
Abstract
Regulatory T cells (Tregs) are important members of the immune system regulating the host responses to infection and neoplasms. Tregs prevent autoimmune disorders by protecting the host-cells from an immune response, related to the peripheral tolerance. However, tumor cells use Tregs as a shield to protect themselves against anti-tumor immune response. Thus, Tregs are a hurdle in achieving the complete potential of anti-cancer therapies including immunotherapy. This has prompted the development of novel adjuvant therapies that obviate their negative effects thereby enhancing the therapeutic efficacy. Our earlier studies have shown the efficacy of the glycolytic inhibitor, 2-deoxy-D-glucose (2-DG) by reducing the induced Tregs pool and enhance immune stimulation as well as local tumor control. These findings have suggested its potential for enhancing the efficacy of immunotherapy, besides radiotherapy and chemotherapy. This review provides a brief account of the current status of Tregs as a component of the immune-biology of tumors and various preclinical and clinical strategies pursued to obviate the limitations imposed by them in achieving therapeutic efficacy.
Video abstract
Point your SmartPhone at the code above. If you have a QR code reader the video abstract will appear. Or use:
Introduction
Cancer accounts for the major cause of death after cardiovascular disorders worldwide.Citation1 Cancer primarily is a disease that arises due to the deregulation of the growth of functionally matured (somatic) cells leading to a state of “malignant” behavior, which is reflected in the well-established hallmarks of the disease as described by Hanahan and Weinberg.Citation2 Several pioneering studies over the past few years have established immune evasion as one of the key events for the successful establishment of tumors.Citation3 Cancer cells modulate several pathways leading defective antigen presentation, secretion of immunosuppressive mediators, tolerance and immune deviation, apoptosis and release of immunosuppressive cells to evade immune responsesCitation4 (). Recruitment of immunosuppressive cells like myeloid-derived suppressor cells (MDSCs), tumor-derived macrophages, modulated dendritic cells (DCs) and T-regulatory cells (Tregs), are important mechanisms underlying the immune evasion achieved by cancer cells. Among these immunosuppressive cells, the master regulatory cells, Tregs not only secrete molecules that promote initiation and progression of tumors, but also induce neoangiogenesis facilitating metastasis.Citation5–Citation7 The role of Tregs has also been well established in pathogenic infections and allergic response.Citation8,Citation9 Despite more than 20 years of their identification, unraveling of their role in many disease states, the precise mechanisms underlying their suppressive function remains to be completely understood.Citation10 In a disease state such as cancer, Tregs become an impediment as they compromise the anti-tumor response of the host by dampening the efficiency of T-effector (Teff) cells. Therefore, maintaining an optimum balance between Treg and Teff cells is vital in not only avoiding autoimmunity, but also keeping in check the progression of malignancy, avoiding therapeutic resistance, leading to better prognosis of patients (). Emerging evidences suggest that the avoidance of tumor cell death from therapeutic agents is linked to the up-regulation of the Treg pool and escape from immune response.Citation11–Citation18 Therefore, therapeutic approaches, which also modify Tregs appear to be successful in the management of tumors. Several mechanisms appear to be involved in Treg-mediated immunosuppression including the down-regulation of MHC complexes, shedding of antigens, induction of immune checkpoints like programmed death protein 1 (PD-1) and cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), reduction in co-stimulatory molecules (GITR and OX40), release of various cytokines and factors such as IL-10, VEGF, TGF-β, indoleamine 2,3 dioxygenase (IDO).Citation19 Hence, targeting Tregs associated mechanisms have been considered a major strategy in immunotherapy. Many agents such as ipilimumab (anti-CTLA-4 antibody, brand name: Yervoy), which are in different phases of pre-clinical and clinical trials for metastatic renal cell carcinoma and other cancers, are also known to target Tregs.Citation20
Figure 1 Model of immune evasion by tumor cells. Cancer cells modulate several pathways leading to defective antigen presentation, secretion of immunosuppressive mediators [immunosuppressive cytokines like IL-10, vascular endothelial growth factor (VEGF), transforming growth factor (TGF-β), immunosuppressive enzymes like indoleamine 2,3 dioxygenase (IDO), etc], tolerance and immune deviation, apoptosis and release of immunosuppressive cells (Treg cells), which evade immune responses by induction of immune checkpoints like PD-1 and CTLA-4, absence of co-stimulatory molecules like GITR and OX40. These are some of the primary mechanisms involved in tumor cells mediated immune evasion.
![Figure 1 Model of immune evasion by tumor cells. Cancer cells modulate several pathways leading to defective antigen presentation, secretion of immunosuppressive mediators [immunosuppressive cytokines like IL-10, vascular endothelial growth factor (VEGF), transforming growth factor (TGF-β), immunosuppressive enzymes like indoleamine 2,3 dioxygenase (IDO), etc], tolerance and immune deviation, apoptosis and release of immunosuppressive cells (Treg cells), which evade immune responses by induction of immune checkpoints like PD-1 and CTLA-4, absence of co-stimulatory molecules like GITR and OX40. These are some of the primary mechanisms involved in tumor cells mediated immune evasion.](/cms/asset/4e1f77c5-273c-4e86-806f-e92bf586377d/dcmr_a_12186856_f0001_b.jpg)
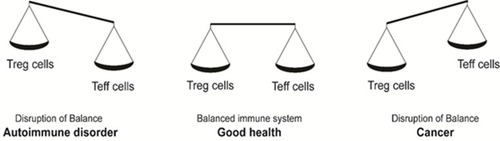
Figure 2 Imbalances in the immune system homeostasis results in a disease state. A balance in the levels of Treg and T effector cells maintains the homeostatic and disease-free state. A shift in the balance towards Tregs causes a decrease in anti-cancer immunity, resulting in cancer. Contrarily, a shift in the balance towards T effector cells causes a decrease in Treg levels and T effector cells hyperactivation leading to auto-immune disorders.
We present here an overview on the existing knowledge about the role of Tregs in tumorigenesis as well as merits and limitations of approaches using conventional chemotherapeutic agents that target Tregs for improving therapeutic gain. Some new agents that also target Tregs and show negligible or absence of any side effects on normal cells are also discussed ().
Table 1 Current Status Of Anti-Cancer Therapies Influencing Treg Levels
Tregs Characterization And Immune Evasion
Treg cells (CD4+CD25+FoxP3+) belong to the family of CD4+ T cells. These cells have a high expression of CD25 (IL-2 receptor) and transcription factor Foxp3 (Forkhead box P3). FoxP3 plays an important role in the generation and production of Tregs, including the maintenance of their functionality; loss of which is associated with immune dysregulation and lymphoproliferative diseases both in mice and humans.Citation21 Tregs have two subtypes on the basis of their origin: naturally occurring Tregs (n-Tregs) and inducible Tregs (i-Tregs). n-Tregs are mainly formed in thymus and need costimulatory molecules for their development and lineage commitment.Citation18 n-Tregs suppress immune effector lymphocytes like T helper cells (Th cells), Th1, Th2, Th17 and follicular Th cells (Tfh cells) in a contact-dependent manner that requires Granzymes B/perforin and Fas/FasL pathways.Citation12 i-Tregs are generated in the periphery and do not require costimulatory molecules for activation, but at least require T cell receptor (TCR), TGF-β and IL-2 under a variety of pathological conditions.Citation8 i-Tregs mediate immunosuppression in multiple ways that involve the secretion of immunosuppressive cytokines (interleukins, IL-10, IL-35, and TGF-β), granzymes-induced cytolysis, metabolic reprogramming of effector cells, and DC-mediated suppression.Citation22,Citation23
In vivo studies in mice models indicate that Tregs regulate concomitant immunity and cross-reactive anti-tumor immunity.Citation24,Citation25 Tregs not only suppress the natural killer (NK) cell-mediated cytotoxicity but also check the proliferation of CD4+ and CD8+ T-cells and inhibit the interferon (IFN)-γ secretion by immune cells, thereby leading to the impairment of effective anti-tumor immune response.Citation20 Indeed, higher Tregs activity has been related to decreased survival and poor prognosis in patients of breast cancer, gastric carcinoma, non-small cell lung cancer (NSCLC), squamous cell carcinoma of head and neck (SCCHN), pancreatic cancer and ovarian cancer.Citation17,Citation26–Citation30
Anti-tumor immunity can be reinforced by the reduction of Treg cells, which are responsible for suppressing immune-response against syngeneic tumors in-vivo and in-vitro.Citation31,Citation32 Several therapeutic agents besides having a direct anti-tumor function also leads to a reduction in Treg numbers by inhibiting the immune checkpoints, CD25 and several chemokine receptors (e.g. CCR5), which induce intra-tumoral recruitment of Tregs.Citation31,Citation33–Citation35 Similarly, the administration of anti-folate receptor monoclonal antibodies (mAbs) also effectively reduces the number of Tregs.Citation36 Anti-GITR or anti-OX40 agonist antibodies based immunotherapy also stimulates anti-tumor immune response via blocking Treg-induced immuno-suppression, resulting in the eradication of established tumors.Citation37,Citation38 Moreover, many chemotherapeutic agents have also been found to deplete Tregs (aromatase inhibitor, cyclophosphamide, mitoxantrone, etc).Citation39–Citation41 Recent studies from our laboratory have demonstrated that glycolytic inhibitor, 2-deoxy-D-glucose (2-DG), which inhibits metabolic modulation in tumors, reduces the i-Tregs frequency. Therefore, 2-DG can be used in combination with immunotherapy in addition to its recognized role as an adjuvant for chemo- and radio-therapies.Citation42,Citation43 Hence, these studies reveal Tregs as a potential target for restoring anti-tumor immunity, thereby improving the anti-tumor response.
