
Abstract
Purpose
Paeonol, a natural product derived from the root of Cynanchum paniculatum (Bunge) K. Schum and the root of Paeonia suffruticosa Andr. (Ranunculaceae) has attracted extensive attention for its anti-cancer proliferation effect in recent years. The present study examined the role of paeonol in suppressing migration and invasion in pancreatic cancer cells by inhibiting TGF-β1/Smad signaling.
Methods
Cell viability was evaluated by MTT and colonial formation assay. Migration and invasion capabilities were examined by cell scratch-wound healing assay and the Boyden chamber invasion assay. Western Blot and qRT-PCR were used to measure the protein and RNA levels of vimentin, E-cadherin, N-cadherin, and TGF-β1/Smad signaling.
Results
At non-cytotoxic dose, 100 μΜ and 150 μΜ of paeonol showed significant anti-migration and anti-invasion effects on Panc-1 and Capan-1 cells (p<0.01). Paeonol inhibited epithelial-mesenchymal-transition by upregulating E-cadherin, and down regulating N-cadherin and vimentin expressions. Paeonol inhibited TGF-β1/Smad signaling pathway by downregulating TGF-β1, p-Smad2/Smad2 and p-Smad3/Smad3 expressions. Further, TGF-β1 attenuated the anti-migration and anti-invasion capacities of paeonol in Panc-1 and Capan-1 cells.
Conclusion
These findings revealed that paeonol could suppress proliferation and inhibit migration and invasion in Panc-1 and Capan-1 cells by inhibiting the TGF-β1/Smad pathway and might be a promising novel anti-pancreatic cancer drug.
Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of the most lethal malignant tumours with growing incidence worldwide.Citation1 Due to the asymptomatic nature of PDAC, more than 80% of patients are in advanced stages and are diagnosed with apparent local invasion and metastasis.Citation2 The treatment outcomes have not improved over the past decade for patients with advanced diseases.Citation3 Therefore, the discovery and the development of new therapeutic strategies for PDAC are warranted.
Epithelial-mesenchymal-transition (EMT) is a critical process involved in the induction of multiple signaling pathways and is thought to play a crucial role in the development and progression of cancer metastasis.Citation4,Citation5 During EMT, cancer cells lose cell polarity and cell-to-cell junctions, resulting in the attaining of invasive phenotype.Citation6,Citation7 Many studies have demonstrated that the reactivation of EMT links cancer-associated inflammation to cancer aggressiveness.Citation6–Citation8 Transforming growth factor-beta (TGF-β1) is a pro-inflammatory cytokine that is frequently over-expressed in neoplasms and inflammatory tissues and is a potent inducer of EMT.Citation8 It has been purposed that EMT is one of the most distinctive features of PDAC and is associated with an unfavourable prognosis.Citation5
Paeonol (Pae, C99H10O3, CAS Number 552-41-0, PubChem CID 11092) is a component and one of the major bioactive constitutes in many Chinese herbs, such as the root of Cynanchum paniculatum (Bunge) K. Schum and the root of Paeonia suffruticosa Andr. (Ranunculaceae).Citation9–Citation11 Previous studies suggested that Paeonol possesses anti-proliferation, apoptosis induction, and cell cycle arresting effects in various types of cancers, such as breast, colorectal, lung, ovarian, gastric cancer, etc.Citation9,Citation12–Citation21 More recent studies reported that Paeonol enhanced paclitaxel-induced apoptosis in breast cancer cells,Citation20,Citation22 reversed Doxorubicin resistance in liver cancer,Citation23 and enhanced the radiosensitivity of lung cancer cells.Citation24 Studies also reported the anti-metastatic activities of Paeonol in human fibrosarcoma, chondrosarcoma and gastric cancer.Citation9,Citation25,Citation26 However, whether Paeonol exerts anti-migration and anti-invasion effects in pancreatic cancer cells and the potential underlying mechanisms of action are yet to be elucidated. In the present study, Panc-1 and Capan-1 cell lines are used to study the effects of Paeonol on migration and invasion, and the potential role of TGF-β1-induced EMT as an underlying mechanism of Paeonol is investigated.
Materials and Methods
Chemicals and Reagents
Paeonol (HOC6H3(OCH3)COCH3, molecular weight: 166.17, purity 99%) was purchased from Sigma-Aldrich Company (Cat. H35803, Sigma-Aldrich, St. Louis, MO, USA). 10M of Paeonol was prepared as a stock concentration in dimethyl sulfoxide (DMSO, Sigma, MO, USA) and stored at −20°C for further use. Recombinant Human Transforming Growth Factor β1 (TGF-β1) was purchased from MedChemExpress (Cat. HY-P7118, Monmouth Junction, NJ, USA). TGF-β1 was reconstituted to 100 µg/mL with sterile ddH2O and store at −20°C for further use.
Cell Line and Cell Culture
Human pancreatic cancer Panc-1 and Capan-1 cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured according to manufacturer’s instruction. In brief, the cells were maintained in high glucose Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, Massachusetts, USA) supplemented with 100 µg/mL each of penicillin and streptomycin (Gibco, Massachusetts, USA), and 10% fetal bovine serum (FBS, Gibco, Massachusetts, USA) under sterilized and humified 5% CO2 condition at 37°C. The morphologies of the cells were assessed periodically and mycoplasma contamination were tested using MycoAlert (Lonza, Rockland, ME, USA).
