
Abstract
Objectives
The objectives of this study were to explore the mechanisms of metformin sensitization to hypoxia-induced gefitinib treatment in resistant head and neck squamous cell carcinoma (HNSCC) and evaluate the effects of this combined treatment strategy.
Methods
The effects of gefitinib treatment on HNSCC were measured under normoxic and hypoxic conditions. The relationship between hypoxia and cell cycle and epithelial–mesenchymal transition (EMT) in tumor cells were analyzed. Palbociclib and LY294002 were used in combination with gefitinib to evaluate the effects on HNSCC cell cytotoxicity during hypoxia. Finally, metformin was used to evaluate the sensitizing effects of gefitinib treatment on HNSCC in vivo and in vitro.
Results
Cell viability and apoptosis assays demonstrated a significant difference in HNSCC cells treated with gefitinib between the normoxia and hypoxia groups. Hypoxia induced the expression of cyclin D1, decreased the percentage of cells in G1, and promoted the EMT of tumor cells. Both palbociclib and LY294002 enhanced gefitinib-induced cytotoxicity of HNSCC cells under hypoxic conditions. Encouragingly, metformin sensitized HNSCC to gefitinib treatment in vivo and in vitro.
Conclusion
Hypoxia promotes G1–S cell cycle progression and EMT in HNSCC, resulting in gefitinib treatment resistance. Metformin sensitizes HNSCC to gefitinib treatment, which might serve as a novel combined treatment strategy.
Introduction
Head and neck squamous cell carcinoma (HNSCC) ranks as the sixth most common cancer worldwide.Citation1 The main treatment strategy for this cancer includes surgery, chemotherapy, and radiotherapy; however, the 5-year survival rate still remains low, ~50%.Citation2,Citation3 Recently, new therapeutic approaches, such as molecular-targeted therapy, have raised more clinical concern. Until now, molecular targeting strategies included treatments targeting EGFR, vascular endothelial growth factor, and vascular endothelial growth factor receptor, as well as intracellular signaling pathway components associated with the phosphatidylinositol 3-kinase (PI3K)/serine/threonine-specific protein kinase (AKT).Citation4–Citation6 Among the abovementioned treatments, EGFR-targeted therapy has been proven to have great clinical value among various cancers, especially lung cancer.Citation7 Nearly 90% of the HNSCC cases are confirmed to have a high expression of EGFR.Citation8 Theoretically, targeting this molecule should result in significant clinical outcomes and improve the prognosis of HNSCC. Unfortunately, this type of therapy has been shown to have limited benefits.Citation9,Citation10 EGFR tyrosine kinase inhibitors (TKIs), including the first-generation TKI – gefitinib, have not been approved by the Food and Drug Administration (FDA) for use in HNSCC. Based on the results of reported clinical trials, HNSCC cells might apply various mechanisms to attain resistance to gefitinib treatment, which greatly reduces treatment efficacy.Citation11
Currently, the molecular mechanisms underlying gefitinib treatment resistance in HNSCC remain unclear. It was reported that acquired resistance is mainly due to a secondary mutation (threonine-to-methionine substitution at amino acid position 790 [T790M]) and amplification of other oncogenes.Citation12–Citation14 Recently, more and more studies have revealed the importance of the tumor microenvironment in promoting tumor progression and treatment resistance.Citation15,Citation16 Hypoxia is a typical feature in the microenvironment and has been demonstrated by several studies to play a pivotal role in acquired gefitinib resistance.Citation17,Citation18 Our previous work showed that chronic inflammation and hypoxic signaling are involved in crosstalk in the HNSCC tumor microenvironment, consequently decreasing the effect of cisplatin treatment.Citation19 Thus, we hypothesize that hypoxia, a dominant tumor microenvironment feature in HNSCC, might have the same effect on gefitinib resistance. It is therefore imperative to explore the mechanisms and overcome hypoxia-induced treatment resistance. There is an urgent need for clinicians to develop an easy and practical way to re-sensitize HNSCC to the effects of gefitinib.
Metformin is a widely used drug for diabetes, and recent reports have confirmed its effects in combating cancer and improving patient survival rates.Citation20 Tseng reported that metformin use is associated with a significantly lower risk of nasopharyngeal cancerCitation21 and Woo et al showed that metformin inhibits the expression of malic enzyme and sensitizes HNSCC cells to therapy-induced senescence.Citation22 Our group also proved that metformin may synergize the effects of cisplatin and hypoxia via the inhibition of hypoxic inducible factor-1α (HIF-1α) expression.Citation23 Hence, in this study, we explored the mechanisms of hypoxia-induced gefitinib resistance and found that this might be explained by the promotion of G1–S cell cycle progression and epithelial–mesenchymal transition (EMT) in tumor cells. We also provide strong evidence for the combined therapy of metformin and gefitinib in overcoming hypoxia-induced gefitinib resistance in HNSCC.
Materials and methods
Cell culture
The human HNSCC cell lines CAL27, HSC3, and SCC4 were obtained from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China); JHU011 was from Johns Hopkins University (Baltimore, MD, USA); Detroit562 and HN4 were purchased from American Type Culture Collection (Manassas, VA, USA). All the cell lines were cultured in DMEM (with 100 µg/mL streptomycin and 100 U/mL penicillin) (KeyGEN Biotech, Nanjing, China) supplemented with 10% FBS (BI, Migdal Haemek, Israel), in a humidified incubator 5% CO2/20% O2 or 5% CO2/1% O2 at 37°C, represented as normoxic and hypoxic tumor environments, respectively. Confluent cells were trypsinized with 0.05% trypsin containing 0.02% EDTA (Thermo Fisher Scientific, Waltham, MA, USA).