Factors Influencing Treg Level, Activity, And Intra-Tumoral Migration
Identification of factors that affect the Treg level, their activity, and migration is essential for developing strategies that target them. Expression of Foxp3 is indispensable for Treg development. Foxp3 when expressed ectopically in conventional T cells can generate a suppressive phenotype and Foxp3 gene mutation results in Treg cells deficiency that leads to severe autoimmune disorders in mice and humans.Citation44 Foxp3 gene locus contains several conserved noncoding sequences (CNS), which are involved with different signaling pathways.Citation45 CNS0 is involved in the activation of Treg-specific super-enhancers involved in Foxp3 expression. CNS1 regulates TGF-β-mediated Foxp3 expression in the periphery. It has a binding site for a nuclear factor of activated T cells and activator protein 1 that regulates TGF-β signaling. CNS2 has binding sites for transcription factors like STAT5, RUNX, cAMP response element-binding protein and is stimulated by TCR expression and IL-2, maintaining a high stability and activity of Foxp3 in Tregs.Citation44,Citation45 CNS3 has binding sites for c-Rel and other transcription factors that mediate expression of Foxp3 during Treg differentiation. Epigenetic regulation of these CNS, mainly demethylation, is important for Foxp3 expression during Treg differentiation.Citation46–Citation53 Thus, genetic and epigenetic factors regulate the Foxp3 level thereby play an important role in Treg levels and activity. Further, a highly dynamic T cell metabolism has a tremendous impact on the ability of T cells to grow, activate and differentiate. Recent observations suggest that Teff cells and Tregs require completely distinct metabolic pathways for their proliferation and activity.Citation54 T effector cells are highly glycogenic and have high Glut-1 levels on their surface.Citation55 In mice and humans, the high glycolytic rate is due to hyper-activation of the mTOR signaling pathway and is operative during nutrient-deprived conditions.Citation54,Citation56 Contrarily, Tregs have higher metabolic flexibility compared to CD4+ Teff cells. During Treg proliferation enhanced glycolysis not only provides energy in the form of ATP and NADH but also relays its metabolic intermediates to nucleotide biosynthetic pathways.Citation54 Also, during low energy conditions, Tregs rely on fatty acid oxidation (FAO), due to the increased level of carnitine palmitoyltransferase 1a (CPT1a), which is critical for FAO as a rate-limiting enzyme. This facilitates the acyl groups to enter into the mitochondria thereby suggesting that Tregs use multiple metabolic pathways needed for extensive proliferation. Apart from development and proliferation, the intra-tumoral migration of Tregs also contributes to its immunosuppressive activity. Tumors secret several chemokines like C-C motif chemokine ligand 22 (CCL22), which is a ligand to C-C chemokine receptor (CCR) 4 present on Tregs, thereby attracting them near to the tumor tissue as observed in human ovarian cancers.Citation52,Citation57–Citation60 Several of these factors have been taken into consideration to develop approaches that target the immunosuppressive Tregs. We present here the existing approaches for reducing Treg activity as well as their numbers ().
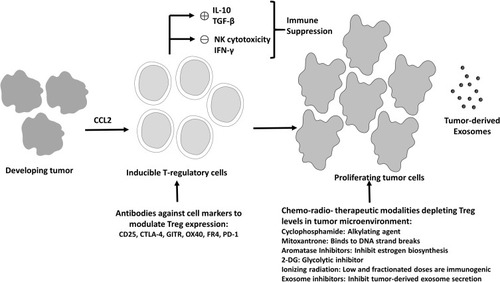
Figure 3 Current Treg therapeutics targeting cancer. Treg cells mediated immunosuppression orchestrated by immunosuppressive mediators like IL-10 and TGF-β leads to reduction in antitumor immunity (IFN-γ production and NK cells induced tumor cell cytotoxicity), resulting in excessive tumor cells proliferation. Several strategies such as antibodies targeting the important molecules for Treg activity or small molecule inhibitors, ionizing radiation and exosome inhibitors, which reduce the Treg cell number and activity represent the currently available modalities for Treg depletion and enhanced anti-tumor activity.
Approaches For Regulating Treg-Mediated Tumor Control
Antibody-Based Treg Modulation
Antibody Against CD25
The alpha chain of heterotrimeric IL-2 receptor complex, CD25 has an inevitable role in both the Treg development and their activity and can, therefore, be exploited as a potential target against Tregs.Citation22 IL-2 is generally essential for proliferation of the activated T cells. Due to the upregulated expression of CD25 on Tregs, most of the IL-2 present in the milieu is utilized by the Tregs present in the vicinity, thereby preventing Teff cells from proliferation and sustained survival.Citation61 The depletion of CD25 positive Treg population via targeted antibody therapy enhanced the anti-tumor immunity that correlated well with the progressive reduction of tumor volume in murine cancer models.Citation62,Citation63 A phase-I/II study with daclizumab (that blocks CD25) in combination with the DC vaccine has shown a transient but complete depletion of CD25high cells in melanoma patients.Citation64 Similarly, in metastatic melanoma patients (with lymphopenia induced via temozolomide), anti-CD25 antibody depleted Tregs significantly without impairing Teff cell functions, thereby augmenting the anti-tumor immune response.Citation65 Administration of daclizumab has also been found to decrease Tregs in the peripheral blood mononuclear cells (PBMCs) of patients with melanoma.Citation66 Since, CD25 is also present on the Teff cells, targeting Tregs with CD25 blockade is also expected to collaterally deplete Teff cells due to IL-2 deprivation, which may lead to opportunistic infections in patients receiving the treatment.Citation67 Furthermore, daclizumab has a short half-life of 20 days and therefore has a transient effect, which is reversible under in-vitro conditions.Citation61 Other limitations of CD25 blockade include side effects such as severe acute hypersensitivity reactions, cytokine release syndrome, infections, and local skin reactions.Citation68 Daclizumab marketed for relapsed multiple sclerosis has been withdrawn from the market due to deadly episodes of encephalitis and meningoencephalitis.Citation69 Interestingly, it has been reported that treating metastatic melanoma patients with anti-CD25 RFT5-SMPT-dgA (IMTOX25) leads to a significant but transient decrease in Treg numbers. However, the desired anti-tumor activity was not observed.Citation70 Despite the preliminary success, many questions still need to be addressed before developing a successful therapeutic strategy exploiting CD25 as a potential target for Tregs depletion.
Antibody Against CTLA-4
CTLA-4 is constitutively expressed in Treg cells but only upregulated in conventional T cells after activation, a phenomenon which is particularly notable in cancers.Citation71 CTLA-4 plays an indispensable function in blocking CD28 (a co-stimulatory receptor on Teff cells) interaction with B7 that is present on antigen-presenting cells (APCs).Citation72,Citation73 The expression of CTLA-4 increases after its binding with B7, delivering a signal that suppresses the proliferation of T-cells.Citation74 Another aspect of CTLA-4 function, which hinders effective anti-tumor immunity is an increase in the levels of IDO in DCs. This consequently leads to the metabolism of tryptophan, producing kynurenines and picolinic acid, thereby abrogating the proliferation and function of Teff cells.Citation75 Possibly Tregs exploit this property of CTLA-4 for implementing immunosuppression as they constitutively express CTLA-4.Citation76,Citation77 Synergistic effects of antibodies against CTLA-4 in combination with other anti-tumor therapeutics are well documented in the recent years.Citation78,Citation79 CTLA-4 deficiency results in lymphoproliferation followed by splenomegaly, development of autoimmune diseases and increased IgE secretion in Treg-specific Foxp3+ CTLA-4 deficient mice, similar to Foxp3 deficient mice.Citation33 Thus, inhibition of CTLA-4 has been found to increase the immune response against pre-established tumors as well as to effectively suppress a secondary tumor challenge i.e. to enhance memory response.Citation80 Co-culturing Tregs with T cells that lack CTLA-4 shows that the absence of CTLA-4 abrogates the immunosuppressive activity of Tregs.Citation72 Treatment of patients having metastatic melanoma with anti-CTLA-4 in a Phase-III clinical trial has shown improved overall survival (OS) with ipilimumab (3 mg/kg), although adverse immune effects were observed at higher doses.Citation34 Similarly, in another clinical trial, targeting CTLA-4 was found to improve the OS but was associated with undesirable toxicity such as autoimmune response (mostly inflammatory bowel disease).Citation81 At present, it is unclear if anti-CTLA-4 treatment enhances anti-tumor immunity by inhibiting the functions of the Tregs or by increasing Teff cells activity and needs further investigations.
Antibody Against Programmed Death Protein 1 (PD-1)
PD-1, a coinhibitory receptor is present on CD4+, CD8+ T cells, and B cells, activated by PDL-1 and PDL-2 ligands. Blocking anti-PD1 antibodies have durable anti-cancer effects and been approved for ipilimumab resistant metastatic melanoma, non-small cell lung cancer (NSCLC), metastatic renal cell carcinoma (RCC), Hodgkin’s Lymphoma (HL), metastatic urothelial carcinoma, metastatic colorectal cancer.Citation82 High PD-1 expression in Tregs is of paramount importance due to their role in blocking CD8+ mediated anti-tumor immunity.Citation21,Citation83 High PD-1 expression on Tregs resulted in the suppression of CD8+ T cell function in various organs like lung, spleen and draining lymph nodes of prostate tumor-bearing mice.Citation84 Further, PD-1 increases the affinity of Tregs to TGF-β-mediated signals and induces the differentiation of naive T cells into Treg cells.Citation85 However, the relationship between PD-1 expression and Treg is still not well understood as a large subset of patients in clinical trials did not respond to PD-1 blockade. The anti-PD-1 antibody (nivolumab) showed an excellent 18–28% objective therapeutic response in patients with advanced non-small-cell lung carcinoma, melanoma, and renal cell carcinoma.Citation86 Interestingly, patients that were cured and showed no signs and symptoms of the disease exhibited high Treg levels after nivolumab therapy while relapsing patients did not.Citation21,Citation87 The differential response was attributed to high pSTAT3 (phospho-signal transducer and activator of transcription) level and IL-10 production following nivolumab.Citation87 Thus, the relationship between anti-PD-1 blockade based immunotherapy and Treg levels need extensive investigation in cancer patients including their response to combinational therapies involving inhibitors of immune checkpoint (agents that inhibit the activity of proteins involved in the suppression of anti-tumor immunity like PD-1 and CTLA-4).Citation88
Antibody Against GITR
GITR, (glucocorticoid-induced TNFR family related gene) is a member of the TNFR superfamily (TNFRSF) and a co-stimulatory molecule that is expressed in resting CD4+ and CD8+ population and increases in expression upon T-cell activation.Citation89 It has also been shown that GITR is expressed constitutively at a high level in TregsCitation36,Citation90 and treatment with anti-GITR agonistic mAb reduces the suppressive activity of Tregs.Citation91 A study in GITR knockout mice, which lack GITR signaling, reported that the responder CD4+CD25− population becomes resistant to the suppressive CD4+CD25+ population.Citation91 Furthermore, the use of anti-GITR mAb has been shown to elevate CD4+ and CD8+ T cell response against tumor cells, an effect especially seen in large tumors.Citation88,Citation92 A combination treatment with anti-GITR antibody and tumor antigen stimulation augments the proliferation of “antigen-primed” Teff cells and furthermore makes them resistant to CD4+CD25+ Tregs mediated-suppression.Citation38 A humanized anti-GITR mAb, TRX518, which is under clinical trials, has shown encouraging results by augmenting immune responses against tumors in murine cancer models.Citation93 Due to a relatively ubiquitous prevalence of GITR on T lymphocyte population including on Teff cells, a local administration of anti-GITR mAb to target Tregs at the site of tumor may be the preferred method of treatment since an intravenous administration may lead to increased autoimmune response.Citation94 In total, anti-GITR therapy is a promising candidate for cancer immunotherapy through depleting Tregs but further investigations for clinical response, dosage, and use in combination therapy as well as normal tissue toxicity are needed.