Cell Viability Assay
The cells were plated in triplicate in 96 well plates at a density of 5 × 103 cells in 0.1 mL and incubated overnight. Serial dilutions of Paeonol at doses ranging from 0–300μM were added and incubated for another 24, 48, or 72 hrs at 37°C. MTT (1-(4,5-Dimethylthiazol-2-yl)-3,5-diphenylformazan, Sigma-Aldrich, St. Louis, MO, USA) was added to each well of the plates followed by incubation at 37°C for 4 hrs. Media were carefully aspired and 200μL of DMSO was added to each well. The plates were then incubated for 15 mins at 37°C, protected from light and measured at 570 nm by a Multiskan MS microplate reader (ThermoScientific, Massachusetts, USA). The growth curves of Paeonol-treated cells were plotted based on OD values obtained from the assay. Subsequently, Paeonol concentrations below the IC50 values after 48 hrs treatment (100 and 150μM, IC10 and IC20, respectively) were selected for further study. Each experiment was performed in triplicate and with at least 3 individual experiments carried out.
Colonial Formation Assay
Approximately 1000 Panc-1 and Capan-1 cells were seeded in 6-well tissue culture plates and incubated overnight. The culture medium was replaced with fresh medium containing 0, 100, 150 µM of Paeonol and then cultured for 12 days. Cells were then fixed in 4% paraformaldehyde (Sigma-Aldrich, St. Louis, MO, USA) and stained with crystal violet (Sigma-Aldrich, St. Louis, MO, USA) for visualization. Images of the colonies were taken after washing and air drying. All experiments were conducted in triplicate.
Scratch-Wound Migration Assay
Panc-1 and Capan-1 cells were seeded in 6-well plates and allowed to grow until confluent. The confluent cells were wounded by scraping with a sterile 1-mL-pipette tip. The culture medium was replaced with fresh medium containing 0, 100, 150µM of Paeonol and cultured for 48 hrs. The rate of wound closure was monitored by images captured with a phase-contrast microscope at 0, 24, and 48 hrs. At least 3 independent experiments were performed.
Boyden Chamber Invasion Assay
Boyden chamber invasion assays were carried out using 8.0-µm cell culture Transwell inserts (Corning, Corning, New York, USA). Transwell chambers were pre-coated with Matrigel (Corning, New York, USA) at 1:5 dilution in serum-free medium for 2 hrs at 37°C. 2 × 104 cells were placed in the upper chambers and incubated overnight. The cells were then treated with 0, 100, and 150μM of Paeonol. After 48 hrs in culture, the cells remaining on the upper surface of the membrane were removed by wiping with cotton swabs. The remaining cells attached to the lower surface of the membrane were fixed 4% paraformaldehyde and stained with crystal violet for 30 mins at room temperature for visualization. Images were acquired under light microscopy, and fully invaded cells were quantitated by counting cells in 5 random fields. Each experiment was performed in triplicate with at least 3 individual experiments and the mean values ± SD were presented.
Real-Time Quantitative Polymerase Chain Reaction
For mRNA analysis, total RNA was isolated using RNA isoPlus (TaKaRa Bio Inc., Shiga, Japan) according to the manufacturer’s protocol. Reverse transcription for cDNA synthesis was performed using a PrimeScript RT reagent Kit (TaKaRa Bio Inc., Shiga, Japan) and amplified by using the ProFlex PCR system (Thermo Fisher, Massachusetts, USA), according to the manufacturer’s instructions. Primers were purchased from Sangon Biotech (Shanghai, China). Quantitative PCR was performed using the SYBR green system (TaKaRa, Japan) according to the manufacturer’s instructions with LightCycler 480 PCR system (Roche, Basel, Switzerland). All reactions were performed in triplicate and the expression of β-actin was used to normalize the gene expression of each sample. The 2−ΔΔCtmethod was used to determine relative gene expression. The gene-specific primers used are listed in .
Table 1 Sequences of PCR Primers for Target Gene Detection
Immunoblotting
The total protein was extracted from cultured cells with ice cold-radio-immunoprecipitation assay (RIPA) extraction buffer (Thermo Fisher, Massachusetts, USA) supplemented with protease and phosphatase inhibitors (Roche, Basel, Switzerland) for 30 mins on ice and followed by centrifugation at 12,500 rpm for 10 mins at 4°C. Protein concentration was quantitated using the Bradford protein assay kit (BioRad, Hercules, CA, USA) according to manufacturer’s instruction. After electrophoresis, protein were transferred to PVDF membranes and were then blocked with 5% bovine serum albumins (BSA, Sigma-Aldrich, St. Louis, MO, USA) for 2 hrs at room temperature. Subsequently, the membrane was incubated with respective primary antibodies overnight at 4°C and then with appropriate secondary antibody (CST, Danvers, MA, USA) at room temperature for 2 hrs. Target protein expression was examined with ECL advanced kit (GE Healthcare, Little Chalfont, UK) and visualized with Tanon 6100 Chemiluminescent Imaging System (Tanon, Shanghai, China) and quantitative analyses were performed using ImageJ software (National Institutes of Health, USA). Primary antibodies used were all purchased from Cell Signaling Technology (Danvers, MA, USA) and were as follows: p-Smad2 (Ser465/467)(#3108), Smad2 (#5339), p-Smad3 (Ser423/425)(#9520), Smad3 (#9523), E-Cadherin (#3195), Vimentin (5741), N-Cadherin (#13116), and β-actin (#4970). All signals were normalized to that of β-actin.
