
Abstract
Purpose
Although increasing lines of evidence showed associations between serum uric acid (UA) levels and schizophrenia, the causality and the direction of the associations remain uncertain. Thus, we aimed to assess whether the relationships between serum UA levels and schizophrenia are causal and to determine the direction of the association.
Patients and Methods
Two-sample bidirectional Mendelian randomization (MR) analyses and various sensitivity analyses were performed utilizing the summary data from genome-wide association studies within the Global Urate Genetics Consortium and the Psychiatric Genomics Consortium. Secondary MR analyses in both directions were conducted within summary data using genetic risk scores (GRSs) as instrumental variables.
Results
Three MR methods provided no causal relationship between serum UA and schizophrenia. Furthermore, GRS approach showed similar results in the three MR methods after adjustment for heterogeneity. By contrast, inverse variance weighted method, weighted median and GRS approach suggested a causal effect of schizophrenia risk on serum UA after adjustment for heterogeneity (per 10-symmetric percentage increase in schizophrenia risk, beta: −0.039, standard error (SE): 0.013, P = 0.003; beta: −0.036, SE: 0.018, P = 0.043; beta: −0.039, SE: 0.013, P = 0.002; respectively). Moreover, in both directions’ analyses, the heterogeneity and sensitivity tests suggested no strong evidence of bias due to pleiotropy.
Conclusion
Schizophrenia may causally affect serum UA levels, whereas the causal role of serum UA concentrations in schizophrenia was not supported by our MR analyses. These findings suggest that UA may be a useful potential biomarker for monitoring treatment or diagnosis of schizophrenia rather than a therapeutic target for schizophrenia.
Introduction
Schizophrenia is a serious and considerably heritable mental illness resulting in severe disability.Citation1,Citation2 Life expectancy for people with schizophrenia is approximately 10–25 years shorter than that for the general people.Citation3,Citation4 Studies have indicated that schizophrenia may be related to oxidative damage, as abnormal levels of major antioxidants were observed in patients with schizophrenia.Citation5–Citation7 Circulating uric acid (UA) is a major antioxidant of plasma and protects cells from oxidative damage.Citation8,Citation9 Study reported approximately 60% of free radical scavenging activity was represented by UA in human blood.Citation10 In addition, patients with schizophrenia were found to have significantly lower UA levels than normal controls, and further decreased UA levels were observed after haloperidol withdrawal in the controlled clinical trial.Citation5 Meanwhile, in a case-control study, significantly lower UA levels were reported in patients with first-episode schizophrenia (FSPR) rather in controls, suggesting oxidative damage remains in the antioxidant system in the early course of schizophrenia.Citation7 Furthermore, a meta-analysis of cross-sectional studies reported every 1 mg/dL lower UA concentration decreased 23–59% FSPR risk.Citation11,Citation12
Although the association between UA and schizophrenia was observed in cumulative studies, whether reduced UA is a cause or a consequence of schizophrenia remains unclear. Reverse causality could be one of the explanations for this association since decreased UA is a defensive consequence of schizophrenia because of the UA antioxidative properties.Citation10 Moreover, it has been demonstrated that serum UA levels were related to numerous factors, such as dietary factors and smoking,Citation13 and such factors may be potential confounders and not all to be adjusted in the reported studies. Thus, understanding the role of UA in schizophrenia and insights into the causal nature of these observed relationships require further investigation.
It is well known that both schizophrenia and UA are highly heritable. A meta-analysis of twin studies and a Swedish population-based study estimated the heritability of schizophrenia ranged from 64% to 81%.Citation14,Citation15 A large genome-wide association study (GWAS) identified 128 single nucleotide polymorphisms (SNPs) significantly associated with schizophrenia which explained approximately 3.4% of the variance in schizophrenia risk.Citation16 As for UA, approximately 40–73% of heritability was estimated by the twin study,Citation17 path analyses and segregation analyses.Citation18,Citation19 Recently, 28 SNPs were identified significantly associated with serum UA level in a large GWAS,Citation20 and the proportion of explained variance was approximately 7%.
For those heritable phenotypes evidenced from GWAS, the causal association between them now can be assessed by Mendelian randomization (MR) study (ie, utilizing genetic variants as an instrumental variable (IV) to estimate the causal relationship between exposure and a medically relevant outcome).Citation21–Citation23 During gamete formation, alleles are randomly allocated and genetic variants have effects that are potentially lifelong. Thus, other confounding factors are probably not going to confound the relationship of genetic variants with risk of a disease outcome.Citation24 Moreover, unlike traditional epidemiology studies, association between genetic variants and outcomes cannot be generated by reverse causality, since genomes are determined at conception. So, we performed the bidirectional MR study to identify whether reverse causality is present in earlier epidemiology studies. MR study has been used for testing a causal effect for a range of traits and diseases,Citation25–Citation32 leading to our understanding for the aetiological architecture of complex diseases. Compared with one-sample MR, the two-sample MR method does not rely on individual-level genotype data and can estimate the causal association between exposure factors and outcome variables through the existing GWAS summarized statistics.Citation33 Thus, the causal association between UA and schizophrenia risk can be assessed by MR. In the present study, we will use GWAS summarized data to analyze the causal relationship between serum UA levels and schizophrenia risk with two-sample bidirectional MR method.
Materials and Methods
The schematic of the two-sample bidirectional MR analysis is shown in . Briefly, serum UA levels affect schizophrenia risk were analyzed first (), then followed the analysis on schizophrenia risk affects serum UA levels ().