Antibody Against OX40
OX40 (CD134), is a co-stimulatory molecule that is expressed transiently on activated T-cells and constitutively on CD4+CD25+ Tregs.Citation95–Citation98 Agonistic mAb against OX40 has been shown to reduce CD4+CD25+ Tregs-mediated immunosuppression.Citation37,Citation99 An intra-tumoral injection of anti-OX40 mAb induces tumor rejection in mice, an effect abrogated by CD8+ T cell depletion.Citation99 Further, the activity of CD25+CD4+ Tregs and CD25−CD4+ cells is altered in OX40 deficient conditions. Moreover, intra-tumoral anti-OX40 injection induces the migration of infiltrating DCs to draining lymph nodes and deploys a population of newly-formed cytotoxic T-lymphocytes (CTLs) that are tumor-specific.Citation99 Recently, an intra-tumoral injection of agonist OX86 mAb that stimulates OX40 significantly reduced the immunosuppression mediated by the Tregs thereby increasing an activation of Teff cells in a murine cancer model.Citation100 Moreover, the tumor-infiltrating Tregs produced less IL-10 in response to the treatment, besides resulting in DC maturation (in-vitro) and DC migration (in-vivo), leading to augmented tumor immunity. Administration of anti-OX40 fusion protein OX40-Fc has been found to result in the regression of murine sarcoma, supporting this proposition.Citation101 The combined use of cyclophosphamide and anti-OX40 mAb has shown a significant increase in immune responses against tumors by depleting Tregs in tumor vicinity and simultaneously inducing an influx of CD8+ Teff cells in B16 melanoma murine models.Citation37 Interestingly, combination therapy increased the number of peripheral Tregs but depleted intra-tumoral Tregs, thereby suggesting that tumor regression largely relies on the depletion of Tregs present in the tumor microenvironment (TME).Citation37 However, another study using a combination therapy with checkpoint inhibitor antibodies (Abs) like PD-1 showed a negative effect on the anti-tumor activity of OX40 agonist Ab, emphasizing the need for the appropriate design of immunotherapy combinations.Citation102 Thus, anti-OX40 therapy enhances the anti-tumor immunity by altering the suppressive function of CD25+CD4+ Tregs although further investigations to establish it as an immunotherapeutic modality are still needed.
Antibody Against FR4
Folate receptor4 or FR4 is a characteristic marker of Rodent Tregs, which express a high level of this receptor. Naïve T cells require folate for protein and nucleic acid synthesis.Citation103 T cell receptor (TCR) stimulation upregulate the expression of FR4 by several folds in Foxp3+ Tregs as compared to Foxp3− T cells.Citation104 Consequently, FR4 molecule can be targeted in anti-FR4 mAb based therapy for specifically depleting suppressive Treg population to promote tumor immunity. However, folate is an important ingredient for proliferation of cells and is, therefore, an essential nutrient.Citation105 Therefore, research on the human counterpart of rodent FR4 is necessary to determine its potential as a target for cancer immunotherapy.
Chemotherapeutic Agents
Several small-molecule chemotherapeutic agents and adjuvant have been found to exert an off-target effect on Tregs that correlate well with their overall anti-cancer efficacy. These include both DNA targeting drugs as well as those that target the biosynthetic pathways or metabolic reprogramming of tumors. In the case of some drugs, the effects on Tregs have been observed at doses significantly lower than the doses used for affecting the primary targets.
Cyclophosphamide
Cyclophosphamide is a DNA crosslinking agent and a widely used anti-cancer drug.Citation106,Citation107 Cyclophosphamide decreases the immune system response and is also used in the treatment of autoimmune disorders such as granulomatosis with polyangiis.Citation108 Cyclophosphamide shows a differential dose-response concerning immune response, where a high dose required for effective anti-tumor chemotherapy may be associated with immunosuppression while a low dose has been demonstrated to show increased anti-tumor immunity in mice models and metastatic melanoma patients.Citation109,Citation110 The low dose immune-stimulatory property of cyclophosphamide was first realized when the regression was observed in a cyclophosphamide-resistant lymphoma (L5178Y) in mice.Citation111,Citation112 Many studies have demonstrated that cyclophosphamide depletes Tregs in normalCitation109 and tumor-bearing mice as well as in rats-bearing chemically induced colon cancer (PROb).Citation25,Citation106,Citation113 However, cyclophosphamide does not specifically deplete Tregs (CD4+CD25+), since it also reduces T cells (CD4+CD25−).Citation114 Interestingly, continuous administration of cyclophosphamide at low doses has been shown to inhibit the renewal of Tregs in multiple myeloma-bearing mice and restore the anti-tumor immunity thereby prolonging the overall survival and preventing the tumor-recurrence.Citation115 We demonstrated that lower doses of cyclophosphamide enhanced radiosensitization by a glycolytic inhibitor 2-DG that could be linked to its ability to deplete iTregs.Citation116,Citation117 Reduced Treg infiltration with an increase in CD8+ T cells has been detected in patients with metastatic carcinoma that were treated with cyclophosphamide and immunotherapy (intra-tumor BCG).Citation118 Recent studies have shown that it is the CD8+ resident DCs that activate Tregs, and the anti-tumor response triggered by cyclophosphamide is due to selective depletion of the CD8+ resident DCs.Citation119 However, selective depletion of CD8+ resident DCs by cyclophosphamide might compromise the immunity of host when exposed to an infectious pathogen as CD8+ resident DCs serve as APCs for exogenous antigens. In a phase-II trial, progression-free survival was shown in more than 50% of the ovarian cancer patients treated with a combination of bevacizumab and low dose cyclophosphamide (50 mg/day), while trials in hepatocellular carcinoma patients using a low dose of cyclophosphamide impaired Treg suppression and unmasked the Teff cell response against α-fetoprotein in 6 out of 13 patients.Citation120,Citation121 The capability of cyclophosphamide to inhibit the suppressive functions of Treg cells, leading to enhanced tumor immunity, entitles it to be an effective chemotherapeutic agent or an adjuvant.Citation107
Mitoxantrone
Mitoxantrone is a DNA binding anthracenedione that binds to deoxyribose sugar leading to a strand break.Citation122 Another marked effect of mitoxantrone, which augments anti-cancer immunity, is through cell surface expression of calreticulin in cancer cells, which in turn promotes phagocytosis of dying cancer cells through DCs.Citation123 Treatment of breast cancer patients with mitoxantrone does not seem to significantly change the CD4+/CD8+ ratio, but lead to depletion of B lymphocytes and Treg population.Citation40 Therefore, mitoxantrone can be combined with other modalities to formulate an effective immunotherapeutic regimen for cancer treatment. However, it should be noted that these effects of mitoxantrone could be non-specific and therefore requires further investigations before it can be used for cancer immunotherapy.
Aromatase Inhibitors
The aromatase enzyme plays a key role in estrogen biosynthesis. Estrogen has been attributed to the promotion of immune tolerance by augmenting Treg proliferation in humans and mice.Citation124,Citation125 It is well established that Tregs infiltration in tumors results in a shorter survival in breast cancer patients.Citation126 Therefore, the blocking of estrogen receptor α-signaling by aromatase inhibitors can be exploited as a therapeutic strategy, especially in estrogen-dependent breast cancer.Citation126 In a phase-II trial, 83 patients with breast cancer were treated with letrozole, an aromatase inhibitor. The results showed that reduced infiltrating Treg numbers in biopsies correlate well with higher immune response in patients.Citation127 Furthermore, in a controlled phase-II trial, letrozole-treated patients with breast cancer showed a marked reduction of tumor-infiltrating Tregs in the primary tumor post-therapy. However, combining cyclophosphamide with letrozole did not further affect the frequency of Tregs in the tumor.Citation127 Furthermore, adverse effects on cognitive functioning have been observed in patients administered with letrozole.Citation128 Thus, these evidences suggest that aromatase inhibitors cause immunomodulatory events resulting in a targeted decrease in proliferation of tumor infiltrating Tregs and therefore have immunotherapeutic potential but their toxic effects need further consideration.
2-Deoxy-D-Glucose (2-DG)
Metabolic reprogramming (MRP) has emerged as one of the vital features of tumors, wherein the tumor cells reprogram their metabolism by switching to glycolysis despite adequate oxygen rather than the much more efficient oxidative phosphorylation pathway, which is known as the Warburg effect.Citation129 MRP purportedly helps in the survival and growth of tumor cells by augmenting energy production, macromolecular (DNA, protein etc.) synthesis and defense against oxidative stress. By targeting this phenotype, among the various glycolytic inhibitors, 2-DG is well established for its cytotoxic and radio/chemo-sensitizing effects on the cancer cells, including limited clinical trials.Citation130,Citation131 Multiple mechanisms underlying this sensitization by 2-DG have been elucidated, which include energy (mainly ATP) crunch, altered anti-oxidant defense, enhanced unfolded protein response (UPR), inhibition of DNA repair, impaired cell cycle regulation, altered calcium influx, and apoptosis.Citation132 Studies in murine tumors have shown enhanced tumor regression and tumor-free survival following treatment with 2-DG and radiation.Citation133,Citation134 Moreover, in phase-I-III clinical trials, negligible normal tissue toxicity, enhanced survival and significant improvement in the quality of life has been observed in cerebral glioma patients following administration of 2-DG with hypofractionated radiotherapy.Citation40,Citation133–Citation137
Results from murine studies have shown that the response of the tumors to the combined treatment of 2-DG and irradiation is heterogeneous with complete (cure; tumor-free survival) and partial (growth delay) response, which correlates well with the immune stimulation, switch of Th2 cells to Th1 cells and a significant reduction in Treg numbers in spleen, peripheral blood, lymph node and tumor.Citation133,Citation134,Citation138 However, significant effects on Treg cells was not observed in the non-tumor bearing mice, suggesting a differential effect of 2-DG on Tregs in normal and tumor-bearing mice. Recently, glycolysis has been shown to regulate the induction of Tregs (iTregs) by modulating the expression of FOXP3 exon2 splice variants.Citation139 Moreover, several studies have shown that 2-DG protects the normal cells from radiation- and chemotherapy-induced damage.Citation140 Optimal TCR stimulation of CD4+ T cells in the presence of glycolysis results in the autocrine secretion of IL-2 and generation of a limited number of Tregs in healthy conditions.Citation141 On the other hand, suboptimal stimulation of TCR in CD4+ T cells results in an enhanced Treg activity via the generation of Foxp3-E2 isoform.Citation140 Treatment with 2-DG or impaired glycolytic conditions such as relapsed, remitting multiple sclerosis reduces the abundance of Foxp3-E2 isoform, necessary for the Treg function.Citation139 Since suboptimal TCR engagement occurs in most tumors, 2-DG appears to be a good component for the combination therapy. Thus 2-DG directly sensitizes tumor cells by compromising the energy dependent repair and recovery processes, and also reduces immune tolerance by reducing Tregs. Although, 2-DG is a well-known adjuvant for radio- and chemotherapy of cancer, further studies are required to assess its combination with immunotherapies.Citation132
It is likely that other anti-cancer therapeutics, besides the one discussed above may also have effects on Tregs, which may partly contribute to their overall efficacy. However, considering them as immune targeting agents is less realistic compared to other immune modulators like checkpoint inhibitors such as anti-PD-1 and anti-CTLA-4 antibodies. Besides, several such agents have shown non-specific effects on Tregs and need further investigations before they can be used to target Tregs.