Statistical Analysis
Data were expressed as the means ± standard deviations. Statistical analyses were performed using analysis of variance or Student’s t-tests. A p-value < 0.05 was considered statistically significant throughout this study.
Results
Paeonol Suppressed Pancreatic Cancer Cells Proliferation in vitro
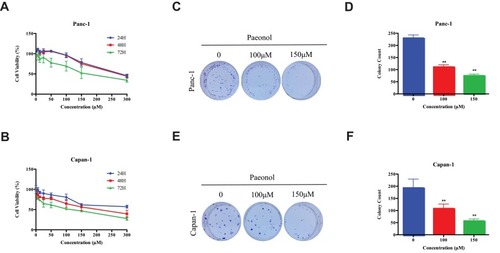
First, we determined the anti-proliferation effect of Paeonol on Panc-1 and Capan-1 cells using MTT assay. As shown in , Panc-1 and Capan-1 cells were treated with various doses of Paeonol for 24, 48 and 72 hrs. It was observed that Paeonol inhibited the proliferation of Panc-1 and Capan-1 cells in a dose- and time-dependent manner. The 50% inhibitions concentration (IC50) of Paeonol on Panc-1 cells were 274.4, 265.5, 175.2μM for 24, 48 and 72 hrs, respectively, and that of the Capan-1 cells were 380.7, 211.7, 90.9μM, respectively. At 48 hrs of incubation, the IC20 and IC10 of Paeonol of Panc-1 cell were 151.6 and 109.2μM, respectively, and that of the Capan-1 cells were 52.9 and 23.5μM, respectively. Colonial formation assay showed that (–) prolonged exposure Paeonol at 100 and 150μΜ for 12 days suppressed colony formation capability of Panc-1 and Capan-1 cells. To further evaluate the anti-migration and anti-invasion effects of Paeonol on PDAC, two doses of 100 and 150μM were selected for subsequent experiments.
Figure 1 The cytotoxicity and effects of Paeonol on PDACs. (A) The cytotoxicity of Paeonol at different concentrations (0, 6.25, 12.5, 25, 50, 100, 150, 300μM) and the cell viability at 24 hrs, 48 hrs and 72 hrs in Panc-1; (B) The cytotoxicity of Paeonol at various concentrations (0, 6.25, 12.5, 25, 50, 100, 150, 300μM) and cell viability for 24 hrs, 48 hrs and 72 hrs in Capan-1; (C) Paeonol (100 or 150μM) reduced clonogenic survival of Panc-1; (D) Compared to non-treated controls, Paeonol (100 or 150μM) suppressed colonial formation of Panc-1; (E) Paeonol (100 or 150μM) reduced clonogenic survival of Capan-1; (F) Compared to non-treated controls, Paeonol (100 or 150μM) suppressed colonial formation of Capan-1. Data were shown as mean ± SD of three independent experiments. **P<0.01.
Paeonol Suppressed Pancreatic Cancer Cell Migration and Invasion in vitro at Non-Cytotoxic Doses
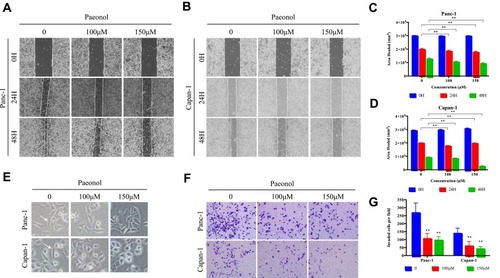
In order to test whether Paeonol exerts anti-migration and anti-invasion effect on PDAC at non-cytotoxic dose, cell scratch-wound migration assay and Transwell invasion assay were performed. The scratch-wound assay results showed that Paeonol treatment reduced Panc-1 and Capan-1 migration (–). Transwell invasion assay results showed that the invasive rate of the Paeonol 100 and 150μM treatment groups was lower than that of the untreated group (). Furthermore, Panc-1 and Capan-1 cells in the control group exhibited mesenchymal-like morphology, and cells treated with Paeonol exhibited epithelial-like morphology (). These results suggested the anti-migration and anti-invasion effects of Paeonol may potentially be related EMT.
Figure 2 Paeonol suppresses the migration and invasion capacities of PDAC cells by promoting EMT. (A, B) Scratch wound-healing assay. Panc-1 and Capan-1 cells were treated with 100 or 150μΜ of Paeonol. The movement of cells into the wounds was shown at 0, 24 hrs, and 48 hrs after scratching (magnification: 100x); (C, D) Panc-1 and Capan-1 cells migration were assessed by restoring scratches and the reduction in the area of the initial scratches were compared. All results are representative of three independent experiments. **P<0.01, as compared Paeonol (100 or 150μΜ) with the non-treated control group; (E) Morphological changes in Panc-1 and Capan-1 cells after 48 hrs of culture with Paeonol (100 or 150μΜ). Arrows indicate cell shapes representing mesenchymal phenotypes; (F) The effect of Paeonol (100 or 150μΜ) treatment for 48 hrs on Panc-1 and Capan-1 cells was detected by chambers assays (magnification: 40x); (G) Histogram, based on five random fields selected for statistical analysis, shows the number of invaded cells. Data are shown as mean ± SD of three independent experiments. **P<0.01.
