
Abstract
Background
Increasing evidence suggests that SALL4 plays oncogenic roles in cancer development and progression. We have previously shown that SALL4 is highly expressed in gastric cancer, and its upregulation is associated with lymph node metastasis and poor prognosis. The role of SALL4 in gastric cancer metastasis and the underlying mechanism remain unclear.
Materials and methods
The biological roles of SALL4 in gastric cancer cell mobility, migration, and invasion were investigated by wound healing, transwell migration assay, and Matrigel invasion assay. The effects of SALL4 on epithelial–mesenchymal transition (EMT) in gastric cancer cells were examined by quantitative real-time PCR and Western blot. The downstream target genes of SALL4 were identified by microarray. The regulation of TGF-β1 by SALL4 in gastric cancer cells was analyzed by luciferase reporter assay and chromatin immunoprecipitation assay.
Results
SALL4 knockdown inhibited, while SALL4 overexpression promoted the motility, migration, and invasion abilities of gastric cancer cells in vitro. SALL4 knockdown also suppressed the peritoneal metastasis of gastric cancer cells in nude mice. SALL4 knockdown suppressed, while SALL4 overexpression induced the activation of TGF-β/SMAD signaling pathway and triggered EMT in gastric cancer cells. TGF-β1 was identified as a direct target gene of SALL4. The results of chromatin immunoprecipitation study and luciferase reporter assay further confirmed that SALL4 bound to the promoter of TGF-b1 gene and activated its expression. TGF-β1 knockdown reversed SALL4-mediated promotion of gastric cancer cell motility, migration, and invasion, indicating that TGF-β1 acts as a downstream effector of SALL4. Furthermore, the expression of TGF-β1 was found to be closely associated with that of SALL4 in gastric cancer tissues.
Conclusion
SALL4 promotes the metastasis of gastric cancer, at least partly, by directly activating TGF-β1, suggesting that SALL4 may serve as a new target for gastric cancer therapy.
Keywords:
Introduction
Gastric cancer is the fourth most common malignancy and the third leading cause of cancer-related deaths worldwide.Citation1 Most patients are diagnosed at a late stage with lymph node or distant metastases or with relapse after prior curative surgical therapy, which makes the 5-year survival rate relatively low.Citation2 Peritoneal metastasis is the most frequent metastatic and recurrent site, which has a very poor prognosis. The identification of molecules that play important roles in metastasis and clarification of their mechanisms will provide novel diagnostic markers and therapeutic targets for gastric cancer.
SALL4 is a zinc finger transcription factor that maintains the self-renewal and pluripotency of embryonic stem cells.Citation3 SALL4 expression gradually decreases during development and is even absent in most adult tissues.Citation4 However, the recent findings show that SALL4 is reexpressed in cancer. The increased expression of SALL4 has been reported in acute myeloid leukemia (AML),Citation5,Citation6 liver cancer,Citation7,Citation8 colon cancer,Citation9 breast cancer,Citation10 endometrial cancer,Citation11 lung cancer,Citation12,Citation13 and glioma.Citation14 Further studies reveal that SALL4 plays important roles in cancer development, progression, and metastasis. For instance, SALL4 is found to be constitutively expressed in AML, in which it inhibits cell apoptosis via interaction with β-catenin.Citation6 In liver cancer, SALL4 inhibits PTEN expression by recruiting nucleosome remodeling and deacetylase complex and activates Akt signaling pathway to promote cell proliferation.Citation15,Citation16 In endometrial cancer, SALL4 promotes cell proliferation, migration, invasion, and drug resistance via the upregulation of c-Myc.Citation11 The potent roles of SALL4 in multiple processes of carcinogenesis have made it a novel biomarker for cancer diagnosis and treatment.Citation17,Citation18
We have previously reported the elevated expression of SALL4 in human gastric cancer.Citation19 SALL4 upregulation was associated with lymph node metastasis and predicted poor prognosis in gastric cancer patients. The functional roles of SALL4 in gastric cancer metastasis and the underlying molecular mechanisms remain unclear. In this study, we reported that SALL4 overexpression promoted, while SALL4 knockdown inhibited the metastatic potential of gastric cancer cells in vitro and in vivo. SALL4 induced epithelial–mesenchymal transition (EMT) in gastric cancer cells through the upregulation of TGF-β1 expression and the subsequent activation of TGF-β/SMAD signaling pathway. TGF-β1 knockdown abrogated the promoting role of SALL4 in gastric cancer metastasis. Our findings indicate that SALL4 promotes gastric cancer metastasis via direct regulation of TGF-β1, a strong EMT inducer, which represents a new mechanism for the oncogenic roles of SALL4 in cancer.
