
Abstract
MicroRNAs (miRNAs) are endogenous, noncoding, single-stranded RNA molecules of 22 nucleotides in length. MiRNAs have both tumor-suppressive properties and oncogenic properties that can control critical processes in tumors. Mature miR-125b originates from miR-125b-1 and miR-125b-2 and leads to the degradation of target mRNAs or the inhibition of translation through binding to the 3′ untranslated regions (3′-UTR) of target mRNAs. Importantly, miR-125b is involved in regulating NF-κB, p53, PI3K/Akt/mTOR, ErbB2, Wnt, and another signaling pathways, thereby controlling cell proliferation, differentiation, metabolism, apoptosis, drug resistance and tumor immunity. This review aims to summarize the recent literature on the role of miR-125b in the regulation of tumorigenesis and to explore its potential clinical application in the diagnosis, prognosis and clinical treatment of tumors.
Introduction
MicroRNAs (miRNAs) are endogenous, noncoding, single-stranded RNA molecules of 22 nucleotides in length that regulate approximately 30% of human genes.Citation1 MicroRNAs are involved in the regulation of multiple cellular functions, including apoptosis,Citation2 metabolism,Citation3 proliferation and differentiation. In addition, miRNAs play a critical role in the communication of tumor microenvironmental cells, influence the tumor microenvironment, and are involved in tumor-related inflammation, hypoxia and immunity.Citation4–Citation7 Meanwhile, miRNAs exist in various biological fluids as circulating miRNAs, and changes in circulating miRNAs are indicative of pathophysiological conditions in cancers. Thus, circulating miRNAs can be effective biomarkers in cancer diagnosis.Citation8 Interestingly, miRNAs have both tumor-suppressive properties and oncogenic properties that can control critical components of signaling pathways. Dysregulation of miRNAs can lead to the generation of surrogate and compensatory signals (parallel or downstream pathways to drug-blocked pathways), thereby maintaining drug resistance.Citation9
Mature miR-125b originates from miR-125b-1 and miR-125b-2. MiR-125b-1 is derived from a long noncoding RNA (lncRNA)–MIR100HG (miR-100/let-7a-2/miR-125b-1, chromosome 11), and miR-125b-2 is derived from a miRNA cluster (miR-99a/let-7c/miR-125b-2, chromosome 21). Recently, miR-125b has emerged as an important regulator in human cancers, and it is incorporated into the RNA-induced silencing complex (RISC), which leads to the degradation of target mRNAs or the inhibition of translation through binding to the 3′ untranslated regions (3′-UTRs) of target mRNAs.Citation10 In addition to targeting mRNAs encoding proteins, miR-125b can also target lncRNAs such as MALAT1 and inhibit its expression. Moreover, miR-125b can produce a synergistic effect when combined with miRNAs from the same miRNA cluster. It has been reported that miR-125b and miR-100 coregulate the resistance of cetuximab or vincristine.Citation11,Citation12
Dysregulation of miR-125b in Cancers
The dysregulation of miR-125b is shown in . Upregulation of miR-125b as an oncogene has been reported in various cancers: nasopharyngeal carcinoma (NPC),Citation13,Citation14 retinoblastoma (RB),Citation15 glioblastoma (GBM),Citation16–Citation20 poorly differentiated non-small-cell lung cancer (NSCLC),Citation21 acute lymphoblastic leukemia (ALL),Citation22 acute myeloid leukemia (AML),Citation23 and gastric cancer.Citation24–Citation26 On the other hand, miR-125b, as a tumor suppressor, is downregulated in the following cancers: non-small-cell lung cancer (NSCLC),Citation27 esophageal squamous cell carcinoma (ESCC),Citation28,Citation29 anaplastic thyroid cancer,Citation30 bladder cancer,Citation31–Citation35 hepatocellular carcinoma (HCC),Citation36–Citation39 melanoma,Citation40,Citation41 ovarian cancer,Citation42–Citation44 osteosarcoma,Citation45–Citation47 chondrosarcoma,Citation48 breast cancer,Citation49–Citation55 gallbladder cancer (GBC),Citation56 endometrioid endometrial cancer (EEC),Citation57 colorectal cancer (CRC),Citation58,Citation59 multiple myeloma (MM),Citation60 and Ewing’s sarcoma (ES).Citation61
Table 1 The Target Genes of miR-125b in Different Cancers
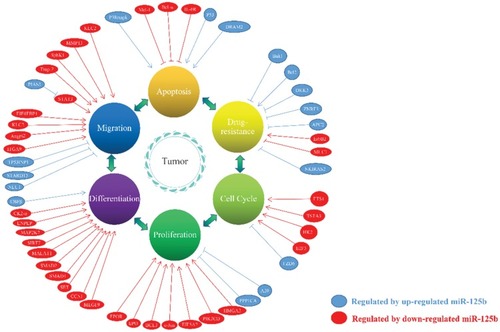
The function of miR-125b diverges in different cancers depending on the different molecular contexts and the tumor microenvironment. At present, many genes have been confirmed as target genes of miR-125b, covering a variety of biological signaling pathways and affecting the formation of many malignant phenotypes such as proliferation, differentiation, migration, apoptosis, cell cycle and drug resistance in different cancers []. In tumors that upregulate miR-125b, tumorigenesis is promoted by inhibiting proapoptotic genes and genes that inhibit proliferation and invasion. Conversely, in tumors where miRNA-125b is downregulated, tumorigenesis is promoted by reducing the inhibition of genes that promote proliferation, differentiation, and apoptosis inhibition.
Figure 1 The role of miR-125b in the development of cancer.
