
Abstract
Antibodies (Abs) containing two different antigen-binding sites in one molecule are called bispecific. Bispecific Abs (BsAbs) were first described in the 1960s, the first monoclonal BsAbs were generated in the 1980s by hybridoma technology, and the first article describing the therapeutic use of BsAbs was published in 1992, but the number of papers devoted to BsAbs has increased significantly in the last 10 years. Particular interest in BsAbs is due to their therapeutic use. In the last decade, two BsAbs – catumaxomab in 2009 and blinatumomab in 2014, were approved for therapeutic use. Papers published in recent years have been devoted to various methods of BsAb generation by genetic engineering and chemical conjugation, and describe preclinical and clinical trials of these drugs in a variety of diseases. This review considers diverse BsAb-production methods, describes features of therapeutic BsAbs approved for medical use, and summarizes the prospects of practical application of promising new BsAbs.
Introduction
Immunoglobulins (antibodies [Abs]) are major protein components of the adaptive immune system, directed against foreign compounds and infectious agents. The IgG molecule consists of two light and two heavy chains connected by disulfide bonds; IgG is a monomer with a molecular weight of 146–160 kDa. Antigen-binding centers of Abs are formed by hypervariable regions of heavy and light chains. In the classical point of view, the Ab molecule contains two identical antigen-binding sites (two HL fragments) and is monospecific and bivalent. Immunoglobulins are expressed as receptors of the cell membrane of B lymphocytes, as well as in the form of soluble molecules secreted by plasma cells. Soluble Abs can bind virtually any natural and artificial molecules (antigens) with high affinity and specificity. The ability of Abs to recognize and bind a broad spectrum of antigens is ensured by their extraordinary diversity, reaching 108–1010 different variants of antigen-binding centers. Bispecific immunoglobulins contain two different antigen-binding sites. In the last 10 years, the number of articles devoted to bispecific Abs (BsAbs) has been steadily increasing. The particular interest in these molecules is due to their potential for therapeutic use. In 2014, the journal Nature Reviews Drug Discovery called BsAbs “next-generation antibodies.”
Monoclonal Abs (mAbs) are secreted by identical immune cells, clones of a single parent cell. mAbs are not just monospecific bivalent molecules, but in contrast to polyclonal Abs, bind the same epitope (antigen fragment recognized by Abs). In this regard, mAbs are widely used for the treatment of cancer: Avastin (bevacizumab, anti-VEGF), Herceptin (trastuzumab, anti-HER2-receptor antagonist), and rituximab (anti-CD20) have been on the pharmaceutical market for more than 15 years. However, these and other drugs based on mAbs are not able to cure some cases of cancer in monotherapy, particularly due to T lymphocytes not taking an active part in the destruction of tumors, while mAb molecules only prevent the binding of growth factors to the receptors. mAbs blocking the inhibitory signals that protect tumors from immune cells show excellent results in the treatment of particular types of tumors. Nevertheless, high expectations are focused on Abs binding two or more antigens (BsAbs), as well as conjugated to agents for chemo- and radiotherapy.Citation1
BsAbs have been developed in which one antigen-binding site is directed against the CD3 receptor (activates cytotoxic T lymphocytes) and the other against specific antigens of tumor cells (CD19, CD20, CD33, CD123, HER2, epithelial cell adhesion molecule [EpCAM], BCMA, CEA, and others).Citation2 The convergence of cytotoxic T lymphocytes and tumor cells due to BsAb binding activates cytotoxic T cells and promotes the destruction of tumor cells. In addition to a wide range BsAbs directed against tumors, several bispecific molecules for the treatment of other diseases have been developed. The BsAb for the treatment of osteoporosis blocks the factors of Wnt signal-transduction pathway (sclerostin and Dkk1); it enhances the formation of osteoblasts and growth of bone tissue.Citation3 ACE910 binds blood-coagulation factors IX and X and is designed to reduce bleeding rate in hemophilia A. The convergence of coagulation factors enhances the coagulation cascade.Citation4 A BsAb against the transferrin receptor (provides passage through the blood–brain barrier) and protease BACE1 (accumulates amyloid peptides) is a candidate for an anti-Alzheimer’s disease drug.Citation5 BsAbs that are focused on autoimmune diseases usually bind cytokines: TNF, IL1, IL4, IL14, IL17, IL23, and others.Citation2,Citation6 It has been shown that simultaneous use of two mAbs against cytokines in autoimmune diseases has severe side effects without superior efficiency. In this regard, BsAbs against autoimmune diseases usually combine two anticytokine antigen-binding sites and provide higher therapeutic potential than a mixture of two mAbs.Citation7,Citation8 In particular, the most therapeutically important cytokines in psoriasis are IL17, IL23, IL6, and TNF.Citation9 ABT122 against TNFα and IL17A has clinical effects in rheumatoid arthritis and psoriatic arthritis.Citation10 In contrast, Phase I/II clinical trials of COVA322 (same specificity as ABT122) in psoriasisCitation11 were preliminary terminated, due to safety concerns.Citation8 Antigen-binding sites of ABT981 are directed against IL1α and IL1β, inflammatory cytokines found in the cartilage and synovial fluid of patients with osteoarthritis.Citation12
BsAbs have several significant advantages over monospecific Abs. BsAbs direct specific effectors of the immune system to target tumor cells, enhancing their cytotoxicity. BsAbs can provide higher binding specificity, since in contrast to monospecific Abs, they interact with two different surface antigens. The use of BsAbs compared to combination therapy with two monospecific drugs makes it possible to optimize expenses by reducing the cost of development and clinical trials. Since one disease modulator may play an essential role in several independent pathways and coexpression of different receptors has been found in many tumors, targeting of two different growth-promoting receptors on a single tumor cell may increase the antiproliferative effect and help to avoid the development of resistance.Citation13,Citation14
The sales of more than 50 mAbs presented on the pharmaceutical market have reached more than US$60 billion per year. According to a 2014 estimation, the market of therapeutic BsAbs will grow up to $5.8 billion per year by 2024.Citation15 Therapeutic BsAbs approved for medical use are directed for the treatment of “liquid” tumors: blood cancer (leukemia and lymphoma). The specialty of “liquid tumors” is that palpable tumors that can be mechanically probed do not form in the body, unlike solid tumors (breast, uterus, rectum). Leukemia and lymphoma never develop symptoms typical of other oncological pathologies, since malignant leukocytes proliferate in the bone marrow and come into the bloodstream as individual cells, making them available for BsAb therapy. There are other diseases for which BsAbs are being developed: autoimmune (arthritis, asthma, diabetes), infectious (pneumonia), hemophilia, and Alzheimer’s disease.Citation6
Therapeutic bispecific antibodies
To date, two BsAbs have been approved for use in the US (blinatumomab; Amgen) and Europe (catumaxomab; Trion Pharma). More than 60 drugs are in preclinical and 30 in clinical trials, and two-thirds of them are focused on cancer treatment.Citation16 Two therapeutic BsAbs that are on the market and some of the drugs in clinical and preclinical trials bring T lymphocytes (or natural killers) closer to cells expressing specific antigens on the surface, or simultaneously bind two antigens on the surface of a target cell.
