
Abstract
Purpose
Colorectal cancer (CRC) is a heterogeneous disease that can be classified into distinct molecular subtypes. The aims of this study were 1) to compare claudin (CLDN) gene expression in CRC samples and normal colon mucosa, and then in the different CRC molecular subtypes, and 2) to assess their prognostic value.
Patients and methods
CLDN expression in CRC samples was analyzed using gene expression data for a cohort of 143 primary CRC samples, and compared in the same CRC samples classified into different molecular subtypes (C1 to C6 according to the Marisa’s classification, and CMS1 to CMS4 of the consensus classification). Comparison of CLDN expression in normal and tumor colon samples was also made on a smaller number of samples. Then, the relationship between CLDN expression profiles and overall survival (OS) and progression-free survival was examined.
Results
Compared with normal mucosa, CLDN1 and CLDN2 were upregulated, whereas CLDN5, 7, 8, and 23 were downregulated in CRC samples. Variations in CLDN expression profiles were observed mainly in the CMS2/C1 and CMS4/C4 subtypes. Overall, expression of CLDN2 or CLDN4 alone had a strong prognostic value that increased when they were associated. In the CMS4/C4 subtypes, lower expressions of CLDN11, CLDN12, and CLDN23 were associated with longer OS. Conversely, in the CMS2 and C1 subtypes, low CLDN23 expression was associated with shorter OS and progression-free survival, suggesting a dual role for CLDN23 as a tumor suppressor/promoter in CRC. CLDN6 and CLDN11 had a prognostic value in the CMS2 and C4 subtypes, respectively.
Conclusion
This analysis of CLDN gene expression profiles and prognostic value in CRC samples classified according to their molecular subtype shows that CRC heterogeneity must be taken into account when assessing CLDN potential value as prognostic markers or therapeutic targets.
Introduction
Colorectal cancer (CRC) originates from the oncogenic transformation of the intestinal epithelium that physiologically acts as a functional barrier between the intestinal mucosa and the luminal environment. It is now becoming clear that epithelial cell polarity is a major gatekeeper against cancer initiation and metastasis formation.Citation1 Epithelial cell polarity depends on the establishment of the apical junctional complex that includes tight junctions (TJs) and adherens junctions.Citation2 TJs, the most apical of these intercellular junctions, play an essential role in maintaining cell polarity and in the regulation of paracellular permeability.Citation3 Alterations in TJs by downregulation or upregulation of TJ proteins can trigger malignant transformation and influence cancer progression.Citation4
Claudins (CLDNs) are TJ core componentsCitation5 that are essential for TJ formationCitation6 and contribute to their selectivity.Citation7 In mammals, the CLDN family includes 27 members divided into two groups: classic and nonclassic CLDNs.Citation8 Almost all CLDNs have a short intracellular N-terminal domain, four transmembrane domains, two extracellular loops, and an intracellular C-terminal domain that contains a PDZ-domain-binding motif for linking to TJ-associated proteins, such as MUPP1, PATJ, ZO-1, ZO-2 and ZO-3, MAGUKs, PAR3, PAR6, and PALS.Citation9,Citation10 These proteins function as adaptors at the cytoplasmic surface of TJ strands and can directly or indirectly interact with cytosolic and nuclear proteins, for instance cytoskeletal molecules, regulatory proteins, tumor suppressors, and transcription factors.Citation11 Finally, some CLDNs interact with cell adhesion proteins (eg, EPCAM) or receptors (eg, EPHA and EPHB).Citation12 The cytoplasmic tail of most CLDNs contains a large number of predicted phosphorylation sites that could be involved in molecular interactions.Citation13 Accumulated evidence indicates that CLDNs are associated with various pathways, including the WNT/β-catenin, JAK-STAT3, and Notch signaling cascades.Citation7,Citation13,Citation14
CLDNs are expressed in a cell- and tissue-specific manner. In the intestine, CLDNs display specific spatiotemporal expression profiles with variations along the crypt–lumen axis.Citation15 Their expression can be regulated by various mechanisms at the transcriptional or posttranscriptional level, but also via mRNA stability modulationCitation16 and through epigenetic mechanisms.Citation16–Citation19
CLDN expression is altered in several cancer types in a tumor-specific manner, and can vary according to the tumor stage.Citation11 CLDN aberrant expression in tumors may have opposite functions (promotion of tumorigenesis and metastasis formation, or suppressive effects).Citation13,Citation20 For example, CLDN-1 is a cancer invasion/metastasis suppressor in lung adeno-carcinoma,Citation21 while in CRC, CLDN1 expression enhances the invasive ability and metastatic properties.Citation22 Moreover, some CLDNs have an important regulatory role in the epithelial–mesenchymal transition (EMT).Citation23–Citation25 CLDNs can also serve as a hub for different signaling proteins, and therefore could have a critical role in the regulation of carcinogenesis or cancer progression.Citation26 Finally, CLDN expression has been associated with patient survival, suggesting that they could be prognostic markers and/or therapeutic targets.Citation14,Citation27,Citation28
In CRC, research has focused mainly on CLDN1,Citation22,Citation28,Citation29 CLDN2,Citation18,Citation30,Citation31 and CLDN7.Citation24,Citation32,Citation33 Changes in the expression of TJ-related genes, including CLDNs, have been reported in CRC;Citation34,Citation35 however, these studies did not consider CRC heterogeneity, which can be described using molecular subtypes based on gene expression profiles.Citation36–Citation39
In this study, CLDN gene expression profiles were investigated in a cohort of 143 primary CRC samples classified according to their molecular subtype and for which gene expression and clinical data were available.Citation40–Citation42 The expression of each CLDN gene was first compared in normal and tumor colon samples, and then among the different CRC molecular subtypes. Finally, the prognostic value of the different expression profiles was evaluated.