Upregulation of miR-125b in Cancer
In previous studies of many cancers, miR-125b participated in tumor proliferation and cell cycle regulation as a suppressor mechanism. In NPC and ALL, miR-125b was shown to directly suppress A20, which inhibits proliferation and induces apoptosis by negatively regulating the NF-κB signaling pathway.Citation13,Citation22,Citation62 The study showed that downregulation of CBFβ by miR-125b prevents granulocyte differentiation and maturation, indicating that miR-125b promotes hematopoietic malignancies through the CDX2/miR-125b/CBFβ pathway.Citation23 Overexpression of miR-125b in gastric cancer results in downregulation of PPP1CA and then promotes phosphorylation of Rb protein and activation of E2F, a transcription factor, to promote gene transcription. This indicates that miR-125b participates in the pathogenesis of gastric cancer through the PPP1CA-Rb-E2F signaling pathway.Citation26 Upregulation of miR-125b decreased the level of FZD6 in GBM. Unexpectedly, in this mechanism, FZD6 acted as a negative regulator to inhibit the Wnt pathway while promoting STAT3 and NF-kB signaling, and miR-125b was shown to be an important regulator of GBM tumorigenesis. NEU1 and STARD13 are direct targets of miR-125b. NEU1 acts as a tumor suppressor gene to regulate downstream molecules by encoding NEU1 sialidase. One of the functions of NEU1 is sialylation, which can reduce protein phosphorylation and attenuate FAK, ERK1/2, and MMP17.Citation55 Our data suggest that autophagy is reduced in triple-negative breast cancer (TNBC), likely leading to the deregulation of the EGFR-MUC1-NEU1 complex and its associated cellular pathways. Another function of NEU1 is to form the EGFR-MUC1-NEU1 complex that controls autophagy and associated cellular pathways.Citation63 Zheng et al indicated that the mRNA of StarD13 as a ceRNA regulated the expression of TP53INP1, thereby regulating the migration and invasion of breast cancer cells.Citation64
MiR-125b can inhibit apoptosis to promote tumorigenesis. In RB and GMB, miR is significantly upregulated, and then it apparently suppresses DRAM2, p53, and P38.Citation15,Citation16,Citation65 DRAM2 is a crucial component of the p53 signaling pathway, and the induction of autophagy by DRAM is a potential mechanism contributing to cell death. DRAM2 encodes a transmembrane lysosomal protein that is thought to play a key role in autophagy initiation, and the autophagy induced by DRAM2 is an underlying mechanism of p53-mediated cell death.Citation15,Citation65 Both p53 and p38MAPK are direct target genes of miR-125b, and p53 is known to activate the apoptotic pathway, while p38MAPK can induce mitochondrial apoptosis.Citation16,Citation66 Zheng et al reported that miR-125b expression was significantly higher in poorly differentiated NSCLC cells with high metastatic potential and that it reduced the level of TP53INP1. TP53INP1 is considered a tumor suppressor gene with proapoptotic and antimetastatic functions.Citation64
Downregulation of miR-125b in Cancer
In terms of the downregulation of miR-125b in various tumors, the low expression of miR-125b promotes cell proliferation, migration and invasion by controlling HMGA2 and STAT3 in ESCC.Citation28,Citation29 HMGA2 alters the structure of DNA and regulates gene transcription. In both Ewing’s sarcoma and ATC, miR-125b regulates the PI3K/Akt/mTOR pathway by regulating PIK3CD. MiR-125b in bladder cancer upregulates Trop-2, SphK1, E2F3, SIRT7, MALAT1, and MMP13 to promote cancer tumorigenesis and metastasis.Citation67,Citation68
In HCC, miR-125b regulates many genes involved in various mechanisms. The downregulation of miR-125b causes the overexpression of EIF5A2, which is known to be associated with proliferation and poor prognosis in patients with CRC and NSCLC.Citation39,Citation69–Citation71 MiR-100 reduces the protein level of Angpt2, which is an essential molecule for angiogenesis, by blocking the mTOR/p70S6K signaling pathway. It has been demonstrated that miR-125b can cause excessive angiogenesis by affecting the proliferation of endothelial cells.Citation36 MiR-125b downregulation facilitates the epithelial-mesenchymal transition (EMT) of HCC cells by targeting SMAD2 and SMAD4, which are both signal transducers of the TGFβ signaling pathway.Citation37,Citation72 Moreover, miR-125b is downregulated in macrophage exosomes of HCC and can target CD90 to mediate the stem cells of HCC.Citation73 EMT can increase tumor stem cell-like properties, such as migratory and invasive capabilities and chemoresistance. Furthermore, the levels of Mcl-1, Bcl-xL and Bcl-w, the antiapoptotic members of the Bcl-2 family, are increased via miR-125b. The Bcl-2 family plays a crucial role in suppressing apoptosis largely through binding to proapoptotic proteins (Bax and Bak1).Citation38,Citation74
ITGA9, NEDD9, and c-jun are controlled by miR-125b and contributed to melanoma development. One study showed that ITGA9 promotes EMT by promoting the dissociation of E-cadherin and beta-catenin.Citation75 The NEDD9/Src/FAK signaling complex plays an important role in mediating the movement of growth factors and converting mechanical forces into biochemical signals. These signals promote cancer metastasis by regulating migration, invasion, and infiltration.Citation76 Interestingly, miR-125b can bind within the coding sequence (CDS) of c-Jun mRNA and thus regulate c-Jun protein expression.Citation41 In ovarian cancer, SET, EIF4EBP1, and BCL3 are the targets of miR-125b.Citation43 Zhao et al indicated that BCL3 serves as an oncogene and is regulated by its downstream target gene, STAT3.Citation77 SET proteins, also known as TAF-1β, I2PP2A and INHAT, belong to the multifunctional protein family which is involved in apoptosis, transcription, nucleosome assembly and histone binding.Citation42
The decrease in miR-125b expression in breast cancer may be due to the enhancement of DNA methylation in the promoter region.Citation52 Downregulation of miR-125b induces cell survival by inducing the expression of erythropoietin (EPO) and its receptor EPOR through the ERBB2/Her2 pathway.Citation78 In addition, miR-125b downregulates the expression of the oncogene ETS1 involved in cell cycle transformation and proliferation. Further, ENPEP is thought to be an important and highly specific enzyme that metabolizes and inactivates bioactive peptides such as angiotensin II (AngII) in the renin-angiotensin system. ENPEP is expressed in cervical neoplasm lesions and is upregulated as lesions develop from invasive intraepithelial neoplasias to invasive squamous cell carcinomas.Citation79 In addition, MAP2K7 and TSTA3 are upregulated by miR-125b. MiR-125b-mediated EMT may occur through the MAP2K7–JNK cascade in breast cancer. TSTA3 is an NADP(H)-binding protein that is involved in electron carrier activity, GDP-l-fucose synthase activity, glycoprotein biosynthesis, cell-cell adhesion, and biopolymer glycosylation.Citation49 Wang et al reanalyzed GEO data and speculated that the TSTA3-related network regulates cell apoptosis, cyclin-dependent protein kinase activity, cell migration, insulin secretion, transcription, and cell proliferation in HCC.Citation80 MiR-125b is also downregulated in EECs, and Shang et al demonstrated that miR-125b inhibits EEC cell invasion by downregulating ErbB2. Several studies have indicated ErbB2 as the target of miR-125b that mediates tumor development and invasion.Citation57,Citation81 It is worth considering that patients with overexpression of ERBB2 have an important genotype in breast cancer. It has been reported that miR-125b is involved in drug resistance in these patients.Citation48 However, the significance of miR-125b in the diagnosis and treatment of breast cancer patients deserves further study.
