
Abstract
Background
Increasing evidence indicates that circular RNAs (circRNAs) act as vital regulators in various cancers. Nevertheless, the effect of circCSNK1G1 on gastric cancer (GC) is still unknown.
Methods
The mRNA levels of circCSNK1G1, miR-758, and ZNF217 were measured by RT-qPCR. The protein levels of ZNF217 were evaluated by Western blotting. Cell migration, invasion, proliferation, and apoptosis were detected by Transwell, CCK-8, and flow cytometry assays. The association between miR-758 and circCSNK1G1/ZNF217 was confirmed by RIP and luciferase reporter assays. Xenograft assay was employed for in vivo experiment.
Results
In the current study, it was demonstrated that the expression levels of circCSNK1G1 and ZNF217 were upregulated in GC tissues and cells, while the level of miR-758 was declined. Furthermore, functional assays indicated that circCSNK1G1 depletion suppressed GC progression in vitro and in vivo. In addition, circCSNK1G1 directly interacted with miR-758, and the supplementation of miR-758 suppressed the development of GC, which was abolished following pcDNA3.1-circCSNK1G1 transfection. Then, we explored the downstream mechanism of miR-758 and found that miR-758 could target the 3′UTR of ZNF217 mRNA. The overexpression of miR-758 neutralized the ZNF217-mediated effects on facilitating the progression of GC. Finally, we revealed that circCSNK1G1 could upregulate ZNF217 expression by sponging miR-758 in GC cells.
Conclusion
Our study revealed that circCSNK1G1 accelerated GC progression via the miR-758/ZNF217 axis, suggesting that circCSNK1G1 might be a potential biomarker for GC diagnosis and treatment.
Keywords:
Introduction
Gastric cancer (GC) is the third most frequent malignancy in the world Citation1 and nearly a million new cases of GC are reported annually, with two-thirds of all cases in developing countries. Citation2,Citation3 Despite great progress has been achieved in the diagnosis and therapy of GC, the five-year survival rate remains unsatisfactory owing to the high metastasis and recurrence of GC. Citation4,Citation5 Therefore, the identification of novel possible biomarkers for GC diagnosis and treatment is critical.
Circular RNAs (circRNAs) are a category of RNA with a covalent closed loop structure. Citation6–8 Increasing research has indicated that circRNAs are involved in the development of various cancers through the regulation of cellular processes, such as cell growth, migration, apoptosis, and cell cycle. Citation9 Liu et al disclosed that the upregulation of circRNA-5692 attenuated hepatocellular carcinoma cell proliferation, migration, and invasion in vitro and inhibited tumor growth in vivo. Citation10 Zeng et al revealed that c-Myb-induced circHIPK3 facilitated cell growth and metastasis by regulating miR-7 expression in colorectal cancer. Citation11 Moreover, many circRNAs have been identified as important regulators in GC. To cite an instance, Yu et al identified down-regulated expression and vital diagnostic value of circ_0067582 in GC. Citation12 A report from Sun et al uncovered that circMAN2B2 facilitated GC cell proliferation and metastasis by acting as a sponge for miR-145. Citation13 It was also revealed that circ_0023642 contributed to GC development via interaction with miRNA-223. Citation14 Ding et al reported that circCSNK1G1 was one of the markedly upregulated circRNAs in GC. Citation15 However, the biological role of circCSNK1G1 in GC is still unclear.
MicroRNAs (miRNAs), another type of ncRNAs, frequently participate in the regulation of the tumorigenesis of multiple human cancers, including GC. Citation16 For example, the supplementation of miR-129 suppressed the tumorigenesis of GC by targeting COL1A1. Citation17 MiR-4317 impeded GC cell proliferation by inhibiting ZNF322. Citation18 miR-197 repressed GC cell viability and invasion through mediating MTDH. Citation19 The dysregulation of miR-758 has been reported to participate in the development of various human cancers, including bladder, Citation20 ovarian, Citation21 and cervical cancers. Citation22 Moreover, Guo et al identified that miR-758 played an antitumor role in the GC progression. Citation23 Nonetheless, the exact function and molecular mechanism of miR-758 in GC need to be further explained.
In this research, we uncovered a new mechanism that circCSNK1G1 modulated the miR-758/ZNF217 axis to facilitate GC progression. Our findings suggest that circCSNK1G1 may be a novel biomarker for GC diagnosis and treatment.
Materials and Methods
Clinical Samples
From the First People’s Hospital of Huzhou, 48 GC tissues and 48 corresponding normal tissues were collected. Besides, serum samples were also collected from 48 healthy volunteers as a healthy control grouThis study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the First People’s Hospital of Huzhou, and each participant provided the informed consent. None of the patients received preoperative treatment or had been diagnosed with other cancers. The specimens were quickly frozen in liquid nitrogen and stored at −80°C for further study.