Flow cytometry
HNSCC cell lines were harvested and washed with PBS containing 3% BSA. After washing, the monoclonal anti-human PE-conjugated EGFR antibody (BioLegend, San Diego, CA, USA) was added to the cells and incubated for 30 minutes at 4°C in the dark. The cells were washed three times with PBS and analyzed using a FACSVerse flow cytometer (BD Biosciences, San Jose, CA, USA) to measure the expression of EGFR. The data obtained were analyzed by FlowJo (Tree Star, Inc., Ashland, OR, USA). Apoptosis assays and cell cycle analysis were detected by flow cytometry as described in detail below.
Quantitative real-time PCR assay
Total RNA was isolated using TRIzol reagent. Reverse transcription was performed using a PrimeScript™ RT Reagent Kit with gDNA Eraser (Takara Biotechnology, Dalian, China) according to the manufacturer’s instructions. RT-PCR was performed on ViiA™ 7 (Thermo Fisher Scientific) using the ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech, Nanjing, China) according to the manufacturer’s protocol. β-Actin was used as the reference gene, and results were expressed as the relative expression ratio of target gene to reference gene. Data were analyzed with the 2−ΔΔCt method. The sequences for the primers used in this study are listed in .
Table 1 Sequences of primers used in this study
Western blot assay
Following the appropriate treatments, the cells were lysed on ice using the mammalian protein extraction reagent RIPA (Beyotime, Shanghai, China) supplemented with a protease inhibitor cocktail (Hoffman-La Roche Ltd, Basel, Switzer-land) and phenylmethylsulfonyl fluoride (Beyotime). The supernatants were collected by centrifugation at 12,000× g at 4°C for 25 minutes. Total protein concentrations were determined with a bicinchoninic acid protein assay kit (KeyGEN Biotech). Protein samples were mixed with 5× loading buffer (GenScript, Nanjing, China) and heated at 95°C for 10 minutes. Equal amounts of protein were separated by SDS-PAGE, transferred to a 0.22 mm itrocellulose membrane (EMD Millipore, Billerica, MA, USA) and blocked by incubation with 5% fat-free milk in TBST buffer (150 mM NaCl, 50 mM Tris-HCl, 0.5% Tween 20, pH 7.6) at room temperature for 2 hours. The membranes were incubated with primary antibodies at 4°C overnight and then with horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 2 hours, prior to being exposed with ECL reagent (EMD Millipore). The pictures were captured by a Tanon 6200 Luminescent Imaging Workstation (Tanon, Shanghai, China). The following primary antibodies were used to detect proteins: rabbit anti-cyclin D1 (1:10,000; Abcam, Cam-bridge, UK), E-cadherin (1:500; Abcam), vimentin (1:2,000; Abcam), slug (1:1,000; Abcam), α-smooth muscle actin (α-SMA; 1:2,000; Abcam), phospho-AKT (1:1,000; Cell Signaling Technology, Danvers, MA, USA), AKT (1:1,000; Cell Signaling Technology), phospho-ERK (1:1,000; Cell Signaling Technology), ERK (1:1,000; Cell Signaling Technology), mouse anti-twist (1:500; Abcam), and anti-β-actin (1:2,000; Proteintech, Rosemont, IL, USA).
Patient cohort
A total of 30 patients diagnosed with HNSCC at the Department of Oral and Maxillofacial Surgery, Nanjing Stomatological Hospital, Medical School of Nanjing University between 2007 and 2011 were included in this study. All patients provided their written informed consent. The mean and median age at diagnosis was 61.17 and 61 years old, respectively (range, 46–81 years). The detailed clinicopathological parameters are provided in .
Table 2 Clinical and pathological characteristics of HNSCC
Histopathological analysis and immunohistochemistry
Samples from clinical patients and animal models were collected. Tissue sections (4 µm thick) were obtained, deparaffinized, and subjected to antigen recovery treatment with 100 mM citrate buffer target retrieval solution, pH 6.0 at 95°C, in a water bath for 20 minutes. Endogenous peroxidase activity was blocked by incubating with PBS and 3% hydrogen peroxidase for 30 minutes. After washing with PBS, the sections were incubated with rabbit anti-cyclin D1 (1:500; Abcam), E-cadherin (1:1,600; Abcam), HIF-1α (1:400; Abcam), and Ki67 (Typing, Nanjing, China) overnight at 4°C, followed by the Envision Dual Link System HRP method (Dako Denmark A/S, Glostrup, Denmark). All the antibodies were diluted in Dako antibody diluent. Reactions were revealed by incubating the sections with 3,3´-diaminobenzidine tetrahydrochloride (Dako Denmark A/S). Three pathologists independently scored the immunohistochemically stained slides. The scoring was based on the extent (E) of staining (percentage of positive tumor cells graded on a scale from 0 to 3: 0, none; 1, 1%–25%; 2, 26%–50%; 3, 51%–75%; 4, 75%–100%) and the intensity (I) of staining (graded on a scale of 0–3: 0, none; 1, weak staining; 2, moderate staining; 3, strong staining). Finally, the scores were calculated using the formula: scores = ∑(E×I). In vivo, hypoxia was detected by Hypoxyprobe™-1 Plus Kits (EMD Millipore) according to the manufacturer’s instructions. In brief, 15 minutes before being sacrificed, mice were intraperitoneally injected with a Hypoxyprobe™-1 (pimonidazole HCl) solution at a dosage of 100 mg/kg body weight. The xenograft tumors were removed for formalin fixation, paraffin-embedded, followed by immunostaining analysis with FITC-MAb1 (primary antibody, 1:100) and peroxidase-conjugated anti-FITC secondary reagent (1:100). The remaining steps were carried out as mentioned earlier.