Patients and methods
Gene expression analysis
In this study, expression data for tumor samples from 143 patients coming from three cohorts (REG/P,Citation40 COSIVAL, and BIOCOLONCitation41,Citation42) were used. These three studies were approved by the relevant ethics committees and all participants were informed about the study, and they signed a written informed consent before enrolment. All patients selected for this study had metastatic colorectal cancer (mCRC), and did not receive any chemotherapy treatment before primary tumor resection. Colon samples (normal colon, primary tumor, and hepatic metastasis samples from the REG/P cohort, and only primary tumor specimens from the COSIVAL and BIOCOLON cohorts) were collected at the time of surgery, following a standardized procedure to obtain high-quality RNA.Citation43 Samples were then hybridized to human genome U133 Plus 2.0 arrays (Affymetrix Inc., Santa Clara, CA, USA). The gene expression data can be found online at the Gene Expression Omnibus under the accession numbers GSE62080 and GSE72970.
All 143 CRC samples were classified using the molecular classifications based on gene expression profiles that have been proposed by Marisa et alCitation36 and Guinney et alCitation39 (), as described in each reference publication.Citation42 Briefly, Marisa et al described six molecular subtypes (C1 to C6) with the following main features: C1= CIN and immune pathway downregulation; C2= MSI; C3= mutated KRAS; C4= stem cell phenotype-like; C5= CIN and upregulation of the WNT pathways; and C6= CIN and normal-like gene expression profile.Citation36 The consensus classification includes four subtypes: CMS1 (microsatellite instability [MSI]-immune), CMS2 (epithelial and canonical), CMS3 (epithelial and metabolic), and CMS4 (mesenchymal). Most of the MSI-high tumors belong to the CMS1 subtype that has the best survival without recurrence. Chromosomal instability (CIN) tumors have a more heterogeneous gene expression pattern and, therefore, can be classified from CMS2 to CMS4. CMS4 tumors have a significantly higher risk of distant relapse.Citation38,Citation39
Table 1 Distribution of patients with mCRC according to the tumor molecular subtype
The Tsuji cohort (GSE28702)Citation44 includes 83 stage IV CRC samples from patients treated with the FOLFOX regimen. Dr Shingo Tsuji kindly provided the overall survival (OS) data for this series.
Statistical analyses
For gene expression analyses, differences between groups were determined using the Kruskal–Wallis/Dunn’s test.
Progression-free survival (PFS) was defined as the time from the beginning of first-line treatment for mCRC until recurrence or death. Alive patients without progression were censored at the date of last contact. OS was calculated from the beginning of first-line treatment until death. Correlations between CLDN gene expression and PFS or OS were evaluated in the entire group (n=143 patients) and according to the tumor molecular subtype. In each subtype, CRC samples were divided into two groups (high/low expression) based on the median CLDN gene expression. The Kaplan–Meier method was used to compare PFS and OS values, and the log-rank test was used to assess differences between survival distributions. For all experiments, differences were considered to be significant when P<0.05.
Results
CLDN gene expression patterns in colon
Analysis of the CLDN gene expression levels from Affymetrix data for 17 normal colon mucosa, 20 primary CRC samples, and 19 hepatic metastases (REG/P cohort)Citation40,Citation41 showed that 8 CLDN genes (CLDN6, 9, 10, 11, 14, 16, 17, and 18) were weakly expressed in all samples. Among the other ten claudins, CLDN1, CLDN3, CLDN4, and CLDN7 were strongly expressed in primary CRC and metastatic samples, and CLDN3, CLDN4, CLDN7, CLDN8, and CLDN23 in normal mucosa ().
Figure 1 Expression of CLDN genes.
Notes: (A) Mean expression of 18 CLDN genes in NM (n=17), PT (n=20), and HM (n=19). The Affymetrix RNA expression data were log2 transformed. (B) Detailed analysis of the RNA expression levels of six CLDN genes in the NM, PT, and HM samples described in (A). Horizontal bars indicate the mean value. ***P<0.001= (Kruskal–Wallis test).
Abbreviations: CRC, colorectal cancer; HM, hepatic metastases; NM, normal mucosa; PT, primary CRC tumors.