Therapy with BsAbs constructed on whole IgG molecules shows active immunization against specific tumors, which in the future can lead to increased use of this format to provide long-lasting antitumor immunity in the organism. In the case of catumaxomab, the induction of a prolonged immunoresponse after treatment is due to the affinity of fragment crystallizable region (Fc) for dendritic cells presenting the antigen.Citation17 In the case of BsAbs generated by chemical hetero-conjugation (when Fc of two IgG molecules are covalently attached), immunization occurs due to the formation of an immunoactivating environment that attracts and activates dendritic cells.Citation18
Blinatumomab
The drug blinatumomab (Amgen) is the first representative of bispecific T-cell engagers (BiTEs) authorized for use in the US. The efficacy of blinatumomab as a therapeutic drug against B-cell tumors was first shown in 2008 in 38 patients with refractory non-Hodgkin’s lymphoma,Citation19 and the results of other preclinical and clinical studies have been published in a number of works.Citation20–Citation23 At the end of 2014, the US Food and Drug Administration approved the treatment with blinatumomab of acute lymphoblastic leukemia without the Philadelphia chromosome as a second-line drug.Citation24 In the EU, the drug was registered in 2015. Therapy with blinatumomab leads to the depletion of B lymphocytes and precursors in peripheral blood, which is gradually restored after the end of treatment.Citation19
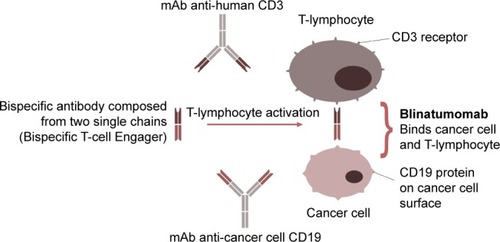
The bispecific blinatumomab molecule has been developed using diabody technology: the first antigen-binding site is directed against a CD19 protein on the surface of B lymphocytes, the second against the CD3 receptor on the surface of cytotoxic T lymphocytes (). The single-stranded structure of blinatumomab allows easy protein expression in monomeric form in significant amounts and provides broad therapeutic potential for use in lymphoma and leukemia.Citation25 Unfortunately, this feature is the reason continuous intravenous administration of the drug is required. The blinatumomab molecule directs primary CD3+ T cells against CD19+ lymphoma cells, and provides cytotoxicity at very low concentrations (~10–100 pg/mL).Citation26 The direction of cytotoxic T lymphocytes to tumor B cells, bypassing T-cell receptors and major histocompatibility complex, is a significant advantage of the drug.Citation27 Blinatumomab increases the secretion of anti-inflammatory cytokines (IL2, IL4, IL6, IL10, IFNγ, TNFα).Citation26 Data from animal models also confirm the high efficacy of blinatumomab against tumor cells in leukemia and lymphoma at very low concentrations.Citation26,Citation28
Figure 1 Mechanism of blinatumomab therapeutic action: recruitment of T cells to tumors through binding of tumor-cell-surface antigens to immune cells.
Blinatumomab therapy in adult patients with recurrent acute lymphoblastic leukemia leads to entirely positive results in 72%, achievement of minimal residual disease (tumor cells remaining in the organism after remission) in 88%, and average life expectancy after therapy of 9 months.Citation21 Therapy for non-Hodgkin’s lymphoma with blinatumomab also shows good efficacy: in monotherapy clinical trials, blinatumomab significantly exceeded the effect of mAb therapy at much-lower final blood concentrations.Citation22 Treatment of non-Hodgkin’s lymphoma patients with blinatumomab results in minimal residual disease, even after induction and consolidation,Citation20 but clinical trials are still under way. In refractory and recurrent acute lymphoblastic leukemia in cases of CD19 absence on the lymphocyte surface and extramedullary hematopoiesis (formation of lymphocytes outside the bone marrow), therapy with blinatumomab is ineffective.Citation29
After initiation of blinatumomab administration, the number of B lymphocytes decreases to under one cell/μL for 2 days and remains virtually undetectable until the end of therapy. On the contrary, the number of T lymphocytes lowers in all patients to a minimum level for 1 day and is then restored to normal within a few days. Moreover, within 2–3 weeks, the number of T cells doubles in most patients, dominated by an expansion of memory T cells expressing CD45RA. This can be explained by differences in signaling pathways used by memory T cells and naïve T cells.Citation30 Rapid removal of blinatumomab from the bloodstream due to low molecular weight and the necessity of regular intravenous administration is partially resolved in newer therapeutics of tetravalent Abs – AFM11 and AFM13, developed by AffimedCitation31,Citation32 – which can be administered once or twice a week. T lymphocytes with chimeric CD19 receptors (CAR-T) provide complete remission in 90% of patients with refractory acute lymphoblastic leukemia, and in vivo such cells can proliferate up to 1,000 times.Citation33 However, in the case of CAR-T therapy, the incidence of lymphocyte-release syndrome is much higher (up to 27%) than in the case of blinatumomab (up to 2%).Citation26 Abs against other antigens (eg, CD79B) developed using BiTE technology directed to the treatment of myeloid leukemia and lymphoma are currently undergoing clinical trials.Citation34
Catumaxomab
Catumaxomab (Removab, Trion) was the first bispecific trifunctional drug approved in 2009 by the European Medicines Agency for the treatment of malignant ascites.Citation35–Citation37 Catumaxomab redirects T cells to tumor cells, expressing EpCAM, ascites secondary to epithelial forms of cancer, especially gastric cancer. The results of clinical and preclinical studies of catumaxomab have been described in detail in many reviews.Citation37–Citation42 Catumaxomab is produced using the “quadroma” technology: HL fragments of mouse mAbs against CD3 (IgG2a) and rat mAbs (IgG2b) against EpCAM secreted by the corresponding hybridomas are combined in one bispecific molecule, which also binds the Fc-receptor.Citation43 The Fc of catumaxomab preferentially binds to FcγRI, FcγRIIa, and FcγRIII activation receptors, but not to the FcγRIIb inhibitory receptor. This determines the recruitment and activation of macrophages, NK cells, and dendritic cells (FcγR+), leading to a complex immunoreaction.Citation40 The use of HL fragments obtained from different host organisms reduces the possibility of BsAb formation with mismatched light chains, since light chains of rat Abs predominantly interact with rat heavy chains, and vice versa; light chains of mouse Abs are preferably associated with heavy mouse chains.Citation43
The application of catumaxomab in clinical trials and therapy proves to be successful, since there is no barrier for T lymphocytes or catumaxomab molecules to penetrating ascites tumors. Ascites tumors are represented by separate cells that float separately from one another in liquid, which unites them with lymphoma and leukemia, two other liquid tumors, against which is directed blinatumomab. As in the case of blinatumomab, the antitumor effect of catumaxomab is due to the colocalization of a T-lymphocyte, of a tumor cell expressing EpCAM, and of the cell with the Fc receptor on the surface (macrophage, dendritic cell, natural killer) (). Therefore, catumaxomab enhances activation of the patient’s immune system against the tumor. Binding of catumaxomab to Fcγ receptors of antigen-presenting cells turns inflammatory monocytes to express costimulatory molecules, which are necessary for T-cell activation.Citation44,Citation45 During preclinical studies, it was shown that catumaxomab could provide immunoresponse due to B- and T-memory cells activated by the Fc receptor.Citation46 In a Phase IIIB clinical trial, intraperitoneal infusion of catumaxomab favored accumulation of and activated macrophages, NK cells, and CD4+ and CD8+ T-cells in ascites of peritoneal cavities.Citation47,Citation48
Figure 2 Mechanism of action: catumaxomab is a trifunctional antibody that accelerates the recognition and destruction of tumor cells by different immune cells.