Materials and methods
Clinical specimens
A total of 19 paired gastric cancer and adjacent noncancerous tissues (5 cm away from the tumor edge) were obtained from the Department of General Surgery, the Affiliated People’s Hospital of Jiangsu University, between April 2015 and September 2016. Written informed consent was obtained from all the patients, and this study was approved by the Institutional Ethical Committee of Jiangsu University. All of the tissues were frozen in liquid nitrogen and then stored at −80°C for further use. The patients included in this study had not received any preoperative therapies.
Cell culture
Human gastric cancer cell lines MGC-803 and HGC-27 were purchased from the Institutes for Biological Sciences at the Chinese Academy of Sciences (Shanghai, China) and cultured in high-glucose Dulbecco’s modified Eagle’s medium, supplemented with 10% fetal bovine serum (FBS; Gibco®, Invitrogen Life Technologies, Carlsbad, CA, USA). All the cells were cultured in a humidified incubator with 5% CO2 at 37°C. For TGF-β1 inhibitor experiment, the cells were cultured in the presence or absence of Disitertide (HY-P0118, 50 µg/mL; MedChemExpress, Shanghai, China).
Gene transfection
The cells were seeded in 6-well plates at a density of 2×105/well and cultured in 37°C incubator overnight. The over-expressing plasmid and silencing siRNAs (GeneChem, Shanghai, China) were transfected into the cells by using LipoFiter™ transfection reagent (Hanbio, Shanghai, China) in a serum-free medium. The cells were changed to a complete medium at 6 hours after transfection and cultured for another 30 hours. The SALL4-targeting shRNA lentivirus was provided by GeneChem. The cells were transfected with lentivirus at a multiplicity of infection of 100 for 24 hours and then selected with puromycin (0.8 µg/mL) for 3 days. lists the target sequences of shRNAs and siRNAs.
Luciferase reporter assay
In total, 293 T cells were cotransfected with SALL4-overex-pressing plasmid and the luciferase reporter vector containing the promoter region of TGF-β1 as indicated. At 36 hours after transfection, the cells were collected and lysed. The luciferase activity was detected by using the dual luciferase assay kit (Promega, Madison, WI, USA).
Microarray analysis
RNA samples from control and SALL4-targeting shRNA-transfected MGC-803 cells (three samples/group) were collected and used for gene expression analysis by using Agilent Human lncRNA microarray v2.0 4 × 180K (OE Biotech, Shanghai, China).
Chromatin immunoprecipitation assay
The chromatin immunoprecipitation assay was performed in MGC-803 cells by using a commercial kit (Millipore, Darmstadt, Germany). After cross-linking with 1% formaldehyde at 37°C for 10 minutes, the cells were harvested in sodium dodecyl sulfate lysis buffer, and the DNA was shredded to fragments of 200 bp by sonication. The precleared chromatin was incubated with the antibodies against SALL4 or nonspecific IgG overnight. Protein G Agarose beads were added and incubated at 4°C for 1 hour. After reversing the cross-links, the DNA was isolated and used for PCR. shows the specific primers for PCR detection of the responsive element in TGF-b1 gene promoter.
Cell scratch assay
The confluent cell monolayers were wounded by scratching with a 10-µL pipette tip and then cultured for 36 hours. Cell migration over the scraped area was photographed at 0 and 36 hours, respectively.
Transwell migration assay
The transfected cells were collected and seeded into the upper chamber (8 µm) at a density of 1×105 cells/well (Corning Inc., Corning, NY, USA). The lower chamber was filled with 500 µL culture medium supplemented with 10% FBS; 12 hours later, the cells on the upper surface of the membrane were removed with a cotton swab. Then, the lower cells were fixed with formaldehyde and stained with crystal violet for 30 minutes. The number of migrated cells was counted under a microscope.
Matrigel invasion assay
The diluted basement Matrigel (BD Biosciences, Franklin Lakes, NJ, USA) was added into each chamber and let to polymerize at 37°C for 30 minutes. The transfected cells were seeded into the upper chamber at a density of 2×105 cells/well. The lower chamber was filled with 500 µL culture medium supplemented with 10% FBS. The cells were allowed to invade to the lower membrane for 24 hours. Subsequently, the cells on the upper surface of the membrane were removed with a cotton swab. The lower cells were then fixed with formaldehyde and stained with crystal violet for 30 minutes. The number of migrated cells was counted under a microscope.
Quantitative real-time (qRT) PCR
Total RNA was extracted using TRIzol reagent (Invitrogen Life Technologies) and reverse-transcribed into cDNA using miScript reverse transcription kit (Bio-Rad Laboratories Inc., Hercules, CA, USA). The relative expression of target genes was detected on a Bio-Rad CFX96 qRT-PCR system with the SYBR Green method (β-actin served as an internal control). lists the sequences of the primers.