Figure 1 Two-sample bidirectional Mendelian randomization study of the association of serum uric acid (UA) and schizophrenia.
Abbreviations: SNP, single nucleotide polymorphism; GUGC, Global Urate Genetics Consortium; PGC, Psychiatric Genomics Consortium; MR, Mendelian randomization.
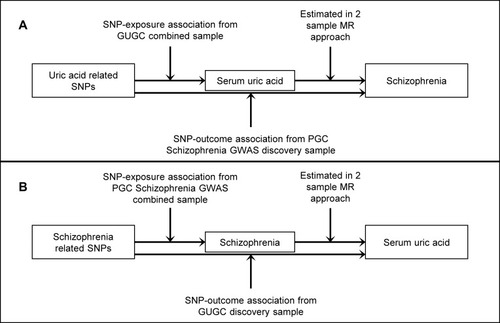
Selection of Datasets and Genetic Variants
Serum UA to Schizophrenia
The datasets from the serum UA Global Urate Genetics Consortium (GUGC) GWAS and the Psychiatric Genomics Consortium (PGC) GWAS were used for the analysis. The GUGC GWAS comprised more than 140 000 individuals of European ancestry.Citation20 Twenty-eight SNPs associated with serum UA levels were selected from GUGC GWAS, which were identified to be of genome-wide significance (P<5×10−8) in the combined sample (discovery sample + replication sample). These variants are not in linkage disequilibrium and were used in the previous MR studies.Citation34,Citation35 A central assumption of MR is that the SNPs included in the genetic instrument are uncorrelated with confounders.Citation22 In the context of UA – schizophrenia/schizophrenia – UA relationship, smoking is most likely a very important confounder.Citation36,Citation37 Information queried from publicly available databases of human genotype-phenotype associations, PhenoScanner V2 and GWAS Catalog, was used to assess associations of these SNPs with smoking confounder.Citation38,Citation39 Of the 28 SNPs related to serum UA therein, we excluded 1 SNP (rs653178) for being related with smoking confounder (Table S1) and 1 SNP (rs17632159) for being palindromic.Citation40 Finally, 26 independent SNPs were used as IVs for serum UA. All these 26 SNPs associated with serum UA were directly matched in the summary data of gene (SNP)–outcome (schizophrenia risk) association estimates.
We obtained the summary data of gene (SNP)–outcome (schizophrenia risk) association estimates from the PGC GWAS discovery sample,Citation16 which included 35,476 cases of schizophrenia and 46,839 controls. The summary statistics were downloaded from the public access (http://www.med.unc.edu/pgc/downloads (SCZ2)).
We also conducted data harmonization to prevent bias due to errors according to a previous report (Tables S2 and S3).Citation40
Schizophrenia to Serum UA
We utilized variants at genome-wide significance (P<5×10−8) as genetic instrument for schizophrenia; these variants were from the combined sample (discovery sample + replication sample) of the PGC schizophrenia GWAS, which comprised 36,989 cases and 113 075 controls (principally of European ancestry).Citation16 A total of 128 SNPs were identified genome-wide significance and 2 SNPs (rs11210892, rs8042374) were excluded for being related with smoking confounder queried from PhenoScanner V2 and GWAS Catalog (Table S4),Citation38,Citation39 and 7 SNPs (rs11139497, rs12325245, rs12522290, rs215411, rs2851447, rs4240748, rs9841616) were excluded for being palindromic.Citation40 Of the remaining 119 SNPs, 45 SNPs were directly matched in the summary data of gene (SNP)–outcome (serum UA) association estimates. Finally, 45 independent SNPs were used as IVs for schizophrenia.
We obtained the summary data of gene (SNP)–outcome (serum UA) association estimates from the GUGC GWAS discovery sample of up to 110 347 individuals from 48 studies.Citation20
We also conducted data harmonization to prevent bias due to errors according to a previous report (Tables S5 and S6).Citation40
Statistical Analyses
Statistical Analyses for MR
Three MR methods were used in two sets of two-sample MR analyses: (1) inverse variance weighted (IVW) method,Citation41 in which the SNP–outcome estimation is regressed on the SNP–exposure estimation. This approach assumes that all SNPs are valid instruments; thus, the overall bias is zero; (2) MR Egger regression analysis, whose slope represents the causal effect estimate,Citation42 is robust to invalid instruments against directional pleiotropy;Citation43 and (3) weighted median approach, which assesses consistently regardless of whether up to half of the weight comes from invalid instruments.Citation44
In the MR analyses, ln (exposure) was used in both directions. That is, in the calculation of the effect size values for every exposure SNP, a unit increment in ln (exposure) is equivalent to a 10-symmetric percentage (s%) increment in exposure level, which corresponds to the beta coefficient (ie, logit of risk estimate) for a binary outcome or a unit change of a continuous outcome.Citation45 Therefore, the causal estimate value for every exposure SNP is displayed for the outcome, which corresponds to a 10-s% increment in real exposure level.
We conducted all analyses utilizing R (version 3.5.3) with the R package “TwosampleMR”.Citation33 We transformed all the causal effect estimates (beta coefficients) to odds ratios (ORs). Two-tail P<0.05 was considered statistically significant. We used only freely accessible summarized data in this study; therefore, this work did not require ethical approval.
