
Abstract
Purpose
The effects of miR-139 on the tumorigenicity of triple negative breast cancer (TNBC) and the underlying mechanisms were investigated.
Methods
Normal human breast epithelial (MCF-10A) and TNBC cell lines (HCC1806 and BT549) were used for microRNA (miR)-139 overexpression, SOX8 overexpression, and knockdown studies as in vitro models of TNBC. The expression of SOX8 and miR-139 was detected by reverse transcription-polymerase chain reaction. CCK8 and clone formation assays were used to evaluate cell proliferation ability. Transwell assays and flow cytometry were used to test cell migration and apoptosis, respectively. Cell tumorigenicity was examined by tumor sphere formation assays. The interaction between miR-139 and SOX8 was examined by dual-luciferase reporter assays. The expression of SOX8, cleaved caspase-3, and cleaved caspase-9 was analyzed by Western blotting. The findings were validated in vivo using a nude mouse transplanted tumor model.
Results
SOX8 expression was higher (P < 0.05) and miR-139 expression was lower (P < 0.05) in HCC1806 and BT549 cells than in MCF-10A cells. SOX8 overexpression significantly enhanced cell proliferation and migration, reduced the rate of cell apoptosis, and increased tumor sphere formation (P < 0.05) compared with the control group, whereas SOX8 knockdown had the opposite effect (P < 0.05). Overexpression of miR-139 markedly decreased cell proliferation and migration, increased cell apoptosis in vitro, and decreased tumor angiogenesis and volume in vivo (P < 0.05).
Conclusion
miR-139 suppressed the tumorigenicity of TNBC cells by targeting SOX8.
Introduction
Breast cancer is one of the most common cancers in women worldwide and its incidence is increasing.Citation1 Triple negative breast cancer (TNBC) is a form of breast cancer characterized by negative estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor 2 (HER2) expression. It accounts for approximately 15–20% of all breast cancers, and is the most invasive and aggressive type of breast cancer.Citation2 Because of limited treatment options, poor response to systemic therapy, and high aggressiveness and early widespread metastasis, the overall prognosis of patients with TNBC remains dismal.Citation3 Moreover, the high degree of heterogeneity and the complexity of molecular mechanisms limit the development of novel therapeutic strategies.Citation4,Citation5 Currently, adjuvant chemotherapy for TNBC is based on standard chemotherapy regimens including anthracyclines, taxol, and cyclophosphamide.Citation6 Investigating how to prevent, delay, or even suppress the rapid growth of TNBC has become a hot area of basic and clinical research in recent years.
MicroRNAs (miRNAs) are a group of small non-protein-coding RNA molecules of 20–22 nucleotides in length that can regulate gene expression by binding to specific target sites of mRNAs. They have been implicated in various diseases and are regarded as potential biomarkers or key regulators of numerous cellular processes.Citation7,Citation8 Many miRNAs are involved in the biological process of breast cancer.Citation7 The dysregulation of miRNA expression is closely related to the mechanism of disease development, especially the occurrence of invasion and migration in breast cancer.Citation9,Citation10 Accumulating evidence indicates that miR-139-5p may play a significant role in cancer biology, diagnosis, prognosis, and therapy.Citation11 A previous study demonstrated that miR-139-5p may serve as a novel serum biomarker for colorectal cancer recurrence and metastasis.Citation12 Watanabe et al reported that silencing miR-139 by histone methylation promotes non-small-cell lung cancer cell invasion.Citation13 Pajic et al reported that miR-139 suppresses proliferation, migration, and invasion of tumor cells in epithelial ovarian cancer and in breast cancer.Citation4 However, there have been few studies on the effects of miR-139 in TNBC.
In this study, we investigated the role of miR-139 in TNBC and the underlying molecular mechanisms in vitro and in vivo.
Materials and Methods
Cell Culture
The normal human breast epithelial cell line MCF-10A and the TNBC cell lines HCC1806 and BT549 were purchased from Shanghai Institute of Biochemistry and Cell Biology affiliated with the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in DMEM/F12 medium (Gibco, Invitrogen, Grand Island, NY, USA) supplemented with 1% penicillin/streptomycin and 10% fetal bovine serum in a constant temperature incubator (DRH-A100, Shanghai, China) with 5% CO2 at 37°C. Cells were collected at the logarithmic growth phase when reaching approximately 80% confluence.