Cell Line Culture
Human GC cell lines (AGS, HGC-27, and MKN45) and normal gastric epithelial cell line (GES-1) were purchased from ATCC. Cells were incubated in RPMI 1640 (HyClone, UT, USA) containing 10% FBS (Gibco, Rockville, MD, USA) maintained in a 37°C, 5% CO2 incubator.
Cell Transfection
siRNA targeting circCSNK1G1 (si-circCSNK1G1#1: 5ʹ- GGUACCGACGACUAGUCUUGU-3ʹ; si-circCSNK1G1#2: 5ʹ- UGGCCCUCGUAACCCAGCUGAU-3ʹ) with its control group (si-NC), miR-758 mimics (5′-UUUGUGACCUGGUCCACUAACC-3′) with its control (NC mimics, 5′-ACAUCUGCGUAAGAUUCGAGUCUA-3′), miR-758 inhibitor (5′-GGUUAGUGGACCAGGUCACAAA-3′) with its control (NC inhibitor, 5′-GCGUAACUAAUACAUCGGAUUCGU-3′) were obtained from GenePharma (Shanghai, China). circCSNK1G1 or ZNF217 overexpression plasmid (pcDNA3.1-circCSNK1G1 or pcDNA3.1-ZNF217) was established by inserting the full-length circCSNK1G1 or ZNF217 sequence into pcDNA3.1 vector. Lipofectamine 2000 (Invitrogen, CA, USA) was performed to transfect these oligonucleotides or vectors into GC cells.
RT-qPCR
Trizol reagent (Invitrogen) was used for the extraction of RNA. cDNA was then synthesized using PrimeScript RT Reagent kit (Takara, Otsu, Japan) qPCR was performed using the SYBRGreen PCR kit (Takara) and a 7500 Fast PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.). GAPDH or U6 was used as reference control, and the relative gene expression was calculated by the 2−ΔΔCq method. The primer sequences used were as follows: circCSNK1G1 forward, 5′-GCCATCACAACAGCAGCCT-3′ and reverse, 5′-AGGTCAAACAAGTCCTCCAAG-3′; miR-758 forward, 5′-ACACTCCAGCTGGGTTTGTGACCTGGTCCA-3′ and reverse, 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGGTTAGTG-3′; ZNF217 forward, 5ʹ-GAGAAGCGAATGGTGAAAGC-3ʹ, and reverse, 5ʹ-CAGCGCTCAAGTATGCAAAA-3ʹ; GAPDH forward, 5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′; and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′
Transwell Assay
The abilities of invasion and migration of GC cells were detected using transwell chambers (Corning, NY, USA). For detecting invasion ability, the transwell chambers were coated with Matrigel (BD Biosciences). The transfected cells (1x105 cells) were suspended in RPMI 1640 without serum and added to the upper chambers. Then, the lower chamber was filled with 600μL RPMI 1640 supplemented with 10% FBS. After 24 h, the invaded cells were stained with 0.5% crystal violet. The procedure for cell migration was the same as for cell invasion, except that the transwell chambers were not coated with Matrigel.
CCK-8 Assay
Transfected cells were seeded onto 96-well plates and then incubated for 0, 24, 48, and 72 h. The CCK-8 reagent was added to each well and then incubated for 4 h. The absorbance value of each well was detected at 490 nm.
Xenograft Assay in Nude Mice
5-week-old BALB/c nude mice bought from Shanghai Laboratory Animal Center (Shanghai, China) were used for the experiment. 1×107 AGS transfected with si-circCSNK1G1#1 or si-NC were subcutaneously injected into mice. Every 7 days, the tumor volume was monitored. 28 days after injection, mice were sacrificed and tumors were weighted. The animal experiment was performed according to the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and approved by the Ethics Committee of the First People’s Hospital of Huzhou.
Immunohistochemistry (IHC)
Tumor tissues collected from mice were processed by paraformaldehyde (4%) for fixation, and then the fixed tissues were dehydrated and embedded in paraffin. Subsequently, sections (4-mm-thick) were obtained via intersecting the paraffin-embedded tissues. Then, sections were deparaffinized and incubated at 4°C overnight with Ki67 antibody (Abcam). After incubation with a secondary antibody, sections were stained by hematoxylin, and photographed under a microscope.
RIP
RIP assay was applied using EZ-Magna RiP Kit (Millipore). Cells were lysed and incubated by anti-Ago2 antibody, with magnetic beads (Millipore, USA) or control antibody (Millipore, USA). The RT-qPCR was conducted after the immune precipitated RNA was purified.
Flow Cytometry
The transfected cells were digested with 0.25% trypsin, and then suspended in 100 μL binding buffer to produce 1×106 cells/mL suspension. Subsequently, the suspension was added with 10 μL AnnexinV-FITC and 10 μL PI for 10 min in darkness, and the apoptosis of cells was analyzed using FACScan flow cytometer (BD Biosciences).