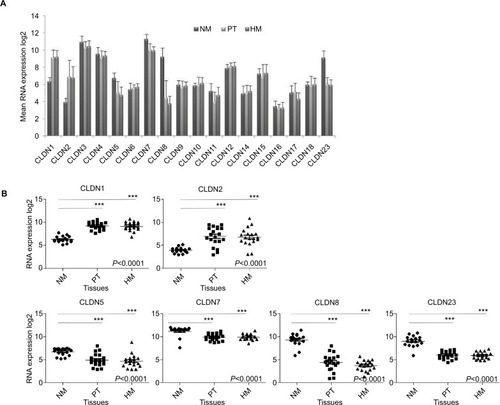
To determine whether CLDN gene expression levels change during tumorigenesis, CLDN expression profiles in normal mucosa, primary tumor, and hepatic metastases were compared. CLDN4, 6, 9, 10, 12, 14, 15, 16, 17, and 18 displayed similar expression levels in all tissue samples, whereas CLDN3 and CLDN11 showed a tendency to downregulation in primary tumor samples (Figure S1). On the other hand, CLDN1 and CLDN2 were significantly upregulated, and CLDN5, 7, 8, and 23 downregulated in primary tumor and hepatic metastasis samples compared with normal mucosa (). These results were validated in 15 matched samples from the study cohort (normal mucosa and primary tumor from the same patient; Figure S2). The largest variation of expression between paired samples was observed for CLDN2 and CLDN8.
Comparison of CLDN gene expression in the different CRC molecular subtypes
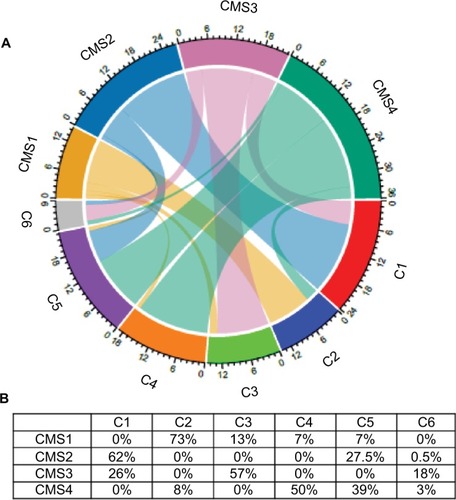
CLDN expression was then analyzed in all 143 primary CRC samples classified in molecular subtypes according to Marisa et al (C1 to C6) and the consensus classification (CMS1 to 4; see and for the relationships between classifications).Citation42
Figure 2 Interrelationships between the consensus classification (four subtypes: CMS1 to 4) and the Marisa’s classification (six molecular subtypes: C1 to C6) of the 143 CRC samples included in this study.
Notes: (A) Chord diagram to visualize the relationships among subtypes (in different colors) of the two CRC classifications arranged radially. Data are connected to each other using arcs; each connection is proportional to the arc size. (B) Percentage of CRC samples classified as C1 to C6 included in each CMS subtype.
Abbreviation: CRC, colorectal cancer.
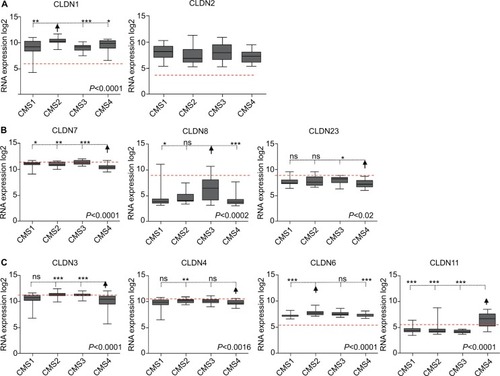
Among the CLDN genes that were upregulated in tumors compared with normal mucosa, CLDN1 expression was significantly higher in the CMS2 subtype and in the C1 and C5 subtypes, while CLDN2 expression was similar in all subtypes (Figure 3A and Figure S3A). Among the CLDN genes downregulated in tumors, CLDN8 expression reduction was less pronounced in the CMS3 and C6 subtypes, while CLDN7 and CLDN23 were more downregulated in the CMS4 and C4 subtypes ( and Figure S3Bb). Among the CLDN genes with comparable expression in normal and tumor tissues, CLDN3 and CLDN4 showed lower expressions in the CMS4 subtype (not significant for CLDN4) and in the C4 subtype ( and Figure S3C). Among the CLDN genes that were weakly expressed in primary tumors, CLDN6 expression level was higher in the CMS2 and C1 subtypes, and CLDN11 expression was strongest in the CMS4 and C4 subtypes ( and Figure S3C). The other CLDN genes did not show any significant expression level difference among CRC subtypes.
Figure 3 Differential CLDN expression in CRC samples classified using the CMS classification.
Notes: (A) CLDN genes upregulated in tumor samples compared with normal mucosa. (B) CLDN genes downregulated in tumors compared with normal mucosa. (C) CLDN genes with similar expression in normal mucosa and tumor samples. The red horizontal line indicates CLDN expression in normal mucosa within the CRC samples. The arrow highlights the subtype where CLDN expression is the most significantly different compared with the other subtypes (Kruskal–Wallis/Dunn’s test). *P<0.05; **P<0.01; ***P<0.001.
Abbreviation: CRC, colorectal cancer.
Association between CLDN expression level and OS and PFS
Then, the correlation between CLDN gene expression level and PFS and OS rates was evaluated in the 143 patients divided into two groups (high/low expression relative to the median CLDN gene expression) of the same size. This analysis found a significant correlation only between CLDN2 gene expression (low/high) and OS (P=0.03) and PFS (P=0.005), and between CLDN4 gene expression and OS (P=0.01; and ). These results were confirmed in an independent cohort of 80 patients (Tsuji’s cohort;Citation44 Figure S4A).
Figure 4 Clinical value of CLDN gene expression in the 143 CRC samples.