Detection of Sry-Like High-Mobility Group Box (SOX8) mRNA Expression in Cells
Cells were centrifuged at 4°C (12,000 rpm, 15 min) after harvesting and total RNA was extracted and isolated using the TRIzol Kit (Takara, Dalian, China). The OD260/OD280 value (between 1.8 and 2.0) was used as an indicator of RNA purity. cDNA was transcribed using a reverse transcription kit (Roche, 11939823001, Shanghai, China). A Mastercycler® nexus X2 (Eppendorf, Guangzhou, China) was used to perform reverse transcription-PCR (RT-PCR). The reaction conditions were as follows: 15 s at 95°C, 60 s at 60°C, and 40 s at 72°C (35 cycles). The data were organized and analyzed using the 2−ΔΔCt method. The relative expression level was calculated after normalizing to GAPDH mRNA as the internal control. The sequences of the primers (Shenggong Bioengineering Technology Service Co., Ltd., Shanghai, China) used were the following.
miR-139: 5′-GCCTCTACAGTGCACGTGTCTC-3′ (F), 5′-CGCTGTTCTCATCATCTGTCTCGC-3′ (R);
U6: 5′-GACCTCTATGCCAACACAGT-3′(F), 5′-AGTACTTGCGCTCAGGAGGA-3′ (R);
SOX8: 5′-CGAGAGAAGACGCCTGCT-3′ (F),
5′-CGTGTTGGAGAATGAGGG-3′ (R);
GAPDH: 5′-TGACTTCAACAGCGACACCCA-3′ (F), 5′-CACCCTGTTGCTGTAGCCAAA-3′ (R).
Dual-Luciferase Reporter Assay
Wild-type (wt) and mutant (mut) 3ʹ untranslated regions (3ʹ-UTRs) of SOX8 were amplified in the pGL3/luciferase vector (Promega, E1751, Madison, WI, USA) and cloned downstream of the luciferase gene. The luciferase activities of HCC1806 and BT549 cells were detected using the dual luciferase reporter system following the manufacturer’s instructions (Promega) at 48 h after transfection.Citation14
Cell Transfection
Cells were cultured 1 day before transfection and inoculated into 6-well plates for lentivirus transfection.Citation15 Full length SOX8 cDNA cloned into pcDNA3.1 mammalian expression plasmid (Shanghai GeneChem Co., Ltd, China). TNBC cells (HCC1806 and BT549) were divided into the following five groups according to treatment: 1) blank control group (BC group, no transfection), 2) SOX8 overexpression group (SOX8 group, cells were transfected with SOX8 pcDNA3.1 using Lipofectamine 3000), 3) SOX8 overexpression negative control group (NC1 group, cells transfected with SOX8 negative control), 4) SOX8 silencing group (si-S group, stable shSOX8 cells were constructed using short hairpin RNA (shRNA) with GeneChem), and 5) SOX8 silencing negative control group (NC2 group, cells transfected with shSOX8 negative control). RT-PCR and Western blotting were used to detect the transfection efficiency.
To investigate the effects of miR-139 on TNBC cells, the cells were grouped as follows: 1) blank control group (BC, no transfection), 2) miR-139 mimic group (miR-139, transfected with miR-139 mimic); and 3) and miR-139 analogue negative control group (miR-NC, transfected with unrelated mimic as negative control). The transfection efficiency and the expression of SOX8 in cells were detected by RT-PCR. miR-139 mimic and its negative control (NC), synthesized by Life Technologies Corporation, were transfected into cells using Lipofectamine 3000 reagent and the transfected cells were harvested at 72 h post-transfection.
To further explore the role of miR-139 in targeting SOX8, cells were divided into four groups as follows: 1) blank control group (BC), 2) miR-139 mimic group (miR-139), 3) SOX8 silencing group (si-S), and 4) miR-139 mimics + SOX8 overexpression group (miR+S), in which cells were transfected with miR-139 mimic and SOX8 pcDNA3.1. Experiments were performed in triplicate for each group.
Cell Proliferation Assay
Transfected cells in the logarithmic growth stage were seeded into 96-well plates at a density of 2 × 104 cells/mL with each well containing 100 μL. Cells were incubated in a 5% CO2 incubator at 37°C for 24, 48, 72, and 96 h, and CCK-8 solution (10 μL, Tongren Institute of Chemistry, Japan) was added to each well. The solution was mixed and cultured for 4 h. The absorbance (OD) of each well was quantified by adjusting to the blank control as zero at 450 nm.
Plate Clone Formation Assay
Cells were grown to the logarithmic growth phase, harvested, and digested with 0.25% trypsin to generate single cell suspensions. Cell density was adjusted to 250 cells/mL, and 2 mL of suspension was seeded into each well of a 6-well plate. The plate was incubated at 37°C under 5% CO2 for 2–3 weeks, and the culture media were replaced every 3 days. Cells were then fixed with methanol, and 1 mL Giemsa working fluid (G1015, Solarbio, Beijing, China) was added to each well and incubated for 30 min. Finally, the plate was washed with ultrapure water and photographed using a camera after the water around the plate was removed with filter paper.