Luciferase Reporter Assay
Wild-type circCSNK1G1 (circCSNK1G1-WT), mutant-type (circCSNK1G1-Mut), wild-type ZNF217 (ZNF217-WT), and mutant-type ZNF217 (ZNF217-Mut) were constructed by GenePharma. The reporter plasmids were co-transfection with miR-758 mimics or NC mimics via Lipofectamine 2000. The relative luciferase activity was detected by a dual-luciferase reporter assay system (Promega Corporation).
Western Blotting
Total protein was isolated from GC cells by using RIPA lysis buffer. The protein was separated with 10% SDS-PAGE and then moved to the PVDF membrane. After being blocked with 5% skimmed milk, the membrane was mixed with primary antibodies (anti-ZNF217: ab136678 and anti-GAPDH: ab181602; Abcam, USA) at 4°C all night. Then, the membrane was cultured with horseradish peroxidase-labeled secondary antibodies. Subsequently, the bands were evaluated with the enhanced chemiluminescence (ECL) Kit (Pierce, Thermo Fisher Scientific, IL, USA).
Statistical Analysis
The experiments were all conducted 3 times. Data were presented as mean ± standard deviation. SPSS 17.0 was performed for statistical analysis. Student’s t-test and ANOVA were utilized to measure the differences. Kaplan-Meier method was used to calculate the overall survival curve. P< 0.05 was considered as statistical significance.
Results
CircCSNK1G1 Was Upregulated in GC and Associated with the Poor Prognosis of GC Patients
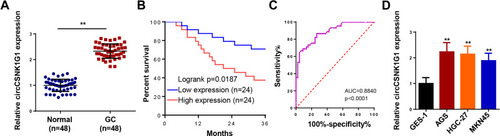
Firstly, the levels of circCSNK1G1 in GC tissues and adjacent normal tissues were detected by RT-qPCR and the data showed that circCSNK1G1 was considerably upregulated in GC tissues (). In the clinical sample, Kaplan–Meier survival analysis showed that the GC patients with high expression of circCSNK1G1 exhibited a lower survival rate than those with low expression of circCSNK1G1 (). Besides, the upregulation of circCSNK1G1 was related to tumor size, histological grades and TNM stage, but not with patients’ ages, gender or lymph node metastasis (). In addition, we also assessed the diagnostic value of serum circCSNK1G1, and the results showed that area under curve (AUC) was 0.8840, implying that circCSNK1G1 might be used as an indicator for GC diagnosis (). Moreover, we further observed that GC cell lines (AGS, HGC-27, and MKN45) established higher circCSNK1G1 expression than GES-1 (). Hence, we speculated that circCSNK1G1 might be an oncogene of GC.
Table 1 The Relationship Between circCSNK1G1 Expression Level and the Clinicopathologic Features of GC
Figure 1 CircCSNK1G1 was upregulated in GC and associated the prognosis of GC patients. (A) The level of circCSNK1G1 in GC tissues and paired normal tissues was measured using RT-qPCR. (B) Kaplan-Meier analysis reveals the association of high crcCSNK1G1 expression with poor overall survival in patients with GC. (C) ROC curve analysis was used to assess the diagnostic values of the circCSNK1G1. (D) The expression of circCSNK1G1 in GC cell lines and normal gastric epithelial cell line was detected using RT-qPCR. **P < 0.01.
Interference of circCSNK1G1 Inhibited the Progression of GC in vitro
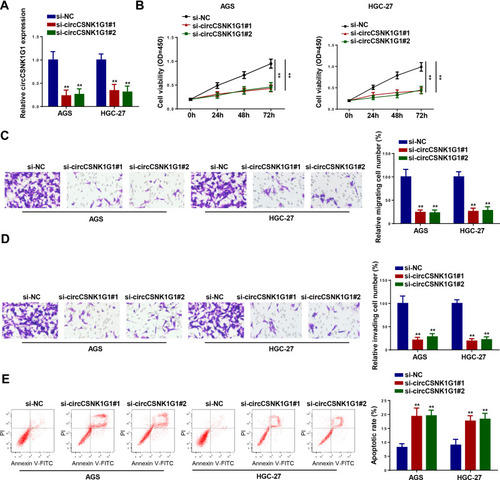
To determine the function of circCSNK1G1 in GC, AGS and HGC-27 cells were transfected with si-circCSNK1G1#1, si-circCSNK1G1#2, or si-NC, and a remarkable downregulation of circCSNK1G1 level in cells transfected with si-circCSNK1G1#1 and si-circCSNK1G1#2 was detected by RT-qPCR (). CCK-8 assay revealed that circCSNK1G1 depletion restrained the proliferation of GC cells (). Furthermore, transwell assays indicated that the interference of circCSNK1G1 attenuated the migration and invasion of GC cells (). In addition, flow cytometry assay indicated that circCSNK1G1 knockdown promoted the cell apoptosis of GC (). These results demonstrated that the knockdown of circCSNK1G1 could inhibit GC development.