Paeonol Suppressed EMT in Pancreatic Cancer Cells Through the Inhibition of TGF-β1/Smad Signaling
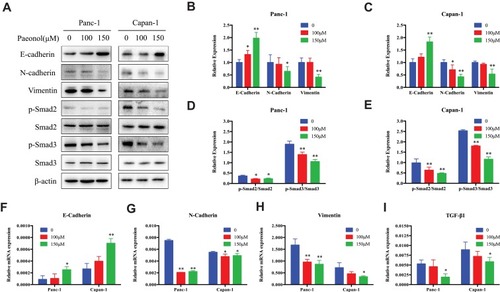
EMT had been associated with the increased metastatic potential in PDAC. Since Paeonol could inhibit the migration and invasion capabilities of Panc-1 and Capan-1 cells, we sought to investigate the potential mechanism and characterization of EMT by detecting the expression of EMT-related genes and proteins in Paeonol-treated cells. As shown in , Paeonol-treated Panc-1 and Capan-1 cells showed increased protein expression of the epithelial marker E-cadherin, and decreased protein expression of mesenchymal markers, Vimentin and N-cadherin (–). Consistently, Paeonol treatment reduced mRNA expression of Vimentin and N-cadherin, while it increased the expression level of E-cadherin (–). These results indicated that Paeonol could effectively inhibit PDAC via suppression EMT. Considering that EMT is frequently induced by TGF-β1 and the TGF-β1/Smad signaling pathway is a critical pathway triggered by phosphorylation of Smads, we sought to determine whether Paeonol possesses its anti-migration and anti-invasion effect by suppression of the active status of Smads signaling pathway. Not surprisingly, our results showed that Paeonol inhibited the expression of TGF-β1 and phosphorylation of Smad2 and Smad3 in Panc-1 and Capan-1 cells ().
Figure 3 Paeonol reduces the mesenchymal characteristics and promotes epithelial characteristics of Panc-1 and Capan-1 cells and modulates the TGF-β1/Smad signaling. (A) Western blot analysis of protein levels of E-cadherin, N-cadherin, Vimentin, p-Smad2, Smad2, p-Smad3, Smad3 in Panc-1 and Capan-1 cells treated with either 100 or 150μΜ of Paeonol for 48 hrs; (B, C) Histograms show the change of relative protein expression of E-cadherin, N-cadherin, Vimentin in Panc-1 and Capan-1 cells; (D, E) Histograms show the change of relative protein expression of p-Smad2/Smad2, p-Smad3/Smad3 in Panc-1 and Capan-1 cells; (F–I) qRT-PCR analysis of mRNA levels of E-cadherin, N-cadherin, Vimentin and TGF-β1 in Panc-1 and Capan-1 cells treated with either 100 or 150μΜ of Paeonol for 48 hrs. Data are shown as mean ± SD of three independent experiments. *P < 0.05, **P < 0.01.
Paeonol Abrogated TGF-β1-Induced Migration and Invasion Effects of in Pancreatic Cancer Cells
Since Paeonol inhibited the migration and invasion of Panc-1 and Capan-1 cells by altering cell morphology, we sought to determine whether the anti-migration and anti-invasion effects of Paeonol were related to TGF-β1-induced EMT. We introduced TGF-β1 cytokine (HY-P7118) and incubated at 10ng/mL with or without 100μM of Paeonol for 48 hrs. Scratch wound assay and Boyden chamber invasion assay results showed that TGF-β1 remarkably induced the migration and invasion of Panc-1 and Capan-1 cells, and the addition of Paeonol reversed TGF-β1-induced migration and invasion (). As shown in , the morphology of Panc-1 and Capan-1 cells showed a more mesenchymal phenotype after treatment with 10ng/mL TGF-β1. We also found that when Paeonol and TGF-β1 were added simultaneously, the levels of E-cadherin turned back to the former level without any treatment (–). These results indicated that Paeonol inhibited migration and invasion of Panc-1 and Capan-1 cells by suppressing TGF-β1/SMAD signaling pathway activation ().
Figure 4 Paeonol attenuated the migration-promoting ability of TGF-β1. (A, B) Scratch wound-healing assay. Cell migration of Panc-1 and Capan-1 cells treated with or without Paeonol (100μΜ) and with or without TGF-β1 (10ng) for 48 hrs was assessed by restoring scratches and comparing with the area of the initial scratch (magnification: 100x); (C) The effects of Paeonol (100μΜ) treated with or without TGF-β1 (10ng) for 48 hrs on Panc-1 and Capan-1 cells migration were examined using chamber assays (magnification: 40x); (D, E) Histogram, based on five random fields of three independent experiments selected for statistical analysis, showed the number of invaded cells; (F, G) Histogram, based on Panc-1 and Capan-1 cell migrations were assessed by restoring scratches and the repair of the initial scratched area. All results are representative of three independent experiments. *P<0.05, **P<0.01, as compared with the non-treated control group.
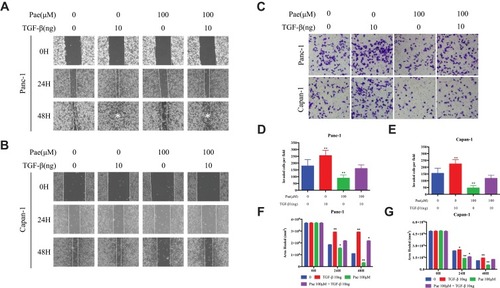
Figure 5 Paeonol attenuated mesenchymal characteristics promoting the effect of TGF-β1. (Α) Morphological changes of Panc-1 and Capan-1 cells after 48 hrs of treatment with or without Paeonol (100μΜ) and with or without TGF-β1 (10ng). Arrows indicate cell shapes representing mesenchymal phenotypes; (B, C) Western blot analysis of protein levels of E-cadherin, Vimentin in Panc-1 and Capan-1 cells treated with or without Paeonol (100μΜ) and with or without TGF-β1 (10ng) for 48 hrs; (D, E) Histograms show the change of relative protein expression of E-cadherin, Vimentin in Panc-1 and Capan-1 cells. Data are shown as mean ± SD of three independent experiments. *P < 0.05, **P < 0.01.