Cell Migration
Pre-warmed DMEM medium (50 μL per well) was evenly spread on the bottom of the upper chambers of a Transwell plate (3413, Corning, Beijing, China). Next, 100 μL of cell suspension (5 × 104) was added to each well and incubated for 4 h at 37°C. In the lower chamber, 600 μL DMEM/F12 medium was added and cultured for 72 h in the same incubator. The chamber was then washed twice with PBS and fixed with 5% glutaraldehyde at 4°C. After two washes, the plates were stained with 0.1% Crystal Violet for 30 min and five fields (400×) of view were randomly selected using an inverted microscope (Olympus, Japan) to calculate the mean value of migrated cells.
Apoptosis Detection
Cells treated as indicated were incubated for 24 h, collected, resuspended in 1× precooled PBS (4°C), and centrifuged for 5–10 min at 1000 rpm/min. After washing, cells were suspended with 1× binding buffer (300 μL), and 5 μL of Annexin V-APC (CA1020, Solarbio, Beijing, China) was added and mixed well. Cells were kept in the dark and incubated for 15 min at room temperature. Propidium iodide (5 μL) was added 5 min before loading, followed by the addition of 1× binding buffer (200 μL). Finally, flow cytometry (Beckman Coulter, Brea, CA, USA) was used to analyze samples, and the data were analyzed using CellQuest software (BD Bioscience, San Diego, CA, USA).
Western Blotting Analysis
Protein lysates were obtained from cells using a protein extraction kit (W037, Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Total proteins (30 μg) were subjected to 10% SDS-PAGE electrophoretic separation and the resolved proteins were transferred onto a polyvinylidene difluoride (PVDF) membrane (W016-1-1, Nanjing Jiancheng Bioengineering Institute). Following blocking with non-fat milk powder for 2 h, membranes were incubated with rabbit anti-human SOX8 (1:1000, ab226983, Abcam, Bejing, China), rabbit anti-human cleaved caspase-3 (1:4000, ab49822, Abcam), rabbit anti-human cleaved caspase-9 (1:20,000, ab2324, Abcam), rabbit anti-human β-actin (1:20,000, ab2324, Abcam) at 4°C for 12 h. Subsequently, membranes were incubated with a goat anti-rabbit HRP-conjugated IgG H&L (1:20,000, ab205718, Abcam) secondary antibody for 50 min. The protein bands were visualized using an ECL chemiluminescence kit (32132, ThermoFisher, Shanghai, China) and β-actin was selected as a control to calculate the relative expression of the target proteins.
Tumor Sphere Formation Assays
DMEM/F12 serum-free medium (containing 1% penicillin-streptomycin, 20 ng/mL EGF, 20 ng/mL bFGF, and 2% B27) was used to resuspend the cells, and the concentration was adjusted to 1 × 104/mL. Cells were inoculated in a low adhesion 6-well plate at a density of 5000 cells per well, and the solution was changed every other day. After 7 days of culture, tumor spheres were collected when they reached a diameter of 50 μm, digested, dispersed into single cells, and reinoculated to form new tumor spheres. After 1 week, the number of tumor spheres with a diameter >50 μm was counted under an inverted microscope. The expression of miR-139 in cells was detected by RT-PCR following RNA extraction of tumor spheres.
In vivo Experiment Using a Murine TNBC Model
Specific pathogen-free grade BALB/C female nude mice (n=48) (4-weeks-old, 16–18 g body weight) were obtained from the Beijing Vital River Laboratory Animal Technology Co., Ltd. [SCXK (Jing) 20190006, Beijing, China] and maintained in a sterile cage in an air laminar flow purifying room. The room was kept under constant temperature (26–28°C) and humidity (40–60%), and the feed, drinking water, and bedding were sterilized.
The 48 mice were randomly divided into a blank control group (BC), miR-139 mimic group (miR-139), SOX8 silencing group (si-SOX8), miR-139 mimic and SOX8 pcDNA3.1 group (miR+S), with 12 mice in each group. TNBC HCC1806 and BT549 cells in the logarithmic growth stage were digested with 0.25% trypsin, collected, and counted. The cell concentration was adjusted to 5 × 107/mL, and the cell suspension (0.1 mL) was injected into the soft part of the skin on the back of the right forelimb of nude mice.
The tumor volume was calculated by measuring tumor length (L) and short diameter (W) using a Vernier caliper every 7 days as follows: (V) = (long diameter × short diameter2)/2. After 28 days, mice were intraperitoneally anesthetized with 0.6% pentobarbital sodium (40 mg/kg) and euthanized. The tumor tissue was extracted and tumors were weighed.