Figure 2 Interference of CircCSNK1G1 inhibited the progression of GC. (A) The expression of circCSNK1G1 was detected by RT-qPCR in AGS and HGC-27 cells transfected with si-circCSNK1G1#1, si-circCSNK1G1#2, or si-NC. (B) The proliferation of AGS and HGC-27 cells was measured by CCK-8 assay. (C and D) Transwell was used to detect the migration and invasion of AGS and HGC-27 cells transfected with si-circCSNK1G1#1, si-circCSNK1G1#2, or si-NC. (E) Flow cytometry assay showed the apoptosis of AGS and HGC-27 cells transfected with si-circCSNK1G1#1, si-circCSNK1G1#2, and si-NC. **P < 0.01.
CircCSNK1G1 Knockdown Represses GC Tumorigenesis in vivo
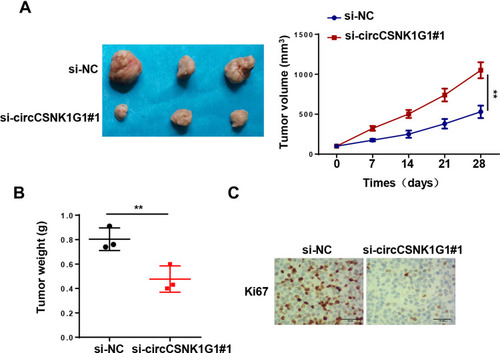
To further elaborate the biological role of circCSNK1G1 in GC in vivo, AGS cells transfected with si-NC or si-circCSNK1G1#1 were subcutaneously injected into nude mice. As presented in , circCSNK1 depletion significantly restrained tumor growth in mice. Moreover, IHC staining showed that circCSNK1 knockdown decreased Ki67 expression with comparison of the control group (). Altogether, these results implied that circCSNK1G1 knockdown suppressed GC tumorigenesis in vivo.
Figure 3 CircCSNK1G1 knockdown represses GC tumorigenesis in vivo. (A) AGS cells stably transfected with si-circCSNK1G1#1 or si-NC were subcutaneously injected into nude mice. Tumor growth curves indicated that circCSNK1G1 knockdown led to suppression on GC growth in mice. (B) The subcutaneous tumors were harvested and weighted. (C) IHC staining showed that Ki67 expression in AGS cells transfected with si-circCSNK1G1 and si-NC. **P < 0.01.
CircCSNK1G1 Was a Sponge of miR-758
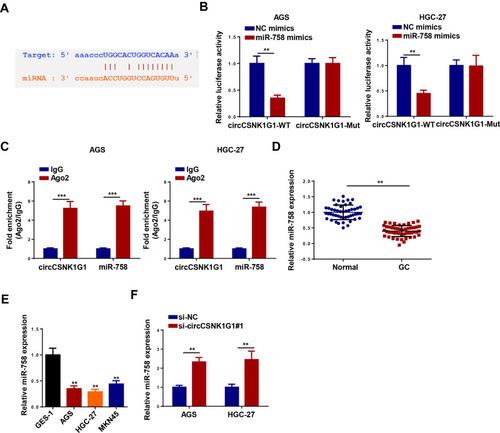
The downstream target genes were screened via the StarBase website to further explore the molecular mechanism of circCSNK1G1 in GC, and the results indicated that miR-758 might be a candidate downstream target of circCSNK1G1 (). Moreover, it was found that miR-758 mimics transfection remarkably diminished the luciferase activity of circCSNK1G1-WT reporter, but not the activity of circCSNK1G1-Mut in AGS and HGC-27 cells (). RIP assay showed that circCSNK1G1 and miR-758 were significantly enriched in the Ago2 group compared with the IgG group (). Furthermore, miR-758 expression was distinctly highly expressed in normal tissues and cells compared to GC tissues and cells (). Besides that, the deletion of circCSNK1G1 upregulated miR-758 expression (). These data showed that circCSNK1G1 could directly interact with miR-758.
Figure 4 CircCSNK1G1 was a sponge of miR-758. (A) Putative binding regions of circCSNK1G1 in miR-758 predicted with biological software. (B) Luciferase activity of wild-type or mutant circCSNK1G1 in AGS and HGC-27 cells following NC mimics, or miR-758 mimics transfection determined by luciferase reporter assay. (C) Correlations between circCSNK1G1 and miR-758 detected by RIP assay. (D) The level of miR-758 in GC tissues and paired normal tissues was measured using RT-qPCR. (E) miR-758 in GC cell lines and normal gastric epithelial cell line was detected using RT-qPCR. (F) The expression of miR-758 was detected by RT-qPCR in AGS and HGC-27 cells transfected with si-circCSNK1G1 and si-NC. **P < 0.01 and ***P < 0.001.