Western blot analysis
The cells were washed twice with PBS and lysed with radioimmunoprecipitation assay buffer containing 1% protease inhibitors. Equal amounts of proteins were separated on 12% sodium dodecyl sulfate–polyacrylamide gels and transferred onto polyvinylidene fluoride membranes, followed by blocking with 5% nonfat milk for 1 hour. The membranes were incubated with primary antibodies overnight at 4°C. The following primary antibodies were used: anti-E-cadherin (4695S; Cell Signaling Technology, Beverly, MA, USA), anti-N-cadherin (4370S; Cell Signaling Technology), anti-Slug (9585S; Cell Signaling Technology), anti-Vimentin (5741S; Cell Signaling Technology), anti-Twist (46702S; Cell Signaling Technology), anti-p-SMAD2 (3108S; Cell Signaling Technology), SMAD2 (5399S; Cell Signaling Technology), anti-p-SMAD3 (9520S; Cell Signaling Technology), SMAD3 (9523S; Cell Signaling Technology), and anti-GAPDH (MB001; Bioworld Technology, St. Louis Park, MN, USA). After incubation with the secondary antibodies (Bioworld Technology) at 37°C for 1 hour, the bands were visualized with a chemiluminescent detection system.
Animal study
BALB/c nude mice aged 4–6 weeks were purchased from the Shanghai Laboratory Animal Center (Shanghai, China) and maintained in accordance with the institutional policies. Control or sh-SALL4 MGC-803 cells were collected in PBS and intraperitoneally injected into the mice (2×106 cells/mice, n=5). At 6 weeks after injection, the mice were sacrificed, and the number of metastatic nodules was counted. The protocol was approved by the Animal Use and Care Committee of Jiangsu University.
Immunofluorescence
For immunofluorescent staining, the cells seeded on cover slips were fixed and incubated with primary monoclonal antibody against N-cadherin and p-SMAD3 (Cell Signaling Technology) followed by incubation with fluorescence-labeled secondary antibody for 30 minutes at room temperature. The cells were counterstained with Hoechst33342 for 30 seconds. Finally, the cells were photographed under a microscope (DeltaVision OMX SR; GE Healthcare BioSciences, Piscataway, NJ, USA).
Statistical analysis
All the results were expressed as mean ± SD. Statistical analyses were performed using Student’s t-test with GraphPad Prism Version 5.0 software (GraphPad Software, La Jolla, CA, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
SALL4 gene silencing inhibits, while gene overexpression promotes the migration and invasion of gastric cancer cells
To test the importance of SALL4 in gastric cancer metastasis, we knocked down SALL4 gene expression by lentivirus-mediated shRNA in MGC-803 cells (with a relatively high level of SALL4). The efficiency of gene silencing by SALL4 shRNA was verified by qRT-PCR and Western blot (). Cell scratch assay results showed that SALL4 knockdown reduced the motility of MGC-803 cells (). Transwell migration and Matrigel invasion assay results showed that MGC-803 cells in SALL4 knockdown group had lower migration rate and invasion abilities than that in the control group (). On the contrary, SALL4 overexpression by plasmid transfection promoted the migration and invasion abilities of HGC-27 cells (with a relatively low level of SALL4; ). These data suggest that SALL4 knockdown inhibits, while overexpression promotes the migration and invasion of gastric cancer cells.
Figure 1 SALL4 knockdown inhibits, while overexpression promotes the migration and invasion of gastric cancer cells.
Notes: (A) The efficiency of SALL4 knockdown was verified by using qRT-PCR and Western blot. (B) Cell scratch assay for the motility of MGC-803 cells with SALL4 gene knockdown. (C) Transwell migration and Matrigel invasion assays for the migration and invasion abilities of MGC-803 cells with SALL4 gene knockdown. (D) The efficiency of SALL4 overexpression was verified by using qRT-PCR and Western blot. (E) Cell scratch assay for the motility of SALL4-overexpressing HGC-27 cells. (F) Transwell migration and Matrigel invasion assays for the migration and invasion abilities of SALL4-overexpressing HGC-27 cells. **P<0.01.
Abbreviation: qRT-PCR, quantitative real-time PCR.