Effects Of Ionizing Radiation On Tregs
Radiation enhances anti-tumor immunity which is associated with alterations in tumor cell phenotype and increased activity of immune cells like CD4+, CD8+ T cells, macrophages, APCs, NK cells, etc. Irradiation of tumors releases tumor-specific antigens leading to the immune recognition of tumor-related new peptides by increasing major histocompatibility complex-I molecules (MHC-I), besides up-regulating the expression of Fas antigen, to activate the T-cell mediated cytotoxicity.Citation12,Citation142,Citation143 Radiation up-regulates the expression of various adhesion molecules like vascular cell adhesion protein 1 (VCAM-1), E-selectin, and intercellular adhesion molecule 1 (ICAM-1) on endothelial cells, enhancing the leukocyte adhesion and migration, leading to alterations in the TME.Citation144 Contrarily, radiation enhances the proliferative capacity and activity of Tregs at therapeutic doses leading to immunosuppression and tumor relapse. Recent studies suggest that the major mechanism involved in radio-resistance of Tregs is intrinsically through the high expression of GITR and extrinsically through radiation-induced production of TGF-β; an important cytokine needed for Treg proliferation and activity.Citation145,Citation146 Stereotactic radiotherapy has been suggested to enhance the functionality of Treg cells in the TME, which is independent of TGF-β and IL33 thereby indicating the involvement of multiple mechanisms in Tregs radioresistance.Citation147 Moreover, Tregs show a differential response to a low and high dose of radiation. CTLA4 was found to be upregulated at a low dose (1.8 Gy) and decreased at high dose (30 Gy) suggesting low dose enhanced Treg cell proliferation and activity whereas high dose abolished the suppressive capacity of Tregs attributed to increased apoptosis and pro-apoptotic protein Bax.Citation12,Citation146 Tregs have been considered as a major hurdle in realizing the efficacy of radiotherapy and therefore Treg cell-based immunotherapy is recommended in combination with radiation to achieve maximum therapeutic gain. STAT3 inhibition in combination with radiation has been recently shown to enhance delay in the tumor growth, which is accompanied by a decrease in Tregs, MDSCs and M2 macrophages while an increase in the effector T cells and M1 macrophages in an orthotopic mouse model of Head and Neck Cancer.Citation147 Blockade of CD25 results in better local tumor control with enhanced T cell-mediated immune responses against the tumor, whereas blockade of CTLA-4 in combination with radiation does not appear to be effective.Citation148 Taken together, a greater caution must be exercised during planning strategies aimed at combining radiation with Treg-targeting agents.
Other Approaches
Exosomes secreted by several cell types contain many inter-cellular communication-related molecules, including components of the immune network, facilitating the communication among various immune cells.Citation149,Citation150 Tumor-derived exosomes contribute to increased status of Tregs through their surface-bound TGF-β1 that mediates Foxp3 expression and the associated suppressive functions in the malignant effusions.Citation151,Citation152 Attenuating the impact of the tumor-derived exosome or controlling their release has been suggested as an immunotherapy strategy in the management of advanced cancer with malignant effusions.Citation152 Since exosomes can contain part of the overall genomic DNA and proteins, targeting them or their effects may have far-reaching consequences, besides Treg related effects.
Limitations Of Approaches Targeting Tregs
Impact of Tregs on anti-tumor immunity can be reduced by their depletion, interference in their trafficking into tumors, or attenuating their differentiation thereby provoking or enhancing anti-tumor immunity, through the development of tumor-specific effector cells. However, the complete ablation of CD25+CD4+ Treg population disturbs natural self-tolerance and leads to several chronic and destructive autoimmune disorders.Citation146–Citation161 Freshly isolated or ex-vivo expanded donor-derived Tregs have been shown to delay or even prevent graft-versus-host disease (GVHD) in murine models of allogeneic bone marrow transplantation, whereas the depletion of Tregs in the transplant results in increased severity of acute GVHD.Citation162–Citation165 Depletion of CD4+CD25+ Treg cells boosts anti-tumor immunity,Citation30,Citation62,Citation63,Citation166 enhances immune responses against invading pathogens, triggers allergic responses to environmental substances and causes autoimmune disorders or uncontrolled pathological immune responses and breaches feto-maternal tolerance during pregnancy.Citation167 More recently apoptotic death of Tregs has been shown to enhance immune suppression through the A2A pathway that is triggered by adenosine (generated from the ATP released by apoptotic cells).Citation168 This suggests that approaches that lead to functional inactivation of Tregs may be more promising in achieving better tumor control and therapeutic gain since eliminating or depleting Tregs by inducing apoptosis may be counterproductive.
Conclusions
Taken together, existing and emerging evidence suggests that the depletion of Treg cells via targeted therapy as well as by chemo-radio-therapeutic modalities result in enhanced anti-tumor immunity and therapeutic outcome. However, these modalities also have normal tissue toxicity as the same target protein or receptor is also shared by other effector T cells. Moreover, their depletion below a particular level may result in the generation of autoimmunity. Therefore, targeting of Tregs induced by tumors should be thoroughly investigated and the threshold of depletion should be carefully taken into consideration. Our recent results suggest that metabolic inhibitors like 2-DG, which can reduce the impact of Tregs with negligible effects on the normal cells, may demonstrate higher success as adjuvants to immunotherapy/radiotherapy to overcome immune suppression by Tregs. Thus, the Treg chariot that the tumor cells ride is a pertinent cellular target in the battlefield of cancer but needs to be cautiously maneuvered.
Abbreviations
2-DG, 2-deoxy-D-glucose; APCs, antigen-presenting cells; CD, cluster of differentiation; CTLs, cytotoxic T lymphocytes; DC, Dendritic cell; FAO, fatty acid oxidation; FR4, folate Receptor 4; Foxp3, Forkhead box P3; GVHD, graft-versus-host disease; IL, Interleukin; IDO, indoleamine 2,3 dioxygenase; i-Tregs, inducible Tregs; mAb, monoclonal antibody; NKT, natural killer T cells; MRP, metabolic reprogramming; n-Tregs, naturally occurring Tregs; PBMC, peripheral blood mononuclear cells; SCCHN, squamous cell carcinoma of head and neck; TCR, T cell receptor; Teff, T-effector; TLRs, toll-like receptors; TGF-β, transforming growth factor-beta; Tregs, T regulatory cells; VEGF, vascular endothelial growth factor.
Author Contributions
All authors contributed to conceptualization of the review, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of this review.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
This work is supported by the grants INM-301 and INM-311 from DRDO, Government of India. BSD thanks Dr. Jiade Lu, and Dr. Xiaodong Wu, Shanghai Proton and Heavy Ion Center, Shanghai, China for their encouragement and support. All authors thank Dr. Abdullah Farooque, who met with an untimely death, for his help and support during the initial conception and drafting of the review.