CircCSNK1G1 Overexpression Counteracted miR-758-Mediated Effects on the Progression of GC
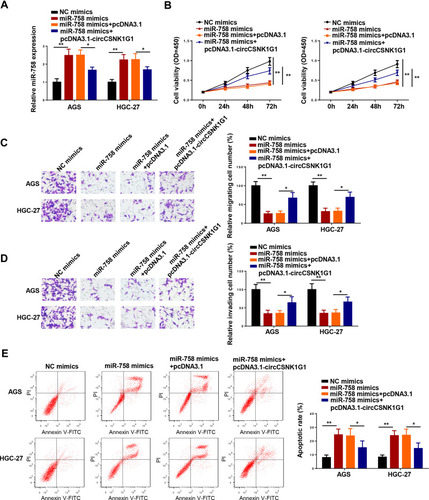
In order to deeply explore the interaction between circCSNK1G1 and miR-758 in GC, the level of miR-758 was assessed in AGS and HGC-27 cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-circCSNK1G1. RT-qPCR assay indicated that the addition of circCSNK1G1 counteracted the promotive effects of miR-758 mimics on miR-758 expression (). Moreover, the supplementation of miR-758 hindered the progression of GC, which was neutralized following pcDNA3.1-circCSNK1G1 transfection (–). The above data indicated that the opposite functions of circCSNK1G1 and miR-758 in GC progression.
Figure 5 CircCSNK1G1 overexpression counteracted miR-758-mediated effects on the progression of GC. (A) The expression of miR-758 was measured by RT-qPCR in AGS and HGC-27 cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-circCSNK1G1. (B) The proliferation of AGS and HGC-27 cells was measured by CCK-8 assay. (C and D) Transwell was used to detect the migration and invasion of AGS and HGC-27 cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-circCSNK1G1. (E) Flow cytometry assay showed the apoptosis of AGS and HGC-27 cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-circCSNK1G1. *P < 0.05 and **P < 0.01.
ZNF217 Was Directly Targeted by miR-758
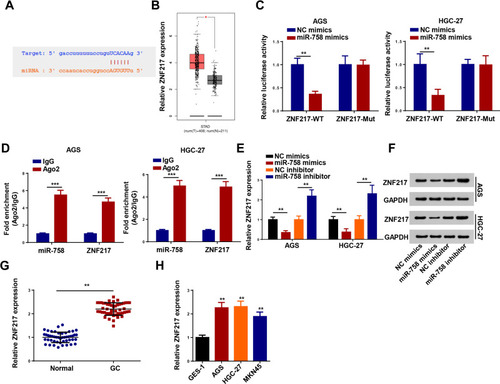
According to the prediction of TargetScan, ZNF217 might be a target of miR-758 (). Though GEPIA database, ZNF217 expression was higher expressed in stomach adenocarcinoma (STAD) tissues than that in normal tissues (). To validate the binding ability between miR-758 and ZNF217, luciferase reporter and RIP assays were conducted. The results revealed that miR-758 mimics resulted in an obvious reduction in relative luciferase activity of ZNF217-WT but not in ZNF217-Mut (). Meanwhile, miR-758 and ZNF217 were both enriched in the Ago2 group (). Moreover, RT-qPCR and Western blotting revealed that the upregulation of miR-758 decreased ZNF217 expression, and the downregulation of miR-758 increased ZNF217 expression (). After that, an increased ZNF217 expression was observed in GC tissues and cells (). To summarize, miR-758 negatively regulated ZNF217 level by directly targeting 3ʹ-UTR of ZNF217 mRNA.
Figure 6 ZNF217 was directly targeted by miR-758. (A) Putative binding regions of miR-758 in ZNF217 3ʹUTR. (B) GEPIA database showed ZNF217 expression in stomach adenocarcinoma (STAD) tissues compared with that in normal tissues. (C) Luciferase activity of wild-type or mutant ZNF217 in AGS and HGC-27 cells following NC mimics, or miR-758 mimics transfection determined by luciferase reporter assay. (D) Correlations between ZNF217 and miR-758 detected by RIP assay. (E and F) The expression of ZNF217 was measured by RT-qPCR and Western blotting in AGS and HGC-27 cells transfected with miR-758 mimics or miR-758 inhibitor. (G and H) The expression of ZNF217 in GC tissues and cells detected by RT-qPCR. **P < 0.01 and ***P < 0.001.