MiR-125b Controls Drug Resistance in Cancer
As in the mechanism of tumorigenesis, miR-125b also has two sides in the mechanism of drug resistance. Multidrug resistance (MDR) is a major clinical obstacle in the successful treatment of patients with NPC. In a study by Yuan et al a significant reduction in miR-125b expression levels was observed in cisplatin (DDP)-resistant cells compared with parental NPC cells. The results of the study suggest that miR-125b may regulate the sensitivity of NPC cells to DDP by modulating the expression levels of the antiapoptotic factor BCL-2. Additionally, Tang et al found that miR-125b acts as a tumor suppressor to inhibit glucose metabolism by targeting ErbB2 in chondrosarcoma, thereby inhibiting glucose metabolism. The results showed that miR-125b is downregulated in chondrosarcoma patient tissues and doxorubicin-resistant cells compared to normal tissues. MiR-125 overexpression enhances doxorubicin sensitivity in parental and doxorubicin-resistant cells by directly targeting ErbB2-mediated upregulation of glycolysis.Citation48
In contrast, miR-125b was found to be overexpressed in glioblastoma stem cells (GSCs) compared to normal brain tissues and GBM tissues. The repression of miR-125b before temozolomide (TMZ) treatment in GSCs resulted in increased inhibition of cell proliferation compared to that with TMZ treatment alone, and a study indicated that miR-125b enhances TMZ resistance by targeting Bak1.Citation20 Similarly, Haemmig et al showed that GBM cells overexpressing miR-125b exhibit increased NF-κB activity and antiapoptotic activity and upregulation of cell cycle genes by targeting TNFAIP3 and NKIRAS2, both of which contribute to NF-κB activity. This is significantly related to the resistance of GBM cells to TNFα- and TMZ-induced apoptosis.Citation17 Furthermore, Shi et al showed that a miR-125b inhibitor enhanced the anti-invasion activity of TMZ in GSCs by targeting PIAS3, which contributed to reduced STAT3 transcriptional activity and subsequently decreased the expression of MMP-2 and MMP-9.Citation18 Remarkably, miR-125b overexpression was clearly associated with shorter overall survival of patients treated with TMZ. The evidence suggests that miR-125b may be a predictor of TMZ response in GBM patients.
Recently, studies have shown that miR-125b is associated with resistance to ERBB-targeted therapies by activating downstream signal transducers or parallel pathways.Citation9,Citation24,Citation82 In research on cetuximab resistance in CRC, miR-100 and miR-125b derived from lncRNA MIR100HG were found to be overexpressed in cetuximab-resistant cells. MiR-100 and miR-125b coordinately repressed five Wnt/β-catenin negative regulators, resulting in increased Wnt signaling and cetuximab resistance. In this mechanism, miR-125b inhibited DKK3, ZNRF3, RNF43, and APC2. LncRNA TINCR has been reported to sponge miR-125b and release HER-2, resulting in trastuzumab resistance in breast cancer.Citation83
The Role of miR-125b in Tumor Immunity
MiR-125b also plays an important role in tumor immunity. Studies have shown that miR-125b is involved in the regulation of immune cells such as T cells, B cells, DC cells and macrophages.Citation84–Citation87 MiR-125b regulates T cell proliferation and activation, is highly expressed in naive CD4 T cells and can inhibit the T cell immune response, while miR-125b can promote T cell apoptosis.Citation84,Citation88 Overexpression of miR-125b in the B cells of tumor patients prevents the release of immature B cells from the bone marrow into the blood. Overexpression of miR-125b also leads to defects in the development of pre-B cells, and miR-125b inhibits differentiation of primary B cells into plasma cells.Citation89
Macrophages can be polarized into two different types of macrophages (M1 and M2). The M1 type promotes the inflammatory response, while the M2 type inhibits the inflammatory response. MiR-125b regulates the polarization of macrophages and is highly expressed in M1 macrophages and promotes tumor immune responses by targeting IRF4.Citation85 In an ovarian cancer study, nanoparticles coated with miR-125b repolarized tumor-associated macrophages (TAMs) in the peritoneal cavity to M1 macrophages. Nanoparticles combined with paclitaxel enhanced anti-ovarian cancer efficacy compared to that with pacitaxel alone.Citation90 Nanoparticles encapsulating miR-125b also successfully reprogrammed TAMs to the M1 phenotype in non-small-cell lung cancer (NSCLC), which may play an important role in immunotherapy.Citation91
MiR-125b in Cancer Diagnosis and Prognosis
MiRNAs in solid tissues or circulating miRNAs can be used as a basis for the diagnosis of tumors and prognosis. Currently available tumor markers for HCC are of little clinical relevance. Zuo et al suggested that a panel of serum miRNAs (including miR-125b) combined with alpha-fetoprotein (AFP) had a higher sensitivity (82%) and specificity (75%) for the diagnosis of early-stage HCC compared to that of a single marker.Citation92 In 150 patients, 90 patients with HCC and 60 patients without cancer, serum expression of four miRNAs was assessed. The results indicated that the combination of miR-125b, miR-223, miR-27a, and miR-26a could serve as a valuable biomarker with an AUC of 0.874 in distinguishing the HCC group from the noncancer group in AFP-negative subjects. Moreover, the diagnostic accuracy of miR-125b in cancer diagnosis was assessed by a meta-analysis, which included 695 patients with various kinds of cancers and 370 healthy controls from 8 qualified studies. In this meta-analysis, the sensitivity and specificity of miR-125b in cancer diagnosis were 82% (95% CI, 76–87%) and 77% (95% CI, 70–84%), respectively. A high AUC of 0.84 reflected the high overall level of diagnostic accuracy.Citation93
Studies have suggested that the high expression of miR-125b could be an independent and poor prognostic factor in patients with HER2-positive gastric cancer. Wu et al detected the expression of miR-125b in fresh tissues of 50 gastric cancer patients and 6 gastric cancer cell lines. MiR-125b was detected by in situ hybridization, and its clinicopathological diagnosis and clinical parameters were studied. The results demonstrated that the 5-year survival rate of patients with gastric cancer and with high levels of miR-125b expression was significantly lower than that of patients with low levels of expression in stages I, II, and III. In addition, this high expression level of miR-125b in gastric cancer tissues was associated with lymph node and distant metastases.Citation26 Sui et al collected a total of 132 samples of gastric cancer and 38 noncancerous samples, and the analyzed results showed that the OS rate of patients with HER2-positive gastric cancer and higher miR-125b expression was significantly reduced compared to those with lower miR-125b expression (P=0.034).