Cell viability assay
The cell viability of HNSCC cells following treatment of gefitinib (Selleck Chemicals, Houston, TX, USA), palbociclib (Selleck Chemicals), LY294002 (Selleck Chemicals), and metformin (Sigma Aldrich Co, St Louis, MO, USA) was tested by using cell counting kit-8 (CCK-8) (Bimake, Houston, TX, USA) following the manufacturer’s instructions. Cells were plated in 96-well plates at a density of 3,000 cells in 100 µL complete culture medium. Blank control wells containing the same volume of complete culture medium were included in each assay. The microplate was incubated overnight at 37°C until cells adhered to the plates. The cells were then washed with PBS and incubated with gefitinib or metformin, which were serially diluted in complete culture medium before use. The plate was further incubated for 48 hours. CCK-8 (10 µL) was added to each well. The OD of formazan at 450 nm was recorded every 0.5 hour until the OD reached 1.0–2.0. Six wells corresponded to each concentration of the abovementioned drugs. Cell viability was calculated as follows: cell viability = ([OD] test − [OD] control)/([OD] control − [OD] blank) ×100%. The IC50 value was calculated using GraphPad Prism 7 (GraphPad Software Inc, La Jolla, CA, USA).
Apoptosis detection
HNSCC cells were seeded into 6-well plates. The cells were incubated overnight and then treated with different agents under normoxic or hypoxic conditions. After 48 hours, the apoptotic cells were quantified by Annexin V-FITC/Propidium Iodide (PI) Cell Apoptosis Detection Kits (KeyGEN Biotech) according to the manufacturer’s protocol. Samples were detected with a FACSVerse flow cytometer (BD Bio-sciences), and the data obtained were analyzed by FlowJo. The apoptosis rate in every group was described by Annexin V+ PI+ (late apoptosis) + Annexin V+ PI− (early apoptosis).
Cell cycle analysis
HNSCC cells were seeded into 6-well plates. The cells were incubated overnight and then treated with different agents under normoxic or hypoxic conditions. After 48 hours, flow cytometry was used to detect the cell cycle using Cell Cycle Detection Kits (KeyGEN Biotech). Samples were detected with a FACSVerse flow cytometer (BD Biosciences), and the data obtained were analyzed by FlowJo.
In vivo assay
All the animals used in this study were from the Nanjing Biomedical Research Institute of Nanjing University (Nanjing, China). Mice were maintained under specific pathogen-free conditions (N=24, six mice/group). The Immunocompetent nude mice (BALB/cJNju-Foxn1nu/Nju) were treated with 30 µL HSC3 (2×107/mL; six mice/group) injected into the tongue. Tumor volume was measured every 3 days with calipers and calculated using the formula (volume = long diameter × short diameter2/2). After 10 days, the mice were orally gavaged every day with PBS, gefitinib (60 mg/kg), metformin (200 mg/kg), or gefitinib plus metformin. After 3 weeks of treatment, the mice were killed and the xenograft tumors were removed and fixed in formalin, paraffin-embedded, and H&E staining was performed. Immunostaining analysis for HIF-1α, cyclin D1, E-cadherin, and Ki67 protein expression was also carried out, and apoptosis was observed by TUNEL Apoptosis Detection Kits (Sigma-Aldrich Co) according to the manufacturer’s instructions. Hypoxic areas in the tumors were detected by Hypoxyprobe™-1 Plus Kits (EMD Millipore) as previously described.
Statistical analyses
All data are expressed as mean ± SD. Statistical differences were measured using an unpaired two-sided Student’s t-test or one-way ANOVA for multiple comparisons when appropriate. All statistical analyses were performed using SPSS 17.0 statistical software package (SPSS Inc, Chicago, IL, USA). Statistical significance was set at an alpha value of P<0.05. All the graphs were drawn using GraphPad Prism 7 (GraphPad Software Inc).
Results
Hypoxia induced HNSCC gefitinib treatment resistance in vitro
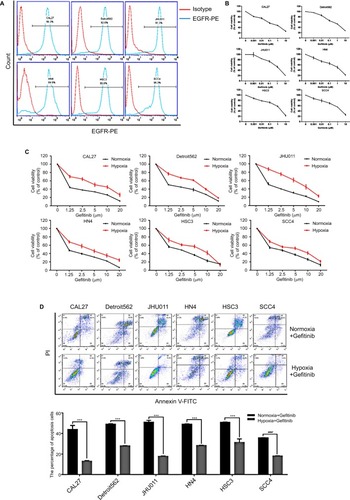
We first evaluated the expression level of EGFR in HNSCC. A total of six HNSCC cell lines (CAL27, Detroit562, JHU011, HN4, HSC3, and SCC4) were included in this experiment. Flow cytometry results showed that all six of the HNSCC cell lines exhibited high expression levels of EGFR (>90%) (), which is in accordance with previous reports.Citation8 Then, we examined the ability of gefitinib to inhibit the proliferation of HNSCC cell lines. Cell viability assay results showed that gefitinib alone could inhibit the proliferation of HNSCC cell lines to a certain degree under normoxic conditions, illustrating the drug’s efficacy and effectiveness in tumor control (). The IC50 values of gefitinib in the HNSCC cell lines, as calculated by GraphPad Prism 7, are listed in . To further explore the effect of tumor hypoxia on the regulation of gefitinib sensitivity, HNSCC cell lines were cultured with gefitinib under normoxic and hypoxic conditions. The results of the cell viability assay demonstrated a significant increase in cell survival during hypoxia (P<0.05), suggesting the role of hypoxia in desensitizing cells to treatment with gefitinib (). The results of the cell apoptosis assay also showed a significant difference between the normoxic and hypoxic groups (, P<0.05), confirming that hypoxia can induce gefitinib treatment resistance in vitro.