CircCSNK1G1 Regulated ZNF217 by Sponging miR-758 in GC Cells
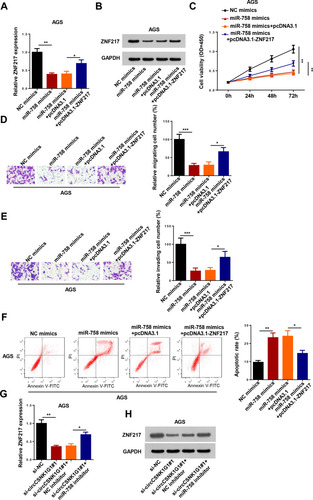
Subsequently, we examined whether ZNF217 was a downstream regulator of circCSNK1G1/miR-758 axis. RT-qPCR and Western blotting indicated that ZNF217 was notably downregulated by miR-758 mimics transfection, which could be abolished by pcDNA3.1-ZNF217 in AGS cells (). Moreover, the effect of miR-758 mimics on proliferation, migration, invasion, and apoptosis of AGS cells was abolished by upregulating ZNF217 expression (–). Furthermore, we investigated whether circCSNK1G1 could regulate ZNF217 expression via miR-758, and the results indicated that interference of circCSNK1G1 downregulated the expression of ZNF217, which was abolished following miR-758 inhibitor transfection (). To sum up, these results demonstrated that circCSNK1G1 regulated GC progression via miR-758/ZNF217 axis.
Figure 7 CircCSNK1G1 regulated ZNF217 by sponging miR-758 in GC cells. (A and B) The expression of ZNF217 was measured by RT-qPCR and Western blotting in AGS cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-ZNF217. (C–F) CCK-8, Transwell and flow cytometry assays were used to detect the proliferation, migration and invasion and apoptosis of AGS cells transfected with NC mimics, miR-758 mimics, miR-758 mimics+pcDNA3.1, and miR-758 mimics+pcDNA3.1-ZNF217. (G and H) The expression of ZNF217 was measured by RT-qPCR and Western blotting in AGS cells transfected with si-NC, si-circCSNK1G1#1, si-circCSNK1G1#1+NC inhibitor, and si-circCSNK1G1+miR-758 inhibitor. *P < 0.05, **P < 0.01 and ***P < 0.001.
Discussion
With the continuous development of next-generation sequencing technology, increasing circRNAs have been identified, as well as their function in human cancers, including GC. For instance, circ-HuR was downregulated in GC, and overexpression of circ-HuR inhibited the growth and metastasis of GC cells. Citation24 circRNA_001569 increased proliferation and decreased apoptosis of GC cells via inhibiting miR-145 expression. Citation25 CircCSNK1G1, a novel circRNA, was confirmed to be involved in the development of tumors. For example, Yao et al revealed that circCSNK1G1 facilitated hepatocellular carcinoma development by upregulating TRAF6 and MAPK11. Citation26 Huang et al reported that circCSNK1G1 accelerated colorectal cancer progression by elevating MYO6 level via interaction with miR-455-3p. Citation27 In this research, we focused on the role of circCSNK1G1 in GC. The results showed that circCSNK1G1 was upregulated in GC and deletion of circCSNK1G1 restrained GC progression in vitro and in vivo.
Previous researches have indicated that circRNAs could act as a competing endogenous RNA (ceRNA) of miRNA to exert its effect in cellular physiology. Citation28 For example, circCACTIN served as a ceRNA of miR-331 to regulate TGFBR1 level and accelerated GC development. Citation29 CircMAT2B facilitated glycolysis and growth of GC through increasing HIF-1α expression by restraining miR-515-5p. Citation30 CircLDLRAD3 promoted GC progression and could function as ceRNA to target NRP2 by sponging miR-224. Citation31 In our study, miR-758 was identified as a downstream gene of circCSNK1G1. miR-758 has been verified as a downstream regulator of circRNAs in the tumorigenesis of human cancers. For instance, circRNA RBM33 contributed to the occurrence and development of cervical cancer by sponging miR-758. Citation32 Hsa_circ_0002483 facilitates acute myeloid leukemia progression by downregulating miR-758 expression. Citation33 Here, we observed that supplementation of miR-758 restrained cell proliferation, migration and invasion, and promoted the apoptosis of GC cells, while circCSNK1G1 overexpression neutralized the effects on GC progression induced by miR-758.
ZNF217 is a potential oncogene located at chromosome 20q13.2, a region that is frequently involved in tumorigenesis of many cancers. Citation34 For example, the upregulation of ZNF217 predicted poor prognosis in breast cancer and promoted breast cancer progression. Citation35 LncRNA CTBP1-AS2 enhanced cervical cancer cell proliferation and migration via upregulation of ZNF217. Citation36 Moreover, Shida et al indicated high expression of ZNF217 contributed a worse relapse-free survival and overall survival of patients with GC. Citation37 In this study, we confirmed that ZNF217 was a target gene of miR-758. Functional assays indicated that the suppressive effect of miR-758 overexpression on GC progression was counteracted by ZNF217 overexpression. In addition, the absence of circCSNK1G1 resulted in a downregulation ZNF217, and miR-758 inhibition abolished the repressive effect of circCSNK1G1 inhibition on ZNF217 expression.