Several studies have indicated miR-125b as a biomarker to predict prognosis in breast cancer.Citation49,Citation94–Citation96 Sun et al performed a study to analyze miR-125b in breast cancer, and they followed up with the patients who provided 175 pairs of fresh breast cancer and normal control samples for 80 months. The results showed that TSTA3 was highly regulated by miR-125b, highly expressed in breast cancer tissues and tumor cells, and closely related to TNM stage. TSTA3-overexpressing patients had low survival rates.Citation49 In 2019, a study showed that miR-125b is upregulated in the blood of untreated breast cancer patients. We were able to differentiate between breast cancer patients and normal women by the amount of miR-125b in plasma (AUC = 0.85).Citation97 In addition, the combination of miR-125b and CA153 provided better diagnostic accuracy (89% vs 70%) than CA153 alone. In the prognosis of breast cancer, analysis in the combined TCGA and METABRIC databases showed that patients with high miR-125b had a better prognosis (HR = 0.72, P = 0.0015).Citation97
Moreover, in small intestinal neuroendocrine tumors (SBNETs), four serum miRNAs (miR-125b, −362, −425 and −500a) had high diagnostic value (AUC=0.951) in SBNETs. The effectiveness after surgical resection of SBNET was evaluated, and the results showed that miR-125b-5p returns to normal levels in patients without disease and remains upregulated in patients with residual tumors one month after surgery.Citation98
Signaling Pathways Regulated by miR-125b
As mentioned above, miR-125b may play different roles in different cancers, and miR-125b’s anticancer or procancer function may also be regulated by different signaling pathways. In terms of cancer promotion, miR-125b can target DRAM2 and p53 and then inhibit the P53 tumor suppressor signaling pathway.Citation16,Citation66 In addition, miR-125b can target A20 to activate the NF-κB signaling pathway and target FZD6 to activate STAT3 and the Wnt signaling pathway.Citation13,Citation19,Citation22,Citation62 As tumor suppressors, miR-125b can promote the PI3K/Akt, TGFβ, and ERBB2/Her2 signaling.Citation67,Citation68,Citation72,Citation78
Conclusions and Future Directions
In conclusion, emerging evidence further demonstrates that miR-125b dysregulation is a general feature in many cancers and plays an important role in tumorigenesis and clinical therapy. MiR-125b may serve as a potent tumor promoter or inhibitor in different tumors, depending on the different molecular contexts. MiR-125b is involved in various aspects of tumor cell proliferation, differentiation, invasion, migration, drug resistance, and tumor immunity. Importantly, miR-125b can serve as a diagnostic and prognostic marker for multiple tumors. However, the roles of miR-125b have not been fully elucidated in different tumor subtypes, and defining its potential molecular mechanisms in the pathological processes of tumors will be helpful to provide an opportunity for possible intervention in cancer therapy by targeting either the regulatory pathways or the miRNAs themselves. The current problem encountered with miR-125b is that it has dual functions in tumor regulation. We need to adopt different analysis and interference strategies for miR-125b in different tumors. However, at present, miR-125b as a therapeutic target needs to overcome many obstacles, such as the stability of the drug, the efficiency of the delivery system, and the off-target effect of the drug.
In the future, miR-125b can be studied more to determine the interaction between miR-125b and other functions in immunotherapy or as a biomarker for diagnosis and therapy and to determine the more precise regulation of miR-125b in different cancers. Therefore, it is proposed that miR-125b could be regarded as a new therapeutic target in the treatment of cancer.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-514744438
- Garofalo M, Condorelli GL, Croce CM, Condorelli G. MicroRNAs as regulators of death receptors signaling. Cell Death Differ. 2010;17(2):200–208. doi:10.1038/cdd.2009.10519644509
- Chan B, Manley J, Lee J, Singh SR. The emerging roles of microRNAs in cancer metabolism. Cancer Lett. 2015;356(2Pt A):301–308. doi:10.1016/j.canlet.2014.10.01125451319
- Rupaimoole R, Calin GA, Lopez-Berestein G, Sood AK. miRNA deregulation in cancer cells and the tumor microenvironment. Cancer Discov. 2016;6(3):235–246. doi:10.1158/2159-8290.CD-15-089326865249
- Chiba T, Marusawa H, Ushijima T. Inflammation-associated cancer development in digestive organs: mechanisms and roles for genetic and epigenetic modulation. Gastroenterology. 2012;143(3):550–563. doi:10.1053/j.gastro.2012.07.00922796521
- Mehta A, Baltimore D. MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol. 2016;16(5):279–294. doi:10.1038/nri.2016.4027121651
- Contreras J, Rao DS. MicroRNAs in inflammation and immune responses. Leukemia. 2012;26(3):404–413. doi:10.1038/leu.2011.35622182919
- D’Angelo B, Benedetti E, Cimini A, Giordano A. MicroRNAs: a puzzling tool in cancer diagnostics and therapy. Anticancer Res. 2016;36(11):5571–5575. doi:10.21873/anticanres27793880
