
Abstract
Purpose:
Numerous studies have shown that the expression of microRNA-181c (miR-181c) is inhibited in various cancers, which suggests that it has a cancer suppressive effect. In the current study, we evaluated the regulation and characteristics of miR-181c in human hepatocellular carcinoma (HCC).
Materials and methods:
Samples of tumor tissues and adjacent non-tumor tissues were collected from 52 patients with HCC, and expression levels of miR-181c in these samples were investigated via quantitative real-time polymerase chain reaction. HCC cell migration and invasion were investigated via wound healing assays and transwell assays. HCC cell apoptosis rates were assessed via flow cytometry, and HCC proliferation was assessed via 5-ethynyl-20-deoxyuridine assays. In vivo tumors were initiated by subcutaneously inoculating HCC cells into nude mice. And various biomarkers were investigated via western blotting.
Results:
In microarray datasets and tumor tissues, significant downregulation of miR-181c was apparent compared with non-tumorous adjacent tissues. Expression of miR-181c in HCC cells was also significantly lower than it was in normal human liver cells. miR-181c regulated the migration, invasion, apoptosis, and proliferation of HCC cell lines in vitro, and tumor development in vivo. Observations also suggest that miR-181c regulates NCAPG in HCC cells, and its expression affects cellular invasion, migration, proliferation, and apoptosis. There was a negative correlation between miR-181c expression and NCAPG in HCC tissue samples.
Conclusion:
miR-181c exhibits tumor-suppression via the regulation of NCAPG levels.
Introduction
Liver cancer is a fatal disease, and according to the latest statistics it is the sixth most common cancer and the fourth highest cause of cancer death worldwide. Among liver cancers, hepatocellular carcinoma (HCC) is the most common and reportedly accounts for 75–85% of primary liver cancers.Citation1 The main treatment strategies for patients with HCC are currently surgical resection and liver transplantation, but most HCC patients are diagnosed at an advanced stage, and thus the clinical efficacy of these treatments and patient prognoses are not satisfactory.Citation2,Citation3 Therefore, it is important to elucidate the molecular mechanisms involved in the progression of HCC and to identify new HCC biomarkers and therapeutic targets.
Micro-RNAs (miRNAs) are small non-coding RNA molecules that contain approximately 22 nucleotides and play a role in RNA silencing and post-transcriptional regulation of gene expression.Citation4–Citation6 miRNAs act via base pairing with complementary sequences in mRNA molecules.Citation7 They play an important role in a variety of HCC biological and pathological processes.Citation8–Citation11 Several studies have reported abnormal expression of miRNA-181c (miR-181c) in a variety of tumors including endometrial carcinoma, prostate cancer, glioma, and ovarian cancer, and it is reportedly involved in multiple processes associated with tumor progression.Citation12–Citation17 However, the characteristics of miR-181c in HCC have not yet been elucidated.
NCAPG is a condensed protein complex subunit responsible for the concentration and stabilization of chromosomes during mitosis and meiosis.Citation18 Protein phosphorylation activates the condensin complex.Citation19 In recent years there has been an increase in the number of studies reporting abnormal expression of NCAPG in various tumors, including prostate cancer, glioma, and lung adenocarcinoma, and NCAPG is evidently involved in multiple biological functions.Citation20–Citation22 Moreover, in previous studies knockdown of NCAPG affected HCC cell cycle, apoptosis, and metastasis.Citation23 However, the relationship between NCAPG and miRNAs in HCC has not been elucidated.
The aims of the current study were to investigate the roles of miR-181c and NCAPG in HCC.
Materials and methods
Tissue specimens and microarray data
HCC and histologically confirmed paracancerous tissue specimens were collected from 52 patients at the Second Affiliated Hospital of Nanchang University in China. The patients did not undergo radiotherapy, chemotherapy, or immunotherapy before surgery. Immediately after the removal of tissue specimens during surgery, HCC and paracancerous tissue samples were placed in liquid nitrogen and stored. The study was conducted in accordance with the Helsinki Declaration of 1964 and all subsequent amendments, it was approved by the Ethics Committee of the Second Affiliated Hospital of Nanchang University in China, and all patients provided written informed consent. NCAPG mRNA expression levels of 225 HCC samples and 220 corresponding matched normal samples were downloaded from the Oncomine (Roessler Liver 2) database.Citation24 The miR-181c expression levels of 370 HCC samples and 50 corresponding matched normal samples of were downloaded from The Cancer Genome Atlas (TCGA) database. The Log2 median center intensity ratio was used. Differences in expression between HCC and normal liver tissues were analyzed via Student’s t-tests.
Cell lines and cell culture
The normal human liver cell line HL-7702 and two HCC cell lines, SMMC-7721 and MHCC-97H, were purchased from the Shanghai Institute of Cell Biology (Shanghai, China). HL-7702 cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS; Biological Industries, Israel). SMMC-7721 and MHCC-97H cells were cultured in high glucose Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS. All cells were maintained at 37 ℃ in a cell culture incubator with 5% CO2 and 95% humidity. Cells in log phase were used in all experiments.