Notes: Association between CLDN2 and CLDN4 gene expression levels and (A) OS and (B) PFS (log-rank test). High and low: higher or lower expressions relative to the median expression for that CLDN gene. (C) Association of combined CLDN2 and CLDN4 gene expressions with OS.
Abbreviations: CRC, colorectal cancer; OS, overall survival; PFS, progression-free survival.
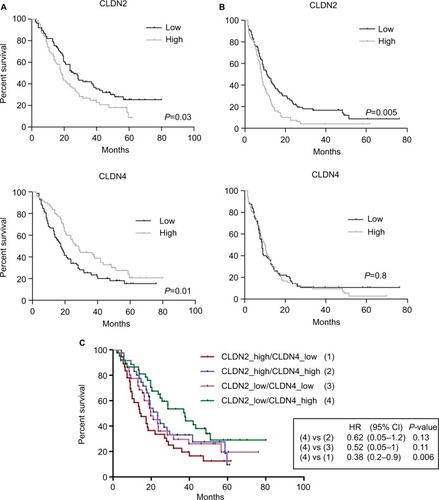
As low CLDN2 expression and high CLDN4 tumor expression were good prognostic factors in patients with CRC, the next step was to assess whether OS was correlated with the expression of both CLDNs. OS rate was significantly higher in patients with CRC displaying concomitant low CLDN2 and high CLDN4 expressions, particularly compared with patients with high CLDN2 and low CLDN4 tumor expressions (P=0.006; ). OS time was more than twofold longer in patients with low CLDN2/high CLDN4 tumors than in those with high CLDN2/low CLDN4 tumors (38 months vs 14.4 months). Moreover, analysis of the correlation between OS and CLDN2 and CLDN4 gene expressions according to the CRC molecular subtype confirmed the significant associations between CLDN2 and CLDN4 expressions and OS for the CMS3 and C3 and the CMS2 and C1 subtypes, respectively (Figure S4B).
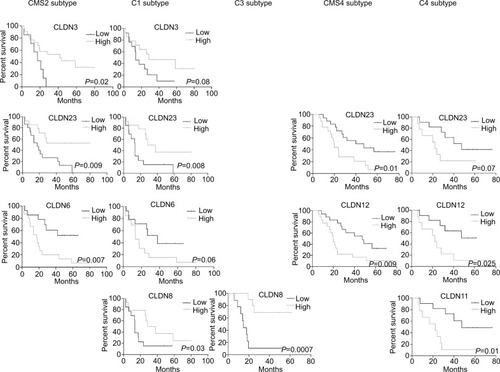
Similarly, high expressions of CLDN3 and CLDN23 and low expression of CLDN6 in the CMS2 subtype were associated with longer OS (). Low CLDN6 expression in the CMS2 subtype and high CLDN23 expression in the CMS2 and C1 subtypes were correlated with better PFS rates (Figure S5). In the CMS4 and C4 subtypes, low expression of CLDN12 and CLDN23 was associated with longer OS (). In the C4 subtype, low CLDN11 expression was correlated with longer OS () and also PFS (Figure S5). Finally, high CLDN8 expression was associated with longer OS in the C1 and C3 subtypes ().
Figure 5 Association of CLDN gene expression with OS in the different CRC subtypes.
Notes: Log-rank test. High/low: expression higher/lower than the median value for that CLDN gene.
Abbreviation: CRC, colorectal cancer; OS, overall survival.
Discussion
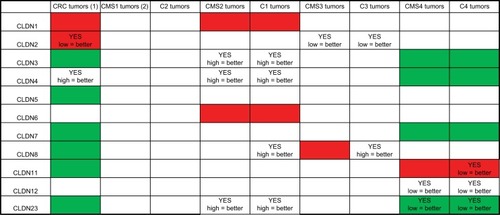
In this study, using Affymetrix gene expression data from 143 patients with CRC categorized according the CRC molecular subtypes,Citation42 we showed that CLDN gene expression profiles vary according to the tumor stage (normal tissue, primary tumor, and metastasis) and also the molecular subtype. We assessed CLDN gene expression in the four subtypes proposed by the consensus molecular classification,Citation39 and in the six subtypes defined by Marisa et alCitation36 to refine the tumor type identification. Our analysis (summary in ) indicated that eight CLDNs were differentially expressed in CRC tumors compared with normal mucosa. CLDN expression alterations were mainly seen in the CMS2/C1 and CMS4/C4 subtypes. The changes in the expression of these eight CLDN genes were associated with a prognostic value in the whole cohort of CRC samples and also in specific molecular subtypes. Conversely, we did not detect any differential expression or prognostic value of CLDN expression in the CMS1 and C2 subtypes. This could be due to the low number of tumors in these two groups (n=15 and 17, respectively).
Figure 6 Summary of CLDN gene expression and prognostic value in CRC.
Notes: (1) Expression in CRC samples (n=20) compared with normal mucosa samples (n=17); red = upregulated; green = downregulated; white, no expression difference with normal mucosa. (2) Comparison of CLDN expression in the different CRC subtypes; red = subtype where CLDN expression is strongest; green = subtype where the expression of that CLDN gene is lowest compared with the other subtypes; white = no expression difference among subtypes. YES indicates that the expression level of that CLDN gene shows prognostic value, with the correlation between CLDN expression level (high/low relative to the median expression of that gene) and improved prognosis.
Abbreviation: CRC, colorectal cancer.