- Migliore C, Giordano S. Resistance to targeted therapies: a role for microRNAs? Trends Mol Med. 2013;19(10):633–642. doi:10.1016/j.molmed.2013.08.00224012193
- Maqbool R, Ul Hussain M. MicroRNAs and human diseases: diagnostic and therapeutic potential. Cell Tissue Res. 2014;358(1):1–15. doi:10.1007/s00441-013-1787-324493638
- Lu Y, Zhao X, Liu Q, et al. lncRNA MIR100HG-derived miR-100 and miR-125b mediate cetuximab resistance via Wnt/beta-catenin signaling. Nat Med. 2017;23(11):1331–1341. doi:10.1038/nm.442429035371
- Akbari Moqadam F, Lange-Turenhout EA, Aries IM, Pieters R, den Boer ML. MiR-125b, miR-100 and miR-99a co-regulate vincristine resistance in childhood acute lymphoblastic leukemia. Leuk Res. 2013;37(10):1315–1321. doi:10.1016/j.leukres.2013.06.02723915977
- Zheng Z, Qu JQ, Yi HM, et al. MiR-125b regulates proliferation and apoptosis of nasopharyngeal carcinoma by targeting A20/NF-kappaB signaling pathway. Cell Death Dis. 2017;8(6):e2855. doi:10.1038/cddis.2017.21128569771
- Yuan TZ, Zhang HH, Lin XL, et al. microRNA-125b reverses the multidrug resistance of nasopharyngeal carcinoma cells via targeting of Bcl-2. Mol Med Rep. 2017;15(4):2223–2228. doi:10.3892/mmr.2017.623328260044
- Bai S, Tian B, Li A, Yao Q, Zhang G, Li F. MicroRNA-125b promotes tumor growth and suppresses apoptosis by targeting DRAM2 in retinoblastoma. Eye (Lond). 2016;30(12):1630–1638. doi:10.1038/eye.2016.18927518550
- Wu N, Lin X, Zhao X, et al. MiR-125b acts as an oncogene in glioblastoma cells and inhibits cell apoptosis through p53 and p38MAPK-independent pathways. Br J Cancer. 2013;109(11):2853–2863. doi:10.1038/bjc.2013.67224169356
- Haemmig S, Baumgartner U, Gluck A, et al. miR-125b controls apoptosis and temozolomide resistance by targeting TNFAIP3 and NKIRAS2 in glioblastomas. Cell Death Dis. 2014;5:e1279. doi:10.1038/cddis.2014.24524901050
- Shi L, Wan Y, Sun G, Zhang S, Wang Z, Zeng Y. miR-125b inhibitor may enhance the invasion-prevention activity of temozolomide in glioblastoma stem cells by targeting PIAS3. BioDrugs. 2014;28(1):41–54. doi:10.1007/s40259-013-0053-223857508
- Huang T, Alvarez AA, Pangeni RP, et al. A regulatory circuit of miR-125b/miR-20b and Wnt signalling controls glioblastoma phenotypes through FZD6-modulated pathways. Nat Commun. 2016;7:12885. doi:10.1038/ncomms1288527698350
- Chen J, Fu X, Wan Y, Wang Z, Jiang D, Shi L. miR-125b inhibitor enhance the chemosensitivity of glioblastoma stem cells to temozolomide by targeting Bak1. Tumour Biol. 2014;35(7):6293–6302. doi:10.1007/s13277-014-1821-424643683
- Li Q, Han Y, Wang C, et al. MicroRNA-125b promotes tumor metastasis through targeting tumor protein 53-induced nuclear protein 1 in patients with non-small-cell lung cancer. Cancer Cell Int. 2015;15:84. doi:10.1186/s12935-015-0233-x26388699
- Liu Z, Smith KR, Khong HT, et al. miR-125b regulates differentiation and metabolic reprogramming of T cell acute lymphoblastic leukemia by directly targeting A20. Oncotarget. 2016;7(48):78667–78679. doi:10.18632/oncotarget.1201827637078
- Lin K-Y, Zhang X-J, Feng -D-D, et al. miR-125b, a target of CDX2, regulates cell differentiation through repression of the core binding factor in hematopoietic malignancies. J Biol Chem. 2011;286(44):38253–38263. doi:10.1074/jbc.M111.26967021903586
- Sui M, Jiao A, Zhai H, et al. Upregulation of miR-125b is associated with poor prognosis and trastuzumab resistance in HER2-positive gastric cancer. Exp Ther Med. 2017;14(1):657–663. doi:10.3892/etm.2017.454828672982
- Chang S, He S, Qiu G, et al. MicroRNA-125b promotes invasion and metastasis of gastric cancer by targeting STARD13 and NEU1. Tumour Biol. 2016;37(9):12141–12151. doi:10.1007/s13277-016-5094-y27220320
- Wu J-G, Wang -J-J, Jiang X, et al. MiR-125b promotes cell migration and invasion by targeting PPP1CA-Rb signal pathways in gastric cancer, resulting in a poor prognosis. Gastric Cancer. 2015;18(4):729–739. doi:10.1007/s10120-014-0421-825240408
- Wang M, Zhu X, Sha Z, Li N, Li D, Chen L. High expression of kinesin light chain-2, a novel target of miR-125b, is associated with poor clinical outcome of elderly non-small-cell lung cancer patients. Br J Cancer. 2015;112(5):874–882. doi:10.1038/bjc.2015.325668010
- Mei LL, Wang W-J, Qiu Y-T, Xie X-F, Bai J, Shi Z-Z. miR-125b-5p functions as a tumor suppressor gene partially by regulating HMGA2 in esophageal squamous cell carcinoma. PLoS One. 2017;12(10):e0185636. doi:10.1371/journal.pone.018563628968424
- Feng J, Fan Y, Ayiheng Q, Zhang H, Yong J, Hu B. MicroRNA-125b targeted STAT3 to inhibit laryngeal squamous cell carcinoma cell growth and motility. Oncol Lett. 2017;14(1):480–486. doi:10.3892/ol.2017.615528693195
- Bu Q, You F, Pan G, et al. MiR-125b inhibits anaplastic thyroid cancer cell migration and invasion by targeting PIK3CD. Biomed Pharmacother. 2017;88:443–448. doi:10.1016/j.biopha.2016.11.09028122310
- Avellini C, Licini C, Lazzarini R, et al. The trophoblast cell surface antigen 2 and miR-125b axis in urothelial bladder cancer. Oncotarget. 2017;8(35):58642–58653. doi:10.18632/oncotarget.v8i3528938585