References
- Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6 ):394–424. doi:10.3322/caac.v68.630207593
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. doi:10.1016/j.cell.2011.02.01321376230
- Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331:1565–1570. doi:10.1126/science.120348621436444
- Cavallo F, De Giovanni C, Nanni P, et al. 2011: the immune hallmarks of cancer. Cancer Immunol Immunother. 2011;60:319–326. doi:10.1007/s00262-010-0968-021267721
- Mougiakakos D, Choudhury A, Lladser A, et al. Regulatory T cells in cancer. Adv Cancer Res. 2010;107:57–117.20399961
- Dwarakanath BS, Farooque A, Gupta S. Targeting regulatory T cells for improving cancer therapy: challenges and prospects. Cancer Rep. 2018;1:e21105. doi:10.1002/cnr2.1105
- Halvorsen EC, Mahmoud SM, Bennewith KL. Emerging roles of regulatory T cells in tumor progression and metastasis. Cancer Metastasis Rev. 2014;33(4 ):1025–1041. doi:10.1007/s10555-014-9529-x25359584
- Akdis M, Blaser K, Akdis CA. T regulatory cells in allergy: novel concepts in the pathogenesis, prevention, and treatment of allergic diseases. J Allergy Clin Immunol. 2005;116:961–968. doi:10.1016/j.jaci.2005.09.00416275361
- Demengeot J, Zelenay S, Moraes-Fontes MF, et al. Regulatory T cells in microbial infection. Springer Semin Immunopathol. 2006;28:41–50. doi:10.1007/s00281-006-0024-516874537
- Shevach EM. Foxp3+ T regulatory cells: still many unanswered questions-A perspective after 20 years of study. Front Immunol. 2018;9:1048. doi:10.3389/fimmu.2018.0104829868011
- Antony PA, Piccirillo CA, Akpinarli A, et al. CD8+ T cell immunity against a tumor/self-antigen is augmented by CD4+ T helper cells and hindered by naturally occurring T regulatory cells. J Immunol. 2005;174:2591–2601. doi:10.4049/jimmunol.174.5.259115728465
- Grauer OM, Nierkens S, Bennink E, et al. CD4+FoxP3+ regulatory T cells gradually accumulate in gliomas during tumor growth and efficiently suppress anti-glioma immune responses in vivo. Int J Cancer. 2007;121:95–105. doi:10.1002/(ISSN)1097-021517315190
- Laronne-Bar-On A, Zipori D, Haran-Ghera N. Increased regulatory versus effector T cell development is associated with thymus atrophy in mouse models of multiple myeloma. J Immunol. 2008;181:3714–3724. doi:10.4049/jimmunol.181.5.371418714048
- Liyanage UK, Moore TT, Joo HG, et al. Prevalence of regulatory T cells is increased in peripheral blood and tumor microenvironment of patients with pancreas or breast adenocarcinoma. J Immunol. 2002;169:2756–2761. doi:10.4049/jimmunol.169.5.275612193750
- Marshall NA, Christie LE, Munro LR, et al. Immunosuppressive regulatory T cells are abundant in the reactive lymphocytes of Hodgkin lymphoma. Blood. 2004;103:1755–1762. doi:10.1182/blood-2003-07-259414604957
- Ormandy LA, Hillemann T, Wedemeyer H, et al. Increased populations of regulatory T cells in peripheral blood of patients with hepatocellular carcinoma. Cancer Res. 2005;65:2457–2464. doi:10.1158/0008-5472.CAN-04-323215781662
- Sasada T, Kimura M, Yoshida Y, et al. CD4+CD25+ regulatory T cells in patients with gastrointestinal malignancies: possible involvement of regulatory T cells in disease progression. Cancer. 2003;98:1089–1099. doi:10.1002/(ISSN)1097-014212942579
- Schaefer C, Kim GG, Albers A, et al. Characteristics of CD4+CD25+ regulatory T cells in the peripheral circulation of patients with head and neck cancer. Br J Cancer. 2005;92:913–920. doi:10.1038/sj.bjc.660240715714205
- Shitara K, Nishikawa H. Regulatory T cells: a potential target in cancer immunotherapy. Ann N Y Acad Sci. 2018;1417(1 ):104–115. doi:10.1111/nyas.2018.1417.issue-129566262
- Luther C, Swami U, Zhang J, et al. Advanced stage melanoma therapies: detailing the present and exploring the future. Crit Rev Oncol Hematol. 2019;133:99–111. doi:10.1016/j.critrevonc.2018.11.00230661664
- Zhao H, Liao X, Kang Y. Tregs: where we are and what comes next? Front Immunol. 2017;8. doi:10.3389/fimmu.2017.01578
- Han S, Toker A, Liu ZQ, Ohashi PS. Turning the tide against regulatory T cells. Front Oncol. 2019;9:279 eCollection 2019. doi:10.3389/fonc.2019.0027931058083
- Schmidt A, Oberle N, Krammer PH. Molecular mechanisms of treg-mediated T cell suppression. Front Immunol. 2012;3:51 eCollection 2012. doi:10.3389/fimmu.2012.0005122566933
- Lin YC, Chang LY, Huang CT, et al. Effector/memory but not naive regulatory T cells are responsible for the loss of concomitant tumor immunity. J Immunol. 2009;182:6095–6104. doi:10.4049/jimmunol.080382919414761
- Turk MJ, Guevara-Patino JA, Rizzuto GA, et al. Concomitant tumor immunity to a poorly immunogenic melanoma is prevented by regulatory T cells. J Exp Med. 2004;200:771–782. doi:10.1084/jem.2004113015381730
- Bates GJ, Fox SB, Han C, et al. Quantification of regulatory T cells enables the identification of high-risk breast cancer patients and those at risk of late relapse. J Clin Oncol. 2006;24:5373–5380. doi:10.1200/JCO.2006.05.958417135638
- Okita R, Saeki T, Takashima S, et al. CD4+CD25+ regulatory T cells in the peripheral blood of patients with breast cancer and non-small cell lung cancer. Oncol Rep. 2005;14:1269–1273.16211295
- Bergmann C, Strauss L, Wang Y, et al. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: mechanisms of suppression and expansion in advanced disease. Clin Cancer Res. 2008;14:3706–3715. doi:10.1158/1078-0432.CCR-07-512618559587
- Hiraoka N, Onozato K, Kosuge T, et al. Prevalence of FOXP3+ regulatory T cells increases during the progression of pancreatic ductal adenocarcinoma and its premalignant lesions. Clin Cancer Res. 2006;12:5423–5434. doi:10.1158/1078-0432.CCR-06-036917000676
- Curiel TJ, Coukos G, Zou L, et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med. 2004;10:942–949. doi:10.1038/nm109315322536
- Tanaka H, Tanaka J, Kjaergaard J, et al. Depletion of CD4+ CD25+ regulatory cells augments the generation of specific immune T cells in tumor-draining lymph nodes. J Immunother. 2002;25:207–217. doi:10.1097/00002371-200205000-0000312000862
- Prasad SJ, Farrand KJ, Matthews SA, et al. Dendritic cells loaded with stressed tumor cells elicit long-lasting protective tumor immunity in mice depleted of CD4+CD25+ regulatory T cells. J Immunol. 2005;174:90–98. doi:10.4049/jimmunol.174.1.9015611231
- Wing K, Onishi Y, Prieto-Martin P, et al. CTLA-4 control over Foxp3+ regulatory T cell function. Science. 2008;322:271–275. doi:10.1126/science.116006218845758
- Callahan MK, Wolchok JD, Allison JP. Anti-CTLA-4 antibody therapy: immune monitoring during clinical development of a novel immunotherapy. Semin Oncol. 2010;37:473–484. doi:10.1053/j.seminoncol.2010.09.00121074063
- Knutson KL, Disis ML, Salazar LG. CD4 regulatory T cells in human cancer pathogenesis. Cancer Immunol Immunother. 2007;56:271–285. doi:10.1007/s00262-006-0194-y16819631
- McHugh RS, Whitters MJ, Piccirillo CA, et al. CD4(+)CD25(+) immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity. 2002;16:311–323. doi:10.1016/S1074-7613(02)00280-711869690
- Ko K, Yamazaki S, Nakamura K, et al. Treatment of advanced tumors with agonistic anti-GITR mAb and its effects on tumor-infiltrating Foxp3+CD25+CD4+ regulatory T cells. J Exp Med. 2005;202:885–891. doi:10.1084/jem.2005094016186187
- Hirschhorn-Cymerman D, Rizzuto GA, Merghoub T, et al. OX40 engagement and chemotherapy combination provides potent antitumor immunity with concomitant regulatory T cell apoptosis. J Exp Med. 2009;206:1103–1116. doi:10.1084/jem.2008220519414558
- Hegde U, Chhabra A, Chattopadhyay S, et al. Presence of low dose of fludarabine in cultures blocks regulatory T cell expansion and maintains tumor-specific cytotoxic T lymphocyte activity generated with peripheral blood lymphocytes. Pathobiology. 2008;75:200–208. doi:10.1159/00012498118550918
- Correale P, Cusi MG, Tsang KY, et al. Chemo-immunotherapy of metastatic colorectal carcinoma with gemcitabine plus FOLFOX 4 followed by subcutaneous granulocyte macrophage colony-stimulating factor and interleukin-2 induces strong immunologic and antitumor activity in metastatic colon cancer patients. J Clin Oncol. 2005;23:8950–8958. doi:10.1200/JCO.2005.12.14716061910
- Barni S, Lissoni P, Paolorossi F, et al. Mitoxantrone as a single agent in pretreated metastatic breast cancer: effects on T lymphocyte subsets and their relation to clinical response. Tumori. 1991;77:227–231. doi:10.1177/0300891691077003081862550
- Dwarakanath BS, Singh D, Banerji AK, et al. Clinical studies for improving radiotherapy with 2-deoxy-D-glucose: present status and future prospects. J Cancer Res Ther. 2009;5 Suppl 1:S21–S26. doi:10.4103/0973-1482.5513620009289