Table 3 IC50 values for HNSCC cell lines treated with gefitinib under normoxic conditions
Figure 1 Hypoxia induced HNSCC gefitinib treatment resistance.
Notes: (A) The expression levels of EGFR in HNSCC cell lines were detected by flow cytometry analysis. (B) Cell viability of HNSCC cell lines treated with the indicated doses of gefitinib in normoxia for 48 hours was assessed with the CCK-8 assay. (C) After being treated with the indicated doses of gefitinib in normoxia or hypoxia for 48 hours, cell viability was assessed by the CCK-8 assay. (D) Cell apoptosis of HNSCC cell lines was assessed by flow cytometry following treatment with their own normoxia IC50 value in normoxia or hypoxia for 48 hours. Images are representative of three independent experiments (***P<0.001 compared with normoxia).
Abbreviations: HNSCC, head and neck squamous cell carcinoma; CCK-8, cell counting kit-8; HNSCC, head and neck squamous cell carcinoma; PI, propidium iodide; PE, Phycoerythrin..
Hypoxia regulated HNSCC cell cycle progression and EMT in vitro and in vivo
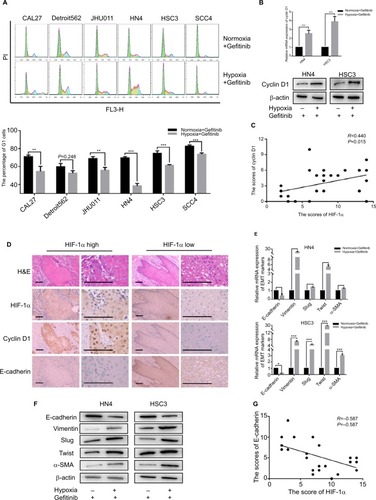
Since hypoxia improved cell viability in cells co-cultured with gefitinib, we hypothesized that it might function via regulation of the cell cycle. Our in vitro experiment showed that the percentage of HNSCC cells in the G1 phase decreased to varying degrees in response to hypoxic stimulation and 1 µM gefitinib treatment (). It is well known that cyclin D1 is a key factor in cell cycle regulation; therefore, we chose two cell lines among the six available HNSCC cell lines, HN4 and HSC3, to detect the expression of cyclin D1 at the mRNA and protein level when the cells were cultured with gefitinib under hypoxic conditions. The results showed that hypoxia significantly promoted the expression of cyclin D1, which may lead to regulation of the cell cycle (). To further explore the relationship between hypoxia and cyclin D1 expression, 30 HNSCC clinical samples were included in this study. Immunohistochemical staining analysis demonstrated that the expression of HIF-1α was positively correlated with cyclin D1 (R=0.44, P=0.015) (). The representative pictures are shown in . Shien K et al reported that EMT is strongly related to gefitinib resistance;Citation24 Kohnoh et al claimed that hypoxia can induce lung cancer cell EMT.Citation25 Therefore, it was reasonable for us to further analyze the status of EMT treated with gefitinib in cells treated with gefitinib under hypoxic conditions. Encouragingly, compared with normoxic cells, hypoxic HNSCC cells demonstrated significantly higher levels of EMT markers (E-cadherin, vimentin, slug, twist, and α-SMA) at the mRNA and protein level in HN4 and HSC3 cell lines (). Further study showed that in clinical samples, the expression of E-cadherin was negatively correlated with the expression of HIF-1α (R=−0.587, P=0.001) (). These data indicated that hypoxia regulates the cell cycle and EMT in vitro and in vivo, which might partially explain the mechanisms behind hypoxia-induced gefitinib resistance.
Figure 2 Hypoxia promoted G1–S cell cycle progression and EMT.
Notes: After being treated with 1 µM gefitinib in normoxia or hypoxia for 48 hours, (A) cell cycle was detected by flow cytometry. (B) The expression of cyclin D1 in mRNA and protein level was analyzed by qPCR and Western blot (**P<0.01 and ***P<0.001 compared with normoxia). (C) Relativity between the expression of HIF-1α and cyclin D1 in clinical HNSCC specimens (n=30, R=0.440, P=0.015). (D) Representative images of H&E staining and IHC of HIF-1α, cyclin D1, E-cadherin in HNSCC specimens (scale plate represents 200 µm). (E and F) Expression of EMT markers in mRNA and protein level was analyzed by qPCR and Western blot following treatment with their own IC50 value in normoxia or hypoxia for 48 hours (*P<0.05, **P<0.01, and ***P<0.001 compared with normoxia). (G) Relativity between the expression of HIF-1α and E-cadherin in clinical HNSCC specimens (n=30, R=0.587, P=0.001).
Abbreviations: EMT, epithelial–mesenchymal transition; HNSCC, head and neck squamous cell carcinoma; HIF-1α, hypoxic inducible factor-1α; IHC, immunohistochemistry; PI, propidium iodide; qPCR, quantitative PCR; α-SMA, α-smooth muscle actin; PI, propidium iodide.