Statistical Analyses
Analysis was performed using SPSS, version 19.0 (IBM, Chicago, IL, USA), and results are presented as the mean ± standard deviation (SD). The differences among multiple groups were calculated by one-way analysis of variance (ANOVA) and Dunnett’s t-test. A value of P < 0.05 was considered statistically significant.
Results
miR-139 Bound to the 3ʹUTR of SOX8 in Dual Luciferase Reporter Assay
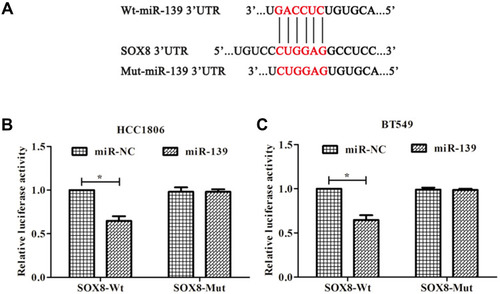
SOX8 was predicted as a candidate target of miR-139 using TargetScan (http://www.targetscan.org) (). A dual-luciferase reporter assay was performed to confirm that miR-139 targeted SOX8 in HCC1806 () and BT549 cell lines (). The results showed that when miR-139 mimic was co-transfected with wild-type SOX8 3ʹ-UTR, the luciferase activity decreased (P < 0.05), while a reversal of luciferase expression was observed in experiments where miR-139 mimic was co-transfected with mut-SOX8.
Figure 1 SOX8 is a target gene of miR-139 in TNBC cells. (A) The predicted binding sites of miR-139 and the SOX8 3ʹ-UTR region according to TargetScan. (B, C) Luciferase activity in cells following co-transfection with miR-139 mimic and luciferase reporters containing wild type (wt)- or mutated (mut)-SOX8 transcript in HCC1806 cells and BT549 cells. *P < 0.05.
SOX8 Promoted the Proliferation of TNBC Cells
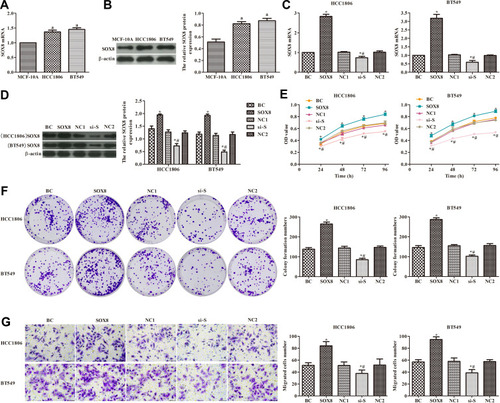
As shown in , the expression of SOX8 mRNA and protein was significantly higher in HCC1806 and BT549 cells than in MCF-10A cells (P < 0.05). Differences in expression of SOX8 in HCC1806 and BT549 cells were observed by RT-PCR. In the SOX8 group, the expression of SOX8 was significantly increased compared with the control group (P < 0.05, ), and the expression of SOX8 was clearly decreased in the si-SOX8-treated group compared with the control group (P < 0.05, ). In order to analyze the effects of SOX8 on the proliferation of TNBC cells, cell proliferation was measured by CCK-8 expression (), plate clone formation assays (), and the transwell migration assay (). The results showed that SOX8 overexpression notably increased the TNBC cell viability and migration, and silencing SOX8 expression significantly decreased the TNBC cell viability and migration, when compared with control cells (P < 0.05). These data suggested that higher expression of SOX8 in TNBC cells promoted cell proliferation.
Figure 2 Effect of SOX8 on the proliferation of TNBC cells. (A and B) Expression of SOX8 mRNA in cells measured by RT-PCR and Western blotting analysis. (C and D) Transfection efficiency of SOX8 evaluated by RT-PCR and Western blotting analysis. (E) Cell viability detected by the CCK8 assay. (F) Cell proliferation was measured by plate clone formation. (G) Cell migration ability detected by transwell assay (400×). In each group, triplicate experiments were performed. The differences among groups were analyzed by ANOVA followed by the Dunnett’s t-test. Compared with MCF-10A cells, aP < 0.05; compared with the BC group, *P < 0.05; compared with the SOX8 group, #P < 0.05.