- Farooque A, Singh N, Adhikari JS, et al. Enhanced antitumor immunity contributes to the radio-sensitization of ehrlich ascites tumor by the glycolytic inhibitor 2-deoxy-D-glucose in mice. PLoS One. 2014;9:e108131. doi:10.1371/journal.pone.010813125248151
- Szylberg L, Karbownik D, Marszalek A. Role of FOXP3 in human cancers. Anticancer Res. 2016;36(8 ):3789–3794.27466478
- Lee W, Lee GR. Transcriptional regulation and development of regulatory T cells. Exp Mol Med. 2018;50:e456. doi:10.1038/emm.2017.31329520112
- Kitagawa Y, Ohkura N, Kidani Y, et al. Guidance of regulatory T cell development by Satb1-dependent super-enhancer establishment. Nat Immunol. 2016;18:173–183. doi:10.1038/ni.364627992401
- Zhao D-M, Xue -H-H. MLL4 keeps Foxp3 in the loop. Nat Immunol. 2017;18:957–958. doi:10.1038/ni.381128829449
- Tone Y, Furuuchi K, Kojima Y, et al. Smad3 and NFAT cooperate to induce Foxp3 expression through its enhancer. Nat Immunol. 2008;9:194–202. doi:10.1038/ni154918157133
- Zheng Y, Josefowicz S, Chaudhry A, et al. Role of conserved non-coding DNA elements in the Foxp3 gene in regulatory T-cell fate. Nature. 2010;463:808–812. doi:10.1038/nature0875020072126
- Kim H-P, Leonard WJ. CREB/ATF-dependent T cell receptor-induced FoxP3 gene expression: a role for DNA methylation. J Exp Med. 2007;204:1543–1551. doi:10.1084/jem.2007010917591856
- Burchill MA, Yang J, Vogtenhuber C, et al. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells. J Immunol. 2007;178:280–290. doi:10.4049/jimmunol.178.1.28017182565
- Kitoh A, Ono M, Naoe Y, et al. Indispensable role of the Runx1-Cbfbeta transcription complex for in vivo-suppressive function of FoxP3+ regulatory T cells. Immunity. 2009;31:609–620. doi:10.1016/j.immuni.2009.09.00319800266
- Rudensky AY. Regulatory T cells and Foxp3. Immunol Rev. 2012;241:260–268. doi:10.1111/j.1600-065X.2011.01018.x
- Galgani M, Rosa VD, Cava AL, Matarese G. Role of metabolism in the immunobiology of regulatory T cells. J Immunol. 2016;197(7 ):2567–2575. doi:10.4049/jimmunol.160024227638939
- Jacobs SR, Herman CE, Maciver NJ, et al. Glucose uptake is limiting in T cell activation and requires CD28- mediated AKT-dependent and independent pathways. J Immunol. 2008;180:4476–4486. doi:10.4049/jimmunol.180.7.447618354169
- Gupta S, Roy A, Dwarakanath BS. Metabolic cooperation and competition in the tumor microenvironment: implications for therapy. Front Oncol. 2017;7:68.28447025
- Liu S, Sun X, Luo J, et al. Effects of radiation on T regulatory cells in normal states and cancer: mechanisms and clinical implications. Am J Cancer Res. 2015;5(11 ):3276–3285.26807310
- Takeuchi Y, Nishikawa H. Roles of regulatory T cells in cancer immunity. Int Immunol. 2016;28:401–409. doi:10.1093/intimm/dxw02527160722
- Ohue Y, Nishikawa H, Regulatory T. (Treg) cells in cancer: can Treg cells be a new therapeutic target? Cancer Sci. 2019;110(7 ):2080–2089. doi:10.1111/cas.1406931102428
- Pandiyan P, Lenardo MJ. The control of CD4+CD25+Foxp3+ regulatory T cell survival. Biol Direct. 2008;3:6. doi:10.1186/1745-6150-3-618304352
- Vincenti F, Kirkman R, Light S, et al. Interleukin-2-receptor blockade with daclizumab to prevent acute rejection in renal transplantation. Daclizumab Triple Therapy Study Group. N Engl J Med. 1998;338:161–165. doi:10.1056/NEJM1998011533803049428817
- Shimizu J, Yamazaki S, Sakaguchi S. Induction of tumor immunity by removing CD25+CD4+ T cells: a common basis between tumor immunity and autoimmunity. J Immunol. 1999;163:5211–5218.10553041
- Onizuka S, Tawara I, Shimizu J, et al. Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor alpha) monoclonal antibody. Cancer Res. 1999;59:3128–3133.10397255
- Jacobs JF, Punt CJ, Lesterhuis WJ, et al. Dendritic cell vaccination in combination with anti-CD25 monoclonal antibody treatment: a phase I/II study in metastatic melanoma patients. Clin Cancer Res. 2010;16:5067–5078. doi:10.1158/1078-0432.CCR-10-175720736326
- Mitchell DA, Cui X, Schmittling RJ, et al. Monoclonal antibody blockade of IL-2 receptor alpha during lymphopenia selectively depletes regulatory T cells in mice and humans. Blood. 2011;118:3003–3012. doi:10.1182/blood-2011-02-33456521768296
- De Vries IJ, Castelli C, Huygens C, et al. Frequency of circulating Tregs with demethylated FOXP3 intron 1 in melanoma patients receiving tumor vaccines and potentially Treg-depleting agents. Clin Cancer Res. 2011;17:841–848. doi:10.1158/1078-0432.CCR-10-222721177412
- Gabler C, Blank N, Hieronymus T, et al. Extranuclear detection of histones and nucleosomes in activated human lymphoblasts as an early event in apoptosis. Ann Rheum Dis. 2004;63:1135–1144. doi:10.1136/ard.2003.01145215308524
- Hansel TT, Kropshofer H, Singer T, et al. The safety and side effects of monoclonal antibodies. Nat Rev Drug Discov. 2010;9:325–338. doi:10.1038/nrd300320305665
- Martinelli V, Dalla Costa G, Sangalli F, et al. Severe disease activity in a patient with multiple sclerosis after daclizumab discontinuation. Mult Scler Relat Disord. 2019;28:57–59. doi:10.1016/j.msard.2018.11.03430554038
- Powell DJ Jr., Attia P, Ghetie V, et al. Partial reduction of human FOXP3+ CD4 T cells in vivo after CD25-directed recombinant immunotoxin administration. J Immunother. 2008;31:189–198. doi:10.1097/CJI.0b013e31815dc0e818481388
- Syn NL, Teng MWL, Mok TSK, et al. De-novo and acquired resistance to immune checkpoint targeting. Lancet Oncol. 2017;18(12 ):e731–e741. doi:10.1016/S1470-2045(17)30607-129208439
- Takahashi T, Tagami T, Yamazaki S, et al. Immunologic self-tolerance maintained by CD25(+)CD4(+) regulatory T cells constitutively expressing cytotoxic T lymphocyte-associated antigen 4. J Exp Med. 2000;192:303–310. doi:10.1084/jem.192.2.30310899917
- Read S, Malmstrom V, Powrie F. Cytotoxic T lymphocyte-associated antigen 4 plays an essential role in the function of CD25(+)CD4(+) regulatory cells that control intestinal inflammation. J Exp Med. 2000;192:295–302.10899916
- Greenfield EA, Nguyen KA, Kuchroo VK. CD28/B7 costimulation: a review. Crit Rev Immunol. 1998;18:389–418. doi:10.1615/CritRevImmunol.v18.i59784967
- Fallarino F, Grohmann U, You S, et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cells. J Immunol. 2006;176:6752–6761. doi:10.4049/jimmunol.176.11.675216709834
- Walker LS. Treg and CTLA-4: two intertwining pathways to immune tolerance. J Autoimmun. 2013;45:49–57. doi:10.1016/j.jaut.2013.06.00623849743
- Kavanagh B, O’Brien S, Lee D, et al. CTLA4 blockade expands FoxP3+ regulatory and activated effector CD4+ T cells in a dose-dependent fashion. Blood. 2008;112:1175–1183. doi:10.1182/blood-2007-11-12543518523152
- Hurwitz AA, Foster BA, Kwon ED, et al. Combination immunotherapy of primary prostate cancer in a transgenic mouse model using CTLA-4 blockade. Cancer Res. 2000;60:2444–2448.10811122
- Kwon ED, Foster BA, Hurwitz AA, et al. Elimination of residual metastatic prostate cancer after surgery and adjunctive cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) blockade immunotherapy. Proc Natl Acad Sci U S A. 1999;96:15074–15079. doi:10.1073/pnas.96.26.1507410611340
- Leach DR, Krummel MF, Allison JP. Enhancement of antitumor immunity by CTLA-4 blockade. Science. 1996;271:1734–1736. doi:10.1126/science.271.5256.17348596936
- Hodi FS. Overcoming immunological tolerance to melanoma: targeting CTLA-4. Asia Pac J Clin Oncol. 2010;6 Suppl 1:S16–S23. doi:10.1111/j.1743-7563.2010.01271.x20482528
- Robert C, Ribas A, Wolchok JD, et al. Anti-programmed-death-receptor-1 treatment with pembrolizumab in ipilimumab-refractory advanced melanoma: a randomised dose-comparison cohort of a phase 1 trial. Lancet. 2014;384(9948 ):1109–1117. doi:10.1016/S0140-6736(14)60958-225034862
- Topalian SL, Drake CG, Pardoll DM. Targeting PD-1/B7-H1 (PD-L1) pathway to activate antitumor immunity. Curr Opin Immunol. 2012;24:207–212. doi:10.1016/j.coi.2011.12.00922236695
- Kumar S, Malik S, Singh UP, et al. PD-1 expression on Foxp3+ Treg cells modulates CD8+ T cell function in prostate tumor microenvironment. J Immunol. 2017;198(1 Supplement ):155.11.
- Francisco LM, Salinas VH, Brown KE, et al. PD-L1 regulates the development, maintenance, and function of induced regulatory T cells. J Exp Med. 2009;206:3015–3029. doi:10.1084/jem.2009084720008522
- Topalian SL, Hodi FS, Brahmer JR, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012;366:2443–2454. doi:10.1056/NEJMoa120069022658127
- Woods DM, Ramakrishnan R, Sodré AL, et al. PD-1 blockade induces phosphorylated STAT3 and results in an increase of Tregs with reduced suppressive function. J Immunol. 2017;198:56.7.