Heterogeneity and Sensitivity Tests
Cochran’s Q test was used to estimate heterogeneities between SNPs.Citation46 This test assumes that all valid IVs assess the equivalent effect.Citation47 A P-value of <0.1 was considered statistically significant (ie, indicating the possibility of pleiotropy) for the Cochran’s Q test. We removed stepwise the SNPs with potential pleiotropic effects (ie, after adjustment for heterogeneity) to limit false positives,Citation48 and then repeated the analyses until the Cochran’s Q test did not differ from the null. We then performed MR Egger regression, whose intercept represents bias due to directional pleiotropy.Citation42 For MR Egger regression, intercept P<0.05 (the MR Egger regression) indicates SNPs with directional pleiotropy.Citation42 Furthermore, a “leave-one-out” analysis was conducted to assess whether the causal estimate was driven by a single SNP.
Power Calculation and Weak Instrument Bias
For the forward-direction MR analysis (SNPs–serum UA–schizophrenia), the power was assessed at a two-sided α of 0.05 in accordance with the method of Brion et al (https://shiny.cnsgenomics.com/mRnd/) (Table S7).Citation49 The forward-direction MR analysis would have 100% power to detect a statistically significant effect. The power level ensured that the associations did not arise from chance.
In MR, an F-statistic value of more than 10 reflects as a strong instrument.Citation50 The F-statistic value for each of the 45 SNP IVs for schizophrenia was more than 10 in the reverse-direction MR (Table S8), which meant the IVs for schizophrenia were strong and a causal estimate was unlikely to be biased due to weak instruments.
Sample Overlap
Conventionally, if overlap data sets on the SNP–exposure and SNP–outcome are used, the estimates can be biased and a spurious causal estimate can be yielded.Citation51 In the present study, data from the serum UA GUGC GWAS and the PGC GWAS on the exposure and outcome were taken from non-overlapping datasets,Citation16,Citation20 suggesting that the estimate was less biased and any bias was in the direction of the null.
Associations Between Exposure Genetic Risk Score (GRS) and Outcome in Both Directions
To obtain the combined estimate of the relationship of exposure-influencing alleles with the outcome, MR analyses were performed using weighted GRS as IVs in both directions by using the same summary data described above. In forward-direction MR, the GRS consisted of 21 SNPs (after removal of five SNPs with potential pleiotropic effects from the 26 IVs (SNPs)) that were related to serum UA level (P<5×10−8) in the combined sample (discovery sample + replication sample) of the GUGC GWAS (GRSUA).Citation20 In the reverse-direction MR, the GRS comprised 44 SNPs (after removal of one SNP with potential pleiotropic effects from the 45 IVs (SNPs)) that were related to schizophrenia (P<5×10−8) in the combined sample (discovery sample + replication sample) of the PGC GWAS (GRSSCZ).Citation16 We conducted the analyses utilizing R (version 3.5.3) with the “gtx” R package (version 0.0.8 for Windows), whose grs.summary module has the GRS function. The grs.summary module merely used single SNP association summarized data obtained from the results of the GWAS analysis, which is similar to a method which regresses an outcome onto an additive GRS. For uncorrelated SNPs, the causal estimate α value can be estimated by , and the standard error seα can be estimated by
. Here, ω denotes the estimated effects on the intermediate trait or biomarker, and β values are estimated effects on the response variable or outcome with standard errors seβ. This approach is described in detail elsewhere.Citation52 Furthermore, previous studies reported that this MR method utilizing meta-GWAS summary data was equally efficient to that using individual-level data.Citation41,Citation53 Moreover, goodness-of-fit tests were conducted to adjust for the heterogeneity of SNPs in both GRSs.
Results
MR Results of Serum UA to Schizophrenia
All the three MR approaches (IVW method, MR Egger regression and weighted median approach) provided no causal relationship between serum UA and schizophrenia (per 10-s% increment in UA, OR: 0.942, 95% confidence interval (95% CI): 0.866–1.025, P = 0.163; OR: 0.964, 95% CI: 0.854–1.087, P = 0.555; OR: 0.965, 95% CI: 0.908–1.026, P = 0.260; respectively) (). Similar results were observed by using the three different MR methods after removing the five SNPs with potential pleiotropic effects (rs1165151, rs3741414, rs478607, rs6770152, rs7953704), indicating the lack of causal association between serum UA and schizophrenia (per 10-s% increment in UA, OR: 0.962, 95% CI: 0.908–1.020, P = 0.195; OR: 0.972, 95% CI: 0.895–1.054, P = 0.496; OR: 0.968, 95% CI: 0.908–1.031, P = 0.307; respectively) ().
Table 1 Results of Two-Sample Bidirectional MR Analyses on the Causal Effects Between Serum UA and Schizophrenia
MR Results of Schizophrenia to Serum UA
The results of IVW method showed a causal effect of schizophrenia risk on serum UA before and after removal of the SNP (rs10803138) with potential pleiotropic effects (per 10-s% increase in schizophrenia risk, beta: −0.045, standard error (SE): 0.014, P = 0.002; beta: −0.039, SE: 0.013, P = 0.003; respectively) (). Meanwhile, a causal relationship between schizophrenia risk and serum UA was demonstrated by the weighted median approach before and after removal of the SNP (rs10803138) with potential pleiotropic effects as well (per 10-s% increase in schizophrenia risk, beta: −0.038, SE: 0.018, P = 0.036; beta: −0.036, SE: 0.018, P = 0.043; respectively) (). However, no causal association was observed between schizophrenia risk and serum UA by using MR Egger regression analysis before and after removal of the SNP (rs10803138) with potential pleiotropic effects (per 10-s% increase in schizophrenia risk, beta: 0.143, SE: 0.099, P = 0.159; beta: 0.134, SE: 0.091, P = 0.147; respectively) (). The result of MR Egger regression was inconsistent with those findings from IVW and weighted median method.