Notes: Catumaxomab binds the EpCAM on the surface of a cancer cell, CD3 on T lymphocytes, and Fc on Fcγ-receptor positive accessory cell. Immunoeffector cells interact with each other, leading to the elimination of tumor cells by the mechanisms of T-cell cytotoxicity, cytokine cytotoxicity, phagocytosis, or antibody-dependent cellular toxicity. Adapted by permission from Macmillan Publishers Ltd: Nat Biotechnol. Walsh G. Biopharmaceutical benchmarks 2010. 2010;28(9):917–924. Copyright 2010 Nature Publishing Group. Available from: https://www.nature.com/nbt/.Citation48
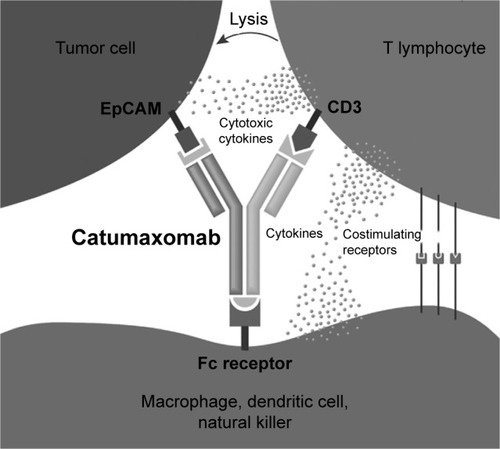
The interaction of patient immune cells with tumor cells leads to a complicated reaction, resulting in the elimination of tumor cells. Studies have shown several mechanisms of cytotoxicity: lymphocyte-mediated lysis, cytokine action (IL1β, IL2, IL6, IL12, CCL18 chemokine), phagocytosis, and Ab-dependent cellular toxicity. Compared to BsAbs, individual mouse and rat mAbs (anti-CD3 and anti-EpCAM) show significantly lower antitumor potential.Citation37 Catumaxomab has high therapeutic potential with acceptable safety: intraperitoneal administration of low doses (10–100 mg) of the drug are carried out four to five times with an interval of 10–14 days. Catumaxomab is approved for the treatment of malignant ascites and is currently undergoing clinical trials in ovarian, stomach, and epithelial cancer. At the same time, a similar therapeutic effect is observed in some other tumors, eg, in ovarian carcinoma, catumaxomab reduces the formation of ascitic fluid.Citation49 Interestingly, one of the catumaxomab-therapy side effects is the formation of Abs against mouse and rat Abs, and notably immunoresponse against mouse Abs correlates with positive response to treatment.Citation50
Other BsAbs undergoing preclinical and clinical trials
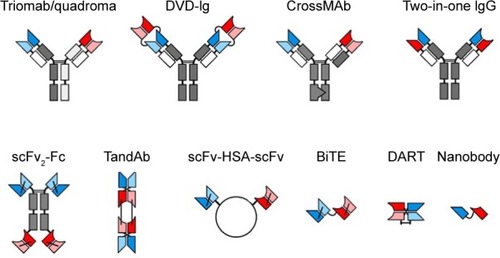
Most of the antitumor BsAbs currently undergoing clinical and preclinical trials, such as blinatumomab and catumaxomab, contain one anti-CD3 antigen-binding site, attracting the T lymphocyte to the tumor cell. The second antigen-binding site can be directed against CD19, CD20, CD33, CD123, HER1, HER2, CEA, disialoganglioside GD2, PSMA, gpA33, and other proteins (). Also, many BsAbs with other combinations of antigen-binding sites (usually anticytokines) are in trials: HER2 + HER3, IL1α + IL1β, IL13 + IL17, IL17A/IL17F, and CD30 + CD16A. Data on BsAbs undergoing clinical trials have been published in many reviews and are combined in ; the list of terminated clinical trials is combined in . The structures of most common BsAbs formats are presented in .
Table 1 Clinical trials of bispecific antibodies
Table 2 Terminated clinical trials of therapeutic bispecific antibodies
Figure 3 Architecture of common BsAbs formats.
Abbreviations: scFv, single-chain variable fragment; DART, dual-affinity retargeting; BsAbs, bispecific antibodies.
Adverse effects of BsAb treatment
Like other methods of therapy for severe diseases, therapeutic BsAbs cause different side effects, the most common of which are nausea, vomiting, abdominal pain, fatigue, leukopenia, neutropenia, and thrombopenia. In many patients, Abs against therapeutic BsAbs appear in the blood during treatment. Most adverse events occur during the beginning of therapy, and in most cases side effects normalize under continued treatment. The majority of data on therapeutic BsAb adverse effects are available on blinatumomab and catumaxomab, since these drugs have undergone numerous clinical trials. A common side effect of blinatumomab and catumaxomab therapy is “cytokine storm”, elevation of cytokine levels. Cytokine release-related symptoms are general side effects of many therapeutic mAbs, and occur due to specific mechanisms of action: use of cytotoxic T cells as effectors. Minimizing cytokine-release syndrome is possible with a low initial dose of the drug in combination with subsequent high doses,Citation54 as well as corticosteroid (dexamethasone)Citation40 and antihistamineCitation55 premedication.
The most common side effects during blinatumomab treatment are hepatotoxicity, leukopenia, lymphopenia, thrombocytopenia, CRP increase, chills, fever, pyrexia, nausea, and vomiting.Citation44 There are also noted neurological and psychiatric side effects, which are completely reversible after the end of therapy.Citation56 In catumaxomab treatment, the most frequent adverse events are cytokine release-related symptoms, which may have some predictive value for treatment efficacy. Some disorders of liver parameters and white blood cells are often observed, but usually these changes are reversible.Citation40 Adverse events in cases of Mus110 (BiTEs BsAb anti-EpCAM and anti-CD3) are in general due to activation of T cells. During treatment with MEDI565 (AMG211, MT111: BiTEs BsAb anti-CEA and anti-CD3), the most common side effects are nausea, vomiting, abdominal pain, and fatigue. Significant increases in antidrug Abs are detected in the blood of half the patients.Citation57 The main adverse event during TF2 (anti-CEA Fabs and anti-histamine–succinyl–glycine Fab) therapy is bone-marrow toxicity.Citation57
History and methods of bispecific antibody generation
The first work on BsAb generation was published in 1961 and described the production of chimeric BsAbs containing simultaneously two different antigen-binding sites from a mixture of two monospecific Abs. Such BsAbs simultaneously precipitated both antigens specific to the original Abs.Citation58 In 1983–2007, it was shown that IgG4 molecules stochastically exchanged HL fragments; this posttranslational modification results in a generation of chimeric bispecific molecules.Citation59,Citation60 In 2012–2016, it was shown that the bispecific IgG molecules of all four subclasses are present in blood,Citation61 placenta,Citation62 and milk (IgG and sIgA)Citation63,Citation64 of healthy donors.
The explosive growth of publications on BsAb generation happened in the 1980s and 1990s, when preparations of mAbs obtained by hybridoma technology became available. In 1983, hybrid hybridomas, called quadromas, expressing two types of light and heavy chains simultaneously and producing BsAbs were described.Citation65 In 1985, bispecific Fab2 was obtained by chemical cross-linking.Citation66 In 1987, Fab molecules were first cross-linked with thioesters.Citation67
In 1984, mAbs against the T-lymphocyte receptor and leukemia cell-line antigen were cross-linked with a heterobifunctional linker and it was shown that generated BsAbs attract T-lymphocytes and stimulate T-cell mediated cytotoxicity.Citation68 In 1985, a bispecific Fab2 was constructed from HL fragments against human CD3 receptors and HL fragments against H2k mouse protein; the resulting construct directed cytotoxic human T-lymphocytes to murine tumor cells with surface KK antigen and destroyed them by an Ab-dependent cytotoxicity mechanism.Citation69 In 1987, it was shown that BsAbs containing HL fragments, anti-T-cell antigen receptors, and antihemagglutinin of influenza virus demonstrate aspecific cytotoxic T-lymphocyte-mediated lysis of infected cells.Citation70 A detailed review of BsAb development history has been published.Citation71
The first papers on perspectives on BsAbs in clinical use were published in the 1990s. In 1992, BsAbs were shown to direct monocytes (carriers of FcγRI) against CD15+ breast carcinoma cells PM81, acute myeloid leukemia, and small-cell lung and intestinal carcinoma.Citation72 In 1995, MDX210 BsAbs against FcγR1 on the surface of monocytes and macrophages and against HER2+ cells were described; the construct was clinically active in breast and ovarian tumors.Citation73,Citation74 Clinical studies of 2B1 specific against tumor cells expressing c-erbB-2 and cells bearing FcyRIII showed the possibility of BsAb-mediated targeted lysis of HER2+ cells (breast, intestine, lung, kidney, prostate cancer) by natural killers and phagocytes.Citation75
The first generation of BsAbs was obtained by chemical cross-linking or from hybridomas. Now, most BsAbs are generated by three methods: by chemical conjugation with cross-linkers, by somatic fusion of two hybridoma lines (quadroma), and by genetic (protein/cell) engineering. Large pharmacological and biotechnological companies are developing new techniques for therapeutic BsAb generation, and more than 60 different technological platforms have been developed in the last 15 years.Citation16 Depending on the production method and structure, BsAbs vary in the number of antigen-binding sites, geometry, half-life in the blood serum, and effector functions. According to the mechanism of action, most modern BsAbs undergoing preclinical and clinical studies are classified into four formats: BiTEs, dual-affinity retargeting Abs, homodimeric “knob-in-hole” Abs, and trifunctional BsAbs.