- Kamada T, Togashi Y, Tay C, et al. PD-1+ regulatory T cells amplified by PD-1 blockade promote hyperprogression of cancer. Proc Natl Acad Sci U S A. 2019;116(20 ):9999–10008. doi:10.1073/pnas.182200111631028147
- Shevach EM, Stephens GL. The GITR-GITRL interaction: co-stimulation or contrasuppression of regulatory activity? Nat Rev Immunol. 2006;6:613–618. doi:10.1038/nri186716868552
- Kanamaru F, Youngnak P, Hashiguchi M, et al. Costimulation via glucocorticoid-induced TNF receptor in both conventional and CD25+ regulatory CD4+ T cells. J Immunol. 2004;172:7306–7314. doi:10.4049/jimmunol.172.12.730615187106
- Ephrem A, Epstein AL, Stephens GL, et al. Modulation of Treg cells/T effector function by GITR signaling is context-dependent. Eur J Immunol. 2013;43:2421–2429. doi:10.1002/eji.20134345123722868
- Cohen AD, Diab A, Perales MA, et al. Agonist anti-GITR antibody enhances vaccine-induced CD8(+) T-cell responses and tumor immunity. Cancer Res. 2006;66:4904–4912. doi:10.1158/0008-5472.CAN-05-281316651447
- Zappasodi R, Sirard C, Li Y, et al. Rational design of anti-GITR-based combination immunotherapy. Nat Med. 2019;25:759–766. doi:10.1038/s41591-019-0420-831036879
- Pruitt SK, Boczkowski D, de Rosa N, et al. Enhancement of anti-tumor immunity through local modulation of CTLA-4 and GITR by dendritic cells. Eur J Immunol. 2011;41:3553–3563. doi:10.1002/eji.v41.1222028176
- Nocentini G, Giunchi L, Ronchetti S, et al. A new member of the tumor necrosis factor/nerve growth factor receptor family inhibits T cell receptor-induced apoptosis. Proc Natl Acad Sci U S A. 1997;94:6216–6221. doi:10.1073/pnas.94.12.62169177197
- Kwon B, Yu KY, Ni J, et al. Identification of a novel activation-inducible protein of the tumor necrosis factor receptor superfamily and its ligand. J Biol Chem. 1999;274:6056–6061. doi:10.1074/jbc.274.10.605610037686
- Croft M. Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol. 2003;3:609–620. doi:10.1038/nri114812974476
- Watts TH. TNF/TNFR family members in costimulation of T cell responses. Annu Rev Immunol. 2005;23:23–68. doi:10.1146/annurev.immunol.23.021704.11583915771565
- Piconese S, Valzasina B, Colombo MP. OX40 triggering blocks suppression by regulatory T cells and facilitates tumor rejection. J Exp Med. 2008;205:825–839. doi:10.1084/jem.2007134118362171
- Burocchi A, Pittoni P, Gorzanelli A, et al. Intratumor OX40 stimulation inhibits IRF1 expression and IL-10 production by Treg cells while enhancing CD40L expression by effector memory T cells. Eur J Immunol. 2011;41:3615–3626. doi:10.1002/eji.v41.1222229156
- Pardee AD, McCurry D, Alber S, et al. A therapeutic OX40 agonist dynamically alters dendritic, endothelial, and T cell subsets within the established tumor microenvironment. Cancer Res. 2010;70:9041–9052. doi:10.1158/0008-5472.CAN-10-136921045144
- Shrimali R, Ahmad S, Verma V, et al. concurrent PD-1 blockade negates the effects of OX40 agonist antibody in combination immunotherapy through inducing T-cell apoptosis. Cancer Immunol Res. 2017;5(9 ):755–766. doi:10.1158/2326-6066.CIR-17-029228848055
- Walker LS. Regulatory T cells: folate receptor 4: a new handle on regulation and memory? Immunol Cell Biol. 2007;85:506–507. doi:10.1038/sj.icb.710011517710105
- Yamaguchi T, Hirota K, Nagahama K, et al. Control of immune responses by antigen-specific regulatory T cells expressing the folate receptor. Immunity. 2007;27:145–159. doi:10.1016/j.immuni.2007.04.01717613255
- Stover PJ. Physiology of folate and vitamin B12 in health and disease. Nutr Rev. 2004;62(6 Pt 2 ):S3–S12; discussion S13. doi:10.1111/j.1753-4887.2004.tb00070.x
- Ghiringhelli F, Larmonier N, Schmitt E, et al. CD4+CD25+ regulatory T cells suppress tumor immunity but are sensitive to cyclophosphamide which allows immunotherapy of established tumors to be curative. Eur J Immunol. 2004;34:336–344. doi:10.1002/(ISSN)1521-414114768038
- Proietti E, Greco G, Garrone B, et al. Importance of cyclophosphamide-induced bystander effect on T cells for a successful tumor eradication in response to adoptive immunotherapy in mice. J Clin Invest. 1998;101:429–441. doi:10.1172/JCI13489435316
- Lynch JP, Derhovanessian A, Tazelaar H, et al. Granulomatosis with polyangiitis (Wegener’s Granulomatosis): evolving concepts in treatment. Semin Respir Crit Care Med. 2018;39(4 ):434–458. doi:10.1055/s-0038-166087430404111
- Motoyoshi Y, Kaminoda K, Saitoh O, et al. Different mechanisms for anti-tumor effects of low- and high-dose cyclophosphamide. Oncol Rep. 2006;16(1 ):141–146.16786137
- Berd D, Mastrangelo MJ. Effect of low dose cyclophosphamide on the immune system of cancer patients: depletion of CD4+, 2H4+ suppressor-inducer T-cells. Cancer Res. 1988;48:1671–1675.2830969
- Awwad M, North RJ. Cyclophosphamide-induced immunologically mediated regression of a cyclophosphamide-resistant murine tumor: a consequence of eliminating precursor L3T4+ suppressor T-cells. Cancer Res. 1989;49:1649–1654.2522344
- Awwad M, North RJ. Cyclophosphamide (Cy)-facilitated adoptive immunotherapy of a Cy-resistant tumour. Evidence that Cy permits the expression of adoptive T-cell mediated immunity by removing suppressor T cells rather than by reducing tumour burden. Immunology. 1988;65:87–92.2972604
- Ercolini AM, Ladle BH, Manning EA, et al. Recruitment of latent pools of high-avidity CD8(+) T cells to the antitumor immune response. J Exp Med. 2005;201:1591–1602.15883172
- Zhang H, Chua KS, Guimond M, et al. Lymphopenia and interleukin-2 therapy alter homeostasis of CD4+CD25+ regulatory T cells. Nat Med. 2005;11:1238–1243. doi:10.1038/nm131216227988
- Sharabi A, Haran-Ghera N. Immune recovery after cyclophosphamide treatment in multiple myeloma: implication for maintenance immunotherapy. Bone Marrow Res. 2011;2011:269519. doi:10.1155/2011/26951922046558
- Farooque A, Singh N, Verma A, et al. Immuno-modulation linked to the depletion of T regulatory cells contributes to the radio-sensitization of tumors by the glycolytic inhibitor 2-Deoxy-D-Glucose. J Cancer Res Therap. 2012;8:502.
- Farooque A, Verma A, Adhikari JS, et al. Targeting T-regulatory cells (CD4+CD25highFoxP3+) by the glycolytic inhibitor 2-Deoxy-D-Glucose (2-DG): implications in radiotherapy. J Adv Biotech. 2010;10:113.
- Audia S, Nicolas A, Cathelin D, et al. Increase of CD4+ CD25+ regulatory T cells in the peripheral blood of patients with metastatic carcinoma: a Phase I clinical trial using cyclophosphamide and immunotherapy to eliminate CD4+ CD25+ T lymphocytes. Clin Exp Immunol. 2007;150:523–530. doi:10.1111/(ISSN)1365-224917956583
- Nakahara T, Uchi H, Lesokhin AM, et al. Cyclophosphamide enhances immunity by modulating the balance of dendritic cell subsets in lymphoid organs. Blood. 2010;115:4384–4392. doi:10.1182/blood-2009-11-25123120154220
- Garcia AA, Hirte H, Fleming G, et al. Phase II clinical trial of bevacizumab and low-dose metronomic oral cyclophosphamide in recurrent ovarian cancer: a trial of the California, Chicago, and Princess Margaret Hospital phase II consortia. J Clin Oncol. 2008;26:76–82. doi:10.1200/JCO.2007.12.193918165643
- Greten TF, Ormandy LA, Fikuart A, et al. Low-dose cyclophosphamide treatment impairs regulatory T cells and unmasks AFP-specific CD4+ T-cell responses in patients with advanced HCC. J Immunother. 2010;33:211–218. doi:10.1097/CJI.0b013e3181bb499f20139774
- Nizar S, Copier J, Meyer B, et al. T-regulatory cell modulation: the future of cancer immunotherapy? Br J Cancer. 2009;100:1697–1703. doi:10.1038/sj.bjc.660504019384299
- Obeid M, Tesniere A, Ghiringhelli F, et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat Med. 2007;13:54–61. doi:10.1038/nm152317187072
- Polanczyk MJ, Hopke C, Vandenbark AA, et al. Estrogen-mediated immunomodulation involves reduced activation of effector T cells, potentiation of Treg cells, and enhanced expression of the PD-1 costimulatory pathway. J Neurosci Res. 2006;84:370–378. doi:10.1002/(ISSN)1097-454716676326
- Prieto GA, Rosenstein Y. Oestradiol potentiates the suppressive function of human CD4 CD25 regulatory T cells by promoting their proliferation. Immunology. 2006;118:58–65. doi:10.1111/j.1365-2567.2006.02339.x16630023
- Jiao J, Xiang H, Liao Q. Recent advancement in nonsteroidal aromatase inhibitors for treatment of estrogen-dependent breast cancer. Curr Med Chem. 2010;17:3476–3487. doi:10.2174/09298671079292787720738251
- Generali D, Bates G, Berruti A, et al. Immunomodulation of FOXP3+ regulatory T cells by the aromatase inhibitor letrozole in breast cancer patients. Clin Cancer Res. 2009;15:1046–1051. doi:10.1158/1078-0432.CCR-08-150719188178
- Phillips KA, Ribi K, Fisher R. Do aromatase inhibitors have adverse effects on cognitive function? Breast Cancer Res. 2011;13:203. doi:10.1186/bcr280621392408
- Warburg O. The Metabolism of Tumors. London: Constable and Co.; 1930.
- Adhikari JS, Dwarakanath BS, Mathur R, Ravindranath T. Alterations in radiation induced cell cycle perturbations by 2-deoxy-D-glucose in human tumor cell lines. Indian J Exp Biol. 2003;41:1392–1399.15320491
- Dwarakanath B, Jain V. Targeting glucose metabolism with 2-deoxy-D-glucose for improving cancer therapy. Future Oncol. 2009;5:581–585. doi:10.2217/fon.09.4419519197
- Dwarakanath BS. Cytotoxicity, radiosensitization, and chemosensitization of tumor cells by 2-deoxy-D-glucose in vitro. J Cancer Res Ther. 2009;5 Suppl 1:S27–S31. doi:10.4103/0973-1482.5513720009290
- Dwarakanath BS, Singh S, Jain V. Optimization of tumour radiotherapy: part V Radiosensitization by 2-deoxy-D-glucose and DNA ligand Hoechst-33342 in a murine tumour. Indian J Exp Biol. 1999;37:865–870.10687280
- Gupta S, Farooque A, Adhikari JS, Singh S, Dwarakanath BS. Enhancement of radiation and chemotherapeutic drug responses by 2-deoxy-D-glucose in animal tumors. J Cancer Res Ther. 2009;5 Suppl 1:S16–S20. doi:10.4103/0973-1482.5513520009287
- Mohanti BK, Rath GK, Anantha N, et al. Improving cancer radiotherapy with 2-deoxy-D-glucose: phase I/II clinical trials on human cerebral gliomas. Int J Radiat Oncol Biol Phys. 1996;35:103–111. doi:10.1016/S0360-3016(96)85017-68641905
- Singh D, Banerji AK, Dwarakanath BS, et al. Optimizing cancer radiotherapy with 2-deoxy-d-glucose dose escalation studies in patients with glioblastoma multiforme. Strahlenther Onkol. 2005;181:507–514. doi:10.1007/s00066-005-1320-z16044218
- Venkataramanaa NK, Venkatesh PK, Dwarakanath BS, Vani S. Protective effect on normal brain tissue during a combinational therapy of 2-deoxy-d-glucose and hypofractionated irradiation in malignant gliomas. Asian J Neurosurg. 2013;8:9–14. doi:10.4103/1793-5482.11027423741257
- Gupta S, Mathur R, Dwarakanath BS. The glycolytic inhibitor 2-deoxy-D-glucose enhances the efficacy of etoposide in ehrlich ascites tumor-bearing mice. Cancer Biol Ther. 2005;4:87–94. doi:10.4161/cbt.4.1.138115711125
- De Rosa V, Galgani M, Porcellini A, et al. Glycolysis controls the induction of human regulatory T cells by modulating the expression of FOXP3 exon 2 splicing variants. Nat Immunol. 2015;16:1174–1184. doi:10.1038/ni.326926414764
- Farooque A, Afrin F, Adhikari JS, Dwarakanath BS. Protection of normal cells and tissues during radio- and chemosensitization of tumors by 2-deoxy-D-glucose. J Cancer Res Ther. 2009;5 Suppl 1:S32–S35. doi:10.4103/0973-1482.5513820009291
- Almeida L, Lochner M, Berod L, et al. Metabolic pathways in T cell activation and lineage differentiation. Semin Immunol. 2016;28(5 ):514–524. doi:10.1016/j.smim.2016.10.00927825556
- Persa E, Balogh A, Safrany G, et al. The effect of ionizing radiation on regulatory T cells in health and disease. Cancer Lett. 2015;368(2 ):252–261. doi:10.1016/j.canlet.2015.03.00325754816
- Kachikwu EL, Iwamoto KS, Liao YP, et al. Radiation enhances regulatory T cell representation. Int J Radiat Oncol Biol Phys. 2011;81(4 ):1128–1135. doi:10.1016/j.ijrobp.2010.09.03421093169
- Kershaw MH, Devaud C, John LB, Westwood JA, Darcy PK. Enhancing immunotherapy using chemotherapy and radiation to modify the tumor microenvironment. Oncoimmunology. 2013;2:e25962. doi:10.4161/onci.2596224327938
- Patel MA, Kim JE, Theodros D, et al. Agonist anti-GITR monoclonal antibody and stereotactic radiation induce immunemediated survival advantage in murine intracranial glioma. J Immuno Ther Cancer. 2016;4:28. doi:10.1186/s40425-016-0132-2
- Cao M, Cabrera R, Xu Y, et al. Gamma irradiation alters the phenotype and function of CD4+CD25+ regulatory T cells. Cell Biol Int. 2009;33:565–571. doi:10.1016/j.cellbi.2009.02.00719268553
- Muroyama Y, Nirschl TR, Kochel CM, et al. Stereotactic radiotherapy increases functionally suppressive regulatory T cells in the tumor microenvironment. Cancer Immunol Res. 2017;5(11 ):992–1004. doi:10.1158/2326-6066.CIR-17-004028970196
- Oweida AJ, Darragh L, Phan A, et al. STAT3 modulation of regulatory T cells in response to radiation therapy in head and neck cancer. J Natl Cancer Inst. 2019; djz036 [Epub ahead of print]. doi:10.1093/jnci/djz036.