SOX8 Overexpression Inhibited Apoptosis of TNBC Cells and Promoted Tumor Sphere Formation
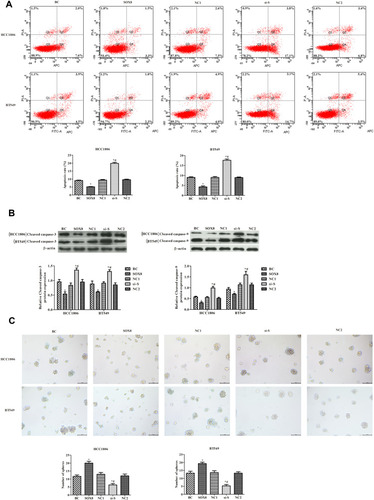
The effects of SOX8 expression on TNBC cell apoptosis were analyzed by flow cytometry (). The apoptosis rate was markedly decreased in the SOX8 group and increased in the si-SOX8-treated group, when compared with the control group (P < 0.05). The expression of cleaved caspase-3 and cleaved caspase-9 was observed using Western blotting analysis (). Consistent with the results of the apoptosis rate, the levels of cleaved caspase-3 and cleaved caspase-9 were significantly increased in the si-S group and decreased in the SOX8 group, compared with the control group (P < 0.05). The formation of tumor spheres of HCC1806 and BT549 cells was also observed (). Compared with the control group, the number of tumor spheres was significantly higher in the SOX8 group overexpressing SOX8, while the number of tumor spheres was significantly lower in the si-S group (P < 0.05). These results indicated that higher expression of SOX8 in TNBC cells inhibited cell apoptosis and promoted tumor sphere formation in vitro.
Figure 3 Effects of SOX8 on TNBC cell apoptosis and tumor sphere formation. (A) Apoptosis was detected by flow cytometry. (B) The expression of cleaved caspase-3 and cleaved caspase-9 was analyzed by Western blotting analysis. (C) Tumorigenicity of cells was determined by in vitro tumor sphere formation assays (100×). In each group, experiments were performed in triplicate. The differences among groups were analyzed by ANOVA followed by the Dunnett’s t-test. Compared with the BC group, *P < 0.05; compared with the SOX8 group, #P < 0.05.
Overexpression of miR-139 Inhibited the Proliferation of TNBC Cells
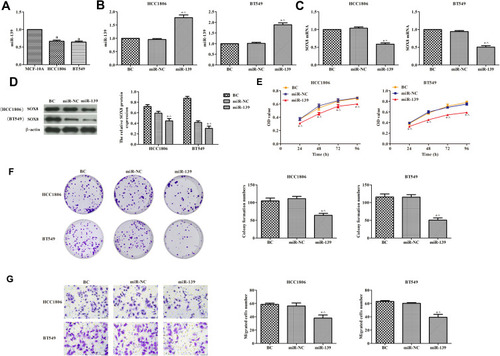
As shown in , the expression of miR-139 was remarkably lower in HCC1806 and BT549 cells than in MCF-10A cells (P < 0.05), indicating that miR-139 was downregulated in TNBC cells. In order to evaluate the effects of miR-139 on the proliferation of TNBC cells, the overexpression of miR-139 was induced in HCC1806 and BT549 cells by transfecting miR-139 mimic (). We found that the expression of SOX8 was lower in cells overexpressing miR-139 compared with the control group (, P < 0.05). The proliferation of TNBC cells was tested by measuring CCK-8 (), the plate clone formation assay (), and transwell migration assay (). High expression of miR-139 significantly inhibited the viability of TNBC cells and their migration when compared with control cells (P < 0.05). These data suggested that miR-139 overexpression inhibited the proliferation of TNBC cells.
Figure 4 Effects of miR-139 on the proliferation of TNBC cells. (A) RT-PCR was used to detect the expression of miR-139 in cells. (B) Transfection efficiency of miR-139 evaluated by RT-PCR. (C and D) RT-PCR and Western blotting analysis were used to detect the expression of SOX8 in cells. (E) Cell viability detected by CCK8 assay. (F) Cell proliferation was measured by plate clone formation. (G) Transwell assays were used to detect cell migration ability (400×). In each group, experiments were performed in triplicate. The differences among groups were analyzed by ANOVA with followed by the Dunnett’s t-test. Compared with MCF-10A cells, aP < 0.05; compared with the BC group, *P < 0.05; compared with the miR-NC group, ^P < 0.05.
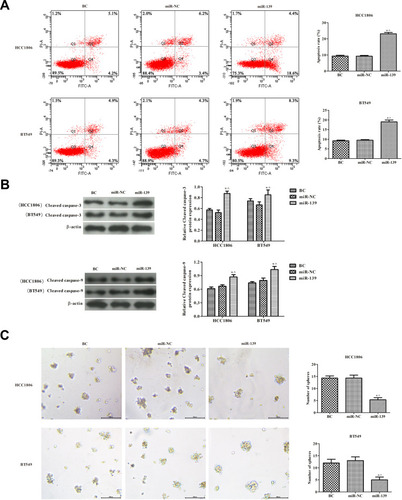
miR-139 Induced Apoptosis of TNBC Cells and Inhibited Tumor Sphere Formation
The effects of miR-139 expression on TNBC cell apoptosis were analyzed by flow cytometry (). The apoptosis rate was markedly higher in the miR-139 group compared with the control group (P < 0.05). The expression of both cleaved caspase-3 and caspase-9 was observed by Western blotting (). Consistent with the results of the apoptosis rate, the levels of cleaved caspases were significantly increased in the miR-139 group, compared with the control group (P < 0.05). The tumor sphere formation of HCC1806 and BT549 cells was also observed (). Compared with the control group, the number of tumor spheres was significantly lower in cells overexpressing miR-139 (P < 0.05). These results revealed that higher expression of miR-139 in TNBC cells promoted cell apoptosis and restrained tumor sphere formation in vitro.