GRSUA and Schizophrenia, GRSSCZ and Serum UA
Consistent with the MR results of serum UA to schizophrenia, the GRSUA revealed no causal effect of serum UA on schizophrenia risk (per 10-s% increment in UA, OR: 0.963, 95% CI: 0.913–1.015, P = 0.154) (, ). Meanwhile, a consistent causal relationship was found with the MR results of schizophrenia to serum UA. The GRSSCZ showed a significant effect of schizophrenia on serum UA (per 10-s% increase in schizophrenia risk, beta: −0.039, SE: 0.013, P = 0.002) (, ).
Figure 2 Genetic risk score GRSUA for schizophrenia.
Abbreviation: OR, odds ratio.
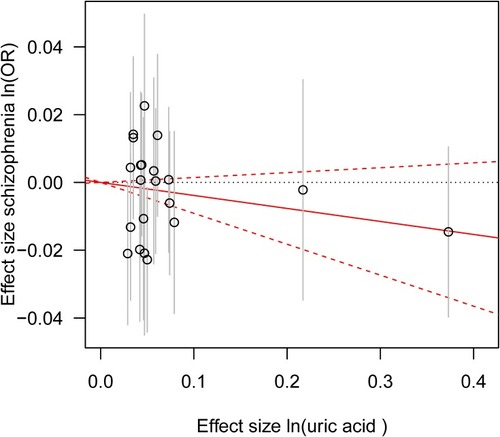
Figure 3 Genetic risk score GRSSCZ for serum UA.
Abbreviation: OR, odds ratio.
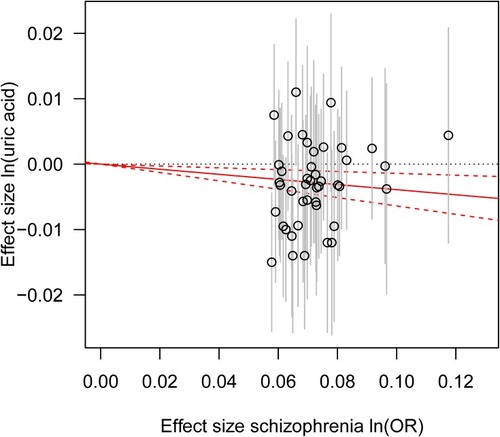
Table 2 The Effect of the GRS Instrument of Serum UA on Schizophrenia and the Effect of the GRS Instrument of Schizophrenia on Serum UA
Heterogeneity and Sensitivity Tests
Cochran’s Q test showed certain heterogeneity among both serum UA IV estimates based on the 26 SNPs and schizophrenia IV estimates based on the 45 SNPs (). The heterogeneity may indicate the possibility of pleiotropic effects of the SNPs. We subsequently performed a stepwise removal of SNPs until the Cochran’s Q test did not differ from the null, and finally, 5 SNPs were removed from the serum UA IV and 1 SNP was removed from the schizophrenia IV (). Of note, the MR Egger regression revealed no evidence that the result was likely to be biased by directional pleiotropy for serum UA to schizophrenia MR before and after removal of the five SNPs with potential pleiotropic effects (intercept = −0.003, P = 0.606; intercept = −0.001, P = 0.742; respectively) and for schizophrenia to serum UA MR before and after removal of the SNP with potential pleiotropic effects (intercept = −0.014, P = 0.063; intercept = −0.013, P = 0.060; respectively) (). The “leave-one-out” analysis showed the IVW causal association estimate was not driven by a single SNP either (Figures S1−S4).
Table 3 Cochran’s Heterogeneity Statistic and MR Egger Intercept, Indicating Horizontal Pleiotropy for Two-Sample Bidirectional MR Analyses Between Serum UA and Schizophrenia
As for GRSexposure and outcome, the results showed no evidence of the heterogeneity effects on outcome risk relative to the estimated effects on exposure in both directions (Qrs = 24.191, P-Het = 0.234 in forward-direction MR; Qrs = 46.941, P-Het = 0.314 in reverse-direction MR) (). Taken together, the results of both directions were unlikely to be biased by directional pleiotropy.
Discussion
To our knowledge, this is the first to use a two-sample bidirectional MR approach with various sensitivity analyses to investigate the causal relationship between serum UA and schizophrenia. Our results provide evidence that the effect of schizophrenia risk on serum UA may be causal, whereas the MR estimate for the serum UA-to-schizophrenia relation is unlikely to be causal.
The published observational studies on the association between serum UA levels and schizophrenia have yielded inconsistent results.Citation54–Citation57 The case-control studies found the schizophrenia group had lower serum UA levels than the healthy control group.Citation56,Citation57 However, Gültekin et al found UA levels in patients with schizophrenia were higher than in healthy males,Citation55 and a cross-sectional study did not find significant differences in blood UA levels between patients with schizophrenia and controls.Citation54 All these studies did not control for the dietary confounding factors which may affect the serum UA levels.Citation13 Confounding factors (dietary factors and smoking) and other residual confounding elements naturally could not be fully excluded in epidemiological studies,Citation13 which might bias the observed estimates. A way to control for confounding factors is MR study, which uses genetic IVs that were randomly allocated during conception; thus, lifestyle or sociodemographic factors are probably not going to confound genetic predictors of exposure, as stated previously.Citation58 Therefore, we used SNPs as IVs to assess whether serum UA is associated with schizophrenia and determine whether the association is causally independent of confounding factors by MR.