Chemical conjugation and covalent attachment of fragments
BsAbs can be generated by the attachment of light or heavy chains, single-domain Abs, single-chain variable fragment (scFv), or other genetic engineering structures with additional antigen-binding sites to the amino or carboxyl ends of monospecific IgG molecules.Citation2 The most widely format used is IgG with dual-variable domains (DVD-Ig),Citation76 in which a variable part of HL fragment is added to another variable part of another Ab with a short peptide linker. The resulting molecules are bispecific and bivalent with regard to each antigen.Citation77 For example, tetravalent tetraspecific Abs binding EGFR, HER2, HER3, and VEGF are constructed by combining DVD-Ig technology with other methods.Citation78 One of the advantages of BsAbs designed with DVD-Ig technology is their ability simultaneously to bind antigens with all variable domains. This is especially true in cases of binding cytokines and other proteins present in the blood in low concentrations. Also, this allows less frequent administration of these BsAbs.Citation79
Chemical conjugation for the BsAb generation was used for the first time in 1985: two Fab2 obtained by pepsinolysis of rabbit IgG were reduced and then oxidized, resulting in bispecific Fab2.Citation66 Subsequently, homo- and heterobifunctional reagents interacting with cysteine residuesCitation67 and Fab obtained by genetic engineeringCitation80 were used. CovX-Body technology is the most modern approach for the preparation of BsAbs based on the site-specific attachment of low-molecular-weight ligands to IgG.Citation81 The half-life of low-molecular-weight drugs increases significantly after attachment to HL fragments. CVX241 BsAbs were produced by the addition of two short peptides that inhibited VEGF or angiopoietin 2 with a branched linker and then with the Abs.Citation82 However, the clinical trials of CVX241 were terminated ahead of schedule, due to a lack of pharmacological effect.Citation2
Chemical conjugation of Abs against CD3 and CD20 (rituximab) was used to obtain T cells with BsAb-coated surfaces.Citation83 Autologous polyclonal activated T lymphocytes were generated by the surface localization of antigen-binding sites against a CD20 receptor. Such cells in the first phase of clinical trials were administered after high-dose chemotherapy and transplantation of peripheral blood stem cells. Injections increased natural and specific cellular response in refractory non-Hodgkin’s lymphoma.Citation84
IMCgp100, developed by ImmTAC technology, contains a single-chain anti-CD3 mAb with an attached mature T-cell receptor that recognizes peptides of human leukocyte antigen. The drug is now undergoing clinical trials against metastatic melanoma.Citation85 IMCgp100 directs and activates CD8 and CD4+ effector cells and memory cells,Citation86 and then after the death of melanoma cells, its antigens are presented by dendritic cells.Citation87
The dock-and-lock method allows the production of polyvalent, multispecific, and multifunctional constructs.Citation88,Citation89 The linker covalently bonds the first Fab to the dimerization and docking domain of cAMP-dependent protein kinase (contains a sulfhydryl group), and the second Fab bonds to the anchoring domain of A-kinase (contains two sulfhydryl groups). The interaction of the two cAMP-dependent protein-kinase domains results in the dimerization of the structures carrying Fab, and then the resulting fragment binds to the protein A-kinase domain bearing the third Fab. Further, the formation of disulfide bonds covalently stabilizes the triple construction.Citation88,Citation90 The trifunctional constructs containing four cytokine IFNα2b molecules linked to the anti-CD20 Ab (veltuzumab) are effective in non-Hodgkin’s lymphoma and multiple myeloma.Citation91,Citation92 The hexavalent construct containing BsAbs against CD20 and CD22 (veltuzumab and epratuzumab) penetrates lipid rafts, stimulates apoptosis, and inhibits tumor growth.Citation90 A hexavalent construct composed of anti-CD22 and anti-CD19 BsAbs (epratuzumab, hA19) enhances the formation of the immunological synapse and trogocytosis (transmission of antigens) of CD19, CD20, CD21, and CD22 between B lymphocytes and antigen-presenting cells and represents a candidate molecule for the treatment of autoimmune diseases.Citation93 Another construct containing a single-chain anti-CD3 Ab covalently bounds the antitumor Fab dimer; bispecific binding of the BsAb to tumor cells and further to T cells results in antitumor T-cell cytotoxicity.Citation94
Therapeutic Ab fragments (scFv, diabody) may be fused with albuminCitation95 or proteins that bind albumin,Citation96 which increases the half-life of the drug in the blood up to five to six times. The construction of such molecules gives unpredictable results, thereby BsAbs generated as the result of different Ab-fragment fusion or binding of Abs to other proteins have limited application in research and development of new therapeutic molecules. Formats of BsAb generation based on Ab or Ab-fragment conjugation today are not used, because of the possibility of stable fusion recombinant protein generation.Citation2
Protein, cellular, and genetic engineering
Coexpression of two heavy- and two light-chain genes in one cell may result in expression of IgG-like molecules. This method allows the creation of a preparation of monoclonal bispecific anti-CD3/anti-EpCAM Abs (catumaxomab).Citation97 Quadroma technology implies fusion of two cell lines producing Abs, generating a hybrid hybridoma. In such cells, heavy and light chains of two different Abs combine, resulting in BsAbs with conventional IgG-like structure, which retain effector functions mediated by Fc. In cases of quadromas, BsAb constant regions of heavy and light chains can be of the same or different isotypes or even from different species (Triomab).Citation6 Several variants of quadroma-producing cell selection have been described. The first method uses two cell lines producing mAbs resistant to different antibiotics. In this case, hybrid cells carrying resistance markers to both antibiotics are selected.Citation98 One of the advantages of this method is the functional stability of fused nuclei in cell hybrids. Another method uses fluorescence-activated cell sorting: one hybridoma cell line is modified with fluorescein isothiocyanate, another with tetramethyl rhodamine isothiocyanate.Citation99 After fusion, hybrid cells carrying two fluorescent markers are selected. An electrofusion technique, which is innocuous and alternative to polyethylene glycol fusion, is also used.Citation100
The main problem of two-Ab coexpression (particularly quadromas) is the formation of up to nine variants of undesirable chimeric Abs along with the target BsAbs. This problem originates due to the homodimerization of heavy chains (instead of heterodimerization) and random binding of light chains to heavy chains. As a result, the significant disadvantage of this method is very low yield of target BsAbs.Citation2 The yield of heterodimers from two different heavy chains can be increased using the knob-in-hole method, in which one heavy chain with the T366W mutation (“knob”: replacement with a sterically bulky amino acid) joins with the second heavy chain with mutations T366S, L368A, or Y407V (“hole”: replacement by a smaller amino acid); formation of heterodimers is thermodynamically favorable.Citation101,Citation102 A variant of method using structural similarity of IgG and IgA CH3 domains ensures the formation of heterodimers of heavy chains.Citation103 Several methods of BsAb production use just one type of light chain.Citation56 However, the most widely used method of BsAb production today is expression of monospecific mAbs in two different cell lines, isolation and subsequent combination in vitro.Citation104,Citation105 The advantage of this approach is using well-characterized Abs, but the significant disadvantage is the high cost and difficulty of obtaining such BsAbs.Citation2
BsAbs containing a paratope recognizing two different antigens have been demonstrated for an anti-HER2 Ab binding VEGFCitation106 and anti-HER3 Ab binding EGFR.Citation107 CrossMAb technology, developed by Roche, made it possible to generate tetraspecific Abs binding EGFR, HER2, HER3, and VEGF.Citation78 DutaMab technology (Creative Biolabs and Roche) uses three complementarity-determining regions (CDRs) in each antigen-binding site to bind one and three other CDRs to bind the second antigen, thus forming two paratopes. This technology allows BsAbs to be produced using methods typical for monospecific Abs, but the drawback of this approach is its nonuniversality: it is not possible for each pair of antigens to match the paratopes within a single antigen-binding site.Citation2
BsAbs that do not contain constant regions have been described, with the most popular and demanded formats diabody and BiTE. Such BsAbs are expressed in one cell, where the fragments of heavy and light chains are connected by short peptide sequences. ScFvs are widely used for generation of such BsAbs. For diabody generation, sequences encoding two different scFvs are combined into one construct in which heavy chains are expressed in a single polypeptide and then joined with the corresponding light chains. The first described diabodies were bivalent BsAbs.Citation108 Heterodimeric constructs were later obtained by the knob-in-hole methodCitation109 and by single-chain diabody generation.Citation110,Citation111 Diabody technology was used to construct the first BiTEs in eukaryotic cells. BiTEs are small scFv molecules tandem connected by flexible peptide linkers. BiTEs usually contain an antigen-binding site against CD3 and another against a high-affinity surface antigen of tumor cells.Citation44,Citation112,Citation113 A disadvantage of such molecules is their short life span in blood serum, which is associated with smallness and lack of Fc. The advantage of this BsAb format is extremely highly specific antitumor activity at concentrations up to 10 pg/mL in cell culture.Citation114 According to some data, one BiTE molecule can be involved several times in elimination of tumor cells by cytotoxic T lymphocytes.Citation115
Single-domain Ab fragments obtained from mouse and human libraries by a phage display are also used to construct BsAbs.Citation116,Citation117 Nanobodies derived from llamas and camels contain only heavy chains; for the production of BsAbs, nanobodies are readily connected by short peptide linkers.Citation118 The advantages of using single-domain Ab fragments are their smallness, easy penetration into cells, and access to antigens hidden for IgG. A significant disadvantage of such small structures is their low half-life in the blood, which requires frequent injection of the drug.Citation2 The most successful representative of the BiTE family is blinatumomab. Amgen has also developed other BiTEs that bind EpCAM, HER2, CEA, EphA2, MCSP, and CD33, some of which are currently undergoing clinical trials.Citation49
The combination of two VL and VH domains in one polypeptide makes it possible to obtain tetravalent TandAb molecules. TandAb AFM13 combines antigen-binding sites against CD16A and CD30, enhancing the response of natural killers in Hodgkin’s lymphoma. Two antigen-binding sites against each of the antigens and the lack of Fc increase the molecular weight and in vivo stability of these BsAbs.Citation119
Perspectives
Blinatumomab and catumaxomab, approved for use as medicines, are designed to treat oncological diseases. In perspective, one can expect the construction of new platforms that will allow development of full processes from BsAb expression to preclinical tests. To develop new antitumor drugs, it is necessary to search for new combinations of BsAb targets to increase the therapeutic effect and reduce side effects; specific features of tumors also should be taken into account. BsAbs can be used in combination with other medications, eg, drugs controlling the cell cycle, indoleamine dioxygenase inhibitors, and vaccines. There is no doubt that continuous development of new approaches to BsAb production is required to control oncological diseases.