CLDN gene expression in normal colon tissues is consistent with a previous study where the expression patterns were analyzed along the proximal–distal axis of the human intestine by real-time PCR.Citation45 In our study, CLDN3, 4, 7, 8, and 23 displayed the highest expressions in normal colon epithelial cells with a predominant expression of CLDN7. CLDN3, 4, and 7 are strongly expressed in several normal tissues, including colon, and their similar expression and localization profiles suggest a coordinated regulation.Citation46 These CLDNs have an important role in the maintenance of homeostasis of colon epithelium. CLDNs 3 and 4 have been classified as colon barrier proteins.Citation47,Citation48 CLDN 7 maintains the barrier function of the intestinal epithelium and regulates epithelial cell renewal.Citation49 CLDN 7 is localized at apical TJs and also in basolateral membranes, and CLDN 7-mediated cell–matrix interaction is indispensable in the intestine.Citation50 Except for CLDN4, their expression was downregulated in CRC samples. CLDN3 and CLDN7 downregulation was more pronounced in the mesenchymal CMS4 and in the stem cell phenotype-like C4 subtypes. CMS4 and C4 tumors are characterized by activation of pathways related to EMT and stemness.Citation36,Citation38,Citation39 Bhat et alCitation24 reported that CLDN7 has a key role in EMT regulation in colon epithelial cells, and that low CLDN7 expression promotes EMT and tumor progression. CLDN7 is also frequently associated with the stem cell marker EPCAM,Citation51 and this association could contribute to EMT.Citation52 CLDN8 displayed the strongest downregulation in CRC samples, but little is known about its function in the colon. CLDN8 was identified as a critical downstream component of the IL9 inflammatory cascade in inflammatory bowel disease.Citation53 Here, we found that in the C1 and C3 subtypes, CLDN8 expression has a prognostic value. The common features of these two subtypes are KRAS mutations and the suppression of pathways associated with activation of the immune system and EMT.Citation36 Moreover, higher expressions of CLDN3 and CLDN4 were associated with a better outcome in patients with CMS2 and C1 cancers. These findings are in agreement with CLDN3 and CLDN7 tumor-suppressive functions;Citation33,Citation54,Citation55 conversely, the possible tumor suppressor role of CLDN4 and CLDN8 remains to be demonstrated.
CLDN23 expression was significantly reduced in CRC samples, as previously reported in different intestinal cancer types,Citation56 including CRC.Citation17,Citation35,Citation57 Like CLDN3, CLDN4, and CLDN7, CLDN23 was only slightly downregulated in the CMS4 and C4 subtypes, and this was correlated with longer OS. We can hypothesize that in these subtypes, CLDN23 expression is regulated by stromal suppressor genes. Conversely, in the canonical CMS2 and C1 subtypes, patients with low CLDN23 tumor expression had shorter OS and PFS, in agreement with a previous analysis in 53 patients with CRC.Citation57 CLDN23 seems to have dual role as a tumor suppressor and a tumor promoter, depending on the CRC subtype where it is expressed. This hypothesis requires additional investigations because very few studies have assessed CLDN23’s role in CRC since its first characteriza tion following the observation that CLDN23 is downregulated in gastric cancer.Citation56
On the other hand, the expressions of CLDN1 and CLDN2 were significantly increased in tumor tissues, compared with normal mucosa. This confirms previous reports on their upregulation in CRC and their involvement in CRC cell tumorigenicity.Citation22,Citation28,Citation58,Citation59 Induction of CLDN1 and CLDN2 expressions has been related to overactivation of WNT/β-catenin signaling in CRC cells.Citation60,Citation61 Moreover, expressions of CLDN1 and CLDN2 have been associated with EMT and cancer progression.Citation25,Citation30 We previously showed that CLDN1 expression is higher in the canonical CMS2 subtype, which includes the C1 and C5 subtypes, and represents epithelial tumors with marked upregulation of WNT and MYC downstream targets, and that PFS is significantly longer in patients with C3 and C5 tumors with low CLDN1 expression.Citation28 Here, we found that CLDN2 level is not different among CRC subtypes, but has a strong prognostic value in all patients and also in the CMS3 and C3 subtypes, which often harbor KRAS-activating mutations. Our findings are in agreement with recent data showing that high CLDN2 expression is linked to posttreatment recurrence in patients with stage II/III CRC.Citation62 This makes CLDN2 a good candidate for therapeutic target in CRC.
Three other claudin genes (CLDN6, CLDN11, and CLDN12) also displayed a prognostic value. CLDN6 was more expressed in the CMS2 and C1 subtypes, and low expression in CMS2 was associated with longer OS and PFS. CLDN6 expression and function have never been studied in CRC. It was described as a cancer-promoting factor in gastric cancer,63 and as a tumor suppressor in breast cancer.64 Low CLDN11 expression correlation with longer OS and PFS in the C4 subtype (at high risk of relapse), and CLDN11 upregulation in the CMS4 and C4 subtypes suggest a pro-tumor function in CRC. CLDN11 could play a role in the TGFβ1-OCLN/CLDN11 paracrine axis between cancer cells and cancer-associated fibroblasts in CRC.65 CLDN12 was strongly expressed in both normal and tumor colon tissues. In the CMS4 and C4 subtypes, low expression was associated with better prognosis. CLDN12 contributes to Ca2+ absorption in intestinal epithelial cells,66 and it is one of the few claudins that do not possess a PDZ binding motif.Citation7
The specific biological features of the CRC subtypes could explain the differences in CLDN gene expression. Indeed, the gene expression profiles of claudins involved in EMT (CLDN7), acting as tumor suppressors (CLDN3, 7, 23) or linked to the stem cell phenotype (CLDN7), were markedly different in CMS4/C4 tumors (). In CMS2/C1 and C5 tumors, CLDN gene expression seemed to be more dependent on WNT signaling, as already described for CLDN1.