Figure 5 Effects of miR-139 on TNBC cell apoptosis and tumor sphere formation. (A) Cell apoptosis was detected by flow cytometry. (B) The expression of cleaved caspase-3 and cleaved caspase-9 was analyzed by Western blotting analysis. (C) Tumorigenicity of cells was determined by in vitro tumor sphere formation assays (100×). In each group, experiments were performed in triplicate. The differences among groups were analyzed by ANOVA followed by the Dunnett’s t-test. Compared with the BC group, *P < 0.05; compared with the miR-NC group, ^P < 0.05.
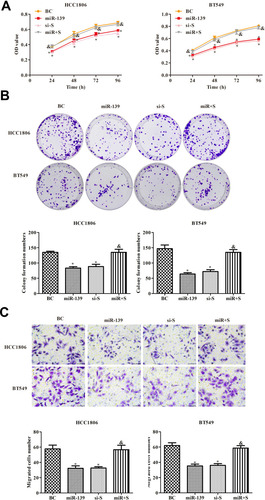
miR-139 Inhibited the Proliferation of TNBC Cells by Targeting SOX8
We further investigated whether miR-139 regulated the proliferation of TNBC cells via targeting SOX8 expression. Overexpression of miR-139 and SOX8 was established in TNBC cells. The proliferation of TNBC cells was analyzed by measuring CCK-8 (), the plate clone formation assay (), and by transwell migration assay (). The results showed that high expression of miR-139 inhibited the proliferation of TNBC cells, and high expression of SOX8 countered the effect of miR-139 overexpression (P < 0.05). These data revealed that miR-139 suppressed proliferation of TNBC cells via regulating SOX8.
Figure 6 Effects of miR-139 on the proliferation of TNBC cells by targeting SOX8. (A) CCK8 was used to detect cell viability. (B) Plate clone formation experiment was used to detect cell proliferation. (C) Transwell assay was used to detect cell migration (400×). In each group, experiments were performed in triplicate. The differences among groups were analyzed by ANOVA followed by the Dunnett’s t-test. Compared with the BC group, *P < 0.05; compared with the miR-139 group, &P < 0.05.
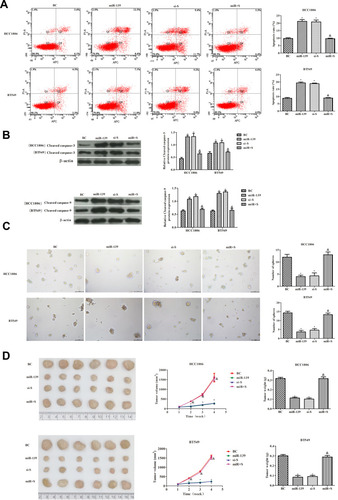
miR-139 Induced Apoptosis of TNBC Cells and Inhibited Tumor Formation by Targeting SOX8
The apoptosis rate of TNBC cells was observed to determine whether miR-139 induced apoptosis via regulating SOX8. As shown in , the apoptosis rate was higher in the miR+S group compared with the miR-139 group (P < 0.05). Similarly, the levels of cleaved caspase-3 and cleaved caspase-9 were clearly decreased in the miR+S group compared with the miR-139 group (P < 0.05, ). Further, the tumor sphere formation of TNBC cells was significantly higher in the miR+S group than in the miR-139 group (P < 0.05, ). In vivo, tumor formation was consistent with the results observed in vitro (). The tumor volume and weight were significantly higher in the miR+S group than those in the miR-139 group (P < 0.05). Overall, these data suggested that miR-139 suppressed the tumorigenicity of TNBC cells by targeting SOX8.
Figure 7 Effects of miR-139 targeting SOX8 on TNBC cell apoptosis and tumor formation. (A) Apoptosis of HCC1806 and BT549 cells measured by flow cytometry. (B) The expression of cleaved caspase-3 and cleaved caspase-9 was analyzed by Western blotting analysis. (C) In vitro, tumor sphere formation assays were used to detect the tumorigenicity of HCC1806 and BT549 cells (100×). (D) In vivo, tumor volume and tumor weight of TNBC cells. In each group, there were six mice. The differences among groups were analyzed by ANOVA followed by Dunnett’s t-test. Compared with the BC group, *P < 0.05; compared with the mir-139 group, &P < 0.05.