The design of new BsAbs will likely include the ability to bind two or more tumor antigens in combination with the attraction of T lymphocytes and assistant cells into the immunosynapse. Of particular importance is the increase in BsAb specificity and sensitivity, as well as reducing cytotoxicity to nontumor cells. Other important tasks are increasing BsAb yield from hybridomas and decreasing the cost of the drugs.
A promising work on short nonviral minicircle DNA for the synthesis of BsAbs in vivo was recently published. BsAbs against CD3 and CD20 efficiently stimulated T-cell cytotoxicity against CD20+ lymphoma cell lines. Introduction of 5 μg plasmid DNA was sufficient for 1 month’s BsAb expression in mice.Citation120 An oncolytic adenovirus expressing a bispecific single-chain Ab in tumor cells has been constructed. The secreted Abs format is BiTE and BsAbs bind EpCAM on a tumor cell and activate CD4+ and CD8+ T-cell cytotoxicity by directing to CD3.Citation121
A relatively universal method of BsAb preparation is the exchange of IgG HL fragments, which occurs stochastically between natural IgG4,Citation60 IgG1 with a mutation in the CH3 domain,Citation105 and IgG2 through disulfide linkers.Citation122 According to the literature, the HL-fragment exchange occurs in human blood, milk, and placenta between IgG of all subclasses, resulting in polyreactive BsAbs.Citation61–Citation64 However, the exchange of HL fragments between the therapeutic molecules of bispecific IgG4 and the patient’s IgG4 leads to the formation of BsAbs that do not possess the original properties,Citation123 so this imposes significant limitations on this method.
In a short time, one may expect the completion of clinical trials and approval of BsAbs directed to treat autoimmune and other diseases. For example, promising results can be obtained in anti-HIV BsAbs. It has been shown that BsAbs directed against the CD4- and V3-binding sites on the Env (gp120) protein exhibit a synergistic effect in vivo and in vitro. Other variants of anti-HIV BsAbs combine antigen-binding sites against gp41, CD4, or CCR5 protein, and can be used to prevent HIV infection.Citation124,Citation125
The design of bispecific molecules demands analysis of the required BsAb properties (affinity to target molecules, pharmacokinetics in blood, and near target cells) and mechanisms of action. The increasing number of therapeutic BsAbs entering clinical trials and results of BsAb use in clinical medicine may improve understanding of their pharmacokinetics in the near future. The ideal therapeutic BsAbs are expected to have a long half-life in human blood, distribution among organs, and sufficient penetration of tissue.Citation126
The use of BsAbs in diagnostic tools is very promising, since BsAbs simplify the detection of target antigens. BsAbs are used in sensitive immunoassays developed for simple and rapid detection of bacterial and viral infectious diseases and in cancer diagnostics.Citation127 BsAbs significantly enhance the quality and reliability of in vivo cancer-diagnostic imaging by positron-emission tomography. The use of BsAbs and 131I-labeled haptens allows pretargeting human prostate cancer xenografts in severe combined immunodeficient mice. With minimal signal background and high sensitivity and specificity, the use of BsAbs in this method is superior to mAbs.Citation128 Lipoarabinomannan is present in the blood of tuberculosis patients and is considered as a disease marker. The BsAb specific to lipoarabinomannan and horseradish peroxidase was developed with quadroma technology. The use of immunoswabs has shown 100% specificity and 64% sensitivity compared with bacterial cultures. Results were obtained within 2 hours of sample collection, which is very competitive compared to the standard laboratory-culture method, where results are obtained 2–6 weeks after sampling.Citation129 The use of anti-HBsAg × antihuman erythrocyte BsAbs allows detection of hepatitis B with 100% specificity and 97.7% sensitivity. The method can be used in actively infected patients since the serum HBsAg detection level ranges from 5 ng/mL to 600 μg/mL.Citation130
The quadroma expressing BsAbs against Escherichia coli lipopolysaccharide and whole bacteria in combination with horseradish peroxidase was used for rapid one-step sandwich enzyme-linked immunosorbent-assay detection of the E. coli strain O157:H7. The sensitivity of the detection was 100 CFU/mL and specific, since it did not detect Salmonella, Pseudomonas, or nonpathogenic E. coli.Citation131 The same approach was used for Bordetella pertussis detection. A quadroma producing anti-B. pertussis lipopolysaccharide and anti-horseradish peroxidase BsAbs was constructed for ultrasensitive immunoassay.Citation132 A similar method was developed for BsAbs detection of Staphylococcus aureus. The assay is highly sensitive and specific, due to release of a bound fluorescent reporter from a BsAb-active center after binding to the thermonuclease-specific antigen of S. aureus.Citation133 BsAbs recognizing several epitopes of the NP antigen of SARS coronavirus (causing severe acute respiratory syndrome) and binding to horseradish peroxidase were used for immunoswab assays, with sensitivity of NP detection 10 pg/mL and high specificity.Citation134
Conclusion
The development of new methods of BsAb generation made it possible to obtain various variants of promising Ab derivatives for use in therapy. The resulting BsAbs differ from natural IgG by pharmacokinetics, blood serum half-life, ability to penetrate tumors, size, valence, and presence of Fc. Simultaneous blocking of several biological pathways allows BsAbs to exhibit a synergistic effect unachievable with a mixture of monospecific Abs. The results of recent years indicate that in the close future, by combinations of methods developed earlier, new BsAbs directed against a variety of diseases in which simultaneous binding of several specific antigens can play a key role will be generated. BsAbs may be used for the development of diagnostic devices of the “next generation”. The potential of simultaneous detection of several antigens or combining antigen-binding sites with assay markers makes BsAbs an important object of further research in biomedicine, pharmacology, and diagnostics.
Acknowledgments
This study was funded by the Russian Foundation for Basic Research, according to the research projects 16-34-60066-mol_a_dk, 16-15-10103-a, and 16-04-00603-a, and with a grant from the Ministry of Education and Science (MK-410.2017.4).
Disclosure
The authors report no conflicts of interest in this work.