CLDN gene downregulation could be explained by alterations in their sequences or in epigenetic regulation mechanisms, such as histone modifications, DNA methylation, and chromatin remodeling. This was demonstrated for CLDN1,Citation16 CLDN2,Citation18 CLDN7,Citation19 and CLDN23.Citation17
Changes in CLDN gene expression were essentially seen in the CMS2 and CMS4 subtypes for which a clear distinction in sensitivity to chemotherapy-induced apoptosis has been reported.Citation67 This suggests that in CRC, claudins could play a role in chemoresistance, as previously shown for CLDN4 and CLDN7 in ovarian cancer,Citation68,Citation69 and more recently, for CLDN6 in triple-negative breast cancer cells.Citation70
Altogether, we showed that CLDN gene expression has a significant clinical relevance. First, in the whole sample, the expressions of CLDN2 and CLDN4 alone showed a strong prognostic value that was increased when both genes were associated (median survival: 14.41 months for patients with high CLDN2/low CLDN4 expression and 38 months for patients with low CLDN2/high CLDN4 expression; HR =0.38). If these expression data are confirmed at the protein level, a prognostic gene/protein expression score can be developed for CRC patients. Second, for almost all CRC subtypes, we identified at least one CLDN gene, the expression of which was correlated with survival (). In addition, we defined a CMS4/C4 tumor subtype signature characterized by low CLDN3, 4, 7, and 23 expressions associated with high CLDN11 expression. Moreover, in the C1 subtype, high CLDN3, 4, 8, and 23 expressions could be associated with good prognosis. In a breast cancer subtype, a signature characterized by low CLDN gene expression was associated with an aggressive phenotype.Citation71 Finally, the best candidates as therapeutic targets seem to be CLDN6 in CMS2 tumors and CLDN11 in C4 tumors because they are overexpressed in these subtypes and low expression was associated with better prognosis. Future studies should thoroughly investigate their value as prognostic markers or therapeutic targets.
Conclusion
CLDN gene expression differences in CRC reflect CRC heterogeneity, and indicate that it is not enough simply to examine CLDN expression level globally. Moreover, claudin functions are not limited to TJs. Indeed, growing evidence shows that claudins are localized to sites outside the TJ complex,Citation72 or even delocalized from the membrane to the nucleus, thus acquiring an important role in tumorigenesis. Finally, our findings demonstrated that CLDN expression is regulated in a CRC molecular subtype manner, and highlighted that to evaluate claudin function and potential value as prognostic markers or therapeutic targets, it is essential to take into account CRC heterogeneity.
Data sharing statement
Transcript profiles: GSE62080, GSE72970, GSE28702.
Acknowledgments
This work was supported by grants from the French National Research Agency under the program “Investissementsd’avenir”, Grant Agreement LabExMAbImprove: ANR-10-LABX-53, and SIRIC Montpellier Cancer Grant: “INCa-DGOS-Inserm 6045”.
Disclosure
The authors report no conflicts of interest in this work.
References
- RoyerCLuXEpithelial cell polarity: a major gatekeeper against cancer?Cell Death Differ20111891470147721617693
- AndersonJMVan ItallieCMPhysiology and function of the tight junctionCold Spring Harb Perspect Biol200912a00258420066090
- ColegioORVan ItallieCMMcCreaHJRahnerCAndersonJMClaudins create charge-selective channels in the paracellular pathway between epithelial cellsAm J Physiol Cell Physiol20022831C142C14712055082
- OshimaTMiwaHGastrointestinal mucosal barrier function and diseasesJ Gastroenterol201651876877827048502
- Van ItallieCMAndersonJMArchitecture of tight junctions and principles of molecular compositionSemin Cell Dev Biol20143615716525171873
- TsukitaSFuruseMPores in the wall: claudins constitute tight junction strands containing aqueous poresJ Cell Biol20001491131610747082
- GünzelDYuASClaudins and the modulation of tight junction permeabilityPhysiol Rev201393252556923589827
- Lal-NagMMorinPJThe claudinsGenome Biol200910823523519706201
- ChenJZhangMThe Par3/Par6/aPKC complex and epithelial cell polarityExp Cell Res2013319101357136423535009
- TsukitaSFuruseMItohMMultifunctional strands in tight junctionsNat Rev Mol Cell Biol20012428529311283726
- SinghABSharmaADhawanPClaudin family of proteins and cancer: an overviewJ Oncol2010201054195720671913
- Van ItallieCMAndersonJMClaudin interactions in and out of the tight junctionTissue Barriers201313e2524724665401
- OsanaiMTakasawaAMurataMSawadaNClaudins in cancer: bench to bedsidePflugers Arch20174691556727624415
- SinghABUppadaSBDhawanPClaudin proteins, outside-in signaling, and carcinogenesisPflugers Arch20174691697527988840