In summary, we demonstrated that circCSNK1G1 stimulated the progression of GC via regulating the miR-758/ZNF217 axis. These findings indicated that circCSNK1G1 might be a potential biomarker for GC diagnosis and therapy.
Disclosure
The authors report no conflicts of interest in this work.
References
- Breugom AJ , Swets M , Bosset J-F , et al. Adjuvant chemotherapy after preoperative (chemo)radiotherapy and surgery for patients with rectal cancer: a systematic review and meta-analysis of individual patient data. Lancet Oncol . 2015;16(2):200–207. doi:10.1016/S1470-2045(14)71199-4 25589192
- Zhao L , Jiang L , He L , et al. Identification of a novel cell cycle-related gene signature predicting survival in patients with gastric cancer. J Cell Physiol . 2019;234(5):6350–6360. doi:10.1002/jcp.27365 30238991
- Chen W , Zheng R , Baade PD , et al. Cancer statistics in China, 2015. CA Cancer J Clin . 2016;66(2):115–132. doi:10.3322/caac.21338 26808342
- Rahman R , Asombang AW , Ibdah JA . Characteristics of gastric cancer in Asia. World J Gastroenterol . 2014;20(16):4483–4490. doi:10.3748/wjg.v20.i16.4483 24782601
- Lasithiotakis K , Antoniou SA , Antoniou GA , et al. Gastrectomy for stage IV gastric cancer. a systematic review and meta-analysis. Anticancer Res . 2014;34(5):2079–2085.24778009
- Li Z , Ruan Y , Zhang H , et al. Tumor-suppressive circular RNAs: mechanisms underlying their suppression of tumor occurrence and use as therapeutic targets. Cancer Sci . 2019;110(12):3630–3638. doi:10.1111/cas.14211 31599076
- Guo JU , Agarwal V , Guo H , et al. Expanded identification and characterization of mammalian circular RNAs. Genome Biol . 2014;15(7):409. doi:10.1186/s13059-014-0409-z 25070500
- Barrett SP , Wang PL , Salzman J . Circular RNA biogenesis can proceed through an exon-containing lariat precursor. eLife . 2015;4:e07540. doi:10.7554/eLife.07540 26057830
- Patop IL , Kadener S . circRNAs in Cancer. Curr Opin Genet Dev . 2018;48:121–127. doi:10.1016/j.gde.2017.11.007 29245064
- Liu Z , Yu Y , Huang Z , et al. CircRNA-5692 inhibits the progression of hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP expression. Cell Death Dis . 2019;10(12):900. doi:10.1038/s41419-019-2089-9 31776329
- Zeng K , Chen X , Xu M , et al. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis . 2018;9(4):417. doi:10.1038/s41419-018-0454-8 29549306
- Yu X , Ding H , Yang L , et al. Reduced expression of circRNA hsa_circ_0067582 in human gastric cancer and its potential diagnostic values. J Clin Lab Anal . 2020;34(3):e23080. doi:10.1002/jcla.23080 31721300
- Sun B , Sun H , Wang Q , et al. Circular RNA circMAN2B2 promotes growth and migration of gastric cancer cells by down-regulation of miR-145. J Clin Lab Anal . 2020;34(6):e23215. doi:10.1002/jcla.23215 32020674
- Zhang Y , Xia L , Wu J , et al. Hsa_circ_0023642 promotes proliferation, invasion, and migration of gastric cancer by sponging microRNA-223. J Clin Lab Anal . 2020;34(10):e23428. doi:10.1002/jcla.23428 32562304
- Ding HX , Xu Q , Wang B-G , et al. MetaDE-based analysis of circRNA expression profiles involved in gastric cancer. Dig Dis Sci . 2020;65(10):2884–2895. doi:10.1007/s10620-019-06014-6 31894486
- You Y , Que K , Zhou Y , et al. MicroRNA-766-3p inhibits tumour progression by targeting Wnt3a in hepatocellular carcinoma. Mol Cells . 2018;41(9):830–841. doi:10.14348/molcells.2018.0181 30145863
- Wang Q , Yu J . MiR-129-5p suppresses gastric cancer cell invasion and proliferation by inhibiting COL1A1. Biochem Cell Biol . 2018;96(1):19–25. doi:10.1139/bcb-2016-0254 28482162
- Hu X , Zhang M , Miao J , et al. miRNA-4317 suppresses human gastric cancer cell proliferation by targeting ZNF322. Cell Biol Int . 2018;42(8):923–930. doi:10.1002/cbin.10870 28880489
- Liao Z , Li Y , Zhou Y , et al. MicroRNA-197 inhibits gastric cancer progression by directly targeting metadherin. Mol Med Rep . 2018;17(1):602–611. doi:10.3892/mmr.2017.7908 29115517