In the present study, the results from all the three MR methods provided no causal relationship between serum UA and schizophrenia risk. Cochran’s Q test suggested heterogeneity in the first forward-direction MR (serum UA to schizophrenia MR before removal of the 5 SNPs with potential pleiotropic effects). After excluding the 5 SNPs with potential pleiotropic effects, the three MR methods’ estimates still provided no causal relationship between serum UA and schizophrenia risk. Moreover, Cochran’s Q test did not show heterogeneity among the serum UA IV estimates, and MR Egger regression result did not show any bias by directional pleiotropy. Therefore, bias may not exist by not violating one of the assumptions of MR that any genetic effect on the outcome was indirect but via exposure. In accordance with these MR results, the results from GRSUA and schizophrenia analysis also supported that serum UA was not causally related to schizophrenia.
Our study also indicated the effect of schizophrenia risk on serum UA may be causal. Both IVW method and weighted median approach suggested a causal effect of the schizophrenia risk on serum UA, and the MR Egger regression analysis suggested a null causal effect. When the genetic variants are uncorrelated, the IVW method using summarized data gives similar estimates to the two-stage least squares estimate that commonly used in the individual-level data.Citation41 Furthermore, the IVW method had greater empirical power than that of two-stage least squares estimate.Citation41 As for the weighted median approach, which assesses consistently when up to half of the weight comes from invalid IVs, it is shown to have better finite-sample Type 1 error rates than the IVW method.Citation44 Moreover, compared to MR Egger regression analysis, the weighted median approach has the strength of holding prominent accuracy in the estimates.Citation44 Thus, the effect of the schizophrenia risk on serum UA was potentially causal, as indicated by the IVW and weighted median approach. When we further analyzed the relationship between GRSSCZ and serum UA, we found GRSSCZ was related to serum UA as well. The schizophrenia SNPs, separately or combined as the GRS, were causally related to serum UA. Taken together, the genetic risk for schizophrenia may causally reduce the serum UA levels.
Recent studies have shown that the prevalence of smoking in patients with schizophrenia was about 60%.Citation37,Citation59 Some evidence has demonstrated that smoking reduced serum UA levels.Citation36,Citation60,Citation61 So smoking may be a potential confounder for the relationship between serum UA and schizophrenia risk. Considering the confounding effects, we removed those SNPs associated with smoking as well as associated with UA and schizophrenia, and the causal relationship between schizophrenia and UA still persisted. Previously, studies have indicated the schizophrenia-to-low serum UA causality is biologically plausible. Schizophrenia is related to a complex pathophysiology and is a result of radical-mediated neurotoxicity.Citation56 The antioxidative properties of UA include reaction with and inactivation of peroxynitrite,Citation62 nitric oxide and hydroxyl radical.Citation63,Citation64 In the deactivating procedure, UA is demolished and easily oxidized at a comparable rate.Citation10 Because of the antioxidative properties, serum UA levels deplete in a state of oxidative stress, as observed in patients with schizophrenia. Furthermore, UA is the final product of purine metabolism (xanthine oxidase catalyzes the conversion of xanthine to UA), in which the conversions of guanosine to guanine or xanthosine to xanthine are reversible.Citation2 In patients with FSPR, significantly higher levels of xanthosine and lower levels of guanine were seen compared to healthy control subjects, and diminished product–precursor ratios indicate a shift favorable to xanthosine production from xanthine, thereby decreasing the UA levels.Citation2 Therefore, a low UA may be a consequence of schizophrenia. A study on the metabolomics of schizophrenia observed a reduction of UA in the patient with schizophrenia group in comparison with the control group, suggesting that UA is a potential biomarker for monitoring the therapy or diagnosis of schizophrenia.Citation65 Although decreased UA is a defensive consequence of schizophrenia, further studies are needed to clarify precise mechanisms.
Our study has several strengths. First, the sample size is large. In particular, the summary data from PGC GWAS comprising a total of 36,989 schizophrenia cases increased our analysis power. Moreover, our study using MR methods as well as GRS analysis increases the reliability of the results, and both showed a consistent result. Our sensitivity analyses also indicated robustness. To the best of our knowledge, no existing MR study has investigated on the causal relationship between serum UA and schizophrenia. Our work is the first to use a two-sample bidirectional MR approach with a variety of sensitivity analyses to investigate the causal relationship between serum UA and schizophrenia.
However, our study has several limitations. First, only 45 SNPs of the 119 remaining non-palindromic SNPs significantly associated with schizophrenia were matched in the outcome data directly and used as IVs for schizophrenia. We also did not find proxy SNPs in the schizophrenia-to-serum UA MR. If the relationships of the excluded SNPs with serum UA differed from the relationships of the included SNPs, then our results would be biased. However, using excessive proxy SNPs may produce unreliable results. Second, the subjects used in our analyses mostly are European ancestry, which minimized the possibility of population stratification bias. However, given that causality may rely upon ethnicity and selection bias, performing further MR studies in other populations is expected. Finally, the summarized data in our MR analyses did not classify schizophrenia (ie, into FSPR and chronic schizophrenia). Hence, further stratification of schizophrenia needs to conduct in future analyses if possible.
Conclusion
We conducted the first two-sample bidirectional MR analysis to assess the causal relationship between serum UA concentrations and schizophrenia. A causal role of serum UA concentrations in schizophrenia was not supported by our MR analyses, suggesting that diminishing serum UA levels may not mean hazard increment for schizophrenia in European individuals and UA does not seem to be a relevant therapeutic target. By contrast, our evidence supported the causal effect of schizophrenia risk on serum UA, suggesting that serum UA may be a potential biomarker for monitoring the therapy or diagnosis of schizophrenia. Earlier observational studies were susceptible to reverse causation because it is the pathophysiology, behavior or treatment of people with schizophrenia seem to influence uric acid levels.