Inhibition of cyclin D1 and EMT re-sensitized HNSCC to gefitinib treatment
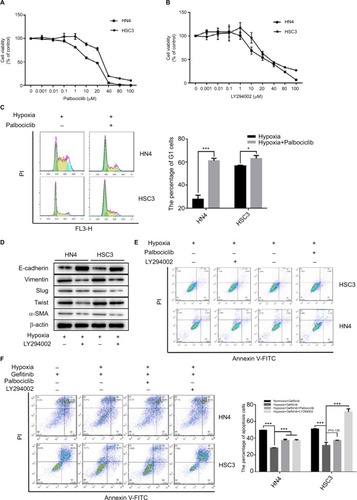
Since we concluded that HNSCC tumor hypoxia might desensitize cells to gefitinib treatment via regulation of the cell cycle and induction of EMT, it was necessary to explore whether inhibition of such signaling might re-establish the efficacy and effectiveness of gefitinib under hypoxic conditions. Palbociclib, a well-known cyclin D1 inhibitor, was used in this study.Citation26 First, we devaluated the effects of palbociclib on the inhibition of cell proliferation. The results demonstrated that palbociclib alone (0.1 µM) did not induce obvious cell cytotoxicity (). Thus, we chose this concentration for the subsequent experiments. For the same reason, we tested the effect of the EMT inhibitor – LY294002,Citation27 and confirmed that a concentration of 1 µM was appropriate (). Our in vitro experiment confirmed that 0.1 µM palbociclib could significantly change the percentage of cells in the G1 phase during hypoxia in HN4 and HSC3 cells (P<0.05) (), and 1 µM LY294002 reversed tumor cell EMT during hypoxia (). To clearly demonstrate the final treatment effect, combined therapy (gefitinib plus palbociclib or gefitinib plus LY294002) was applied to compare the results to the single agent gefitinib results under normoxic and hypoxic conditions. The results proved that the combined treatment significantly enhanced the percentage of apoptotic cells (), while treatment with palbociclib (0.1 µM) or LY294002 (1 µM) alone did not induce significant cell apoptosis (). These results illustrate the synergistic effect of such agents and emphasize the importance of inhibiting the function of cyclin D1 and EMT reversal.
Figure 3 The inhibitor of cyclin D1 and EMT could re-sensitize HNSCC cells to gefitinib in hypoxia.
Notes: (A and B) HN4 and HSC3 cells were treated with the indicated doses of palbociclib or LY294002 for 48 hours. Cell viability was assessed by CCK-8 assay. (C) The cell cycle of HN4 and HSC3 was detected by flow cytometry following treatment with palbociclib (0.1 µM) in hypoxia for 48 hours (*P<0.05 and ***P<0.001 compared with control). (D and E) After treatment with LY294002 (1 µM) in hypoxia for 48 hours, the expression of EMT markers among HN4 and HSC3 cells was analyzed by Western blot; flow cytometry was used to detect the apoptosis condition. (F) The apoptosis condition of HN4 and HSC3 cells was assessed by flow cytometry following treatment with gefitinib (IC50 value) combined with palbociclib (0.1 µM) or LY294002 (1 µM) in hypoxia for 48 hours. Images are representative of three independent experiments (***P<0.001 compared with normoxia).
Abbreviations: HNSCC, head and neck squamous cell carcinoma; CCK-8, cell counting kit-8; EMT, epithelial–mesenchymal transition; HNSCC, head and neck squamous cell carcinoma; PI, propidium iodide; α-SMA, α-smooth muscle actin; PI, propidium iodide.
Metformin re-sensitized cells to hypoxia-induced gefitinib resistance in vitro and in vivo
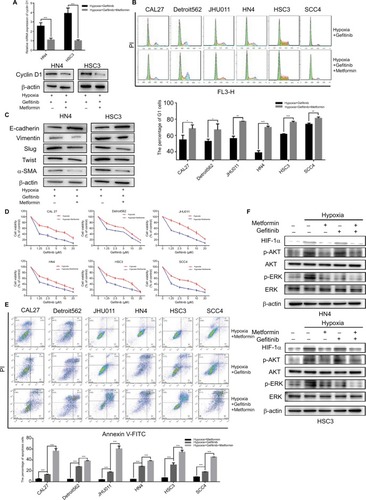
There have been many reports of the versatile nature of metformin in the inhibition of cancer.Citation28–Citation30 Our previous study confirmed the effects of metformin in the suppression of HIF-1α expression and the subsequent enhancement of the therapeutic effects of cisplatin.Citation23 In addition, metformin is a clinically approved drug which may have great potential for future clinical trials if the capacity to re-sensitize tumors to gefitinib is shown. Our in vitro experiment demonstrated that metformin (100 µM) could significantly decrease the expression of cyclin D1 under hypoxic conditions () and change the percentage of G1 cells () when cells were cultured with gefitinib. Meanwhile, metformin also decreased the expression of EMT markers during hypoxia along with the treatment of gefitinib (). More importantly, the combined use of metformin and gefitinib significantly inhibited cell growth and induced apoptosis in all six HNSCC cell lines, compared with metformin and gefitinib alone (, P<0.05). It has been reported that gefitinib alone might activate parallel signaling pathways such as PI3K/AKT, which may partially explain the resistance of EGFR TKIs.Citation31 Hence, we explored the activation status of the aforementioned signaling pathway. Western blot clearly demonstrated that gefitinib could partially increase the expression of p-AKT, while no significant change in p-ERK was observed. Encouragingly, the addition of metformin might suppress HIF-1α expression along with p-AKT in both HSC3 and HN4 cells (). These results provide strong evidence that metformin can regulate oncogenic signaling to induce cell cycle arrest and EMT, as well as that gefitinib itself can activate cell signaling pathways such as PI3K/AKT. The combined treatment strategy may increase apoptosis and achieve far better therapeutic effects in vitro.
Figure 4 Metformin sensitized HNSCC cells to gefitinib in hypoxic condition in vitro.