Discussion
In previous studies, miR-139 has been found to be associated with aggressive tumor behavior and disease progression in breast cancer. MiR-139 expression was thus identified as a surrogate marker for tumor aggressiveness in breast cancer and was found to be downregulated in tumor tissues compared with controls.Citation16–Citation18 SOX8 as a functional oncogene that is involved in the maintenance of stem-like capacities in TNBC cells, and its high expression has been significantly associated with poor prognosis in TNBC patients.Citation19,Citation20 However, whether miR-139 modulates TNBC cells and the underlying molecular mechanisms remain unclear. In this study, the dual-luciferase reporter assay was performed to confirm that miR-139 targets SOX8 in HCC1806 and BT549 cells. Further, we observed higher expression of SOX8 and lower expression of miR-139 in HCC1806 and BT549 cells, when compared with the normal human breast epithelial cell line MCF-10A. These findings support that miR-139 is a potential anti-metastatic miRNA and SOX8 is a potential pro-metastatic gene.
SOX genes have been well studied in many cancers, and have been described as oncogenes or tumor suppressors involved in tumor formation and progression.Citation21 As a member of the SOXE subfamily, SOX8 is an important functional oncogene that promotes tumor growth in tongue squamous cell carcinoma, with poor prognosis.Citation21,Citation22 In the present study, SOX8 overexpression enhanced the proliferation of TNBC and decreased TNBC cell apoptosis. The expression levels of cleaved caspase-3 and cleaved caspase-9 were also evaluated, which were decreased after SOX8 overexpression. When the expression of SOX8 was inhibited, the proliferation of TNBC was suppressed and apoptosis of TNBC increased.
A number of studies have reported that miR-139 plays an important role in the tumor aggressiveness of breast cancer, and is poorly expressed in such tumors.Citation16,Citation23 In breast cancer, cells treated with a miR-139-5p mimic strongly synergized with radiation exposure, resulting in significantly increased oxidative stress, accumulation of unrepaired DNA damage, and induction of apoptosis.Citation4 The present results indicated that miR-139 may be a novel target for the treatment of breast cancer. Consistent with previous studies, our study found that miR-139 was down-regulated in TNBC cells. In addition, overexpression of miR-139 suppressed cell proliferation and promoted apoptosis of TNBC. In order to modify miR-139-inhibited tumor formation via targeting SOX8 in TNBC, cancer cells were transfected with miR-139 mimic and SOX8 pcDNA3.1, simultaneously. The results indicated that the effects of miR-139 were weakened by overexpression of SOX8.
Nonetheless, the mechanisms involved in miR-139 regulation of SOX8 in TNBC are not completely understood. For example, the downstream genes of SOX8 need to be explored in TNBC and effective targeted drugs need to be identified in future studies.
Conclusions
In summary, this study demonstrated that miR-139 inhibited the tumorigenicity of TNBC cells by targeting SOX8, providing a rationale for the clinical diagnosis and development of targeted therapeutic drugs in TNBC.
Abbreviations
TNBC, triple negative breast cancer; ER, estrogen receptor; PR, progesterone receptor; HER2, human epidermal growth factor 2; PVDF, polyvinylidene difluoride; SOX8, Sry-like high-mobility group box.
Statement of Ethics
All experimental protocols were performed according to the National Institutes of Health (NIH Pub. No. 85-23, revised 1996), and the experiments were approved by the Institutional Animal Care and Use Committee of Shandong University (No. 20190703-002).
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; and gave final approval of the version to be published. All authors agreed to be accountable for all aspects of the work.
Disclosure
The authors have no conflicts of interest to declare for this work.