- Garcia-HernandezVQuirosMNusratAIntestinal epithelial claudins: expression and regulation in homeostasis and inflammationAnn N Y Acad Sci201713971667928493289
- KrishnanMSinghABSmithJJHDAC inhibitors regulate claudin-1 expression in colon cancer cells through modulation of mRNA stabilityOncogene201029230531219881542
- MaryanNStatkiewiczMMikulaMRegulation of the expression of claudin 23 by the enhancer of zeste 2 polycomb group protein in colorectal cancerMol Med Rep201512172873625695204
- AhmadRKumarBPanKDhawanPSinghABHDAC-4 regulates claudin-2 expression in EGFR-ERK1/2 dependent manner to regulate colonic epithelial cell differentiationOncotarget2017850877188773629152115
- NakayamaFSembaSUsamiYChibaHSawadaNYokozakiHHypermethylation-modulated downregulation of claudin-7 expression promotes the progression of colorectal carcinomaPathobiology200875317718518550915
- TabarièsSSiegelPMThe role of claudins in cancer metastasisOncogene20173691176119027524421
- ChaoYCPanSHYangSCClaudin-1 is a metastasis suppressor and correlates with clinical outcome in lung adenocarcinomaAm J Respir Crit Care Med2009179212313318787218
- DhawanPSinghABDeaneNGClaudin-1 regulates cellular transformation and metastatic behavior in colon cancerJ Clin Invest200511571765177615965503
- JoyceTCantarellaDIsellaCMedicoEPintzasAA molecular signature for epithelial to mesenchymal transition in a human colon cancer cell system is revealed by large-scale microarray analysisClin Exp Metastasis200926656958719340593
- BhatAAPopeJLSmithJJClaudin-7 expression induces mesenchymal to epithelial transformation (MET) to inhibit colon tumorigenesisOncogene201534354570458025500541
- BhatAAAhmadRUppadaSBSinghABDhawanPClaudin-1 promotes TNF-α-induced epithelial-mesenchymal transition and migration in colorectal adenocarcinoma cellsExp Cell Res2016349111912727742576
- SinghABDhawanPClaudins and cancer: fall of the soldiers entrusted to protect the gate and keep the barrier intactSemin Cell Dev Biol201542586526025580
- JiangLYangLHuangHLiuBYZuGPrognostic and clinical significance of claudin-1 in colorectal cancer: a systemic review and meta-analysisInt J Surg20173921422028192244
- CherradiSAyrolles-TorroAVezzo-ViéNAntibody targeting of claudin-1 as a potential colorectal cancer therapyJ Exp Clin Cancer Res20173618928659146
- OubanAClaudin-1 role in colon cancer: an update and a reviewHistol Histopathol201833101013101929504115
- DhawanPAhmadRChaturvediRClaudin-2 expression increases tumorigenicity of colon cancer cells: role of epidermal growth factor receptor activationOncogene201130293234324721383692
- BuchertMPapinMBonnansCSymplekin promotes tumorigenicity by up-regulating claudin-2 expressionProc Natl Acad Sci U S A201010762628263320133805
- DingLWangLSuiLClaudin-7 indirectly regulates the integrin/FAK signaling pathway in human colon cancer tissueJ Hum Genet201661871172027121327
- DaridoCBuchertMPannequinJDefective claudin-7 regulation by Tcf-4 and Sox-9 disrupts the polarity and increases the tumorigenicity of colorectal cancer cellsCancer Res200868114258426818519685
- WangXTullyONgoBZitinMMullinJMEpithelial tight junctional changes in colorectal cancer tissuesScientificWorldJournal20111182684121479352
- BujkoMKoberPMikulaMLigajMOstrowskiJSiedleckiJAExpression changes of cell-cell adhesion-related genes in colorectal tumorsOncol Lett2015962463247026137091
- MarisaLde ReynièsADuvalAGene expression classification of colon cancer into molecular subtypes: characterization, validation, and prognostic valuePLoS Med2013105e100145323700391
- SadanandamALyssiotisCAHomicskoKA colorectal cancer classification system that associates cellular phenotype and responses to therapyNat Med201319561962523584089
- DienstmannRVermeulenLGuinneyJKopetzSTejparSTaberneroJConsensus molecular subtypes and the evolution of precision medicine in colorectal cancerNat Rev Cancer2017172799228050011
- GuinneyJDienstmannRWangXThe consensus molecular subtypes of colorectal cancerNat Med201521111350135626457759
- Del RioMMolinaFBascoul-MolleviCGene expression signature in advanced colorectal cancer patients select drugs and response for the use of leucovorin, fluorouracil, and irinotecanJ Clin Oncol200725777378017327601
- Del RioMMolleviCVezzio-VieNBibeauFYchouMMartineauPSpecific extracellular matrix remodeling signature of colon hepatic metastasesPLoS One201389e7459924023955
- Del RioMMolleviCBibeauFMolecular subtypes of metastatic colorectal cancer are associated with patient response to irinotecan-based therapiesEur J Cancer201776687528284171
- CopoisVBibeauFBascoul-MolleviCImpact of RNA degradation on gene expression profiles: assessment of different methods to reliably determine RNA qualityJ Biotechnol2007127454955916945445
- TsujiSMidorikawaYTakahashiTPotential responders to FOLFOX therapy for colorectal cancer by Random Forests analysisBr J Cancer2012106112613222095227