References
- RedmanJMHillEMAlDeghaitherDWeinerLMMechanisms of action of therapeutic antibodies for cancerMol Immunol2015672 Pt A284525911943
- SpiessCZhaiQCarterPJAlternative molecular formats and therapeutic applications for bispecific antibodiesMol Immunol2015672 Pt A9510625637431
- FlorioMGunasekaranKStolinaMA bispecific antibody targeting sclerostin and DKK-1 promotes bone mass accrual and fracture repairNat Commun201671150527230681
- ShimaMHanabusaHTakiMFactor VIII-mimetic function of humanized bispecific antibody in hemophilia AN Engl J Med2016374212044205327223146
- YuYJAtwalJKZhangYTherapeutic bispecific antibodies cross the blood-brain barrier in nonhuman primatesSci Transl Med20146261261ra154
- BrinkmannUKontermannREThe making of bispecific antibodiesMAbs20179218221228071970
- GenoveseMCCohenSMorelandLCombination therapy with etanercept and anakinra in the treatment of patients with rheumatoid arthritis who have been treated unsuccessfully with methotrexateArthritis Rheum20045051412141915146410
- BakerKFIsaacsJDNovel therapies for immune-mediated inflammatory diseases: what can we learn from their use in rheumatoid arthritis, spondyloarthritis, systemic lupus erythematosus, psoriasis, Crohn’s disease and ulcerative colitis?Ann Rheum Dis Epub201781
- TorresTRomanelliMChiricozziAA revolutionary therapeutic approach for psoriasis: bispecific biological agentsExpert Opin Investig Drugs2016257751754
- KhatriAGossSJiangPMansikkaHOthmanAAPharmacokinetics of ABT-122, a TNF-α- and IL-17A-targeted dual-variable domain immunoglobulin, in healthy subjects and patients with rheumatoid arthritis: results from three phase I trialsClin Pharmacokinet Epub2017725
- SilacciMLembkeWWoodsRDiscovery and characterization of COVA322, a clinical-stage bispecific TNF/IL-17A inhibitor for the treatment of inflammatory diseasesMAbs20168114114926390837
- KosloskiMPGossSWangSXPharmacokinetics and tolerability of a dual variable domain immunoglobulin ABT-981 against IL-1α and IL-1β in healthy subjects and patients with osteoarthritis of the kneeJ Clin Pharmacol201656121582159027150261
- ZhangXYangYFanDXiongDThe development of bispecific antibodies and their applications in tumor immune escapeExp Hematol Oncol201761228469973
- KontermannREDual targeting strategies with bispecific antibodiesMAbs20124218219722453100
- No authors listedBispecific antibody therapeutics market3rd edition201720302014 Available from: http://www.reportlinker.com/p02530816-summary/bispecific-antibody-therapeutics-market-edition.htmlAccessed November 27, 2017
- ThakurALumLG“NextGen” biologics: bispecific antibodies and emerging clinical resultsExpert Opin Biol Ther201616567568826848610
- RufPInduction of a long-lasting antitumor immunity by a trifunctional bispecific antibodyBlood20019882526253411588051
- SuryadevaraCMGedeonPCSanchez-PerezLAre BiTEs the “missing link” in cancer therapy?Oncoimmunology201546e100833926155413
- BargouRLeoEZugmaierGTumor regression in cancer patients by very low doses of a T cell-engaging antibodyScience2008321589197497718703743
- ToppMSKuferPGökbugetNTargeted therapy with the T-cell-engaging antibody blinatumomab of chemotherapy-refractory minimal residual disease in B-lineage acute lymphoblastic leukemia patients results in high response rate and prolonged leukemia-free survivalJ Clin Oncol201129182493249821576633
- ToppMSGökbugetNZugmaierGLong-term follow-up of hematologic relapse-free survival in a phase 2 study of blinatumomab in patients with MRD in B-lineage ALLBlood2012120265185518723024237
- ToppMSGökbugetNZugmaierGPhase II trial of the anti-CD19 bispecific T cell-engager blinatumomab shows hematologic and molecular remissions in patients with relapsed or refractory B-precursor acute lymphoblastic leukemiaJ Clin Oncol201432364134414025385737
- ToppMSGökbugetNSteinASSafety and activity of blinatumomab for adult patients with relapsed or refractory B-precursor acute lymphoblastic leukaemia: a multicentre, single-arm, phase 2 studyLancet Oncol2015161576625524800
- Nuñez-PradoNCompteMHarwoodSThe coming of age of engineered multivalent antibodiesDrug Discov Today201520558859425757598
- LöfflerAKuferPLutterbüseRA recombinant bispecific single-chain antibody, CD19×CD3, induces rapid and high lymphoma-directed cytotoxicity by unstimulated T lymphocytesBlood20009562098210310706880
- WuJFuJZhangMLiuDBlinatumomab: a bispecific T cell engager (BiTE) antibody against CD19/CD3 for refractory acute lymphoid leukemiaJ Hematol Oncol2015810426337639
- GoebelerMEBargouRBlinatumomab: a CD19/CD3 bispecific T cell engager (BiTE®) with unique anti-tumor efficacyLeuk Lymphoma20165751021103227050240
- SegalDMWeinerGJWeinerLMBispecific antibodies in cancer therapyCurr Opin Immunol199911555856210508714
- AldossISongJStillerTCorrelates of resistance and relapse during blinatumomab therapy for relapsed/refractory acute lymphoblastic leukemiaAm J Hematol201792985886528494518
- KlingerMBrandlCZugmaierGImmunopharmacologic response of patients with B-lineage acute lymphoblastic leukemia to continuous infusion of T cell-engaging CD19/CD3-bispecific BiTE antibody blinatumomabBlood2012119266226623322592608
- ReuschUDuellJEllwangerKA tetravalent bispecific TandAb (CD19/CD3), AFM11, efficiently recruits T cells for the potent lysis of CD19+ tumor cellsMAbs20157358460425875246
- WuJFuJZhangMLiuDAFM13: a first-in-class tetravalent bispecific anti-CD30/CD16A antibody for NK cell-mediated immunotherapyJ Hematol Oncol201589626231785
- BrentjensRJDavilaMLRiviereICD19-targeted T cells rapidly induce molecular remissions in adults with chemotherapy-refractory acute lymphoblastic leukemiaSci Transl Med20135177177ra38
- FrankelSRBaeuerlePATargeting T cells to tumor cells using bispecific antibodiesCurr Opin Chem Biol201317338539223623807
- ChamesPVan RegenmortelMWeissEBatyDTherapeutic antibodies: successes, limitations and hopes for the futureBr J Pharmacol2009157222023319459844
- ScottAMWolchokJDOldLJAntibody therapy of cancerNat Rev Cancer201212427828722437872
- LinkeRKleinASeimetzDCatumaxomab: clinical development and future directionsMAbs20102212913620190561
- SebastianMPasslickBFriccius-QueckeHTreatment of non-small cell lung cancer patients with the trifunctional monoclonal antibody catumaxomab (anti-EpCAM × anti-CD3): a phase I studyCancer Immunol Immunother200756101637164417410361
- HeissMMMurawaPKoralewskiPThe trifunctional antibody catumaxomab for the treatment of malignant ascites due to epithelial cancer: results of a prospective randomized phase II/III trialInt J Cancer201012792209222120473913
- SeimetzDLindhoferHBokemeyerCDevelopment and approval of the trifunctional antibody catumaxomab (anti-EpCAM × anti-CD3) as a targeted cancer immunotherapyCancer Treat Rev201036645846720347527
- StröhleinMALordickFRüttingerDImmunotherapy of peritoneal carcinomatosis with the antibody catumaxomab in colon, gastric, or pancreatic cancer: an open-label, multicenter, phase I/II trialOnkologie201134310110821358214
- Mau-SørensenMDittrichCDienstmannRA phase I trial of intravenous catumaxomab: a bispecific monoclonal antibody targeting EpCAM and the T cell coreceptor CD3Cancer Chemother Pharmacol20157551065107325814216
- LindhoferHMocikatRSteipeBThierfelderSPreferential species-restricted heavy/light chain pairing in rat/mouse quadromas: implications for a single-step purification of bispecific antibodiesJ Immunol199515512192257602098
- BaeuerlePAReinhardtCBispecific T-cell engaging antibodies for cancer therapyCancer Res200969124941494419509221