Cell transfection
NCAPG small interfering RNA (siRNA) and a negative control (NC) were purchased from GenePharma (Shanghai, China). NCAPG overexpression plasmid and control vectors were purchased from GeneChem (Shanghai, China). miR-181c mimics and inhibitors and NC miRNA were purchased from RiboBio (Guangzhou, China). SMMC-7721 and MHCC-97H cell lines were assigned to the NC and treatment groups. Transfection was performed using Lipofectamine 3000 (Invitrogen, Thermo Fisher Scientific, Inc., USA) in accordance with the manufacturer’s instructions. NCAPG siRNA sequences were transfected into the cells, and the sequences were as follows: NCAPG‑s1 sense, 5’‑GGAGUUCAUUCAUUACCUUT T ‑3’ and antisense, 5’‑AAGGUAAUGAAUGAACUCCTT‑3’.
Quantitative real-time polymerase chain reaction and western blotting
Quantitative real-time polymerase chain reaction (qRT-PCR) and western blotting were performed as previously described.Citation25 The relative expression of each gene was normalized to that of the housekeeping gene and calculated using the 2−ΔΔCt method for in vitro studies and the 2−ΔCt method for tissue specimens.Citation26,Citation27 Primers for miR-181c and U6 were designed by RiboBio. The primer sequences of glyceraldehyde 3-phosphate dehydrogenase and NCAPG were purchased from GenScript Biotechnology (Nanjing, China). The primary antibodies, NCAPG (ab226805), Bax (ab32503), Bcl-2 (ab32124), α-tubulin (ab15246), N-cadherin (ab76011), and E-cadherin (ab40772) were purchased from Abcam (Cambridge, UK). Three independent experiments were performed.
Cell wound healing assay
Wound healing assays were performed to assess cell migration. SMMC-7721 and MHCC-97H cells were seeded into 6-well plates and incubated at 37 °C in DMEM containing 10% FBS. After the cells reached ~90% confluence, an artificial wound was created in each well by scraping a 200-μL pipette tip over the cell surface. The wells were then washed with phosphate-buffered saline to remove separated cells, then incubated with fresh complete medium without FBS for 24 hrs. After 24 hrs, the cell migration distance from the edge of the scratch to the center was measured.
Transwell assay
Transwell chambers (Corning Inc., Corning, USA) with 8-μm pores were used to investigate cell invasion and migration. Twenty-four hours after transfection, the cells were resuspended in serum-free medium and inoculated separately into upper chambers with and without Matrigel (10 mg/mL; BD Biosciences, USA). Medium containing 10% FBS was added to the lower chambers. After 24 hrs of cell migration and invasion the cells remaining on the upper surface of the membrane were removed, fixed with polyoxymethylene, stained with 0.1% crystal violet, then photographed under a microscope and counted.
Measurement of apoptosis via flow cytometry
The annexin V‑fluorescein isothiocyanate/propidium iodide apoptosis detection kit (BD Biosciences) was used to assess apoptosis via fluorescence-activated cell sorting, in accordance with the manufacturer’s instructions.
5-ethynyl-20-deoxyuridine assay
Cell proliferation was measured via the 5-ethynyl-20-deoxyuridine (EdU) assay kit (RiboBio) in accordance with the manufacturer’s instructions, and observations were performed with a fluorescence microscope. The percentage of EdU-positive cells relative to the total number of cells was used as the proliferation index.
Tumorigenicity assay
In the in vivo tumorigenicity assays, 1×107 cells in 100 μL of phosphate-buffered saline were subcutaneously injected into the flanks of nude mice (Hunan SJA Laboratory Animal Co., Ltd., Hunan, China). Tumor size was measured at 3-day intervals using a caliper and the following formula: Tumor volume = (shortest diameter2× longest diameter)/2. Four weeks later photographs were taken, mice were anesthetized, and tumors were harvested and weighed. For evaluating lung metastasis, 1×106 cells in 100 μL of phosphate-buffered saline were injected into the tail vein of nude mice. Mice were sacrificed 3 weeks after tumor injection. The lungs were then processed and embedded in paraffin. All animal experiments were approved by the Animal Experimental Ethics Committee of the Second Affiliated Hospital of Nanchang University and was performed in accordance with the “Guide for the Care and Use of Laboratory Animals” (revised 1985).
Hematoxylin & eosin and immunohistochemical staining
After the tissues were fixed in 10% formalin solution, we embedded them in paraffin blocks and then cut the tissue into paraffin sections. Then, the tissue sections were deparaffinized. Some of the deparaffinized sections were stained with hematoxylin & eosin (H&E), which was used to detect morphologic changes, and some were rehydrated and microwave-heated in sodium citrate buffer (10 mmol/L, pH 6.0) for antigen retrieval. The sections were then incubated with 0.3% hydrogen peroxide/phosphate-buffered saline for 30 mins and blocked with Super Blocking solution (Pierce). Subsequently, the tissue sections were incubated with NCAPG antibody at a 5 μg/mL (ab56382; Abcam, Cambridge, UK) at 4 °C overnight. The sections were washed with phosphate‑buffered saline (PBS) 3 times at 5‑min intervals. Then, they were labeled by EnVision HRP kits (DAKO) at room temperature for 30 mins. Next, the sections were stained with diaminobenzidine (DAB) and hematoxylin dyes and were sealed with neutral resins. All of the sections were observed and photographed with a light microscope with a DP70 CCD system (Olympus Corp.).