Abbreviations
UA, uric acid; MR, Mendelian randomization; GRS, genetic risk score; FSPR, first-episode schizophrenia; GWAS, genome-wide association study; SE, standard error; IV, instrumental variable; SNP, single nucleotide polymorphism; GUGC, Global Urate Genetics Consortium; PGC, Psychiatric Genomics Consortium; IVW, inverse variance weighted; s%, symmetric percentage; OR, odds ratio; CI, confidence interval.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work.
Acknowledgments
We thank the GUGC and the PGC for providing the summarized data from the GWAS of UA and schizophrenia for these analyses. This work was supported by National Natural Science Foundation of China [grant number 81770759]; the Major Project of Guangxi Innovation Driven [grant number AA18118016]; and National Key Research and Development Program of China [grant number 2017YFC0908000]. The funding sources had no role in the study design, data collection, data analysis, results interpretation of the present study, and in the writing of the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
- van Dongen J, Boomsma DI. The evolutionary paradox and the missing heritability of schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2013;162B(2):122–136. doi:10.1002/ajmg.b.3213523355297
- Yao JK, Dougherty GG Jr, Reddy RD, et al. Homeostatic imbalance of purine catabolism in first-episode neuroleptic-naive patients with schizophrenia. PLoS One. 2010;5(3):e9508. doi:10.1371/journal.pone.000950820209081
- Laursen TM. Life expectancy among persons with schizophrenia or bipolar affective disorder. Schizophr Res. 2011;131(1–3):101–104. doi:10.1016/j.schres.2011.06.00821741216
- Laursen TM, Munk-Olsen T, Vestergaard M. Life expectancy and cardiovascular mortality in persons with schizophrenia. Curr Opin Psychiatr. 2012;25(2):83–88. doi:10.1097/YCO.0b013e32835035ca
- Yao JK, Reddy R, van Kammen DP. Reduced level of plasma antioxidant uric acid in schizophrenia. Psychiatry Res. 1998;80(1):29–39. doi:10.1016/S0165-1781(98)00051-19727961
- Yao JK, Reddy R, van Kammen DP. Abnormal age-related changes of plasma antioxidant proteins in schizophrenia. Psychiatry Res. 2000;97(2–3):137–151. doi:10.1016/S0165-1781(00)00230-411166086
- Reddy R, Keshavan M, Yao JK. Reduced plasma antioxidants in first-episode patients with schizophrenia. Schizophr Res. 2003;62(3):205–212. doi:10.1016/S0920-9964(02)00407-312837516
- Pingmuangkaew P, Tangvarasittichai O, Tangvarasittichai S. Association of elevated serum uric acid with the components of metabolic syndrome and oxidative stress in abdominal obesity subjects. Indian J Clin Biochem. 2015;30(3):286–292. doi:10.1007/s12291-014-0462-026089614
- Ishizaka Y, Yamakado M, Toda A, Tani M, Ishizaka N. Relationship between serum uric acid and serum oxidative stress markers in the Japanese general population. Nephron Clin Pract. 2014;128(1–2):49–56. doi:10.1159/00036245625342428
- Ames BN, Cathcart R, Schwiers E, Hochstein P. Uric acid provides an antioxidant defense in humans against oxidant- and radical-caused aging and cancer: a hypothesis. Proc Natl Acad Sci U S A. 1981;78(11):6858–6862. doi:10.1073/pnas.78.11.68586947260
- Li X, Meng X, Timofeeva M, et al. Serum uric acid levels and multiple health outcomes: umbrella review of evidence from observational studies, randomised controlled trials, and Mendelian randomisation studies. BMJ. 2017;357:j2376. doi:10.1136/bmj.j237628592419
- Flatow J, Buckley P, Miller BJ. Meta-analysis of oxidative stress in schizophrenia. Biol Psychiatry. 2013;74(6):400–409. doi:10.1016/j.biopsych.2013.03.01823683390
- Pae C-U, Paik I-H, Lee C, Lee S-J, Kim J-J, Lee C-U. Decreased plasma antioxidants in schizophrenia. Neuropsychobiology. 2004;50(1):54–56. doi:10.1159/00007794215179021
- Sullivan PF, Kendler KS, Neale MC. Schizophrenia as a complex trait: evidence from a meta-analysis of twin studies. Arch Gen Psychiatry. 2003;60(12):1187–1192. doi:10.1001/archpsyc.60.12.118714662550
- Lichtenstein P, Yip BH, Bjork C, et al. Common genetic determinants of schizophrenia and bipolar disorder in Swedish families: a population-based study. Lancet. 2009;373(9659):234–239. doi:10.1016/S0140-6736(09)60072-619150704
- Schizophrenia Working Group of the Psychiatric Genomics C. SBiological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511(7510):421–427. doi:10.1038/nature1359525056061
- Kalousdian S, Fabsitz R, Havlik R, Christian J, Rosenman R. Heritability of clinical chemistries in an older twin cohort: the NHLBI Twin Study. Genet Epidemiol. 1987;4(1):1–11. doi:10.1002/gepi.13700401023569874
- Rice T, Vogler GP, Perry TS, Laskarzewski PM, Province MA, Rao DC. Heterogeneity in the familial aggregation of fasting serum uric acid level in five North American populations: the lipid research clinics family study. Am J Med Genet. 1990;36(2):219–225. doi:10.1002/(ISSN)1096-86282368810