- Zhao X, He W, Li J, et al. MiRNA-125b inhibits proliferation and migration by targeting SphK1 in bladder cancer. Am J Transl Res. 2015;7(11):2346–2354.26807182
- Huang L, Luo J, Cai Q, et al. MicroRNA-125b suppresses the development of bladder cancer by targeting E2F3. Int J Cancer. 2011;128(8):1758–1769. doi:10.1002/ijc.2550920549700
- Han Y, Liu Y, Zhang H, et al. Hsa-miR-125b suppresses bladder cancer development by down-regulating oncogene SIRT7 and oncogenic long noncoding RNA MALAT1. FEBS Lett. 2013;587:3875–3882. doi:10.1016/j.febslet.2013.10.02324396870
- Wu D, Ding J, Wang L, et al. microRNA-125b inhibits cell migration and invasion by targeting matrix metallopeptidase 13 in bladder cancer. Oncol Lett. 2013;5(3):829–834. doi:10.3892/ol.2013.112323425975
- Zhou HC, Fang JH, Shang LR, et al. MicroRNAs miR-125b and miR-100 suppress metastasis of hepatocellular carcinoma by disrupting the formation of vessels that encapsulate tumour clusters. J Pathol. 2016;240(4):450–460. doi:10.1002/path.480427577856
- Zhou JN, Zeng Q, Wang HY, et al. MicroRNA-125b attenuates epithelial-mesenchymal transitions and targets stem-like liver cancer cells through small mothers against decapentaplegic 2 and 4. Hepatology (Baltimore, Md). 2015;62(3):801–815. doi:10.1002/hep.27887
- Gong J, Zhang JP, Li B, et al. MicroRNA-125b promotes apoptosis by regulating the expression of Mcl-1, Bcl-w and IL-6R. Oncogene. 2013;32(25):3071–3079. doi:10.1038/onc.2012.31822824797
- Tsang FH, Au V, Lu WJ, et al. Prognostic marker microRNA-125b inhibits tumorigenic properties of hepatocellular carcinoma cells via suppressing tumorigenic molecule eIF5A2. Dig Dis Sci. 2014;59(10):2477–2487. doi:10.1007/s10620-014-3184-524811246
- Zhang J, Na S, Liu C, Pan S, Cai J, Qiu J. MicroRNA-125b suppresses the epithelial-mesenchymal transition and cell invasion by targeting ITGA9 in melanoma. Tumour Biol. 2016;37(5):5941–5949. doi:10.1007/s13277-015-4409-826596831
- Kappelmann M, Kuphal S, Meister G, Vardimon L, Bosserhoff AK. MicroRNA miR-125b controls melanoma progression by direct regulation of c-jun protein expression. Oncogene. 2013;32(24):2984–2991. doi:10.1038/onc.2012.30722797068
- Ying X, Wei K, Lin Z, et al. MicroRNA-125b suppresses ovarian cancer progression via suppression of the epithelial-mesenchymal transition pathway by targeting the SET protein. Cell Physiol Biochem. 2016;39(2):501–510. doi:10.1159/00044564227383536
- Lee M, Kim EJ, Jeon MJ. MicroRNAs 125a and 125b inhibit ovarian cancer cells through post-transcriptional inactivation of EIF4EBP1. Oncotarget. 2016;7(8):8726–8742. doi:10.18632/oncotarget.647426646586
- Guan Y, Yao H, Zheng Z, Qiu G, Sun K. MiR-125b targets BCL3 and suppresses ovarian cancer proliferation. Int J Cancer. 2011;128(10):2274–2283. doi:10.1002/ijc.2557520658525
- Wang F, Yu D, Liu Z, et al. MiR-125b functions as a tumor suppressor and enhances chemosensitivity to cisplatin in osteosarcoma. Technol Cancer Res Treat. 2016;15(6):NP105–NP112. doi:10.1177/153303461561884926744308
- Liu LH, Li H, Li JP, et al. miR-125b suppresses the proliferation and migration of osteosarcoma cells through down-regulation of STAT3. Biochem Biophys Res Commun. 2011;416(1–2):31–38. doi:10.1016/j.bbrc.2011.10.11722093834
- Wu Y, He H, Wu B, et al. miR-125b suppresses the aerobic glycolysis of osteosarcoma HOS cells by downregulating the expression of hexokinase-2. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2017;33(10):1365–1370.29169422
- Tang XY, Zheng W, Ding M, et al. miR-125b acts as a tumor suppressor in chondrosarcoma cells by the sensitization to doxorubicin through direct targeting the ErbB2-regulated glucose metabolism. Drug Des Devel Ther. 2016;10:571–583. doi:10.2147/DDDT.S90530
- Sun Y, Liu X, Zhang Q, et al. Oncogenic potential of TSTA3 in breast cancer and its regulation by the tumor suppressors miR-125a-5p and miR-125b. Tumour Biol. 2016;37(4):4963–4972. doi:10.1007/s13277-015-4178-426531722
- Hong L, Pan F, Jiang H, et al. miR-125b inhibited epithelial-mesenchymal transition of triple-negative breast cancer by targeting MAP2K7. Onco Targets Ther. 2016;9:2639–2648. doi:10.2147/OTT.S10271327226726
- Tang F, Zhang R, He Y, Zou M, Guo L, Xi T. MicroRNA-125b induces metastasis by targeting STARD13 in MCF-7 and MDA-MB-231 breast cancer cells. PLoS One. 2012;7(5):e35435. doi:10.1371/journal.pone.003543522693547
- Zhang Y, Yan LX, Wu QN, et al. miR-125b is methylated and functions as a tumor suppressor by regulating the ETS1 proto-oncogene in human invasive breast cancer. Cancer Res. 2011;71(10):3552–3562. doi:10.1158/0008-5472.CAN-10-243521444677
- Feliciano A, Castellvi J, Artero-Castro A, et al. miR-125b acts as a tumor suppressor in breast tumorigenesis via its novel direct targets ENPEP, CK2-alpha, CCNJ, and MEGF9. PLoS One. 2013;8(10):e76247. doi:10.1371/journal.pone.007624724098452
- Klinge CM, Radde BN, Imbert-Fernandez Y, et al. Targeting the intracellular MUC1 C-terminal domain inhibits proliferation and estrogen receptor transcriptional activity in lung adenocarcinoma cells. Mol Cancer Ther. 2011;10(11):2062–2071. doi:10.1158/1535-7163.MCT-11-038121862684