Dual luciferase reporter gene assay
The Dual‑Luciferase reporter gene assay was performed to ascertain whether NCAPG was the direct target gene of miR‑181c. The NCAPG 3’UTR dual luciferase reporter plasmids (WT and MUT) were constructed by RiboBio. They were co‑transfected using Lipofectamine 3000 (Invitrogen, Thermo Fisher Scientific, Inc., USA) with miR‑181c mimics or Mimic control into 293T cells, respectively. After incubation for 48 h, the luciferase activities were detected using a Dual‑Luciferase Assay System (Promega Corp., Madison, WI, USA).
Statistical analysis
GraphPad Prism 7.0 and SPSS 22.0 software were used for statistical analysis. Results are expressed as mean ± the standard deviation. Separate experiments were performed in triplicate. Bivariate correlations were calculated via the Spearman’s rank correlation coefficient. Comparisons between two groups were analyzed via the Student’s t-test. Multiple sets of comparisons were performed via one-way analysis of variance. Test results were considered significant if p<0.05.
Results
Correlations between miR-181c and NCAPG expression
To investigate expression levels of NCAPG and miR-181c in HCC tissues, and correlations between them, we assessed expression profiles via two databases and performed qRT-PCR in 52 HCC tissues and adjacent non-tumor tissues. Using data derived from Oncomine (Roessler Liver 2), NCAPG levels were compared in 225 HCC tissues and 220 normal liver tissues, and NCAPG was significantly upregulated in HCC tissues (p<0.0001; ). In qRT-PCR, the mean fold change in NCAPG mRNA expression in tumor tissues was significantly upregulated relative to that of paired non-tumor tissues (p<0.001; ). We then analyzed data derived from the TCGA database, including data from 370 HCC patients and 50 normal liver tissues. HCC tissues tended to express lower levels of miR-181c than normal liver tissues (p<0.05; ). In qRT-PCR analyses, miR-181c was significantly decreased in tumor tissues compared with adjacent non-tumor tissues (p<0.01; ). In HCC tissues, NCAPG expression levels were inversely correlated with miR-181c levels (two-tailed Spearman’s correlation, r = −0.496, p<0.001; ).
Figure 1 In microarray datasets and clinical HCC tissues, miR-181c expression levels were negatively correlated with NCAPG expression levels. (A) Levels of NCAPG mRNA expression in 225 HCC tissues and 220 normal liver tissues were obtained from the Oncomine database. (B) NCAPG mRNA expression was determined in 52 HCC tumor tissues and adjacent non‑tumor tissues via qRT-PCR. (C) Levels of miR-181c expression in 370 HCC patients and 50 normal liver tissues were determined via the TCGA database. (D) Expression levels of miR-181c in 52 HCC tumor tissues and adjacent non‑tumor tissues were detected via qRT-PCR. (E) Scatter-plots revealing a negative correlation between NCAPG mRNA and miR‑181c at the RNA level in 52 HCCs (r = ‑0.496, p<0.001). *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviations: HCC, hepatocellular carcinoma; TCGA, The Cancer Genome Atlas.
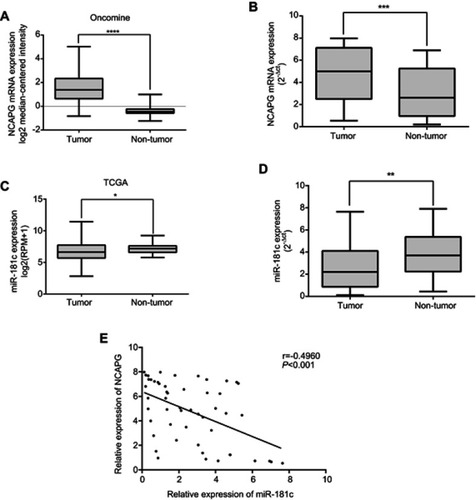
miR‑181c and the regulation of HCC cell migration and invasion in vivo and in vitro
The effects of miR-181c regulation on HCC cell invasion and migration were consistent with the regulation of NCAPG (). Wound healing assays were performed to detect changes in HCC cell migration after transfection with miR-181c mimics and miR-181c inhibitors. After miR-181c upregulation HCC cell migration was reduced, and this was consistent with the effects of NCAPG knockdown (). After miR-181c downregulation HCC cell migration was increased, and this was consistent with the overexpression of NCAPG (). In transwell migration and invasion assays HCC cells exhibited reduced migration and invasion capacity after upregulation of miR-181c and after NCAPG knockdown (). Downregulation of miR-181c and overexpression of NCAPG were associated with increased HCC cell invasion and migration capacity (). To further explore the effects of miR-181c we analyzed the metastasis‑related proteins E cadherin and N cadherin via western blotting. After transfection with miR-181c mimics or NCAPG siRNA, E-cadherin was upregulated and N-cadherin was downregulated (). After transfection with miR-181c inhibitors or vector-NCAPG, E-cadherin expression was downregulated and N-cadherin expression was upregulated (). The dose-dependent effect experiment further demonstrated that miR-181c can regulate the invasion and migration ability of HCC cells (–). Similar to the in vitro results, miR-181c mimic reduced lung metastasis in nude mice compared to the control group (). Compared with the control group, miR-181c inhibitor increased lung metastasis in nude mice ().