- Wilk JB, Djousse L, Borecki I, et al. Segregation analysis of serum uric acid in the NHLBI family heart study. Hum Genet. 2000;106(3):355–359. doi:10.1007/s00439000024310798367
- Kottgen A, Albrecht E, Teumer A, et al. Genome-wide association analyses identify 18 new loci associated with serum urate concentrations. Nat Genet. 2013;45(2):145–154. doi:10.1038/ng.250023263486
- Smith GD, Ebrahim S. Mendelian randomization: prospects, potentials, and limitations. Int J Epidemiol. 2004;33(1):30–42. doi:10.1093/ije/dyh13215075143
- Sheehan NA, VD P, Burton R, Tobin MD. Mendelian randomisation and causal inference in observational epidemiology. PLoS Med. 2008;5(8):1205–1210. doi:10.1371/journal.pmed.0050177
- Bochud M, Rousson V. Usefulness of Mendelian randomization in observational epidemiology. Int J Environ Res Public Health. 2010;7(3):711–728. doi:10.3390/ijerph703071120616999
- Smith GD, Ebrahim S. ‘Mendelian randomization’: can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003;32(1):1–22. doi:10.1093/ije/dyg07012689998
- Qian Y, Ye D, Wu DJ, et al. Role of cigarette smoking in the development of ischemic stroke and its subtypes: a Mendelian randomization study. Clin Epidemiol. 2019;11:725–731. doi:10.2147/CLEP.S21593331616189
- Gao X, Meng LX, Ma KL, et al. The bidirectional causal relationships of insomnia with five major psychiatric disorders: a Mendelian randomization study. Eur Psychiatry. 2019;60:79–85. doi:10.1016/j.eurpsy.2019.05.00431234011
- Polimanti R, Gelernter J, Stein DJ. Genetically determined schizophrenia is not associated with impaired glucose homeostasis. Schizophr Res. 2018;195:286–289. doi:10.1016/j.schres.2017.10.03329092750
- Hartwig FP, Borges MC, Horta BL, Bowden J, Davey Smith G. Inflammatory biomarkers and risk of schizophrenia: a 2-sample Mendelian randomization study. JAMA Psychiatry. 2017;74(12):1226–1233. doi:10.1001/jamapsychiatry.2017.319129094161
- Noyce AJ, Kia DA, Hemani G, et al. Estimating the causal influence of body mass index on risk of parkinson disease: a Mendelian randomisation study. PLoS Med. 2017;14(6):e1002314. doi:10.1371/journal.pmed.100231428609445
- Xu L, Borges MC, Hemani G, Lawlor DA. The role of glycaemic and lipid risk factors in mediating the effect of BMI on coronary heart disease: a two-step, two-sample Mendelian randomisation study. Diabetologia. 2017;60(11):2210–2220. doi:10.1007/s00125-017-4396-y28889241
- Zhang B, Shu X-O, Delahanty RJ, et al. Height and breast cancer risk: evidence from prospective studies and Mendelian randomization. J Natl Cancer Inst. 2015;107:11. doi:10.1093/jnci/djv219
- Casas JP, Bautista LE, Smeeth L, Sharma P, Hingorani AD. Homocysteine and stroke: evidence on a causal link from mendelian randomisation. Lancet. 2005;365(9455):224–232. doi:10.1016/S0140-6736(05)70152-515652605
- Hemani G, Zheng J, Elsworth B, et al. The MR-base platform supports systematic causal inference across the human phenome. Elife. 2018;7:e34408. doi:10.7554/eLife.3440829846171
- Keenan T, Zhao W, Rasheed A, et al. Causal assessment of serum urate levels in cardiometabolic diseases through a Mendelian randomization study. J Am Coll Cardiol. 2016;67(4):407–416. doi:10.1016/j.jacc.2015.10.08626821629
- Yuan H, Yang W. Genetically determined serum uric acid and alzheimer’s disease risk. J Alzheimers Dis. 2018;65(4):1259–1265. doi:10.3233/JAD-18053830149453
- Tsuchiya M, Asada A, Kasahara E, Sato EF, Shindo M, Inoue M. Smoking a single cigarette rapidly reduces combined concentrations of nitrate and nitrite and concentrations of antioxidants in plasma. Circulation. 2002;105(10):1155–1157. doi:10.1161/hc1002.10593511889006
- de Leon J, Diaz FJ. A meta-analysis of worldwide studies demonstrates an association between schizophrenia and tobacco smoking behaviors. Schizophr Res. 2005;76(2–3):135–157. doi:10.1016/j.schres.2005.02.01015949648
- Kamat MA, Blackshaw JA, Young R, et al. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics. 2019;35(22):4851–4853. doi:10.1093/bioinformatics/btz46931233103
- Buniello A, MacArthur JAL, Cerezo M, et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019;47(D1):D1005–D1012. doi:10.1093/nar/gky112030445434
- Hartwig FP, Davies NM, Hemani G, Davey Smith G. Two-sample Mendelian randomization: avoiding the downsides of a powerful, widely applicable but potentially fallible technique. Int J Epidemiol. 2016;45(6):1717–1726. doi:10.1093/ije/dyx02828338968
- Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.2175824114802
- Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x28527048
- Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–525. doi:10.1093/ije/dyv08026050253
- Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40(4):304–314. doi:10.1002/gepi.2016.40.issue-427061298
- Cole TJ. Sympercents: symmetric percentage differences on the 100 log(e) scale simplify the presentation of log transformed data. Stat Med. 2000;19(22):3109–3125. doi:10.1002/1097-0258(20001130)19:22<3109::AID-SIM558>3.0.CO;2-F11113946
- Egger M, Smith GD, Phillips AN. Meta-analysis: principles and procedures. BMJ. 1997;315(7121):1533–1537. doi:10.1136/bmj.315.7121.15339432252
- Greco MFD, Minelli C, Sheehan NA, Thompson JR. Detecting pleiotropy in Mendelian randomisation studies with summary data and a continuous outcome. Stat Med. 2015;34(21):2926–2940. doi:10.1002/sim.v34.2125950993
- Prins BP, Abbasi A, Wong A, et al. Investigating the causal relationship of C-reactive protein with 32 complex somatic and psychiatric outcomes: a large-scale cross-consortium Mendelian randomization study. PLoS Med. 2016;13(6):e1001976. doi:10.1371/journal.pmed.100197627327646
- Brion M-JA, Shakhbazov K, Visscher PM. Calculating statistical power in Mendelian randomization studies. Int J Epidemiol. 2013;42(5):1497–1501. doi:10.1093/ije/dyt17924159078
- Pierce BL, Ahsan H, Vanderweele TJ. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int J Epidemiol. 2011;40(3):740–752. doi:10.1093/ije/dyq15120813862
- Burgess S, Davies NM, Thompson SG. Bias due to participant overlap in two-sample Mendelian randomization. Genet Epidemiol. 2016;40(7):597–608. doi:10.1002/gepi.2199827625185
- Dastani Z, Hivert MF, Timpson N, et al. Novel loci for adiponectin levels and their influence on type 2 diabetes and metabolic traits: a multi-ethnic meta-analysis of 45,891 individuals. PLoS Genet. 2012;8(3):e1002607. doi:10.1371/journal.pgen.100260722479202
- Voight BF, Peloso GM, Orho-Melander M, et al. Plasma HDL cholesterol and risk of myocardial infarction: a mendelian randomisation study. Lancet. 2012;380(9841):572–580. doi:10.1016/S0140-6736(12)60312-222607825
- Fujimaki K, Toki S, Yamashita H, Oyamada T, Yamawaki S. Predictors of negative symptoms in the chronic phase of schizophrenia: a cross-sectional study. Psychiatry Res. 2018;262:600–608. doi:10.1016/j.psychres.2017.09.05128965809
- Gultekin BK, Kesebir S, Kabak SG, Ergun FF, Tatlidil Yaylaci E. Are uric acid levels different from healthy subjects in bipolar affective disorder and schizophrenia?: Relationship between clinical improvement and episode severity in male patients. Noro Psikiyatr Ars. 2014;51(3):229–232. doi:10.4274/npa.y682728360631
- Dadheech G, Sharma P, Gautam S. Oxidative stress-induced response of some endogenous antioxidants in schizophrenia. Indian J Clin Biochem. 2012;27(3):278–283. doi:10.1007/s12291-012-0193-z26405388
- Akanji AO, Ohaeri JU, Al-Shammri SA, Fatania HR. Associations of blood levels of insulin-like growth factor (IGF)-I, IGF-II and IGF binding protein (IGFBP)-3 in schizophrenic Arab subjects. Clin Chem Lab Med. 2007;45(9):1229–1231. doi:10.1515/CCLM.2007.26517635080
- Smith GD, Lawlor DA, Harbord R, Timpson N, Day I, Ebrahim S. Clustered environments and randomized genes: a fundamental distinction between conventional and genetic epidemiology. PLoS Med. 2007;4(12):e352. doi:10.1371/journal.pmed.004035218076282
- Salokangas RK, Honkonen T, Stengard E, Koivisto AM, Hietala J. Cigarette smoking in long-term schizophrenia. Eur Psychiatry. 2006;21(4):219–223. doi:10.1016/j.eurpsy.2005.07.00816360309
- Haj Mouhamed D, Ezzaher A, Neffati F, Douki W, Gaha L, Najjar MF. Effect of cigarette smoking on plasma uric acid concentrations. Environ Health Prev Med. 2011;16(5):307–312. doi:10.1007/s12199-010-0198-221431788
- Gee Teng G, Pan A, Yuan JM, Koh WP. Cigarette smoking and the risk of incident gout in a prospective cohort study. Arthritis Care Res (Hoboken). 2016;68(8):1135–1142. doi:10.1002/acr.2282126714165
- Keller JN, Kindy MS, Holtsberg FW, et al. Mitochondrial manganese superoxide dismutase prevents neural apoptosis and reduces ischemic brain injury: suppression of peroxynitrite production, lipid peroxidation, and mitochondrial dysfunction. J Neurosci. 1998;18(2):687–697. doi:10.1523/JNEUROSCI.18-02-00687.19989425011
- van der Veen RC, Hinton DR, Incardonna F, Hofman FM. Extensive peroxynitrite activity during progressive stages of central nervous system inflammation. J Neuroimmunol. 1997;77(1):1–7. doi:10.1016/S0165-5728(97)00013-19209262
- Davies KJ, Sevanian A, Muakkassah-Kelly SF, Hochstein P. Uric acid-iron ion complexes. A new aspect of the antioxidant functions of uric acid. Biochem J. 1986;235(3):747–754. doi:10.1042/bj23507473753442
- Xuan J, Pan G, Qiu Y, et al. Metabolomic profiling to identify potential serum biomarkers for schizophrenia and risperidone action. J Proteome Res. 2011;10(12):5433–5443. doi:10.1021/pr200679622007635