- Mignot G, Roux S, Thery C, Segura E, Zitvogel L. Prospects for exosomes in immunotherapy of cancer. J Cell Mol Med. 2006;10:376–388. doi:10.1111/jcmm.2006.10.issue-216796806
- Ohno H. Overview: membrane traffic in multicellular systems: more than just a housekeeper. J Biochem. 2006;139:941–942. doi:10.1093/jb/mvj11916788043
- Wada J, Onishi H, Suzuki H, et al. Surface-bound TGF-beta1 on effusion-derived exosomes participates in maintenance of number and suppressive function of regulatory T-cells in malignant effusions. Anticancer Res. 2010;30:3747–3757.20944164
- Onishi H, Morisaki T, Katano M. Immunotherapy approaches targeting regulatory T-cells. Anticancer Res. 2012;32:997–1003.22399623
- Willcox A, Richardson SJ, Bone AJ, Foulis AK, Morgan NG. Analysis of islet inflammation in human type 1 diabetes. Clin Exp Immunol. 2009;155:173–181. doi:10.1111/cei.2009.155.issue-219128359
- Tree TI, Roep BO, Peakman M. A mini meta-analysis of studies on CD4+CD25+ T cells in human type 1 diabetes: report of the immunology of diabetes society T cell workshop. Ann N Y Acad Sci. 2006;1079:9–18. doi:10.1196/annals.1375.00217130526
- Izcue A, Powrie F. Special regulatory T-cell review: regulatory T cells and the intestinal tract–patrolling the frontier. Immunology. 2008;123:6–10. doi:10.1111/imm.2008.123.issue-118154611
- Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003;299:1057–1061. doi:10.1126/science.107949012522256
- Kohm AP, Carpentier PA, Anger HA, Miller SD. Cutting edge: CD4+CD25+ regulatory T cells suppress antigen-specific autoreactive immune responses and central nervous system inflammation during active experimental autoimmune encephalomyelitis. J Immunol. 2002;169:4712–4716. doi:10.4049/jimmunol.169.9.471212391178
- Morgan ME, Sutmuller RP, Witteveen HJ, et al. CD25+ cell depletion hastens the onset of severe disease in collagen-induced arthritis. Arthritis Rheum. 2003;48:1452–1460. doi:10.1002/art.1106312746920
- Buckner JH. Mechanisms of impaired regulation by CD4(+)CD25(+)FOXP3(+) regulatory T cells in human autoimmune diseases. Nat Rev Immunol. 2010;10:849–859. doi:10.1038/nri288921107346
- Yan KX, Fang X, Han L, et al. Foxp3+ regulatory T cells and related cytokines differentially expressed in plaque vs. guttate psoriasis vulgaris. Br J Dermatol. 2010;163:48–56. doi:10.1111/j.1365-2133.2010.09742.x20222932
- Bovenschen HJ, van Vlijmen-willems IM, van de Kerkhof PC, van Erp PE. Identification of lesional CD4+ CD25+ Foxp3+ regulatory T cells in Psoriasis. Dermatology. 2006;213:111–117. doi:10.1159/00009384916902287
- Wood KJ, Sakaguchi S. Regulatory T cells in transplantation tolerance. Nat Rev Immunol. 2003;3:199–210. doi:10.1038/nri102712658268
- Cohen JL, Trenado A, Vasey D, Klatzmann D, Salomon BL. CD4(+)CD25(+) immunoregulatory T cells: new therapeutics for graft-versus-host disease. J Exp Med. 2002;196:401–406. doi:10.1084/jem.2002009012163568
- Edinger M, Hoffmann P, Ermann J, et al. CD4+CD25+ regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation. Nat Med. 2003;9:1144–1150. doi:10.1038/nm91512925844
- Taylor PA, Lees CJ, Blazar BR. The infusion of ex vivo activated and expanded CD4(+)CD25(+) immune regulatory cells inhibits graft-versus-host disease lethality. Blood. 2002;99:3493–3499. doi:10.1182/blood.V99.10.349311986199
- Gallimore A, Sakaguchi S. Regulation of tumour immunity by CD25+ T cells. Immunology. 2002;107:5–9. doi:10.1046/j.1365-2567.2002.01471.x12225357
- Zenclussen AC. CD4(+)CD25+ T regulatory cells in murine pregnancy. J Reprod Immunol. 2005;65:101–110. doi:10.1016/j.jri.2005.01.00315811515
- Maj T, Wang W, Crespo J, et al. Oxidative stress controls regulatory T cell apoptosis and suppressor activity and PD-L1-blockade resistance in tumor. Nat Immunol. 2017;18:1332–1341. doi:10.1038/ni.386829083399
- Chung KY, Gore I, Fong L, et al. Phase II study of the anti-cytotoxic T-lymphocyte-associated antigen 4 monoclonal antibody, tremelimumab, in patients with refractory metastatic colorectal cancer. J Clin Oncol. 2010;28:3485–3490. doi:10.1200/JCO.2010.28.399420498386
- Ralph C, Elkord E, Burt DJ, et al. Modulation of lymphocyte regulation for cancer therapy: a phase II trial of tremelimumab in advanced gastric and esophageal adenocarcinoma. Clin Cancer Res. 2010;16:1662–1672. doi:10.1158/1078-0432.CCR-09-287020179239
- Kirkwood JM, Lorigan P, Hersey P, et al. Phase II trial of tremelimumab (CP-675,206) in patients with advanced refractory or relapsed melanoma. Clin Cancer Res. 2010;16:1042–1048. doi:10.1158/1078-0432.CCR-09-203320086001
- Camacho LH, Antonia S, Sosman J, et al. Phase I/II trial of tremelimumab in patients with metastatic melanoma. J Clin Oncol. 2009;27:1075–1081. doi:10.1200/JCO.2008.19.243519139427
- Rech AJ, Vonderheide RH. Clinical use of anti-CD25 antibody daclizumab to enhance immune responses to tumor antigen vaccination by targeting regulatory T cells. Ann N Y Acad Sci. 2009;1174:99–106. doi:10.1111/j.1749-6632.2009.04939.x19769742
- Mahnke K, Schonfeld K, Fondel S, et al. Depletion of CD4+CD25+ human regulatory T cells in vivo: kinetics of Treg depletion and alterations in immune functions in vivo and in vitro. Int J Cancer. 2007;120:2723–2733. doi:10.1002/ijc.2261717315189
- Powell DJ Jr., Felipe-Silva A, Merino MJ, et al. Administration of a CD25-directed immunotoxin, LMB-2, to patients with metastatic melanoma induces a selective partial reduction in regulatory T cells in vivo. J Immunol. 2007;179:4919–4928. doi:10.4049/jimmunol.179.7.491917878392
- Curtin JF, Candolfi M, Fakhouri TM, et al. Treg depletion inhibits efficacy of cancer immunotherapy: implications for clinical trials. PLoS One. 2008;3:e1983. doi:10.1371/journal.pone.000198318431473
- Garon EB, Rizvi NA, Hui R, et al. Pembrolizumab for the treatment of non-small-cell lung cancer. N Engl J Med. 2015;372(21 ):2018–2028. doi:10.1056/NEJMoa150182425891174
- Oberst MD, Augé C, Morris C, et al. Potent immune modulation by MEDI6383, an engineered human OX40 ligand IgG4P Fc fusion protein. Mol Cancer Ther. 2018;17(5 ):1024–1038. doi:10.1158/1535-7163.MCT-17-020029545330
- Preston CC, Goode EL, Hartmann LC, Kalli KR, Knutson KL. Immunity and immune suppression in human ovarian cancer. Immunotherapy. 2011;3:539–556. doi:10.2217/imt.11.2021463194
- Maker AV, Phan GQ, Attia P, et al. Tumor regression and autoimmunity in patients treated with cytotoxic T lymphocyte-associated antigen 4 blockade and interleukin 2: a phase I/II study. Ann Surg Oncol. 2005;12:1005–1016. doi:10.1245/ASO.2005.03.53616283570