Figure 2 miR‑181c is able to regulate migration and invasion in HCC cells. (A) Wound healing assays were used to detect cell migration after transfection with miR-181c mimics and NCAPG siRNA. (B) Wound healing assays were used to detect cell migration after transfection with miR-181c inhibitors and vector-NCAPG. (C) Transwell assays were used to investigate cell migration and invasion after transfection with miR-181c mimics and NCAPG siRNA. (D) Transwell assays were used to investigate cell migration and invasion after transfection with miR-181c inhibitors and vector-NCAPG. (E) Western blotting analysis of E-cadherin and N-cadherin expression after transfection with miR-181c mimics and NCAPG siRNA. (F) Western blotting analysis of E-cadherin and N-cadherin expression after transfection with miR-181c inhibitors and vector-NCAPG. (G) Dose-dependent effect experiment of miR-181c mimic (left) and NCAPG siRNA (right). (H) Dose-dependent effect experiment of miR-181c inhibitor (left) and Vector-NCAPG (right). (I) Representative H&E–stained sections of the lung tissues collected from Mimic control and miR-181c mimic groups are shown in the left. The incidence of lung metastases in the two groups of nude mice is shown in the right. (J) Representative H&E–stained sections of the lung tissues collected from Inhibitor control and miR-181c inhibitor groups are shown in the left. The incidence of lung metastases in the two groups of nude mice is shown in the right. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviation: HCC, hepatocellular carcinoma.
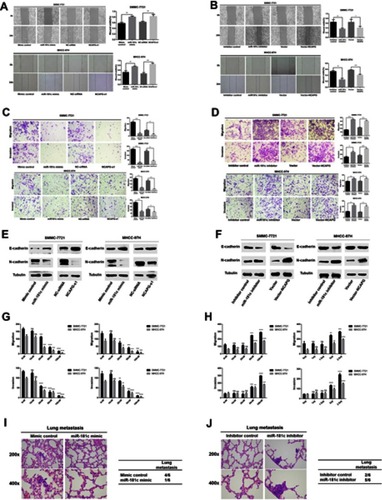
miR‑181c regulation of HCC cell apoptosis
The effects of miR-181c on HCC cell apoptosis were consistent with the regulation of NCAPG (). Flow cytometry was performed to examine changes in rates of apoptosis in HCC cells treated with miR-181c mimics and NCAPG siRNA. Rates of apoptosis in the miR-181C mimics group and the NCAPG siRNA group were higher than they were in the control group (). However, the rates of apoptosis in HCC cells treated with miR-181c inhibitors and in HCC cells treated with vector-NCAPG were lower than the rates of apoptosis in the control group (). Expression levels of apoptosis-associated proteins indicated abnormal alteration in the miR-181c mimics and NCAPG siRNA transfection groups, including increased expression of Bax and decreased expression of Bcl-2 (). After transfection with miR-181c inhibitors or vector-NCAPG, Bax expression was downregulated and Bcl-2 expression was upregulated ().
Figure 3 miR‑181c is able to regulate apoptosis in HCC cells. (A) Apoptosis rates in HCC cells transfected with miR-181c mimics and NCAPG siRNA. (B) Apoptosis rates in HCC cells transfected with miR-181c inhibitors and vector-NCAPG. (C) Western blotting analysis of Bax and Bcl-2 expression after transfection with miR-181c mimics and NCAPG siRNA. (D) Western blotting analysis of Bax and Bcl-2 expression after transfection with miR-181c inhibitors and vector-NCAPG. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviation: HCC, hepatocellular carcinoma.
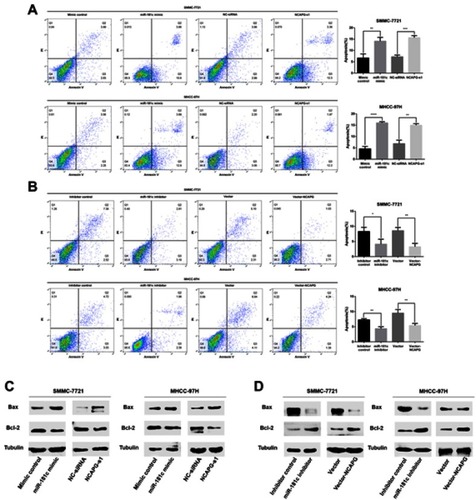
miR-181c regulation of HCC cell proliferation in vivo and in vitro
The EdU proliferation assay was used to study the effects of miR-181c on cell proliferation. HCC cell growth was significantly reduced in the miR-181c mimics group and the NCAPG siRNA group (), but in the miR-181c inhibitors group and the vector-NCAPG group it was significantly increased (). In in vivo experiments tumors formed by miR-181c mimic-treated SMMC-7721 cells were smaller and weighed less than tumors formed by control cells (). Tumors formed by SMMC-7721 cells transfected with miR-181c inhibitors were larger and weighed more than tumors formed by control cells (). Compared with the control group, the miR-181c mimic group showed a significant increase in the expression level of miR-181c and a decrease in the expression level of NCAPG protein (). Compared with the control group, the miR-181c inhibitor group showed a significant decrease in the expression level of miR-181c and an increase in the expression level of NCAPG protein ().