- GoereDFlamentCRusakiewiczSPotent immunomodulatory effects of the trifunctional antibody catumaxomabCancer Res201373154663467323737485
- RufPLindhoferHInduction of a long-lasting antitumor immunity by a trifunctional bispecific antibodyBlood20019882526253411588051
- FossatiMBuzzonettiAMonegoGImmunological changes in the ascites of cancer patients after intraperitoneal administration of the bispecific antibody catumaxomab (anti-EpCAM × anti-CD3)Gynecol Oncol2015138234335126049121
- WalshGBiopharmaceutical benchmarks 2010Nat Biotechnol201028991792420829826
- KontermannREBrinkmannUBispecific antibodiesDrug Discov Today201520783884725728220
- OttMGMarméFMoldenhauerGHumoral response to catumaxomab correlates with clinical outcome: results of the pivotal phase II/III study in patients with malignant ascitesInt J Cancer201213092195220321702044
- TrivediAStienenSZhuMClinical pharmacology and translational aspects of bispecific antibodiesClin Transl Sci201710314716228297195
- YuraszeckTKasichayanulaSBenjaminJETranslation and clinical development of bispecific T cell engaging antibodies for cancer treatmentClin Pharmacol Ther2017101563464528182247
- YangFWenWQinWBispecific antibodies as a development platform for new concepts and treatment strategiesInt J Mol Sci2017181E48
- ZhuMWuBBrandlCBlinatumomab, a bispecific T-cell engager (BiTE) for CD-19 targeted cancer immunotherapy: clinical pharmacology and its implicationsClin Pharmacokinet201655101271128827209293
- ChamesPBatyDBispecific antibodies for cancer therapy: the light at the end of the tunnel?MAbs20091653954720073127
- DhimoleaEReichertJMWorld Bispecific Antibody Summit, September 27–28, 2011, Boston, MAMAbs20124141322327426
- YuSLiALiuQRecent advances of bispecific antibodies in solid tumorsJ Hematol Oncol201710115528931402
- NisonoffARiversMMRecombination of a mixture of univalent antibody fragments of different specificityArch Biochem Biophys196193246046213729244
- AalberseRCvan der GaagRvan LeeuwenJSerologic aspects of IgG4 antibodies – I: prolonged immunization results in an IgG4-restricted responseJ Immunol198313027227266600252
- KolfschotenMNSchuurmanJLosenMAnti-inflammatory activity of human IgG4 antibodies by dynamic Fab arm exchangeScience200731758441554155717872445
- SedykhSELekchnovEAPrinceVVBunevaVNNevinskyGAHalf molecular exchange of IgGs in the blood of healthy humans: chimeric lambda-kappa-immunoglobulins containing HL fragments of antibodies of different subclasses (IgG1-IgG4)Mol Biosyst201612103186319527506137
- LekchnovEASedykhSEDmitrenokPSBunevaVNNevinskyGAHuman placenta: relative content of antibodies of different classes and subclasses (IgG1-IgG4) containing lambda- and kappa-light chains and chimeric lambda-kappa-immunoglobulinsInt Immunol201527629730625644595
- SedykhSEBunevaVNNevinskyGAHuman milk IgGs contain various combinations of different antigen-binding sites resulting in multiple variants of their bispecificityPLoS One201278e4294222912765
- SedykhSEBunevaVNNevinskyGAHuman milk sIgA molecules contain various combinations of different antigen-binding sites resulting in a multiple binding specificity of antibodies and enzymatic activities of abzymesPLoS One2012711e4875610.1371/journal.pone.004875623133657
- MilsteinCCuelloACHybrid hybridomas and their use in immunohistochemistryNature198330559345375406137772
- BrennanMDavisonPFPaulusHPreparation of bispecific antibodies by chemical recombination of monoclonal immunoglobulin G1 fragmentsScience1985229470881833925553
- GlennieMJMcBrideHMWorthATStevensonGTPreparation and performance of bispecific F(ab’ gamma)2 antibody containing thioether-linked Fab’ gamma fragmentsJ Immunol19871397236723752958547
- StaerzUDKanagawaOBevanMJHybrid antibodies can target sites for attack by T cellsNature198531460126286312859527
- PerezPHoffmanRWShawSBluestoneJASegalDMSpecific targeting of cytotoxic T cells by anti-T3 linked to anti-target cell antibodyNature198531660263543563160953
- StaerzUDYewdellJWBevanMJHybrid antibody-mediated lysis of virus-infected cellsEur J Immunol19871745715743494618
- ScottAMAllisonJPWolchokJDMonoclonal antibodies in cancer therapyCancer Immun20121214 http://www.ncbi.nlm.nih.gov/pubmed/2289675922896759
- BallEDGuyrePMMillsLFisherJDincesNBFangerMWInitial trial of bispecific antibody-mediated immunotherapy of CD15-bearing tumors: cytotoxicity of human tumor cells using a bispecific antibody comprised of anti-CD15 (MoAb PM81) and anti-CD64/FcγRI (MoAb 32)J Hematother19921185941365020
- ValoneFHKaufmanPAGuyrePMClinical trials of bispecific antibody MDX-210 in women with advanced breast or ovarian cancer that overexpresses HER-2/neuJ Hematother1995454714758581387
- ValoneFHKaufmanPAGuyrePMPhase IA/IB trial of bispecific antibody MDX-210 in patients with advanced breast or ovarian cancer that overexpresses the proto-oncogene HER-2/neuJ Clin Oncol1995139228122927545221
- WeinerLMClarkJIDaveyMPhase I trial of 2B1, a bispecific monoclonal antibody targeting c-erbB-2 and FcγRIIICancer Res19955520458645937553634
- WuCYingHGrinnellCSimultaneous targeting of multiple disease mediators by a dual-variable-domain immunoglobulinNat Biotechnol200725111290129717934452
- JakobCGEdaljiRJudgeRAStructure reveals function of the dual variable domain immunoglobulin (DVD-Ig) moleculeMAbs20135335836323549062
- HuSFuWXuWFour-in-one antibodies have superior cancer inhibitory activity against EGFR, HER2, HER3, and VEGF through disruption of HER/MET crosstalkCancer Res201575115917025371409
- CorreiaISungJBurtonRThe structure of dual-variable-domain immunoglobulin molecules alone and bound to antigenMAbs20135336437223572180
- ShalabyMRShepardHMPrestaLDevelopment of humanized bispecific antibodies reactive with cytotoxic lymphocytes and tumor cells overexpressing the HER2 protooncogeneJ Exp Med199217512172251346155
- DoppalapudiVRTryderNLiLChemically programmed antibodies: endothelin receptor targeting CovX-BodiesBioorg Med Chem Lett200717250150617055724
- DoppalapudiVRHuangJLiuDChemical generation of bispecific antibodiesProc Natl Acad Sci U S A201010752226112261621149738
- GallJMDavolPAGrabertRCDeaverMLumLGT cells armed with anti-CD3 × anti-CD20 bispecific antibody enhance killing of CD20+ malignant B cells and bypass complement-mediated rituximab resistance in vitroExp Hematol200533445245915781336
- LumLGThakurALiuQCD20-targeted T cells after stem cell transplantation for high risk and refractory non-Hodgkin’s lymphomaBiol Blood Marrow Transplant201319692593323529012
- IamsWTSosmanJAChandraSNovel targeted therapies for metastatic melanomaCancer J2017231545828114255
- BoudousquieCBossiGHurstJMRygielKAJakobsenBKHassanNJPolyfunctional response by ImmTAC (IMCgp100) redirected CD8+ and CD4+ T cellsImmunology2017152342543828640942
- BossiGBuissonSOatesJJakobsenBKHassanNJImmTAC-redirected tumour cell killing induces and potentiates antigen cross-presentation by dendritic cellsCancer Immunol Immunother201463543744824531387
- RossiEAGoldenbergDMCardilloTMMcBrideWJSharkeyRMChangCHStably tethered multifunctional structures of defined composition made by the dock and lock method for use in cancer targetingProc Natl Acad Sci U S A2006103186841684616636283
- ChangCHRossiEAGoldenbergDMThe dock and lock method: a novel platform technology for building multivalent, multifunctional structures of defined composition with retained bioactivityClin Cancer Res200713185586S5591S17875793