- Wu X , Chen B , Shi H , et al. miR-758-3p suppresses human bladder cancer cell proliferation, migration and invasion by targeting NOTCH2. Exp Ther Med . 2019;17(5):4273–4278. doi:10.3892/etm.2019.7400 30988799
- Hu X , Li Y , Kong D , et al. Long noncoding RNA CASC9 promotes LIN7A expression via miR-758-3p to facilitate the malignancy of ovarian cancer. J Cell Physiol . 2019;234(7):10800–10808. doi:10.1002/jcp.27903 30537154
- Song T , Hou X , Lin B . MicroRNA-758 inhibits cervical cancer cell proliferation and metastasis by targeting HMGB3 through the WNT/beta-catenin signaling pathway. Oncol Lett . 2019;18(2):1786–1792. doi:10.3892/ol.2019.10470 31423246
- Guo J , Zhang Z , Pan L , et al. Identification of miR-758-3p as potential modulator of CBX5 expression in gastric cancer. Technol Cancer Res Treat . 2018;17:1533033818816061. doi:10.1177/1533033818816061 30486755
- Yang F , Hu A , Li D , et al. Circ-HuR suppresses HuR expression and gastric cancer progression by inhibiting CNBP transactivation. Mol Cancer . 2019;18(1):158. doi:10.1186/s12943-019-1094-z 31718709
- Shen F , Liu P , Xu Z , et al. CircRNA_001569 promotes cell proliferation through absorbing miR-145 in gastric cancer. J Biochem . 2019;165(1):27–36. doi:10.1093/jb/mvy079 30304349
- Yao Z , Xu R , Yuan L , et al. Circ_0001955 facilitates hepatocellular carcinoma (HCC) tumorigenesis by sponging miR-516a-5p to release TRAF6 and MAPK11. Cell Death Dis . 2019;10(12):945. doi:10.1038/s41419-019-2176-y 31822654
- Huang X , Shen X , Peng L , et al. CircCSNK1G1 contributes to the development of colorectal cancer by increasing the expression of MYO6 via competitively targeting miR-455-3p. Cancer Manag Res . 2020;12:9563–9575. doi:10.2147/CMAR.S262007 33061642
- Qi X , Zhang D-H , Wu N , et al. ceRNA in cancer: possible functions and clinical implications. J Med Genet . 2015;52(10):710–718. doi:10.1136/jmedgenet-2015-103334 26358722
- Zhang L , Song X , Chen X , et al. Circular RNA circCACTIN promotes gastric cancer progression by sponging miR-331-3p and regulating TGFBR1 expression. Int J Biol Sci . 2019;15(5):1091–1103. doi:10.7150/ijbs.31533 31182928
- Liu J , Liu H , Zeng Q , et al. Circular RNA circ-MAT2B facilitates glycolysis and growth of gastric cancer through regulating the miR-515-5p/HIF-1alpha axis. Cancer Cell Int . 2020;20:171. doi:10.1186/s12935-020-01256-1 32467667
- Wang Y , Yin H , Chen X . Circ-LDLRAD3 enhances cell growth, migration, and invasion and inhibits apoptosis by regulating MiR-224-5p/NRP2 axis in gastric cancer. Dig Dis Sci . 2021. doi:10.1007/s10620-020-06733-1
- Ding Y , Yuan X , Gu W . Circular RNA RBM33 contributes to cervical cancer progression via modulation of the miR-758-3p/PUM2 axis. J Mol Histol . 2021;52:173–185. doi:10.1007/s10735-020-09933-1 33398465
- Xiao Y , Ming X , Wu J . Hsa_circ_0002483 regulates miR-758-3p/MYC axis to promote acute myeloid leukemia progression. Hematol Oncol . 2020;39:243–253. doi:10.1002/hon.2829 33283885
- Kallioniemi A , Kallioniemi OP , Piper J , et al. Detection and mapping of amplified DNA sequences in breast cancer by comparative genomic hybridization. Proc Natl Acad Sci U S A . 1994;91(6):2156–2160. doi:10.1073/pnas.91.6.2156 8134364
- Vendrell JA , Thollet A , Nguyen NT , et al. ZNF217 is a marker of poor prognosis in breast cancer that drives epithelial-mesenchymal transition and invasion. Cancer Res . 2012;72(14):3593–3606. doi:10.1158/0008-5472.CAN-11-3095 22593193
- Yang S , Shi F , Du Y , et al. Long non-coding RNA CTBP1-AS2 enhances cervical cancer progression via up-regulation of ZNF217 through sponging miR-3163. Cancer Cell Int . 2020;20:343. doi:10.1186/s12935-020-01430-5 32742190
- Shida A , Fujioka S , Kurihara H , et al. Prognostic significance of ZNF217 expression in gastric carcinoma. Anticancer Res . 2014;34(9):4813–4817.25202062