Figure 4 miR-181c regulates the proliferation of HCC cells in vivo and in vitro. (A) EdU proliferation assays were used to detect cell proliferation after transfection with miR-181c mimics and NCAPG siRNA. (B) EdU proliferation assays were used to detect cell proliferation after transfection with miR-181c inhibitors and vector-NCAPG. (C) Tumor weights and volumes in mice injected with miR-181c mimic-transfected SMMC-7721 cells. (D) Tumor weights and volumes in mice injected with miR-181c inhibitor-transfected SMMC-7721 cells. (E) qRT-PCR, western blot and immunohistochemical analysis of miR-181c mimic group and control group mouse xenografts. (F) qRT-PCR, western blot and immunohistochemical analysis of miR-181c inhibitor group and control group mouse xenografts. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviation: HCC, hepatocellular carcinoma.
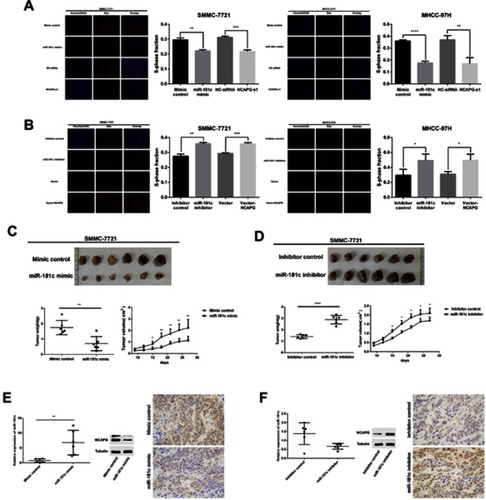
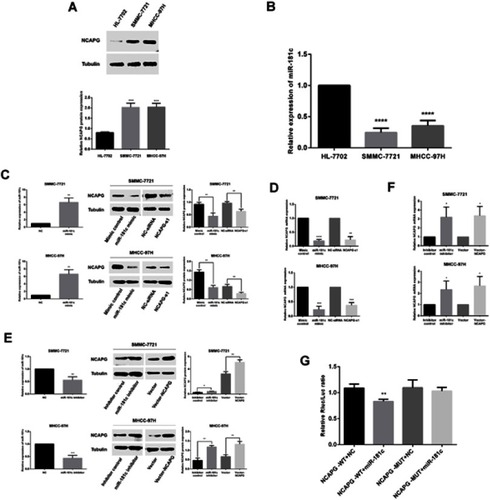
miR-181c regulation of NCAPG in HCC cells
To investigate interactions involving NCAPG and miR-181c in HCC, we first examined the protein expression levels of NCAPG in normal hepatocytes (HL-7702) and two HCC cell lines (SMMC-7721 and MHCC-97H) via western blotting. NCAPG protein expression in HCC cells was higher than it was in normal liver cell lines (p<0.001; ). Expression levels of miR-181c in two HCC cell lines (SMMC-7721 and MHCC-97H) and normal hepatocytes (HL-7702) were investigated via qRT-PCR. Levels of miR-181c expression were lower in HCC cell lines than in HL-7702 cells (p<0.0001; ). Therefore, SMMC-7721 and MHCC-97H cells were selected for subsequent assays. Satisfactory transfection efficiency was confirmed via qRT-PCR performed 36 hrs after transfection with miR-181c mimics, and qRT-PCR and western blotting analysis revealed that NCAPG mRNA and protein expression were reduced; which was consistent with the effects of NCAPG knockdown 48 hrs after transfection ( and ). Satisfactory transfection efficiency was apparent via qRT-PCR at 36 hrs after transfection with miR-181c inhibitors, and in qRT-PCR and western blotting analysis NCAPG mRNA and protein expression were increased, which was consistent with the overexpression of NCAPG 48 hrs after transfection ( and ). Subsequently, we performed a dual luciferase reporter gene assay to confirm the regulation relationship between miR‑181c and NCAPG. The results revealed that the miR‑181c mimics could reduce the luciferase activities of the wild‑type NCAPG reporter vector (NCAPG‑WT) rather than the mutant‑type NCAPG reporter vector (NCAPG‑MUT) (), indicating that NCAPG was the target gene of miR‑181c.