- RossiEAGoldenbergDMCardilloTMSteinRChangCHHexavalent bispecific antibodies represent a new class of anticancer therapeutics – 1: properties of anti-CD20/CD22 antibodies in lymphomaBlood2009113246161617119372261
- RossiEAGoldenbergDMCardilloTMSteinRChangCHCD20-targeted tetrameric interferon-α, a novel and potent immunocytokine for the therapy of B-cell lymphomasBlood2009114183864387119710501
- RossiEARossiDLSteinRGoldenbergDMChangCHA bispecific antibody-IFNα2B immunocytokine targeting CD20 and HLA-DR is highly toxic to human lymphoma and multiple myeloma cellsCancer Res201070197600760920876805
- RossiEAChangCHGoldenbergDMAnti-CD22/CD20 bispecific antibody with enhanced trogocytosis for treatment of lupusPLoS One201495e9831524841238
- RossiDLRossiEACardilloTMGoldenbergDMChangCHA new class of bispecific antibodies to redirect T cells for cancer immunotherapyMAbs20146238139124492297
- MüllerDKarleAMeissburgerBHöfigIStorkRKontermannREImproved pharmacokinetics of recombinant bispecific antibody molecules by fusion to human serum albuminJ Biol Chem200728217126501266017347147
- StorkRMüllerDKontermannREA novel tri-functional antibody fusion protein with improved pharmacokinetic properties generated by fusing a bispecific single-chain diabody with an albumin-binding domain from streptococcal protein GProtein Eng Des Sel2007201156957617982179
- CheliusDRufPGruberPStructural and functional characterization of the trifunctional antibody catumaxomabMAbs20102330931920418662
- BosRNieuwenhuizenWEnhanced transfection of a bacterial plasmid into hybridoma cells by electroporation: application for the selection of hybrid hybridoma (quadroma) cell linesHybridoma199211141511737639
- KarawajewLMicheelBBehrsingOGaestelMBispecific antibody-producing hybrid hybridomas selected by a fluorescence activated cell sorterJ Immunol Methods19879622652702433353
- CaoYSureshMRBispecific antibodies as novel bioconjugatesBioconjug Chem1998966356449815155
- RidgwayJBPrestaLGCarterP“Knobs-into-holes” engineering of antibody CH3 domains for heavy chain heterodimerizationProtein Eng1996976176218844834
- AtwellSRidgwayJBWellsJACarterPStable heterodimers from remodeling the domain interface of a homodimer using a phage display libraryJ Mol Biol1997270126359231898
- DavisJHAperloCLiYSEED bodies: fusion proteins based on strand-exchange engineered domain (SEED) CH3 heterodimers in an Fc analogue platform for asymmetric binders or immunofusions and bispecific antibodies†Protein Eng Des Sel201023419520220299542
- StropPHoWHBoustanyLMGenerating bispecific human IgG1 and IgG2 antibodies from any antibody pairJ Mol Biol2012420320421922543237
- LabrijnAFMeestersJIde GoeijBEEfficient generation of stable bispecific IgG1 by controlled Fab-arm exchangeProc Natl Acad Sci U S A2013110135145515023479652
- BostromJYuSFKanDVariants of the antibody herceptin that interact with HER2 and VEGF at the antigen binding siteScience200932359211610161419299620
- SchaeferGHaberLCrockerLMA two-in-one antibody against HER3 and EGFR has superior inhibitory activity compared with monospecific antibodiesCancer Cell201120447248622014573
- HolligerPProsperoTWinterG“Diabodies”: small bivalent and bispecific antibody fragmentsProc Natl Acad Sci U S A19939014644464488341653
- ZhuZPrestaLGZapataGCarterPRemodeling domain interfaces to enhance heterodimer formationProtein Sci200864781788
- KipriyanovSMMoldenhauerGSchuhmacherJBispecific tandem diabody for tumor therapy with improved antigen binding and pharmacokineticsJ Mol Biol19992931415610512714
- AltMMüllerRKontermannRENovel tetravalent and bispecific IgG-like antibody molecules combining single-chain diabodies with the immunoglobulin γ1 Fc or CH3 regionFEBS Lett19994541–2909410413102
- MackMRiethmüllerGKuferPA small bispecific antibody construct expressed as a functional single-chain molecule with high tumor cell cytotoxicityProc Natl Acad Sci U S A19959215702170257624362
- BaeuerlePAKuferPBargouRBiTE: teaching antibodies to engage T-cells for cancer therapyCurr Opin Mol Ther2009111223019169956
- DreierTLorenczewskiGBrandlCExtremely potent, rapid and costimulation-independent cytotoxic T-cell response against lymphoma cells catalyzed by a single-chain bispecific antibodyInt J Cancer2002100669069712209608
- HaasCKrinnerEBrischweinKMode of cytotoxic action of T cell-engaging BiTE antibody MT110Immunobiology2009214644145319157637
- WardESGüssowDGriffithsADJonesPTWinterGBinding activities of a repertoire of single immunoglobulin variable domains secreted from Escherichia coliNature198934162425445462677748
- DaviesJRiechmannLAntibody VH domains as small recognition unitsBiotechnology (N Y)19951354754799634788
- ConrathKELauwereysMWynsLMuyldermansSCamel single-domain antibodies as modular building units in bispecific and bivalent antibody constructsJ Biol Chem2001276107346735011053416
- RotheASasseSToppMSA phase 1 study of the bispecific anti-CD30/CD16A antibody construct AFM13 in patients with relapsed or refractory Hodgkin lymphomaBlood2015125264024403125887777
- PangXMaFZhangPTreatment of human B-cell lymphomas using minicircle DNA vector expressing anti-CD3/CD20 in a mouse modelHum Gene Ther201728221622527802782
- FreedmanJDHagelJScottEMOncolytic adenovirus expressing bispecific antibody targets T-cell cytotoxicity in cancer biopsiesEMBO Mol Med2017981067108728634161
- PattersonJTGrosEZhouHChemically generated IgG2 bispecific antibodies through disulfide bridgingBioorg Med Chem Lett201727163647365228720505
- LabrijnAFBuijsseAOvan den BremerETTherapeutic IgG4 antibodies engage in Fab-arm exchange with endogenous human IgG4 in vivoNat Biotechnol200927876777119620983
- HuangYYuJLanziAEngineered bispecific antibodies with exquisite HIV-1-neutralizing activityCell201616571621163127315479
- BournazosSGazumyanASeamanMSNussenzweigMCRavetchJVBispecific anti-HIV-1 antibodies with enhanced breadth and potency in brief bispecific anti-HIV-1 antibodies with enhanced breadth and potencyCell201616571609162027315478
- ChenYXuYPharmacokinetics of bispecific antibodyCurr Pharm Rep201733126137
- ByrneHConroyPJWhisstockJCO’KennedyRJA tale of two specificities: bispecific antibodies for therapeutic and diagnostic applicationsTrends Biotechnol2013311162163224094861
- McBrideWJZanzonicoPSharkeyRMBispecific antibody pretargeting PET (immunoPET) with an 124I-labeled hapten-peptideJ Nucl Med200647101678168817015905
- SarkarSTangXLDasDSpencerJSLowaryTLSureshMRA bispecific antibody based assay shows potential for detecting tuberculosis in resource constrained laboratory settingsPLoS One201272e3234022363820
- ChenYPQiaoYYZhaoXHChenHSWangYWangZRapid detection of hepatitis B virus surface antigen by an agglutination assay mediated by a bispecific diabody against both human erythrocytes and hepatitis B virus surface antigenClin Vaccine Immunol200714672072517442848
- GuttikondaSTangXLYangBMArmstrongGDSureshMRMono-specific and bispecific antibodies against E. coli O157 for diagnosticsJ Immunol Methods20073271–21917804009
- TangXLPepplerMSIrvinRTSureshMRUse of bispecific antibodies in molecular velcro assays whose specificity approaches the theoretical limit of immunodetection for Bordetella pertussisClin Vaccine Immunol2004114752757
- WagstaffeSJHillKEWilliamsDWBispecific antibody-mediated detection of the Staphylococcus aureus thermonucleaseAnal Chem201284145876588422650421
- SunwooHHPalaniyappanAGangulyAQuantitative and sensitive detection of the SARS-CoV spike protein using bispecific monoclonal antibody-based enzyme-linked immunoassayJ Virol Methods20131871727822995576