- Rajabi H, Jin C, Ahmad R, McClary C, Joshi MD, Kufe D. Mucin 1 oncoprotein expression is suppressed by the miR-125b oncomir. Genes Cancer. 2010;1(1):62–68. doi:10.1177/194760190935793320729973
- Yang D, Zhan M, Chen T, et al. miR-125b-5p enhances chemotherapy sensitivity to cisplatin by down-regulating Bcl2 in gallbladder cancer. Sci Rep. 2017;7:43109. doi:10.1038/srep4310928256505
- Shang C, Lu YM, Meng LR. MicroRNA-125b down-regulation mediates endometrial cancer invasion by targeting ERBB2. Medi Sci Monitor. 2012;18(4):Br149–155. doi:10.12659/MSM.882617
- Fujino Y, Takeishi S, Nishida K, et al. Downregulation of microRNA-100/microRNA-125b is associated with lymph node metastasis in early colorectal cancer with submucosal invasion. Cancer Sci. 2017;108(3):390–397. doi:10.1111/cas.1315228032929
- Zhang X, Ma X, An H, et al. Upregulation of microRNA-125b by G-CSF promotes metastasis in colorectal cancer. Oncotarget. 2017;8(31):50642–50654. doi:10.18632/oncotarget.1689228881590
- Gao D, Xiao Z, Li HP, Han DH, Zhang YP. The mechanism study of miR-125b in occurrence and progression of multiple myeloma. Cancer Med. 2018;7(1):134–145. doi:10.1002/cam4.118129214735
- Li J, You T, Jing J. MiR-125b inhibits cell biological progression of Ewing’s sarcoma by suppressing the PI3K/Akt signalling pathway. Cell Prolif. 2014;47(2):152–160. doi:10.1111/cpr.2014.47.issue-224517182
- Kim SW, Ramasamy K, Bouamar H, Lin AP, Jiang D, Aguiar RC. MicroRNAs miR-125a and miR-125b constitutively activate the NF-kappaB pathway by targeting the tumor necrosis factor alpha-induced protein 3 (TNFAIP3, A20). Proc Natl Acad Sci U S A. 2012;109(20):7865–7870. doi:10.1073/pnas.120008110922550173
- Garbar C, Mascaux C, Giustiniani J, et al. Autophagy is decreased in triple-negative breast carcinoma involving likely the MUC1-EGFR-NEU1 signalling pathway. Int J Clin Exp Pathol. 2015;8(5):4344–4355.26191126
- Zheng L, Li X, Chou J, et al. StarD13 3ʹ-untranslated region functions as a ceRNA for TP53INP1 in prohibiting migration and invasion of breast cancer cells by regulating miR-125b activity. Eur J Cell Biol. 2018;97(1):23–31. doi:10.1016/j.ejcb.2017.11.00229146309
- El-Asrag ME, Sergouniotis PI, McKibbin M, et al. Biallelic mutations in the autophagy regulator DRAM2 cause retinal dystrophy with early macular involvement. Am J Hum Genet. 2015;96(6):948–954. doi:10.1016/j.ajhg.2015.04.00625983245
- Sui X, Kong N, Ye L, et al. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014;344(2):174–179. doi:10.1016/j.canlet.2013.11.01924333738
- Long J, Xie Y, Yin J, Lu W, Fang S. SphK1 promotes tumor cell migration and invasion in colorectal cancer. Tumour Biol. 2016;37(5):6831–6836. doi:10.1007/s13277-015-4542-426662312
- Tripathi V, Shen Z, Chakraborty A, et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB. PLoS Genet. 2013;9(3):e1003368. doi:10.1371/journal.pgen.100336823555285
- Bao Y, Lu Y, Wang X, et al. Eukaryotic translation initiation factor 5A2 (eIF5A2) regulates chemoresistance in colorectal cancer through epithelial mesenchymal transition. Cancer Cell Int. 2015;15:109. doi:10.1186/s12935-015-0250-926581310
- He LR, Zhao HY, Li BK, et al. Overexpression of eIF5A-2 is an adverse prognostic marker of survival in stage I non-small cell lung cancer patients. Int J Cancer. 2011;129(1):143–150. doi:10.1002/ijc.2566920830705
- Hua S, Quan Y, Zhan M, Liao H, Li Y, Lu L. miR-125b-5p inhibits cell proliferation, migration, and invasion in hepatocellular carcinoma via targeting TXNRD1. Cancer Cell Int. 2019;19:203. doi:10.1186/s12935-019-0919-631384178
- Funa NS, Schachter KA, Lerdrup M, et al. Beta-catenin regulates primitive streak induction through collaborative interactions with SMAD2/SMAD3 and OCT4. Cell Stem Cell. 2015;16(6):639–652. doi:10.1016/j.stem.2015.03.00825921273
- Wang Y, Wang B, Xiao S, Li Y, Chen Q. miR-125a/b inhibits tumor-associated macrophages mediated in cancer stem cells of hepatocellular carcinoma by targeting CD90. J Cell Biochem. 2019;120(3):3046–3055. doi:10.1002/jcb.v120.330536969
- Maes ME, Schlamp CL, Nickells RW. BAX to basics: how the BCL2 gene family controls the death of retinal ganglion cells. Prog Retin Eye Res. 2017;57:1–25. doi:10.1016/j.preteyeres.2017.01.00228064040
- Gupta SK, Oommen S, Aubry MC, Williams BP, Vlahakis NE. Integrin alpha9beta1 promotes malignant tumor growth and metastasis by potentiating epithelial-mesenchymal transition. Oncogene. 2013;32(2):141–150. doi:10.1038/onc.2012.4122370635
- Rambow F, Bechadergue A, Luciani F, et al. Regulation of melanoma progression through the TCF4/miR-125b/NEDD9 cascade. J Invest Dermatol. 2016;136(6):1229–1237. doi:10.1016/j.jid.2016.02.80326968260
- Zhao H, Wang W, Zhao Q, Hu G, Deng K, Liu Y. BCL3 exerts an oncogenic function by regulating STAT3 in human cervical cancer. Onco Targets Ther. 2016;9:6619–6629. doi:10.2147/OTT.S11818427822067