References
- LaBarge MA, Samson S, Miyano M. Breast cancer beyond the age of mutation. Gerontology. 2015;62(4):434–442. doi:10.1159/00044103026539838
- Pogoda K, Niwinska A, Murawska M, Pienkowski T. Analysis of pattern, time and risk factors influencing recurrence in triple-negative breast cancer patients. Med Oncol. 2013;30(1):388. doi:10.1007/s12032-012-0388-423292831
- Cleator S, Heller W, Coombes RC. Triple-negative breast cancer: therapeutic options. Lancet Oncol. 2007;8(3):235–244. doi:10.1016/S1470-2045(07)70074-817329194
- Pajic M, Froio D, Daly S, et al. miR-139-5p modulates radiotherapy resistance in breast cancer by repressing multiple gene networks of DNA repair and ROS defense. Cancer Res. 2018;78(2):501–515. doi:10.1158/0008-5472.CAN-16-310529180477
- Liu D, Huo X, Gao L, Zhang J, Ni H, Cao L. NF-kappaB and Nrf2 pathways contribute to the protective effect of Licochalcone A on dextran sulphate sodium-induced ulcerative colitis in mice. Biomed Pharmacother. 2018;102:922–929. doi:10.1016/j.biopha.2018.03.13029710547
- Joerger M, Thurlimann B. Chemotherapy regimens in early breast cancer: major controversies and future outlook. Expert Rev Anticancer Ther. 2013;13:165–178. doi:10.1586/era.12.17223406558
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281. doi:10.1016/S0092-8674(04)00045-514744438
- Shukla GC, Singh J, Barik S. MicroRNAs: processing, maturation, target recognition and regulatory functions. Mol Cell Pharmacol. 2011;3(3):83–92.22468167
- Zhong S, Li W, Chen Z, Xu J, Zhao J. MiR-222 and miR-29a contribute to the drug-resistance of breast cancer cells. Gene. 2013;531(1):8–14. doi:10.1016/j.gene.2013.08.06223994196
- Le Quesne J, Caldas C. Micro-RNAs and breast cancer. Mol Oncol. 2010;4(3):230–241. doi:10.1016/j.molonc.2010.04.00920537965
- Huang LL, Huang LW, Wang L, Tong BD, Ding XS. Potential role of miR-139-5p in cancer diagnosis, prognosis and therapy (Review). Oncol Lett. 2017;14(2):1215–1222. doi:10.3892/ol.2017.635128789336
- Miyoshi J, Toden S, Yoshida K, et al. MiR-139-5p as a novel serum biomarker for recurrence and metastasis in colorectal cancer. Sci Rep. 2017;7:43393. doi:10.1038/srep4339328262692
- Watanabe K, Amano Y, Ishikawa R, et al. Histone methylation-mediated silencing of miR-139 enhances invasion of non-small-cell lung cancer. Cancer Med. 2015;4(10):1573–1582. doi:10.1002/cam4.50526256448
- Yan G, Li Y, Zhan L, et al. Decreased miR-124-3p promoted breast cancer proliferation and metastasis by targeting MGAT5. Am J Cancer Res. 2019;9(3):585–596.30949412
- Feng X, Luo Q, Wang H, Zhang H, Chen F. MicroRNA-22 suppresses cell proliferation, migration and invasion in oral squamous cell carcinoma by targeting NLRP3. J Cell Physiol. 2018;233(9):6705–6713. doi:10.1002/jcp.26331
- Dai H, Gallagher D, Schmitt S, et al. Role of miR-139 as a surrogate marker for tumor aggression in breast cancer. Hum Pathol. 2017;61:68–77. doi:10.1016/j.humpath.2016.11.00127864119
- Hua W, Sa KD, Zhang X, et al. MicroRNA-139 suppresses proliferation in luminal type breast cancer cells by targeting Topoisomerase II alpha. Biochem Biophys Res Commun. 2015;463(4):1077–1083. doi:10.1016/j.bbrc.2015.06.06126079880
- Gu SQ, Luo JH, Yao WX. The regulation of miR-139-5p on the biological characteristics of breast cancer cells by targeting COL11A1. Math Biosci Eng. 2019;17(2):1428–1441. doi:10.3934/mbe.202007332233587
- Tang H, Chen B, Liu P, et al. SOX8 acts as a prognostic factor and mediator to regulate the progression of triple-negative breast cancer. Carcinogenesis. 2019;40(10):1278–1287. doi:10.1093/carcin/bgz03430810729
- Chen B, Tang H, Chen X, et al. Transcriptomic analyses identify key differentially expressed genes and clinical outcomes between triple-negative and non-triple-negative breast cancer. Cancer Manag Res. 2018;11:179–190. doi:10.2147/CMAR.S18715130613165
- Xie SL, Fan S, Zhang SY, et al. SOX8 regulates cancer stem-like properties and cisplatin-induced EMT in tongue squamous cell carcinoma by acting on the Wnt/β-catenin pathway. Int J Cancer. 2018;142(6):1252–1265. doi:10.1002/ijc.3113429071717
- Chen S, Li H, Li X, et al. High SOX8 expression promotes tumor growth and predicts poor prognosis through GOLPH3 signaling in tongue squamous cell carcinoma. Cancer Med. 2020;9(12):4274–4289. doi:10.1002/cam4.304132307911
- Rask L, Balslev E, Søkilde R, et al. Differential expression of miR-139, miR-486 and miR-21 in breast cancer patients sub-classified according to lymph node status. Cell Oncol (Dordr). 2014;37(3):215–227. doi:10.1007/s13402-014-0176-625027758