagy associated with chemoresistance in human lung adenocarcinomaOncotarget2015632328053282010.18632/oncotarget.535226416454
- FuJZhaoS-DLiuH-JHaoA-JMelatonin promotes proliferation and differentiation of neural stem cells subjected to hypoxia in vitroJ Pineal Res20115110411210.1111/j.1600-079X.2011.00867.x21392094
- KimS-JKangHSLeeJ-HMelatonin ameliorates ER stress-mediated hepatic steatosis through miR-23a in the liverBio-chem Biophys Res Commun2015458346246910.1016/j.bbrc.2015.01.117
- CarloniSFavraisGSalibaEMelatonin modulates neonatal brain inflammation through endoplasmic reticulum stress, autophagy, and miR-34a/silent information regulator 1 pathwayJ Pineal Res201661337038010.1111/jpi.1235427441728
- NakadeOKoyamaHArijiHYajimaAKakuTMelatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitroJ Pineal Res19992710611110496146
- MayoJCSainzRMUriaHIsaac AntolinMMERodriguezCMelatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells_ Implications for Parkinson’s diseaseJ Pineal Res1998241791929551855
- CagnoliCMAtabayCKharlamovaEManevHMelatonin protects neurons from singlet oxygen-induced apoptosisJ Pineal Reurvch19951822222610.1111/j.1600-079X.1995.tb00163.x
- WeiJMaLSLiuDJGuoJJiangWKYuHJMelatonin regulates traumatic optic neuropathy via targeting autophagyEur Rev Med Pharmacol Sci201721214946495129164563
- ZhangSJYangWWangCAutophagy: A double-edged sword in intervertebral disc degenerationClin Chim Acta2016457273510.1016/j.cca.2016.03.01627018178
- HoltSVWyspianskaBRandallKJJamesDFosterJRWilkinsonRWThe development of an immunohistochemical method to detect the autophagy-associated protein LC3-II in human tumor xenograftsToxicol Pathol201139351652310.1177/019262331039690321441228
- LiuWXiaPFengJMicroRNA-132 upregulation promotes matrix degradation in intervertebral disc degenerationExp Cell Res20173591394910.1016/j.yexcr.2017.08.01128793234
- WalkerMHAndersonDGMolecular basis of intervertebral disc degenerationSpine Jl2012154318330
- LiZLiXChenCChanMTVWuWKKShenJMelatonin inhibits nucleus pulposus (NP) cell proliferation and extracellular matrix (ECM) remodeling via the melatonin membrane receptors mediated PI3K-Akt pathwayJ Pineal Res201763310.1111/jpi.12435
- VasiliadisESPneumaticosSGEvangelopoulosDSPapavassiliouAGBiologic treatment of mild and moderate intervertebral disc degenerationMol Med20142040040910.2119/molmed.2014.0014525171110
- LevineBKroemerGAutophagy in the pathogenesis of diseaseCell20081321274210.1016/j.cell.2007.12.01818191218
- KomatsuMWangQJHolsteinGREssential role for autophagy protein Atg7 in the maintenance of axonal homeostasis and the prevention of axonal degenerationProc Natl Acad Sci U S A200710436144891449410.1073/pnas.070131110417726112
- BaydasGReiterRJAkbulutMTuzcuMTamerSMelatonin inhibits neural apoptosis induced by homocysteine in hippocampus of rats via inhibition of cytochrome c translocation and caspase-3 activation and by regulating pro- and anti-apoptotic protein levelsNeuroscience2005135387988610.1016/j.neuroscience.2005.05.04816213988
- CasaoAMata-CampuzanoMOrdasLCebrian-PerezJAMuino-BlancoTMartinez-PastorFCleaved PARP-1, an apoptotic marker, can be detected in ram spermatozoaReprod Domest Anim201550468869110.1111/rda.1254926031316
- Rodriguez-VargasJMRuiz-MaganaMJRuiz-RuizCROS-induced DNA damage and PARP-1 are required for optimal induction of starvation-induced autophagyCell Res20122271181119810.1038/cr.2012.7022525338
- PanYJWeiLLWuXJHuoFCMouJPeiDSMiR-106a-5p inhibits the cell migration and invasion of renal cell carcinoma through targeting PAK5Cell Death Dis2017810e315510.1038/cddis.2017.51829072688
- ZhiFZhouGShaoNmiR-106a-5p inhibits the proliferation and migration of astrocytoma cells and promotes apoptosis by targeting FASTKPLoS One201388e7239010.1371/journal.pone.007239024013584
- HeQYWangGCZhangHmiR-106a-5p suppresses the proliferation, migration, and invasion of osteosarcoma cells by targeting HMGA2DNA Cell Biol201635950652010.1089/dna.2015.312127383537