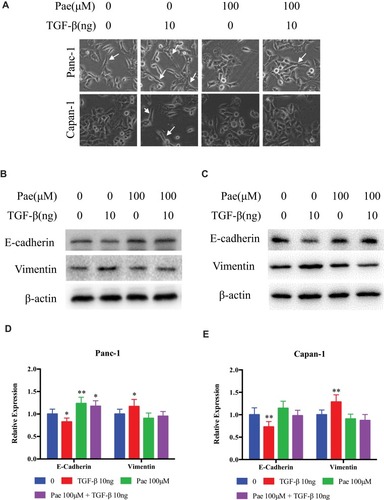
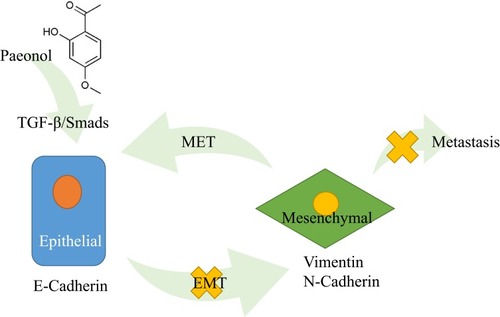
Figure 6 Schematic diagram of the molecular mechanisms of Paeonol’s the anti-migration and anti-invasion effects in PDAC. Paeonol could potentially inhibit migration and invasion of Panc-1 and Capan-1 cells by suppressing the activation of TGF-β1/SMAD signaling pathway and suppressing EMT in PDAC.
Discussion
In this study, we investigated the role of Paeonol in PDAC, focusing on the anti-migration and anti-invasion effects of Paeonol in Panc-1 and Capan-1 cell lines. The results indicated that Paeonol inhibited PDAC cell growth and suppressed colony formation in a time- and dose-dependent manner. Then, to the best of our knowledge, this study demonstrated for the first time that Paeonol inhibited migration and invasion of PDAC cells in non-cytotoxic doses. Our results indicated that Paeonol inhibited TGF-β1-induced EMT in PDAC cells by modulating phosphorylation of the Smad2/3 pathway components. Based on our data, we suggested that Paeonol may be a potential therapeutic agent for the treatment of PDAC; however, further experiments are warranted to confirm its anti-cancer effect in vivo.
Tumour metastasis is a prominent feature of cancer cells and a major contributor to cancer-associated death.Citation27 In PDAC, more than 80% of the patients present with metastasis upon diagnosis.Citation27–Citation29 Therefore, many efforts have been made in aims to suppress the metastatic property of PDAC. Metastasis is a complicated multi-step process involves the ability of cancer cell to detach by losing cell-to-cell junction and cellular polarity, break away from the extracellular matrix, attain increased motility and increased invasive phenotype.Citation30,Citation31 EMT is one of the most distinctive features of PDAC and is associated with metastasis, drug resistance and poor prognosis.Citation32 Accumulating evidence suggests that EMT is the initiation step of cancer metastasis involving the downregulation of epithelial markers, such as E-cadherin and upregulation of mesenchymal markers, such as N-cadherin.Citation33,Citation34 Various signaling pathways are involved in the induction of EMT, include TGF-β1, Wnt-β-catenin, Notch, Hedgehog, and tyrosine kinases.Citation32,Citation35 TGF-β1 is an inflammatory cytokine and a promoter and supporter of EMT.Citation36 In PDAC, TGF-β1 has been shown to induce EMT and association between TGF-β1 induced gene expression and clinical outcomes have been described.Citation37,Citation38 Mechanically, TGF-β1 induces phosphorylation cascade of downstream cytoplasmic substrates, such as the SMAD proteins, and has been purposed to further affects epithelial/mesenchymal differentiation, activation of PI3K-AKT pathway, induction of angiogenesis via activating vascular endothelial growth factor and metalloproteases (MMPs).Citation38–Citation40 It is possible that as a downstream effect, TGF-β1-induced EMT further intensified the inflamed tumour microenvironment and contributes to cancer progression. These suggest that the inhibition of TGF-β1 and suppression of EMT may be effective for the treatment of cancer.
Paeonol is a natural phenolic compound that is often found in various of herbs and has been shown to exert anti-tumour, anti-oxidant and anti-inflammatory effects. It has been reported that the cancer growth inhibition effects of Paeonol involve COX-2/PGE2, TNF-α, caspase-cascade, MAPK pathway, PI3K-AKT pathway, etc.Citation14,Citation17,Citation41,Citation42 Contrary to its relatively well established anti-proliferative and apoptosis-inducing effects in the literature, relatively few works have been published, documenting the role of Paeonol in metastasis. The anti‑metastatic activities of Paeonol on human fibrosarcoma and chondrosarcoma cells have been previously reported.Citation25,Citation26 Furthermore, previous studies have shown that Paeonol inhibits invasion and migration of gastric cancer cells by down-regulating the MMPs.Citation9 However, the anti-metastatic effects of Paeonol, especially in PDAC, lack conclusive evidence. In the absence of other known studies on the anti-migration and anti-invasion activities of Paeonol, we aim to further investigate the activities of Paeonol on migration and invasion and its underlying mechanisms.
