Abstract
Aim
To investigate the effect of endogenous n-3 polyunsaturated fatty acids (PUFAs) on bone marrow adipogenesis under osteoporosis conditions.
Methods
A mouse osteoporosis model overexpressing the FAT1 gene from Caenorhabditis elegans and converting n-6 PUFAs to n-3 PUFAs endogenously was used.
Results
The mice presented significantly lower bone marrow adiposity (adipocyte volume/tissue volume, mean adipocyte number) but increased the bone parameters (bone mineral density, bone mineral content, bone volume/total volume) in the distal femoral metaphysis.
Conclusion
Endogenous n-3 PUFAs protect bone marrow adipogenesis, which provides a novel drug target.
Introduction
Osteoporosis is a common clinical disease in postmenopausal women and aging populations, is caused by an imbalance between bone resorption and bone formation, and leads to osteoporotic fractures and other serious complications.Citation1 Age-induced osteoporosis is mainly associated with increased bone marrow adipocytes and bone loss.Citation2,Citation3 In osteoporotic bone, reduced bone formation is directly correlated with increasing adipose tissue.Citation4
Osteoblasts and marrow adipocytes are both differentiated from a common precursor: bone marrow mesenchymal stem cells (BMSCs).Citation5 An in vitro study has revealed that mesenchymal stem cells (MSCs) derived from postmenopausal women with osteoporosis are more likely to differentiate into adipocytes than osteoblasts, compared with cells isolated from healthy women.Citation6 There is virtual redifferentiation plasticity in both osteoblasts and adipocytes. It has been demonstrated that fully differentiated osteoblasts from human MSCs presented capabilities of differentiation and transdifferentiation into adipocytes and vice versa.Citation7 It also has been reported that the differentiation of MSCs into adipocytes or osteoblasts is competitively balanced within the bone marrow.Citation8 Beresford and coworkers have shown the inverse relationship between the differentiation of adipocytic and osteogenic cells.Citation9 Moreover, Clabaut et al proposed that MSC-derived adipocytes are able to induce MSC-derived osteoblasts to differentiate to an adipocyte phenotype within an in vitro coculture system.Citation10 On the other hand, various studies have indicated that adipocytes can secrete factors, such as hormones, leptin, adiponectin, and fatty acids, which affect the proliferation, apoptosis, and function of neighboring cells.Citation11,Citation12 The evidence from the investigation conducted by Elbaz et al suggests an effect of lipotoxicity from marrow adipocytes on osteoblast function.Citation13 In addition, cross talk between adipocytes and osteoclasts has been unearthed recently. It has been reported that preadipocytes have a stronger ability in supporting osteoclast-like cell formation without prostaglandin E2 than preosteoblasts, suggesting an emerging relationship between bone marrow-cell adipogenesis and osteoclastogenesis.Citation14
The existing evidence delineates that bone marrow-fat accumulation may play a critical role in the onset of osteoporosis, owing to its potential effects on either the commitment of BMSC differentiation or the homeostasis of osteoclast-mediated bone resorption. Based on this notion, reversal bone marrow adipogenesis may be an effective therapeutic strategy for the prevention of age-related osteoporosis.
N-3 long-chain polyunsaturated fatty acids (PUFAs) are essential fatty acids for human beings, and include alpha-linolenic acid, eicosapentaenoic acid, and docosahexaenoic acid, which cannot be self-synthesized in mammals.Citation15 During the past few decades, n-3 PUFAs have been supposed to be beneficial for prevention from bone loss in both in vitro and in vivo studies. However, the underlying mechanisms of this action have not yet been fully defined.Citation16–Citation22 A recent study has identified that inhibition of osteoclast differentiation and activation may be one of the mechanisms involved in preventing osteoporosis by n-3 PUFAs (n-6:n-3 = 1:1).Citation23
To the best of our knowledge, there are no reports regarding the roles and mechanisms of n-3 PUFAs on bone marrow adipogenesis. In the present study, by employing a transgenic (TG) mouse model that overexpresses the FAT1 gene from Caenorhabditis elegans and converts n-6 PUFAs to n-3 PUFAs endogenously at a fixed ratio compared to other diet-formulated approaches, we aimed firstly to investigate the effect of endogenous n-3 PUFAs on bone marrow adipogenesis under osteoporosis conditions, which could give rise to a novel insight into current therapy for antiosteoporosis. Peroxisome proliferator-activated receptor gamma (PPARγ) is an essential transcription factor in adipogenesis,Citation24 the expression and/or activity of which determines the commitment of BMSCs to osteoblasts or adipocyte lineages.Citation25 Runt-related transcription factor 2 (RUNX2) is a key regulator of osteoblast differentiation, playing an important role in bone formation,Citation26 which was used as the measure of adipogenesis/osteogenesis in the present study.