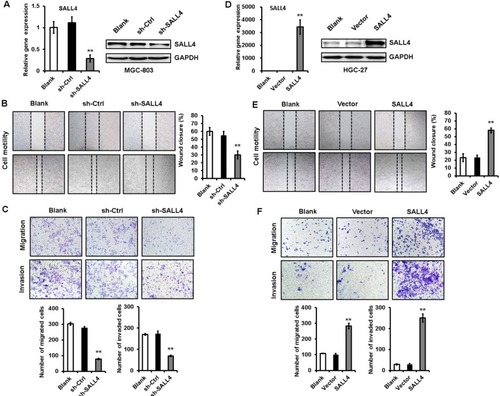
SALL4 gene silencing suppresses, while gene overexpression promotes EMT in gastric cancer cells
EMT has been linked to enhanced metastatic potential in gastric cancer cells. To test whether SALL4 promotes gastric cancer cell migration and invasion by inducing EMT, we observed the morphology changes of SALL4-overexpressing and SALL4-silencing gastric cancer cells and detected the expression of EMT-related genes and proteins in these cells. As shown in , MGC-803 cells in the control group displayed a mesenchymal-like phenotype, while those in SALL4 knockdown group displayed an epithelial-like phenotype. On the contrary, HGC-27 cells in the control group displayed an epithelial-like phenotype, while those in SALL4 overexpression group displayed a mesenchymal-like phenotype. In addition, SALL4 knockdown increased the expression level of epithelial marker E-cadherin and decreased that of mesenchymal markers N-cadherin and Vimentin in MGC-803 cells (). SALL4 knockdown also reduced the expression levels of EMT transcription factors Slug and Twist. However, SALL4 overexpression led to the decreased level of E-cadherin and increased levels of N-cadherin, Vimentin, Slug, and Twist in HGC-27 cells (). The results of fluorescent staining further confirmed a decreased expression of N-cadherin in the SALL4 knockdown group while an increased expression of N-cadherin in the SALL4 overexpression group compared with the control group (). These findings indicate that SALL4 knockdown suppresses, while overexpression induces EMT in gastric cancer cells.
Figure 2 SALL4 knockdown suppresses, while overexpression induces EMT in gastric cancer cells.
Notes: (A) The morphology changes in SALL4-depleted MGC-803 cells and SALL4-overexpressing HGC-27 cells. (B, C) The expression of E-cadherin, N-cadherin, Vimentin, Slug, and Twist in SALL4-depleted MGC-803 cells was determined by using qRT-PCR (B) and Western blot (C). (D, E) qRT-PCR (D) and Western blot (E) assays for the expression of E-cadherin, N-cadherin, Vimentin, Slug, and Twist in SALL4-overexpressing HGC-27 cells. (F) Immunofluorescent staining of N-cadherin in SALL4-depleted MGC-803 cells and SALL4-overexpressing HGC-27 cells. *P<0.05, **P<0.01, ***P<0.001.
Abbreviations: EMT, epithelial–mesenchymal transition; qRT-PCR, quantitative real-time PCR.
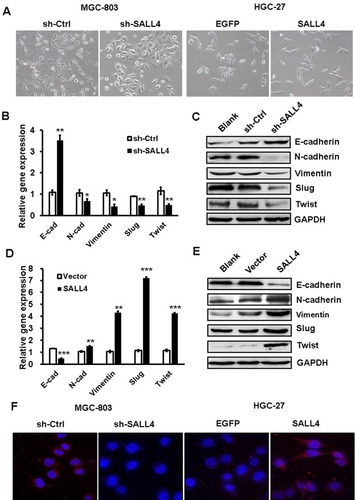
TGF-β1 is identified as a downstream target of SALL4
To demonstrate the mechanism for SALL4-mediated induction of EMT and the promotion of gastric cancer cell migration and invasion, we performed microarray to determine the differentially expressed genes between control and SALL4 knockdown MGC-803 cells. As shown in , 1,539 differentially expressed genes were identified, of which 1,001 genes were upregulated and 538 genes were downregulated. Gene ontology and KEGG pathway analysis results showed that SALL4 was associated with cell adhesion molecules, calcium signaling pathway, and glycolysis (data not shown). In the downregulated genes, we choose TGF-β1 as the potential target for further study as it is a well-known EMT inducer. In consistent with the results of microarray, qRT-PCR and Western blot results confirmed that TGF-β1 expression was decreased in SALL4 knockdown MGC-803 cells (). We confirmed the regulation of TGF-β1 by SALL4 using luciferase reporter and chromatin immunoprecipitation (ChIP) assays. Luciferase reporter assay results showed that SALL4 overexpression increased the luciferase activities of TGF-b1 gene promoter (). ChIP assay results showed that SALL4 could bind to the 540- to −301-bp region of the promoter of TGF-b1 gene (). Finally, we determined the expression of SALL4 and TGF-β1 genes in gastric cancer tissues and found that the expression levels of SALL4 and TGF-β1were closely associated (). Our results suggest that SALL4 may bind to TGF-b1 gene promoter and transactivate its expression.
Figure 3 TGF-β1 is identified as a downstream target of SALL4.