Notes: (A and B) HN4 and HSC3 cells were treated with gefitinib (1 µM) alone or combined with metformin (100 µM) in hypoxia for 48 hours. Whole cell protein was immunoblotted with indicated antibodies, and the cell cycle was analyzed by flow cytometry (*P<0.05, **P<0.01, and ***P<0.001 compared with control). (C) The expression of EMT markers among HN4 and HSC3 cells was analyzed by Western blot following treatment with gefitinib (IC50 value) alone or in combination with metformin (100 µM) in hypoxia for 48 hours. (D) After being treated with the indicated doses of gefitinib combined with metformin (100 µM) in normoxia or hypoxia for 48 hours, cell viability was assessed by the CCK-8 assay. (E) The apoptosis condition of HNSCC cell lines was assessed by flow cytometry after treatment with gefitinib (IC50 value), metformin (100 µM), or in combination in hypoxia for 48 hours. Images are representative of three independent experiments (***P<0.001 compared with control). (F) HN4 and HSC3 cells were treated with gefitinib (IC50 value), metformin (100 µM), or in combination in normoxia or hypoxia for 48 hours. Whole cell protein was immunoblotted with indicated antibodies.
Abbreviations: HNSCC, head and neck squamous cell carcinoma; CCK-8, cell counting kit-8; EMT, epithelial–mesenchymal transition; HIF-1α, hypoxic inducible factor-1α; HNSCC, head and neck squamous cell carcinoma; PI, propidium iodide α-SMA, α-smooth muscle actin; PI, propidium iodide; HIF-1α, hypoxic inducible factor-1α.
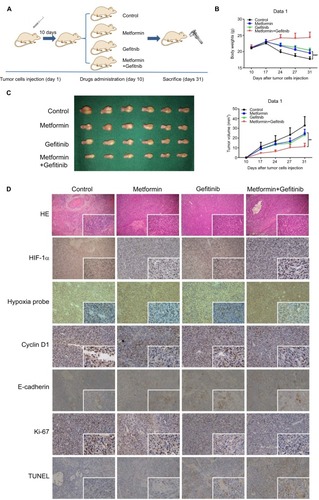
To further examine the in vivo therapeutic effects of our combined treatment strategy, orthotopic HNSCC animal models were established. Ten days after injection of 30 µL HSC3 (2×107/mL), 24 mice were evenly distributed into four groups to receive PBS, metformin, gefitinib, or metformin plus gefitinib. The dose of gefitinib (60 mg/kg/day) and metformin (200 mg/kg/day) used in this study was selected based on studies of Li et al and Al-Wahab et al.Citation32,Citation33 Following treatment, tumor volume and body weight were measured every 3 days. The mice were killed 3 weeks after the treatment (). Our in vivo study revealed a significant increase in body weight in the combined treatment group compared with the single agent and control groups (), which might explain why the combined treatment could exert better tumor inhibition effect in the tongue, and it allowed the mice to have more food intake. Tumor volume was significantly decreased in the combined treatment group. Interestingly, we also found that metformin alone exerted antitumor effects to some degree when compared with the control group (P<0.05). This may be explained by the effect of metformin on the inhibition of hypoxic signaling (). The IHC results supported our hypothesis that the combined treatment may inhibit the expression of HIF-1α and decrease the expression of cyclin D1 and Ki-67. Reversal of EMT, reflected by increased expression of E-cadherin and significant positive TUNEL staining, was also observed (). All these data strongly indicated that metformin might have a synergistic effect with gefitinib in the treatment of HNSCC.
Figure 5 Metformin enhanced the chemotherapeutic efficacy of gefitinib in vivo.
Notes: (A) Schematic figure of the orthotopic xenograft model used in this study. (B) Body weight (g) of mice treated with PBS, metformin (200 mg/kg), gefitinib (60 mg/kg), and metformin plus gefitinib. (C) Macroscopic appearance of the tumors on day 31 after drug administration and tumor volume (mm3) during treatment. (D) Paraffin-embedded sections (4 mm) from tumor tissues were stained for HIF-1α, hypoxia probes, cyclin D1, E-cadherin, Ki67, and TUNEL using IHC (100× and 400×). **P<0.01, and ***P<0.001.
Abbreviations: HIF-1α, hypoxic inducible factor-1α; IHC, immunohistochemistry.
Discussion
Currently, gefitinib is widely regarded as one of the most effective molecular target therapeutic drugs used in lung cancers, especially non-small-cell lung cancer (NSCLC).Citation34 Gefitinib targets EGFR by competitively combining with ATP in the receptor tyrosine kinase of the catalytic center of the Mg-ATP binding site. Consequently, gefitinib inhibits tyrosine kinase activity, blocks intracellular tyrosine kinase phosphorylation, and suspends EGFR signal conduction, thus blocking the growth of tumor cells.Citation35 Despite the effectiveness and efficacy reported in lung cancer, gefitinib has not been suggested as a possible alternative approach in HNSCC treatment guidelines, especially as a first-line drug. Gefitinib is also not yet approved by the FDA, which might be explained by the observed treatment resistance and the limited benefits gained by patients in reported clinical trials.Citation11 Thus, a deeper understanding of the mechanisms behind gefitinib treatment resistance is crucially important. In the present study, HNSCC tumor hypoxia was proven to be one of the major causes of treatment resistance, which is in accordance with previous reports.Citation36–Citation38 An et al reported that hypoxia played an important role in gefitinib adaptive resistance in NSCLC due to the inductive expression of IL-6.Citation39 Murakami et al reported that hypoxia could increase the proportion of cancer stem cells in lung cancer, which in return would generate more gefitinib-resistant cells.Citation17 Our work demonstrated that hypoxia induced HNSCC gefitinib resistance by regulating the cell cycle and inducing EMT.