In our study, we investigated the effects of Paeonol on ameliorating TGF-β1-induced PDAC cell migration, invasion and EMT. First, PDAC cells were exposed to various concentrations of Paeonol for 24, 48 and 72 hrs to determine the appropriate concentrations. Since 100μM and 150μΜ Paeonol effectively inhibited PDAC cell migration and invasion without inducing apparent cell proliferation arrest, these two doses were used in our experiments. Next, as reported in previous studies, 10 ng/mL of TGF-β1 effectively induced epithelial–mesenchymal transition and promoted PDAC cell migration and invasion.Citation43,Citation44 We then affirmed the effect of this dose in both cell lines that TGF-β1 induced PDAC cells to transform their morphology from an epithelial to a mesenchymal morphology. Our results demonstrated that EMT played a crucial role in the malignant behavior of PDAC, and TGF-β1 promoted cell migration, while Paeonol suppressed migration in a concentration-dependent manner.
The TGF-β1/Smad signaling pathway is involved in many diseases, including cardiovascular diseases, neurological disorders and cancers.Citation45–Citation47 In the early stages of tumorigenesis, TGF-β1 acts as a tumour suppressor and inhibits tumour by inducing cell cycle arrest and apoptosis.Citation48 As the tumour progresses, the role of TGF-β1 changes into a tumour promoter and induces EMT, promoting resistance to apoptosis, migration, invasion, and metastasis.Citation45 Nie et al showed that the combination of Paeonol and Panax notoginsenosides can inhibit the TGF-β1/Smad signaling pathway in ventricular remodeling after myocardial infarction in rats.Citation49 Wu et al reported that Paeonol alleviated CCl4-induced liver fibrosis in mice models and inhibited the activation of hepatic stellate cells by inhibiting the TGF-β1/Smad3 signaling pathway.Citation50 In our studies, Paeonol markedly suppressed the phosphorylation of Smad2 and Smad3. Furthermore, our results showed a significant increase in the mesenchymal markers Vimentin and N-Cadherin under TGF-β1 stimulation, which was restrained by Paeonol. These findings indicated that Paeonol may prevent the development of EMT by regulating the TGF-β1/Smad signaling pathway in pancreatic cancer.
Our research has some limitations. First, we investigated the role and mechanism of Paeonol in cultured PDAC cells. However, we did not establish animal models of PDAC, such as murine xenografts or orthotropic models. Second, we elucidated the anti-migration and anti-invasion effects of Paeonol on EMT by a TGF-β1/Smad-dependent pathway; however, other signaling pathways are involved in EMT and cancer migration and invasion. It is necessary to further study the mechanism of action of Paeonol in PDAC cell lines and animal models to further determine its anti-cancer effect and the interaction between tumour inflammatory environment and metastasis. Moreover, whether Paeonol can be applied to the treatment of pancreatic cancer requires further experiments.
Conclusions
In conclusion, in the present study, Paeonol was found to exert anti-migration and anti-invasion effects in PDAC cells, at least in part due to its anti-inflammatory effects. The underlying mechanism of its anti-cancer effect may involve the inhibition of the TGF-β1/Smad signaling and EMT. Our study provided a scientific evidence for the potential therapeutic value of Paeonol in the treatment of pancreatic cancer.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (81573752 and 81603444), Shanghai Science and Technology Committee Project (15401932500) and Workstation of Xia Xiang, National Master of Traditional Chinese Medicine from the State Administration of Traditional Chinese Medicine of the People’s Republic of China. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Disclosure
The authors report no conflicts of interest in this work.
References
- Kamisawa T, Wood LD, Itoi T, Takaori K. Pancreatic cancer. Lancet. 2016;388(10039):73–85. doi:10.1016/S0140-6736(16)00141-026830752
- Vincent A, Herman J, Schulick R, Hruban RH, Goggins M. Pancreatic cancer. Lancet. 2011;378(9791):607–620. doi:10.1016/S0140-6736(10)62307-021620466
- Manji GA, Olive KP, Saenger YM, Oberstein P. Current and emerging therapies in metastatic pancreatic cancer. Clin Cancer Res. 2017;23(7):1670–1678. doi:10.1158/1078-0432.CCR-16-231928373365
- Heerboth S, Housman G, Leary M, et al. EMT and tumor metastasis. Clin Transl Med. 2015;4:6. doi:10.1186/s40169-015-0048-325852822
- Zhou P, Li B, Liu F, et al. The epithelial to mesenchymal transition (EMT) and cancer stem cells: implication for treatment resistance in pancreatic cancer. Mol Cancer. 2017;16(1):52. doi:10.1186/s12943-017-0624-928245823
- Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. doi:10.1038/nrm375824556840
- Jolly MK, Boareto M, Huang B, et al. Implications of the hybrid epithelial/mesenchymal phenotype in metastasis. Front Oncol. 2015;5:155. doi:10.3389/fonc.2015.0015526258068
- Lindsey S, Langhans SA. Crosstalk of oncogenic signaling pathways during epithelial-mesenchymal transition. Front Oncol. 2014;4:358. doi:10.3389/fonc.2014.0035825566498
- Lyu ZK, Li CL, Jin Y, et al. Paeonol exerts potential activities to inhibit the growth, migration and invasion of human gastric cancer BGC823 cells via downregulating MMP2 and MMP9. Mol Med Rep. 2017;16(5):7513–7519. doi:10.3892/mmr.2017.757628944890
- Sun Z, Du J, Hwang E, Yi TH. Paeonol extracted from paeonia suffruticosa Andr. ameliorated UVB-induced skin photoaging via DLD/Nrf2/ARE and MAPK/AP-1 pathway. Phytother Res. 2018;32(9):1741–1749. doi:10.1002/ptr.610029748977
- Guo L, Liang J, Yang Q. Determination of paeonol in the root of cynanchum paniculatum (Bge.) Kitag. by gas chromatography. Zhongguo Zhong Yao Za Zhi. 1996;21(8):484–485, 511.9642412
- Lei Y, Li HX, Jin WS, et al. The radiosensitizing effect of paeonol on lung adenocarcinoma by augmentation of radiation-induced apoptosis and inhibition of the PI3K/Akt pathway. Int J Radiat Biol. 2013;89(12):1079–1086. doi:10.3109/09553002.2013.82505823875954
- Zhou HM, Sun QX, Cheng Y. Paeonol enhances the sensitivity of human ovarian cancer cells to radiotherapy-induced apoptosis due to downregulation of the phosphatidylinositol-3-kinase/Akt/phosphatase and tensin homolog pathway and inhibition of vascular endothelial growth factor. Exp Ther Med. 2017;14(4):3213–3220. doi:10.3892/etm.2017.487728912871
- Yin J, Wu N, Zeng F, Cheng C, Kang K, Yang H. Paeonol induces apoptosis in human ovarian cancer cells. Acta Histochem. 2013;115(8):835–839. doi:10.1016/j.acthis.2013.04.00423768958
- Fu J, Yu L, Luo J, Huo R, Zhu B. Paeonol induces the apoptosis of the SGC7901 gastric cancer cell line by downregulating ERBB2 and inhibiting the NFkappaB signaling pathway. Int J Mol Med. 2018;42(3):1473–1483. doi:10.3892/ijmm.2018.370429845222
- Li N, Fan LL, Sun GP, et al. Paeonol inhibits tumor growth in gastric cancer in vitro and in vivo. World J Gastroenterol. 2010;16(35):4483–4490. doi:10.3748/wjg.v16.i35.448320845518
- Li M, Tan SY, Wang XF. Paeonol exerts an anticancer effect on human colorectal cancer cells through inhibition of PGE(2) synthesis and COX-2 expression. Oncol Rep. 2014;32(6):2845–2853. doi:10.3892/or.2014.354325322760
- Li M, Tan SY, Zhang J, You HX. Effects of paeonol on intracellular calcium concentration and expression of RUNX3 in LoVo human colon cancer cells. Mol Med Rep. 2013;7(5):1425–1430. doi:10.3892/mmr.2013.137223504038
- Ou Y, Li Q, Wang J, Li K, Zhou S. Antitumor and apoptosis induction effects of paeonol on mice bearing EMT6 breast carcinoma. Biomol Ther (Seoul). 2014;22(4):341–346. doi:10.4062/biomolther.2013.10625143814
- Cai J, Chen S, Zhang W, et al. Paeonol reverses paclitaxel resistance in human breast cancer cells by regulating the expression of transgelin 2. Phytomedicine. 2014;21(7):984–991. doi:10.1016/j.phymed.2014.02.01224680370
- Saahene RO, Wang J, Wang ML, Agbo E, Pang D. The antitumor mechanism of paeonol on CXCL4/CXCR3-B signals in breast cancer through induction of tumor cell apoptosis. Cancer Biother Radiopharm. 2018;33(6):233–240. doi:10.1089/cbr.2018.245029847158
- Zhang W, Cai J, Chen S, et al. Paclitaxel resistance in MCF-7/PTX cells is reversed by paeonol through suppression of the SET/phosphatidylinositol 3-kinase/Akt pathway. Mol Med Rep. 2015;12(1):1506–1514. doi:10.3892/mmr.2015.346825760096
- Fan L, Song B, Sun G, Ma T, Zhong F, Wei W. Endoplasmic reticulum stress-induced resistance to doxorubicin is reversed by paeonol treatment in human hepatocellular carcinoma cells. PLoS One. 2013;8(5):e62627. doi:10.1371/journal.pone.006262723658755
- Chen B, Ning M, Yang G. Effect of paeonol on antioxidant and immune regulatory activity in hepatocellular carcinoma rats. Molecules. 2012;17(4):4672–4683. doi:10.3390/molecules1704467222522397
- Lee HJ, Kim SA, Lee HJ, et al. Paeonol oxime inhibits bFGF-induced angiogenesis and reduces VEGF levels in fibrosarcoma cells. PLoS One. 2010;5(8):e12358. doi:10.1371/journal.pone.001235820808805
- Horng CT, Shieh PC, Tan TW, Yang WH, Tang CH. Paeonol suppresses chondrosarcoma metastasis through up-regulation of miR-141 by modulating PKCdelta and c-Src signaling pathway. Int J Mol Sci. 2014;15(7):11760–11772. doi:10.3390/ijms15071176024992595