Notes: (A) The differentially expressed genes between sh-Ctrl- and sh-SALL4-transfected MGC-803 cells were determined by using microarray analyses. The relative fluorescence intensity for TGF-β1 gene was shown. (B) qRT-PCR and Western blot assays for the expression of TGF-β1 gene in MGC-803 cells with SALL4 gene knockdown. (C) Luciferase reporter assays for the relative luciferase activity of TGF-β1 gene promoter. (D) ChIP assay for the binding of SALL4 to the promoter region of TGF-β1 gene. (E) Correlation analysis of SALL4 and TGF-β1 expression in 19 paired gastric cancer and adjacent noncancerous tissues. *P<0.05, **P<0.01, ***P<0.001.
Abbreviations: ChIP, Cardiovascular Health Improvement Project; qRT-PCR, quantitative real-time PCR.
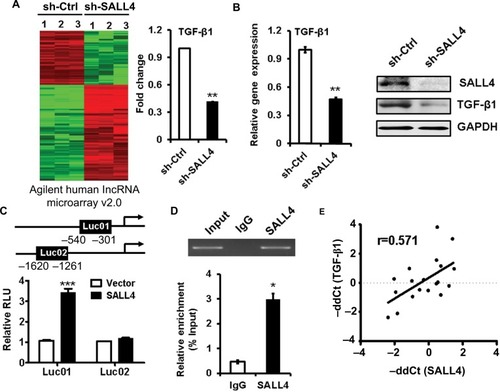
SALL4 gene silencing suppresses, while gene overexpression promotes the activation of TGF-β/SMAD signaling pathway
TGF-β induces EMT mainly through SMAD signaling pathway. We next wanted to know the effects of SALL4 on SMAD signaling pathway. qRT-PCR and Western blot results showed that SALL4 knockdown inhibited the expression of TGF-β1 and the phosphorylation of SAMD2 and SMAD3 in MGC-803 cells (). By contrast, SALL4 overexpression enhanced the expression of TGF-β1 and the phosphorylation of SAMD2 and SMAD3 in HGC-27 cells (). The results of immunofluorescent staining further confirmed the decreased nuclear translocation of p-SMAD3 in SALL4-silencing MGC-803 cells while the increased nuclear translocation of p-SMAD3 in SALL4-overexpressing HGC-27 cells compared with control cells (). These results suggest that SALL4 could control the activation of TGF-β/SMAD signaling pathway.
Figure 4 SALL4 regulates the activation of TGF-β/SMAD signaling pathway.
Notes: (A) qRT-PCR analyses of SALL4 and TGF-β1 expression in SALL4-depleted MGC-803 cells. (B) Western blot assays for the expression of phosphorylated SMA2/3 in SALL4-depleted MGC-803 cells. (C) qRT-PCR analyses of SALL4 and TGF-β1 expression in SALL4-overexpressing HGC-27 cells. (D) Western blot assays for the expression of phosphorylated SMA2/3 in SALL4-overexpressing HGC-27 cells. (E) Immunofluorescent staining of p-SMAD3 in SALL4-depleted MGC-803 cells and SALL4-overexpressing HGC-27 cells. *P<0.05, **P<0.01, ***P<0.001.
Abbreviation: qRT-PCR, quantitative real-time PCR.
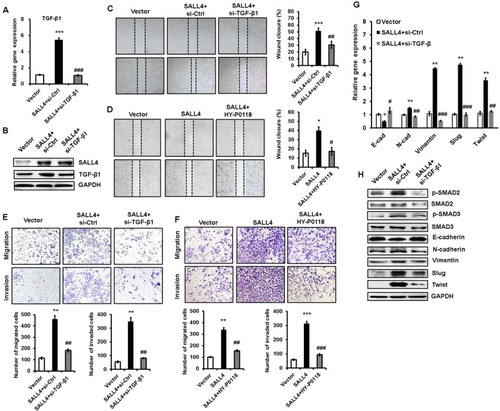
TGF-β1 inhibition abrogates the promotion of gastric cancer cell migration and invasion by SALL4 overexpression
To demonstrate the importance of TGF-β1 in the oncogenic roles of SALL4 in gastric cancer, we depleted TGF-β1 expression in SALL4-overexpressing HGC-27 cells by using siRNA. The efficiency of TGF-β1 gene knockdown was shown in . Compared with SALL4-overexpressing group, TGF-β1 gene knockdown group displayed reduced abilities of cell motility, migration, and invasion (). In addition, we used a TGF-β1 inhibitor (HY-P0118) to block the function of TGF-β1 in SALL4-overexpressing HGC-27 cells. The results of wound healing, transwell migration, and Matrigel invasion assays showed that HY-P0118 treatment remarkably reversed the cell motility, migration, and invasion abilities of SALL4-over-expressing HGC-27 cells (). We also found that the induced EMT phenotype and activation of TGF-β/SMAD signaling pathway in gastric cancer cells by SALL4 overexpression were reversed by TGF-β1 gene knockdown (), indicating that SALL4 promotes gastric cancer cell migration and invasion and induces EMT through the upregulation of TGF-β1.