- Ferracin M, Bassi C, Pedriali M, et al. miR-125b targets erythropoietin and its receptor and their expression correlates with metastatic potential and ERBB2/HER2 expression. Mol Cancer. 2013;12(1):130. doi:10.1186/1476-4598-12-13024165569
- Suganuma T, Ino K, Shibata K, et al. Regulation of aminopeptidase A expression in cervical carcinoma: role of tumor-stromal interaction and vascular endothelial growth factor. Lab Invest. 2004;84(5):639–648. doi:10.1038/labinvest.370007215048132
- Wang L, Huang J, Jiang M, Lin H. Tissue-specific transplantation antigen P35B (TSTA3) immune response-mediated metabolism coupling cell cycle to postreplication repair network in no-tumor hepatitis/cirrhotic tissues (HBV or HCV infection) by biocomputation. Immunol Res. 2012;52(3):258–268. doi:10.1007/s12026-012-8337-z22528125
- Scott GK, Goga A, Bhaumik D, Berger CE, Sullivan CS, Benz CC. Coordinate suppression of ERBB2 and ERBB3 by enforced expression of micro-RNA miR-125a or miR-125b. J Biol Chem. 2007;282(2):1479–1486. doi:10.1074/jbc.M60938320017110380
- Nam S, Long X, Kwon C, Kim S, Nephew KP. An integrative analysis of cellular contexts, miRNAs and mRNAs reveals network clusters associated with antiestrogen-resistant breast cancer cells. BMC Genomics. 2012;13:732. doi:10.1186/1471-2164-13-73223270413
- Dong H, Hu J, Zou K, et al. Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast cancer. Mol Cancer. 2019;18(1):3. doi:10.1186/s12943-018-0931-930621694
- Rossi RL, Rossetti G, Wenandy L, et al. Distinct microRNA signatures in human lymphocyte subsets and enforcement of the naive state in CD4+ T cells by the microRNA miR-125b. Nat Immunol. 2011;12(8):796–803. doi:10.1038/ni.205721706005
- Chaudhuri AA, So AY, Sinha N, et al. MicroRNA-125b potentiates macrophage activation. J Immunol. 2011;187(10):5062–5068. doi:10.4049/jimmunol.110200122003200
- Li G, Zan H, Xu Z, Casali P. Epigenetics of the antibody response. Trends Immunol. 2013;34(9):460–470. doi:10.1016/j.it.2013.03.00623643790
- Wang JK, Wang Z, Li G. MicroRNA-125 in immunity and cancer. Cancer Lett. 2019;454:134–145. doi:10.1016/j.canlet.2019.04.01530981762
- Zhu Y, Zhang S, Li Z, et al. miR-125b-5p and miR-99a-5p downregulate human gammadelta T-cell activation and cytotoxicity. Cell Mol Immunol. 2019;16(2):112–125. doi:10.1038/cmi.2017.16429429995
- Li G, So AY, Sookram R, et al. Epigenetic silencing of miR-125b is required for normal B-cell development. Blood. 2018;131(17):1920–1930. doi:10.1182/blood-2018-01-82454029555645
- Parayath NN, Gandham SK, Leslie F, Amiji MM. Improved anti-tumor efficacy of paclitaxel in combination with MicroRNA-125b-based tumor-associated macrophage repolarization in epithelial ovarian cancer. Cancer Lett. 2019;461:1–9. doi:10.1016/j.canlet.2019.07.00231288064
- Parayath NN, Parikh A, Amiji MM. Repolarization of tumor-associated macrophages in a genetically engineered nonsmall cell lung cancer model by intraperitoneal administration of hyaluronic acid-based nanoparticles encapsulating microRNA-125b. Nano Lett. 2018;18(6):3571–3579. doi:10.1021/acs.nanolett.8b0068929722542
- Zuo D, Chen L, Liu X, et al. Combination of miR-125b and miR-27a enhances sensitivity and specificity of AFP-based diagnosis of hepatocellular carcinoma. Tumour Biol. 2016;37(5):6539–6549. doi:10.1007/s13277-015-4545-126637228
- Wei X, Chen D, Lv T, Li G, Qu S. Serum microRNA-125b as a potential biomarker for glioma diagnosis. Mol Neurobiol. 2016;53(1):163–170. doi:10.1007/s12035-014-8993-125416859
- Luo Y, Wang X, Niu W, et al. Elevated microRNA-125b levels predict a worse prognosis in HER2-positive breast cancer patients. Oncol Lett. 2017;13(2):867–874. doi:10.3892/ol.2016.548228356971
- Vilquin P, Donini CF, Villedieu M, et al. MicroRNA-125b upregulation confers aromatase inhibitor resistance and is a novel marker of poor prognosis in breast cancer. Breast Cancer Res. 2015;17:13. doi:10.1186/s13058-015-0515-125633049
- Liu B, Su F, Chen M, et al. Serum miR-21 and miR-125b as markers predicting neoadjuvant chemotherapy response and prognosis in stage II/III breast cancer. Hum Pathol. 2017;64:44–52. doi:10.1016/j.humpath.2017.03.01628412211
- Incoronato M, Grimaldi AM, Mirabelli P, et al. Circulating miRNAs in untreated breast cancer: an exploratory multimodality morpho-functional study. Cancers. 2019;11:6. doi:10.3390/cancers11060876
- Malczewska A, Frampton AE, Mato Prado M. et al. Circulating microRNAs in small-bowel neuroendocrine tumors: a potential tool for diagnosis and assessment of effectiveness of surgical resection. Ann Surg;2019 1. doi:10.1097/SLA.0000000000003502
- El-Khazragy N, Elshimy AA, Hassan SS, et al. Dysregulation of miR-125b predicts poor response to therapy in pediatric acute lymphoblastic leukemia. J Cell Biochem. 2018.
- Sun B, Zhang Y, Zhou L, et al. The proliferation of cervical cancer is promoted by miRNA-125b through the regulation of the HMGA1. Onco Targets Ther. 2019;12:2767–2776. doi:10.2147/OTT.S19774031043793
- Misso G, Zarone MR, Lombardi A, et al. miR-125b upregulates miR-34a and sequentially activates stress adaption and cell death mechanisms in multiple myeloma. Mol Ther Nucl Acids. 2019;16:391–406. doi:10.1016/j.omtn.2019.02.023