Figure 5 miR-181c regulates NCAPG in HCC cells. (A) Western blotting analysis of NCAPG expression in the normal human hepatocyte cell line HL‑7702, and two HCC cell lines MHCC-97h and SMMC-7721. (B) miR‑181c levels in the HL‑7702, MHCC-97h, and SMMC-7721 cell lines. (C) NCAPG protein expression in HCC cells detected by western blotting after transfection with miR-181c mimics and NCAPG siRNA. (D) mRNA expression in HCC cells detected by qRT-PCR after transfection with miR-181c mimics and NCAPG siRNA. (E) NCAPG protein expression in HCC cells was detected via western blotting after transfection with miR-181c inhibitors and vector-NCAPG. (F) HCC cell mRNA expression was detected via qRT-PCR after transfection with miR-181c inhibitors and vector-NCAPG. (G) Dual‑Luciferase reporter gene assay revealed that miR‑181c mimics could reduce the luciferase activities of NCAPG‑WT reporter vector rather than NCAPG‑MUT, indicating that NCAPG was the targeting gene of miR‑181c. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviation: HCC, hepatocellular carcinoma.
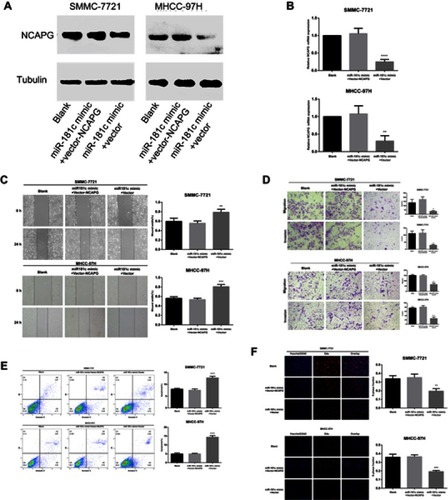
Involvement of NCAPG in miR-181c-mediated effect of cell invasion, migration, proliferation, and apoptosis
After confirming that NCAPG protein and mRNA expression were inhibited by miR-181c mimics, HCC cells were co-transfected with miR-181c mimics and vector-NCAPG. As shown in and , Overexpression of NCAPG restored the low NCAPG levels observed after transfection with miR-181c mimics alone. We also performed a series of rescue experiments involving simultaneous transfection of HCC cells with miR-181c mimics and vector-NCAPG. As shown in –, NCAPG significantly enhanced the invasion, migration, and proliferation capacities that had been suppressed by the miR-181c mimics, and significantly attenuated the ability of miR-181c mimics to promote apoptosis.
Figure 6 Involvement of NCAPG in miR-181c-mediated effect of cell invasion, migration, proliferation, and apoptosis. (A) HCC cell NCAPG protein expression was detected by western blotting after transfection with miR-181c mimics and vector-NCAPG. (B) HCC cell NCAPG mRNA expression was detected by western blotting after transfection with miR-181c mimics and vector-NCAPG. (C) Results of wound healing assays measuring the migration capacity of HCC cells overexpressing miR-181c and transfected with vector-NCAPG. (D) Results of transwell assays measuring the migration and invasion capacities of HCC cells overexpressing miR-181c and transfected with vector-NCAPG. (E) Apoptosis rates of HCC cells transfected with miR-181c mimics and vector-NCAPG. (F) Results of EdU assays measuring the proliferative capacity of HCC cells overexpressing miR-181c and transfected with vector-NCAPG. **p<0.01, ***p<0.001, ****p<0.0001.
Abbreviation: HCC, hepatocellular carcinoma.
Discussion
The results of the current study indicate for the first time that miR-181c plays an important role in HCC. In addition, they suggest that miR-181c regulates NCAPG and affects tumor progression.
The prognoses and survival of patients with HCC are poor.Citation1,Citation28 The low survival rate of HCC patients is due to late diagnosis, so it is necessary to find specific biomarkers that facilitate early detection.Citation2,Citation3,Citation29 Many reports indicate that the initiation and progression of HCC involves the dysregulation of various genes and miRNAs.Citation12,Citation30
miRNAs play important roles in various biological and pathological processes involved in HCC. Therefore, we investigated the effects of miRNA in HCC. miR-181c is strongly associated with the occurrence and development of various tumors and affects cell growth and metastasis. Catanzaro et alCitation31 reported that miR-181c was downregulated in glioma and could inhibit cell proliferation by inhibiting the Notch2 pathway. Jung et alCitation13 reported that NFE2L2/NRF2 activity was linked to mitochondria and AMP-activated protein kinase signaling in cancers via miR-181c/mitochondria-encoded cytochrome c oxidase regulation. Su et alCitation14 reported that miR-181c inhibited the growth and invasion of prostate cancer cells by targeting multiple ERK signaling pathway components. Yao et alCitation15 reported that miR-181c inhibited ovarian cancer metastasis and progression by targeting PRKCD expression. miR-181c can also inhibit chemoresistance by targeting ST8SIA4 in chronic myeloid leukemia.Citation16 However, the biological significance of miR-181c in HCC has not yet been elucidated. Therefore, in the current study we investigated the function of miR-181c in HCC.
In the present study, abnormal expression of miR-181c in HCC tissues was determined via the TCGA database. We then verified the expression of miR-181c in HCC tissue specimens and cell lines. miR-181c expression was decreased in HCC. We investigated the effects of experimentally adjusted miR-181c on the biological functions of HCC cells. The results indicated that miR-181c inhibited the invasion and migration capacities of HCC cells. miR-181c induced apoptosis, and inhibited cell proliferation in vitro and in vivo. Most importantly, miR-181c was negatively correlated with NCAPG expression levels in HCC tissues, which suggests that miR-181c may play a role in the regulation of NCAPG expression.