Figure 5 TGF-β1 mediates the promoting role of SALL4 in gastric cancer cell migration and invasion.
Notes: (A, B) qRT-PCR (A) and Western blot (B) analyses of TGF-β1 expression in SALL4-overexpressing HGC-27 cells with or without TGF-β1 knockdown. (C, D) Cell scratch assay for the motility of SALL4-overexpressing HGC-27 cells with or without TGF-β1 knockdown (C) or HY-P0118 treatment (D). (E, F) Transwell migration and Matrigel invasion assays for the migration and invasion abilities of SALL4-overexpressing HGC-27 cells with or without TGF-β1 knockdown (E) or HY-P0118 treatment (F). (G, H) qRT-PCR (G) and Western blot (H) analyses of EMT markers and SMAD2/3 in SALL4-overexpressing HGC-27 cells with or without TGF-β1 knockdown. *P<0.05, **P<0.01, ***P<0.001, compared to vector group; #P<0.05, ##P<0.01, ###P<0.001, compared to SALL4 group.
Abbreviations: EMT, epithelial–mesenchymal transition; qRT-PCR, quantitative real-time PCR.
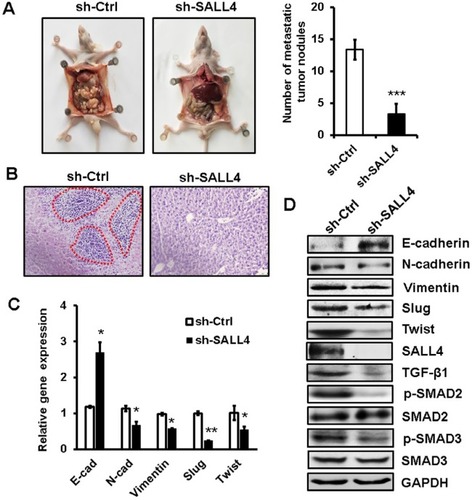
SALL4 knockdown inhibits the peritoneal metastasis of gastric cancer in vivo
To further demonstrate the role of SALL4 in gastric cancer metastasis, we established a peritoneal metastasis model in nude mice. MGC-803 cells with or without SALL4 gene knockdown were injected into the abdominal cavity of nude mice. At 6 weeks after injection, the mice were sacrificed, and the metastatic tumor nodes in the abdomen were carefully examined. As shown in , SALL4 knockdown reduced the number of metastatic tumor nodes in mice. Hematoxylin and eosin staining results showed that there were more hepatic metastases in mice from the control group than that from the SALL4 knockdown group (). qRT-PCR and Western blot results showed that the expression of TGF-β1, p-SMAD2/3, N-cadherin, Vimentin, Slug, and Twist was decreased, while that of E-cadherin was increased in the SALL4 knockdown group (). These data suggest that SALL4 promotes gastric cancer metastasis via the activation of TGF-β/SMAD signaling pathway and induction of EMT.
Figure 6 SALL4 knockdown inhibits the peritoneal metastasis of gastric cancer in vivo.
Notes: (A) The number of metastatic tumor nodes in the abdomen of mice from the control and SALL4 knockdown groups. (B) H&E staining of liver tissues in mice from the control and SALL4 knockdown groups. (C) qRT-PCR analyses of EMT markers in mouse metastatic tumor nodules from the control and SALL4 knockdown groups. (D) Western blot assays for EMT marker and SMAD2/3 expression in mouse metastatic tumor nodules from the control and SALL4 knockdown groups. *P<0.05, **P<0.01, ***P<0.001.
Abbreviations: EMT, epithelial–mesenchymal transition; H&E, hematoxylin and eosin; qRT-PCR, quantitative real-time PCR.
Discussion
In this study, we demonstrate that SALL4 plays a critical role in gastric cancer metastasis. SALL4 overexpression promoted, while SALL4 knockdown inhibited the metastatic potential of gastric cancer cells in vitro and in vivo. Mechanistically, SALL4 bound to the promoter region of TGF-b1 gene and activated its expression, resulting in the activation of TGF-β/SMAD signaling pathway and the induction of EMT. We further provided evidence that TGF-β1 knockdown reversed SALL4 overexpression-mediated promotion of gastric cancer cell migration and invasion, suggesting that TGF-β1 acts as a downstream mediator of SALL4’s function in gastric cancer metastasis. Although the involvement of SALL4 in regulating TGF-β pathway has been implicated in AML,Citation6 to the best of our knowledge, this is the first report to suggest TGF-β1 as a direct target of SALL4 and to reveal the functional importance of TGF-β1 regulation to the oncogenic roles of SALL4 in cancer.