Due to our earlier findings, overcoming hypoxia-induced gefitinib treatment resistance in HNSCC has been our primary goal. Basic principles to re-sensitize tumor cells to gefitinib treatment include simplicity and practicability in the clinic. Recently, many studies have confirmed the anticancer effects of metformin.Citation40–Citation42 In addition, our previous work has also revealed that metformin inhibited the expression of HIF-1α through interference with the nuclear factor-κB pathway under hypoxic conditions and then sensitized oral squamous cell carcinoma to cisplatin.Citation23 Therefore, we hypothesized that the combination of metformin with gefitinib may lead to a better curative effect during hypoxia in HNSCC. In line with our research, there are clinical trials currently in progress utilizing treatment with metformin and gefitinib (NCT01864681, NCT03071705, and NCT01997775; www.ClinicalTrials.gov) in lung cancer patients. To our knowledge, there have been no similar clinical trials yet in HNSCC. In the present study, we showed that metformin could overcome hypoxia-induced gefitinib resistance, which is favorable evidence for possible future application in HNSCC.
Although metformin is a commonly used hypoglycemic drug, the safety and possible systemic toxicity are still of major concern. In our study, the dose of metformin (100 µmol/L) is higher than that used in diabetic patients. Moreover, Iliopoulos et al proved that metformin tends to accumulate in tissues rather than blood; thus, a range of drug concentrations from 1 to 10 mmol/L would not affect normal blood sugar metabolism.Citation43 In addition, Morgillo et al validated the safety of 1,500 mg metformin per day in nondiabetic patients. This dose corresponds to ~308 mg/kg in the mouse, which is higher than what we used in our study.Citation44,Citation45 Thus, we have provided strong evidence for the effectiveness and safety of metformin in our study.
Despite the encouraging outcomes obtained, challenges remain. Drug resistance is a consequence of multiple driving factors. It is difficult to determine the contribution of the microenvironment in this process, especially tumor hypoxia. Therefore, the strategy of inhibiting hypoxia signaling to overcome barriers is unrealistic, and more trials of combined therapy are desperately needed. In addition, although metformin is reported to have anticancer effects, it cannot be included as a first-line therapeutic drug. Hence, it is necessary to further explore the mechanisms of metformin and more novel treatment strategies in the field of HNSCC molecular target therapy.
Conclusion
This study demonstrates that metformin has the potential to synergize with gefitinib to overcome hypoxia-induced treatment resistance in HNSCC via cell cycle regulation and EMT reversal. Thus, a combination of metformin with EGFR TKIs is implicated as a promising therapeutic strategy for HNSCC.
Acknowledgments
This study was supported by Jiangsu Provincial Key Research and Development Program (BE2017741), Nanjing Foundation for Development of Science and Technology (2017sc512031 and 201605042), Jiangsu Provincial Natural Science Foundation (BK20150089 and BK 20160051), the Distinguished Young Investigator Project of Nanjing (JQX14010), and Medical Science and Technology Development Foundation (YKK15116). We would like to thank LetPub (www.letpub.com) for providing linguistic assistance during the preparation of this manuscript.
Author contributions
WH and ZTL conceived and designed the study. XTY, ZW, JQX, CHS, CCT, WGX and YFW performed the experiments and analyzed the data. XTY and ZW wrote the paper; WH and ZTL reviewed and edited the manuscript. All authors contributed toward data analysis, drafting and critically revising the paper, gave final approval of the version to be published and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
- TorreLABrayFSiegelRLFerlayJLortet-TieulentJJemalAGlobal cancer statistics, 2012CA Cancer J Clin20156528710825651787
- MillerKDSiegelRLLinCCCancer treatment and survivorship statistics, 2016CA Cancer J Clin201666427128927253694
- MarurSForastiereAAHead and Neck Squamous Cell Carcinoma: Update on Epidemiology, Diagnosis, and TreatmentMayo Clin Proc201691338639626944243
- MetroGEGFR targeted therapy for lung cancer: are we almost there?Transl Lung Cancer Res20187Suppl 2S142S14529782570
- BhattiMIppolitoTMavisCPre-clinical activity of targeting the PI3K/Akt/mTOR pathway in Burkitt lymphomaOncotarget2018931218202183029774105
- YangJYanJLiuBTargeting VEGF/VEGFR to Modulate Antitumor ImmunityFront Immunol2018997829774034
- ToschiLRossiSFinocchiaroGSantoroANon-small cell lung cancer treatment (r)evolution: ten years of advances and more to comeEcancermedicalscience20171178729225694
- KalyankrishnaSGrandisJREpidermal growth factor receptor biology in head and neck cancerJ Clin Oncol200624172666267216763281
- SoulieresDSenzerNNVokesEEHidalgoMAgarwalaSSSiuLLMulticenter phase II study of erlotinib, an oral epidermal growth factor receptor tyrosine kinase inhibitor, in patients with recurrent or metastatic squamous cell cancer of the head and neckJ Clin Oncol2004221778514701768
- MagnesTEgleAGreilRMelchardtTUpdate on squamous cell carcinoma of the head and neck: ASCO annual meeting 2017Memo201710422022329250200
- CohenEERosenFStadlerWMPhase II trial of ZD1839 in recurrent or metastatic squamous cell carcinoma of the head and neckJ Clin Oncol200321101980198712743152
- MasekiSIjichiKTanakaHAcquisition of EMT phenotype in the gefitinib-resistant cells of a head and neck squamous cell carcinoma cell line through Akt/GSK-3β/snail signalling pathwayBr J Cancer201210661196120422315058