NCAPG is a subunit of the condensed protein complex responsible for the concentration and stabilization of chromosomes during mitosis and meiosis.Citation18 Our previously reported studies shown that the expression of NCAPG in HCC tissue was higher than para-carcinoma tissue. We also found that NCAPG knocked down could affect the cell cycle, apoptosis and metastasis in HCC cells and was significantly associated with prognosis.Citation23 That means, NCAPG might be involved in biological process of tumors. Moreover, recently, accumulating evidence has shown that abnormal expression of NCAPG is associated with progression of HCC. For example, Li et alCitation32 reported that NCAPG is a novel biomarker in hepatocellular carcinoma by weighted gene co-expression network analysis. Wu et alCitation33 reported that NCAPG was identified as a key gene in hepatocellular carcinoma by preliminary bioinformatics analysis. Liu et alCitation34 reported that silencing NCAPG inhibits proliferation and induces apoptosis in hepatocellular carcinoma cells. Zhang et alCitation35 reported that NCAPG is a novel mitotic gene required for hepatocellular cancer cell proliferation and migration. Based on bioinformatics, we predict that NCAPG may be a target gene for miR-181c. In the present study, ectopic expression of miR-181c significantly downregulated endogenous NCAPG mRNA and protein levels in HCC cells, and inhibition of miR-181c upregulated endogenous NCAPG mRNA and protein levels in HCC cells. We also demonstrated that o involvement of NCAPG in miR-181c-mediated effect of cell invasion, migration, proliferation, and apoptosis. Thus, we believe that miR-181c inhibits the growth and metastasis of HCC by regulating NCAPG.
In conclusion, the results of the present study suggest that miR-181c inhibits the growth and metastasis of HCC. Downregulation of NCAPG results from overexpression of miR-181c in HCC. The results of the study provide insight into the molecular mechanism of liver cancer and suggest a potential target for its diagnosis and treatment.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
We express our thankful concerns to the staff and management of Jiangxi Provincial Key Laboratory of Molecular Medicine in the Second Affiliated Hospital of Nanchang University, for providing the experimental facilities. The present study was supported by grants from the Natural Science Foundation of Jiangxi Province, China (no. 20171BAB205063), the Youth Science Foundation of Jiangxi Province, China (no. 20171BAB215037). Jiyuan Ai and Chengwu Gong are co-first authors in this study.
References
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.30207593
- Singh S, Singh PP, Roberts LR, Sanchez W. Chemopreventive strategies in hepatocellular carcinoma. Nat Rev Gastroenterol Hepatol. 2013;11(1):45–54. doi:10.1038/nrgastro.2013.14323938452
- Maluccio M, Covey A. Recent progress in understanding, diagnosing, and treating hepatocellular carcinoma. CA Cancer J Clin. 2012;62(6):394–399. doi:10.3322/caac.2116123070690
- Bartel DP. Metazoan MicroRNAs. Cell. 2018;173(1):20–51. doi:10.1016/j.cell.2018.03.00629570994
- Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–355. doi:10.1038/nature0287115372042
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297.14744438
- Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi:10.1016/j.cell.2009.01.00219167326
- Liang Y, Li E, Min J, et al. miR-29a suppresses the growth and metastasis of hepatocellular carcinoma through IFITM3. Oncol Rep. 2018;40(6):32613272. doi:10.3892/or.2018.6745
- Fang Y, Xue JL, Shen Q, Chen J, Tian L. MicroRNA-7 inhibits tumor growth and metastasis by targeting the phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma. Hepatology. 2012;55(6):1852–1862. doi:10.1002/hep.2557622234835
- Wong CC, Wong CM, Tung EK, et al. The microRNA miR-139 suppresses metastasis and progression of hepatocellular carcinoma by down-regulating Rho-kinase 2. Gastroenterology. 2011;140(1):322–331. doi:10.1053/j.gastro.2010.10.00620951699
- Li N, Fu H, Tie Y, et al. miR-34a inhibits migration and invasion by down-regulation of c-Met expression in human hepatocellular carcinoma cells. Cancer Lett. 2009;275(1):44–53. doi:10.1016/j.canlet.2008.09.03519006648
- Guichard C, Amaddeo G, Imbeaud S, et al. Integrated analysis of somatic mutations and focal copy-number changes identifies key genes and pathw ays in hepatocellular carcinoma. Nat Genet. 2012;44(6):694–698. doi:10.1038/ng.225622561517