EMT is a biological process in which cancer cells lose their epithelial features and gain mesenchymal markers, which enables tumor cells to become more migratory and invasive.Citation20 The hallmark of EMT is the decrease in E-cadherin expression, accompanied by an increase in N-cadherin or Vimentin expression. The process of EMT is controlled by transcription factors such as Slug, Snail, ZEB1, and Twist, as well as certain signaling pathways, including TGF-β/SMAD and Wnt/β-catenin. Therefore, a further exploration of the regulatory mechanism for EMT in cancer metastasis has a great significance. He et al demonstrated that the inhibition of SALL4 reduces tumorigenicity involving EMT via Wnt/β-catenin pathway in esophageal squamous cell carcinoma.Citation21 Liu et al suggested that SALL4 acts as an EMT and drug resistance inducer through the regulation of c-Myc in endometrial cancer.Citation22 Itou et al showed that SALL4 suppresses the expression of E-cadherin while it increases the expression of ZEB1, maintaining cell dispersion in basal-like breast cancer.Citation23 They further demonstrated that SALL4 upregulates integrin α6β1 expression to promote cell migration via the activation of focal adhesion dynamics in basal-like breast cancer cells.Citation24 In this study, we showed that SALL4 overexpression increased TGF-β1 expression and activated TGF-β/SMAD signaling, suggesting that SALL4 acts as a potent EMT inducer via the regulation of distinct gene expression and signaling pathway activation.
SALL4 is involved in tumorigenesis, tumor growth, and tumor metastasis and drug resistance through the regulation of various downstream genes. In AML, Bmi-1 is identified as a target gene of SALL4. SALL4 overexpression induces increased levels of histones H3-K4 and H3-K79 methylation in the Bmi-1 gene promoter, which finally activates Bmi-1 expression.Citation25 There is a strong correlation between SALL4 and Bmi-1 expression in human AML samples. SALL4 has been shown to interact with mixed-lineage leukemia (MLL) and co-occupies the HOXA9 promoter region with MLL in AML cells, leading to enhanced histone activation markers at the HOXA9 promoter region as well as increased HOXA9 expression.Citation26 Li et al suggested that c-Myc is a key target gene of SALL4 in endometrial cancer, which could partially mediate the roles of SALL4 in metastasis and drug resistance.Citation11 Kim et al demonstrated that SALL4 recruits ubiquitin E3 ligase CUL4B to destabilize heterochromatin protein 1α, promoting Glut1 expression and glycolysis and inducing drug resistance by enhancing DNA repair.Citation27 We recently reported that the expression of CD44 and DANCR was controlled by SALL4 in gastric cancer cells to regulate their proliferation, migration, and invasion.Citation28,Citation29 We found in this study that the expression of SALL4 and TGF-β1 was positively correlated in gastric cancer tissues. The elevated expression of TGF-β1 has been linked with a worse overall survival in gastric cancer patients.Citation30 The outcomes of gastric cancer patients with simultaneously high levels of SALL4 and TGF-β1 warrant a further study. Moreover, whether other cofactors are involved in the regulation of TGF-β1 by SALL4 and the epigenetic changes in SALL4 binding site in the TGF-b1 gene promoter need to be investigated in future studies.
Conclusion
In conclusion, our findings show that SALL4 promotes gastric cancer metastasis via the upregulation of TGF-β1 and the activation of TGF-β/SMAD signaling pathway, inducing EMT and enhancing migration and invasion abilities in gastric cancer cells. Our study not only reveals a new mechanism for the oncogenic roles of SALL4 in cancer, but also provides evidence for the potential of SALL4 as a diagnostic and therapeutic target for gastric cancer.
Acknowledgments
The present study was supported by the National Natural Science Foundation of China (grant numbers: 81672416 and 81572075), the Key Research and Development Project of Zhenjiang (grant number: SH2015034), the Major Research and Development Project of Jiangsu Province (grant number: BE2015667), the Starting Foundation for Senior Talents of Jiangsu University (grant number: 13JDG086), the 333 Project of Jiangsu Province, and the Foundation for Young Academic Leader of Jiangsu University.
Supplementary materials
Table S1 Sequences of shRNA and siRNA
Table S2 Sequences of PCR primers for target gene detection
Disclosure
The authors report no conflicts of interest in this work.