- Tamburrino A, Piro G, Carbone C, Tortora G, Melisi D. Mechanisms of resistance to chemotherapeutic and anti-angiogenic drugs as novel targets for pancreatic cancer therapy. Front Pharmacol. 2013;4:56. doi:10.3389/fphar.2013.0005623641216
- Hidalgo M, Alvarez R, Gallego J, et al. Consensus guidelines for diagnosis, treatment and follow-up of patients with pancreatic cancer in Spain. Clin Transl Oncol. 2017;19(6):667–681. doi:10.1007/s12094-016-1594-x27995549
- Peixoto RD, Speers C, McGahan CE, Renouf DJ, Schaeffer DF, Kennecke HF. Prognostic factors and sites of metastasis in unresectable locally advanced pancreatic cancer. Cancer Med. 2015;4(8):1171–1177. doi:10.1002/cam4.45925891650
- Barrallo-Gimeno A, Nieto MA. The snail genes as inducers of cell movement and survival: implications in development and cancer. Development. 2005;132(14):3151–3161. doi:10.1242/dev.0190715983400
- Aigner K, Dampier B, Descovich L, et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene. 2007;26(49):6979–6988. doi:10.1038/sj.onc.121050817486063
- Gaianigo N, Melisi D, Carbone C. EMT and treatment resistance in pancreatic cancer. Cancers (Basel). 2017;9(9):122. doi:10.3390/cancers9090122
- Nakajima S, Doi R, Toyoda E, et al. N-cadherin expression and epithelial-mesenchymal transition in pancreatic carcinoma. Clin Cancer Res. 2004;10(12 Pt 1):4125–4133. doi:10.1158/1078-0432.CCR-0578-0315217949
- Mrozik KM, Blaschuk OW, Cheong CM, Zannettino ACW, Vandyke K. N-cadherin in cancer metastasis, its emerging role in haematological malignancies and potential as a therapeutic target in cancer. BMC Cancer. 2018;18(1):939. doi:10.1186/s12885-018-4845-030285678
- Wang S, Huang S, Sun YL. Epithelial-mesenchymal transition in pancreatic cancer: a review. Biomed Res Int. 2017;2017:2646148. doi:10.1155/2017/264614829379795
- David CJ, Huang YH, Chen M, et al. TGF-beta tumor suppression through a lethal EMT. Cell. 2016;164(5):1015–1030. doi:10.1016/j.cell.2016.01.00926898331
- O’Connor JW, Gomez EW. Biomechanics of TGFbeta-induced epithelial-mesenchymal transition: implications for fibrosis and cancer. Clin Transl Med. 2014;3:23. doi:10.1186/2001-1326-3-2325097726
- Moustakas A, Heldin CH. Mechanisms of TGFbeta-induced epithelial-mesenchymal transition. J Clin Med. 2016;5:7. doi:10.3390/jcm5070063
- Xu W, Yang Z, Lu N. A new role for the PI3K/Akt signaling pathway in the epithelial-mesenchymal transition. Cell Adh Migr. 2015;9(4):317–324. doi:10.1080/19336918.2015.101668626241004
- Haque S, Morris JC. Transforming growth factor-beta: a therapeutic target for cancer. Hum Vaccin Immunother. 2017;13(8):1741–1750. doi:10.1080/21645515.2017.132710728575585
- Zhang L, Tao L, Shi T, et al. Paeonol inhibits B16F10 melanoma metastasis in vitro and in vivo via disrupting proinflammatory cytokines-mediated NF-kappaB and STAT3 pathways. IUBMB Life. 2015;67(10):778–788. doi:10.1002/iub.143526452780
- Sun GP, Wang H, Xu SP, et al. Anti-tumor effects of paeonol in a HepA-hepatoma bearing mouse model via induction of tumor cell apoptosis and stimulation of IL-2 and TNF-alpha production. Eur J Pharmacol. 2008;584(2–3):246–252. doi:10.1016/j.ejphar.2008.02.01618329639
- Ellenrieder V, Hendler SF, Boeck W, et al. Transforming growth factor beta1 treatment leads to an epithelial-mesenchymal transdifferentiation of pancreatic cancer cells requiring extracellular signal-regulated kinase 2 activation. Cancer Res. 2001;61(10):4222–4228.11358848
- Wang H, Wu J, Zhang Y, et al. Transforming growth factor beta-induced epithelial-mesenchymal transition increases cancer stem-like cells in the PANC-1 cell line. Oncol Lett. 2012;3(1):229–233. doi:10.3892/ol.2011.44822740886
- Lebrun JJ. The dual role of TGFbeta in human cancer: from tumor suppression to cancer metastasis. ISRN Mol Biol. 2012;2012:381428.27340590
- Kashima R, Hata A. The role of TGF-beta superfamily signaling in neurological disorders. Acta Biochim Biophys Sin (Shanghai). 2018;50(1):106–120. doi:10.1093/abbs/gmx12429190314
- Pardali E, Ten Dijke P. TGFbeta signaling and cardiovascular diseases. Int J Biol Sci. 2012;8(2):195–213. doi:10.7150/ijbs.8.19522253564
- Jakowlew SB. Transforming growth factor-beta in cancer and metastasis. Cancer Metastasis Rev. 2006;25(3):435–457. doi:10.1007/s10555-006-9006-216951986
- Nie D, Sun H, Shi Z, Zhou X. The effects of combination of paeonol and PNS on TGF-β/smad pathway in ventricular remodeling after myocardial infarction in rats. Tianjin Med J. 2016;44(4):449–452.
- Wu S, Liu L, Yang S, et al. Paeonol alleviates CCl4-induced liver fibrosis through suppression of hepatic stellate cells activation via inhibiting the TGF-beta/smad3 signaling. Immunopharmacol Immunotoxicol. 2019;41(3):438–445. doi:10.1080/08923973.2019.161342731119954