- Jung KA, Lee S, Kwak MK. NFE2L2/NRF2 activity is linked to mitochondria and AMP-activated protein kinase signaling in cancers through miR-181c/Mitochondria-encoded cytochrome c oxidase regulation. Antioxid Redox Signal. 2017;27(13):945–961. doi:10.1089/ars.2016.679728383996
- Su Z, Zhang M, Xu M, et al. MicroRNA181c inhibits prostate cancer cell growth and invasion by targeting multiple ERK signaling pa thway components. Prostate. 2018;78(5):343–352. doi:10.1002/pros.2347829341215
- Yao L, Wang L, Li F, Gao X, Wei X, Liu Z. MiR181c inhibits ovarian cancer metastasis and progression by targeting PRKCD expression. Int J Clin Exp Med. 2015;8(9):15198–15205.26629004
- Zhao L, Li Y, Song X, et al. Upregulation of miR-181c inhibits chemoresistance by targeting ST8SIA4 in chronic myelocytic leukemia. Oncotarget. 2016;7(37):60074–60086. doi:10.18632/oncotarget.1105427527856
- Devor EJ, Miecznikowski J, Schickling BM, et al. Dysregulation of miR-181c expression influences recurrence of endometrial endometrioid adenocarcinoma by modulating NOTCH2 expression: an NRG Oncology/Gynecologic Oncology Group study. Gynecol Oncol. 2017;147(3):648–653. doi:10.1534/genetics.109.10647628969912
- Eberlein A, Takasuga A, Setoguchi K, et al. Dissection of genetic factors modulating fetal growth in cattle indicates a substantial role of the non-SMC condensin I complex, subunit G (NCAPG) gene. Genetics. 2009;183(3):951–964. doi:10.1534/genetics.109.10647619720859
- Murphy LA, Sarge KD. Phosphorylation of CAP-G is required for its chromosomal DNA localization during mitosis. Biochem Biophys Res Commun. 2008;377(3):1007–1011. doi:10.1016/j.bbrc.2008.10.11418977199
- Li S, Xuan Y, Gao B, et al. Identification of an eight-gene prognostic signature for lung adenocarcinoma. Cancer Manag Res. 2018;10:3383–3392. doi:10.2147/CMAR.S17394130237740
- Arai T, Okato A, Yamada Y, et al. Regulation of NCAPG by miR-99a-3p (passenger strand) inhibits cancer cell aggressiveness and is involved in CRPC. Cancer Med. 2018;7(5):1988–2002. doi:10.1002/cam4.145529608247
- Liang ML, Hsieh TH, Ng KH, et al. Downregulation of miR-137 and miR-6500-3p promotes cell proliferation in pediatric high-grade gliomas. Oncotarget. 2016;7(15):19723–19737. doi:10.18632/oncotarget.773626933822
- Liu W, Liang B, Liu H, et al. Overexpression of non-SMC condensin I complex subunit G serves as a promising prognostic marker and therapeutic target for hepatocellular carcinoma. Int J Mol Med. 2017;40(3):731–738. doi:10.3892/ijmm.2017.307928737823
- Roessler S, Jia HL, Budhu A, et al. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res. 2010;70(24):10202–10212. doi:10.1158/0008-5472.CAN-10-260721159642
- Min J, Feng Q, Liao W, et al. IFITM3 promotes hepatocellular carcinoma invasion and metastasis by regulating MMP9 through p38/MAPK signaling. FEBS Open Bio. 2018;8(8):1299–1311. doi:10.1002/2211-5463.12479
- Boufraqech M, Zhang L, Jain M, et al. miR-145 suppresses thyroid cancer growth and metastasis and targets AKT3. Endocr Relat Cancer. 2014;21(4):517–531. doi:10.1530/ERC-14-007724781864
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.126211846609
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. doi:10.3322/caac.2138728055103
- European Association for the Study of the Liver. Electronic address eee, European Association for the Study of the L. EASL clinical practice guidelines: management of hepatocellular carcinoma. J Hepatol. 2018;69(1):182–236. doi:10.1016/j.jhep.2018.03.01929628281
- Baer C, Claus R, Plass C. Genome-wide epigenetic regulation of miRNAs in cancer. Cancer Res. 2013;73(2):473–477. doi:10.1158/0008-5472.CAN-12-373123316035
- Catanzaro G, Sabato C, Russo M, et al. Loss of miR-107, miR-181c and miR-29a-3p promote activation of Notch2 signaling in Pediatric High-Gra de Gliomas (pHGGs). Int J Mol Sci. 2017;18(12):2742. doi:10.3390/ijms18122742
- Li B, Pu K, Wu X. Identifying novel biomarkers in hepatocellular carcinoma by weighted gene co-expression network analy sis. J Cell Biochem. 2019;1–14.doi: 10.1002/jcb.28420
- Wu M, Liu Z, Zhang A, Li N. Identification of key genes and pathways in hepatocellular carcinoma: A preliminary bioinformatics an alysis. Medicine (Baltimore). 2019;98(5):e14287. doi:10.1097/MD.000000000001428730702595
- Liu K, Li Y, Yu B, Wang F, Mi T, Zhao Y. Silencing non-SMC chromosome-associated polypeptide G inhibits proliferation and induces apoptosis in hepatocellular carcinoma cells. Can J Physiol Pharmacol. 2018;1–9. doi:10.1139/cjpp-2018-0016
- Zhang Q, Su R, Shan C, Gao C, Wu P. Non-SMC condensin I complex, subunit G (NCAPG) is a novel mitotic gene required for hepatocellular cancer cell proliferation and migration. Oncol Res. 2018;26(2):269–276. doi:10.3727/096504017X1507596756098029046167