- LamerisALHuybersSKaukinenKExpression profiling of claudins in the human gastrointestinal tract in health and during inflammatory bowel diseaseScand J Gastroenterol2013481586923205909
- HewittKJAgarwalRMorinPJThe claudin gene family: expression in normal and neoplastic tissuesBMC Cancer2006618616836752
- MilatzSKrugSMRosenthalRClaudin-3 acts as a sealing component of the tight junction for ions of either charge and uncharged solutesBiochim Biophys Acta179820101120482057
- HeringNAAndresSFrommATransforming growth factor-β, a whey protein component, strengthens the intestinal barrier by upregulating claudin-4 in HT-29/B6 cellsJ Nutr2011141578378921430244
- TanakaHTakechiMKiyonariHShioiGTamuraATsukitaSIntestinal deletion of claudin-7 enhances paracellular organic solute flux and initiates colonic inflammation in miceGut201564101529153825691495
- DingLLuZForemanOInflammation and disruption of the mucosal architecture in claudin-7-deficient miceGastroenterology2012142230531522044670
- KuhnSKochMNübelTA complex of EpCAM, claudin-7, CD44 variant isoforms, and tetraspanins promotes colorectal cancer progressionMol Cancer Res20075655356717579117
- PhilipRHeilerSMuWBüchlerMWZöllerMThumaFClaudin-7 promotes the epithelial-mesenchymal transition in human colorectal cancerOncotarget2015642046206325514462
- LiLHuangSWangHCytokine IL9 triggers the pathogenesis of inflammatory bowel disease through the miR21-CLDN8 pathwayInflamm Bowel Dis201824102211222329788092
- AhmadRKumarBChenZLoss of claudin-3 expression induces IL6/gp130/Stat3 signaling to promote colon cancer malignancy by hyperactivating Wnt/β-catenin signalingOncogene201736476592660428783170
- OshimaTKunisakiCYoshiharaKReduced expression of the claudin-7 gene correlates with venous invasion and liver metastasis in colorectal cancerOncol Rep200819495395918357381
- KatohMKatohMCLDN23 gene, frequently down-regulated in intestinal-type gastric cancer, is a novel member of CLAUDIN gene familyInt J Mol Med200311668368912736707
- PitulePVycitalOBruhaJDifferential expression and prognostic role of selected genes in colorectal cancer patientsAnticancer Res201333114855486524222123
- KinugasaTHuoQHigashiDSelective up-regulation of claudin-1 and claudin-2 in colorectal cancerAnticancer Res2007276A3729373417970035
- AungPPMitaniYSanadaYNakayamaHMatsusakiKYasuiWDifferential expression of claudin-2 in normal human tissues and gastrointestinal carcinomasVirchows Arch2006448442843416328347
- MiwaNFuruseMTsukitaSNiikawaNNakamuraYFurukawaYInvolvement of claudin-1 in the beta-catenin/Tcf signaling pathway and its frequent upregulation in human colorectal cancersOncol Res20011211–1246947611939410
- MankertzJHillenbrandBTavalaliSHuberOFrommMSchulzkeJDFunctional crosstalk between Wnt signaling and Cdx-related transcriptional activation in the regulation of the claudin-2 promoter activityBiochem Biophys Res Commun200431441001100714751232
- Paquet-FifieldSKohSLChengLTight junction protein clau-din-2 promotes self-renewal of human colorectal cancer stem-like cellsCancer Res201878112925293829510994
- Zavala-ZendejasVETorres-MartinezACSalas-MoralesBFortoulTIMontañoLFRendon-HuertaEPClaudin-6, 7, or 9 overexpression in the human gastric adenocarcinoma cell line AGS increases its invasiveness, migration, and proliferation rateCancer Invest201129111120874001
- WuQLiuYRenYTight junction protein, claudin-6, downregulates the malignant phenotype of breast carcinomaEur J Cancer Prev201019318619420215972
- KaragiannisGSSchaefferDFChoCKCollective migration of cancer-associated fibroblasts is enhanced by overexpression of tight junction-associated proteins claudin-11 and occludinMol Oncol20148217819524268521
- FujitaHSugimotoKInatomiSTight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytesMol Biol Cell20081951912192118287530
- LinnekampJFHooffSRVPrasetyantiPRConsensus molecular subtypes of colorectal cancer are recapitulated in in vitro and in vivo modelsCell Death Differ201825361663329305587
- YoshidaHSumiTZhiXYasuiTHondaKIshikoOClaudin-4: a potential therapeutic target in chemotherapy-resistant ovarian cancerAnticancer Res20113141271127721508375
- KimCJLeeJWChoiJJHigh claudin-7 expression is associated with a poor response to platinum-based chemotherapy in epithelial ovarian carcinomaEur J Cancer201147691892521134740
- YangMLiYRuanYCLDN6 enhances chemoresistance to ADM via AF-6/ERKs pathway in TNBC cell line MDAMB231Mol Cell Biochem20184431–216918029159771
- HerschkowitzJISiminKWeigmanVJIdentification of conserved gene expression features between murine mammary carcinoma models and human breast tumorsGenome Biol200785R7617493263
- HagenSJNon-canonical functions of claudin proteins: beyond the regulation of cell-cell adhesionsTissue Barriers201752e132783928548895