References
- TorreLABrayFSiegelRLFerlayJLortet-TieulentJJemalAGlobal cancer statistics, 2012CA Cancer J Clin20156528710825651787
- Van CutsemESagaertXTopalBHaustermansKPrenenHGastric cancerLancet2016388100602654266427156933
- ZhangJTamWLTongGQSall4 modulates embryonic stem cell pluripotency and early embryonic development by the transcriptional regulation of Pou5f1Nat Cell Biol20068101114112316980957
- OikawaTKamiyaAKakinumaSSall4 regulates cell fate decision in fetal hepatic stem/progenitor cellsGastroenterology200913631000101119185577
- MaYCuiWYangJSALL4, a novel oncogene, is constitutively expressed in human acute myeloid leukemia (AML) and induces AML in transgenic miceBlood200610882726273516763212
- YangJChaiLGaoCSALL4 is a key regulator of survival and apoptosis in human leukemic cellsBlood2008112380581318487508
- OikawaTKamiyaAZeniyaMSal-like protein 4 (SALL4), a stem cell biomarker in liver cancersHepatology20135741469148323175232
- ZengSSYamashitaTKondoMThe transcription factor SALL4 regulates stemness of EpCAM-positive hepatocellular carcinomaJ Hepatol201460112713424012616
- ChengJDengRWuCInhibition of SALL4 suppresses carcinogenesis of colorectal cancer via regulating Gli1 expressionInt J Clin Exp Pathol201589100921010126617716
- KobayashiDKuribayshiKTanakaMWatanabeNSALL4 is essential for cancer cell proliferation and is overexpressed at early clinical stages in breast cancerInt J Oncol201138493393921274508
- LiAJiaoYYongKJSALL4 is a new target in endometrial cancerOncogene2015341637224336327
- KobayashiDKuribayashiKTanakaMWatanabeNOverexpression of SALL4 in lung cancer and its importance in cell proliferationOncol Rep201126496597021725617
- YanagiharaNKobayashiDKuribayashiKTanakaMHasegawaTWatanabeNSignificance of SALL4 as a drugresistant factor in lung cancerInt J Oncol20154641527153425646965
- ZhangLYanYJiangYThe expression of SALL4 in patients with gliomas: high level of SALL4 expression is correlated with poor outcomeJ Neurooncol2015121226126825359397
- YongKJGaoCLimJSOncofetal gene SALL4 in aggressive hepatocellular carcinomaN Engl J Med2013368242266227623758232
- LuJJeongHWJeongHStem cell factor SALL4 represses the transcriptions of PTEN and SALL1 through an epigenetic repressor complexPLoS One200945e557719440552
- ZhangXYuanXZhuWQianHXuWSALL4: an emerging cancer biomarker and targetCancer Lett20153571556225444934
- GaoCDimitrovTYongKJTargeting transcription factor SALL4 in acute myeloid leukemia by interrupting its interaction with an epigenetic complexBlood201312181413142123287862
- ZhangLXuZXuXSALL4, a novel marker for human gastric carcinogenesis and metastasisOncogene201433485491550024276240
- ThieryJPAcloqueHHuangRYNietoMAEpithelial-mesenchymal transitions in development and diseaseCell2009139587189019945376
- HeJZhouMChenXInhibition of SALL4 reduces tumorigenicity involving epithelial-mesenchymal transition via Wnt/β-catenin pathway in esophageal squamous cell carcinomaJ Exp Clin Cancer Res20163519827329034
- LiuLZhangJYangXFangCXuHXiXSALL4 as an epithelial-mesenchymal transition and drug resistance inducer through the regulation of c-Myc in endometrial cancerPLoS One2015109e013851526407074
- ItouJMatsumotoYYoshikawaKToiMSal-like 4 (SALL4) suppresses CDH1 expression and maintains cell dispersion in basal-like breast cancerFEBS Lett2013587183115312123954296
- ItouJTanakaSLiWThe Sal-like 4 – integrin α6β1 network promotes cell migration for metastasis via activation of focal adhesion dynamics in basal-like breast cancer cellsBiochim Biophys Acta2017186417688
- YangJChaiLLiuFBmi-1 is a target gene for SALL4 in hema-topoietic and leukemic cellsProc Natl Acad Sci U S A200710425104941049917557835
- LiAYangYGaoCA SALL4/MLL/HOXA9 pathway in murine and human myeloid leukemogenesisJ Clin Invest2013123104195420724051379
- KimJXuSXiongLYuLFuXXuYSALL4 promotes glycolysis and chromatin remodeling via modulating HP1α-Glut1 pathwayOncogene201736466472647928759035
- YuanXZhangXZhangWSALL4 promotes gastric cancer progression through activating CD44 expressionOncogenesis2016511e26827819668
- PanLLiangWGuJLong noncoding RNA DANCR is activated by SALL4 and promotes the proliferation and invasion of gastric cancer cellsOncotarget2018921915193029416741
- HawinkelsLJVerspagetHWvan DuijnWTissue level, activation and cellular localisation of TGF-beta1 and association with survival in gastric cancer patientsBr J Cancer200797339840417637685