- JamesonMJBecklerADTaniguchiLEActivation of the insulin-like growth factor-1 receptor induces resistance to epidermal growth factor receptor antagonism in head and neck squamous carcinoma cellsMol Cancer Ther201110112124213421878657
- JiaYYunCHParkEOvercoming EGFR(T790M) and EGFR(C797S) resistance with mutant-selective allosteric inhibitorsNature2016534760512913227251290
- OcañaMCMartínez-PovedaBQuesadaARMedinaMÁMetabolism within the tumor microenvironment and its implication on cancer progression: An ongoing therapeutic targetMed Res Rev Epub2018522
- MaSPradeepSHuWZhangDColemanRSoodAThe role of tumor microenvironment in resistance to anti-angiogenic therapyF1000Res2018732629560266
- MurakamiATakahashiFNurwidyaFHypoxia increases gefitinib-resistant lung cancer stem cells through the activation of insulin-like growth factor 1 receptorPLoS One201491e8645924489728
- MinakataKTakahashiFNaraTHypoxia induces gefitinib resistance in non-small-cell lung cancer with both mutant and wild-type epidermal growth factor receptorsCancer Sci2012103111946195422863020
- HanSXuWWangZCrosstalk between the HIF-1 and Toll-like receptor/nuclear factor-κB pathways in the oral squamous cell carcinoma microenvironmentOncotarget2016725377733778927191981
- Wahdan-AlaswadRSEdgertonSMSalemHSThorADMetformin Targets Glucose Metabolism in Triple Negative Breast CancerJ Oncol Transl Res201841pii:129
- TsengCHMetformin and risk of developing nasopharyngeal cancer in patients with type 2 diabetes mellitusMetabolism20188522322629729227
- WooSHYangLPChuangHCDown-regulation of malic enzyme 1 and 2: Sensitizing head and neck squamous cell carcinoma cells to therapy-induced senescenceHead Neck201638Suppl 1E934E94025994759
- QiXXuWXieJMetformin sensitizes the response of oral squamous cell carcinoma to cisplatin treatment through inhibition of NF-κB/HIF-1α signal axisSci Rep201663578827762347
- ShienKToyookaSYamamotoHAcquired resistance to EGFR inhibitors is associated with a manifestation of stem cell-like properties in cancer cellsCancer Res201373103051306123542356
- KohnohTHashimotoNAndoAHypoxia-induced modulation of PTEN activity and EMT phenotypes in lung cancersCancer Cell Int2016163327095949
- FryDWHarveyPJKellerPRSpecific inhibition of cyclin-dependent kinase 4/6 by PD 0332991 and associated antitumor activity in human tumor xenograftsMol Cancer Ther20043111427143815542782
- RekaAKKuickRKurapatiHStandifordTJOmennGSKeshamouniVGIdentifying inhibitors of epithelial–mesenchymal transition by connectivity map-based systems approachJ Thorac Oncol20116111784179221964532
- CamachoLDasguptaAJiralerspongSMetformin in breast cancer - an evolving mysteryBreast Cancer Res2015178826111812
- YouACaoMGuoZMetformin sensitizes sorafenib to inhibit postoperative recurrence and metastasis of hepatocellular carcinoma in orthotopic mouse modelsJ Hematol Oncol201692026957312
- El-ArabeyAANew insight for metformin against bladder cancerGenes Environ2017391328373897
- JanmaatMLKruytFARodriguezJAGiacconeGResponse to epidermal growth factor receptor inhibitors in non-small cell lung cancer cells: limited antiproliferative effects and absence of apoptosis associated with persistent activity of extracellular signal-regulated kinase or Akt kinase pathwaysClin Cancer Res2003962316232612796401
- Al-WahabZMertITebbeCMetformin prevents aggressive ovarian cancer growth driven by high-energy diet: similarity with calorie restrictionOncotarget2015613109081092325895126
- LiLHanRXiaoHMetformin sensitizes EGFR-TKI-resistant human lung cancer cells in vitro and in vivo through inhibition of IL-6 signaling and EMT reversalClin Cancer Res201420102714272624644001
- OnnATsuboiMThatcherNTreatment of non-small-cell lung cancer: a perspective on the recent advances and the experience with gefitinibBr J Cancer200491Suppl 2S11S1715340373
- TetsuOHangauerMJPhuchareonJEiseleDWMccormickFDrug Resistance to EGFR Inhibitors in Lung CancerChemotherapy201661522323526910730
- StegemanHKaandersJHWheelerDLActivation of AKT by hypoxia: a potential target for hypoxic tumors of the head and neckBMC Cancer20121246323046567
- VigneswaranNWuJSongAAnnapragadaAZachariasWHypoxia-induced autophagic response is associated with aggressive phenotype and elevated incidence of metastasis in orthotopic immunocompetent murine models of head and neck squamous cell carcinomas (HNSCC)Exp Mol Pathol201190221522521236253
- LöckSPerrinRSeidlitzAResidual tumour hypoxia in head-and-neck cancer patients undergoing primary radiochemotherapy, final results of a prospective trial on repeat FMISO-PET imagingRadiother Oncol2017124353354028843726
- AnSMLeiHMDingXPInterleukin-6 identified as an important factor in hypoxia- and aldehyde dehydrogenase-based gefitinib adaptive resistance in non-small cell lung cancer cellsOncol Lett20171433445345428927099
- XuSYangZJinPMetformin suppresses tumor progression by inactivating stromal fibroblasts in ovarian cancerMol Cancer Ther20181761291130229545331
- SuiXXuYWangXMetformin: A Novel but Controversial Drug in Cancer Prevention and TreatmentMol Pharm201512113783379126430787
- SivalingamVMcveyRGilmourKA presurgical window-of-opportunity study of metformin in obesity-driven endometrial cancerLancet2015385Suppl 1S9026312913
- IliopoulosDHirschHAStruhlKMetformin decreases the dose of chemotherapy for prolonging tumor remission in mouse xenografts involving multiple cancer cell typesCancer Res20117193196320121415163
- Reagan-ShawSNihalMAhmadNDose translation from animal to human studies revisitedFASEB J200822365966117942826
- MorgilloFFasanoMdella CorteCMResults of the safety run-in part of the METAL (METformin in Advanced Lung cancer) study: a multicentre, open-label phase I-II study of metformin with erlotinib in second-line therapy of patients with stage IV non-small-cell lung cancerESMO Open201722e00013228761738