Materials and methods
Animal model and diet feeding
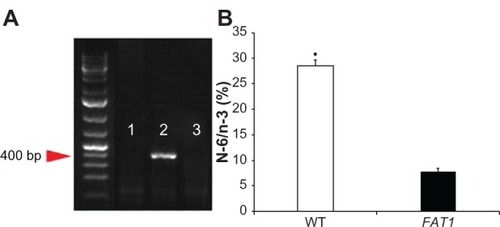
FAT1 TG mice and C57BL/6 wild-type (WT) control mice were donated by Professor Yifan Dai. Since the FAT1 gene is a kind of heterozygote gene, breeder mice were mated with WT C57BL/6 mice to obtain female FAT1 gene-positive C57BL6 mice and FAT1 gene-negative C57BL/6 mice screened by genotyping using a polymerase chain reaction kit (Takara, Dalian, People’s Republic of China) (). The amount of n-3 PUFAs and n-6 PUFAs in mouse tails were measured by gas–liquid chromatography (). The mice were maintained in cages (two to four mice per cage) under controlled laboratory conditions: 12-hour light/dark cycle at 24°C, fed normal diets and tap water. At age 2 months, 40 mice (20 FAT1 mice, 20 WT mice) with matched weight received sham or ovariectomy surgeries. Then they were divided into four groups (ten mice/group): group A, FAT1 mice sham; group B, FAT1 ovariectomized (OVX); group C, WT sham; group D, OVX. All animals were fed with standard high-fat diets (Research Diets, New Brunswick, NJ, USA) containing 44.9 kcal% fat, 35.1 kcal% carbohydrate, and 20 kcal% protein. When the mice were 5 months old, they were killed. The final body weight was measured and, bilateral femurs were excised. Then we stripped the soft tissue on bones and immersed them in 4% paraformaldehyde at 4°C for 24 hours.
Figure 1 (A and B) Genotyping. (A) PCR genotyping confirmation using FAT1 fragment-specific primers indicated FAT1 gene expression (lane 1) and wild-type (WT) gene expression (lane 2); lane 3 is the negative control. (B) N-6/n-3 ratio of FAT1 mice (mean = 7.762) was significantly lower than WT mice (mean = 28.43).
This study was approved by the Animal Ethics Committee of Southern Medical University, and all experiment procedures strictly followed principles of the National Institutes of Health Guide.
Micro-CT analysis
Micro-computed tomography (CT) was performed on each bone and focused on distal femoral metaphysis using a ZKKS-MCT-Sharp-III scanner system (Caskaisheng, Guangzhou, People’s Republic of China). A small field was selected for scanning and was corrected for the CT value, a 70 kV scanning voltage, 30 W power, 429 μA current, and 5 μm scan thickness. The 3D-MED 3.0 (Institute of Automation, Chinese Academy of Sciences, Beijing, People’s Republic of China) was used for 3-D knee reconstruction and image capture. Our analysis included various bone parameters: bone morphometry, bone mineral density (BMD; volumetric), total volume (TV), bone volume (BV), BV/TV, and trabecular number (Tb.N).
Bone marrow adiposity analysis
The samples of femur bone were processed for decalcification, as previously describedCitation28 and then routinely embedded in paraffin. Serial sections (4 μm) were obtained from the sagittal plane of the femur condyle. Hematoxylin and eosin and toluidine blue staining were conducted to quantify proximal metaphyseal adipocyte parameters: adipocyte number (AD#, per mm2), percent adipocyte volume per tissue volume (AV/TV). A uniform number of fields were screened in all sections by three individuals, starting three fields from the left end and three fields from the top endocortical surface,Citation29 excluding the adipocytes with disruption in the fields. To avoid any bias in the final analysis, all sections were read in a blinded way without knowledge of the groups (FAT1 or WT). Pictures were obtained at 20× magnifications using an Olympus (Tokyo, Japan) BX51RF stereomicroscope. All measurements were done at 20× magnification using the Image-Pro Plus analysis software (Media Cybernetics, Rockville, MD, USA).
Immunofluorescence and immunohistochemistry analysis for PPARγand RUNX2 protein expression
The following primary antibodies were used: rabbit antimouse PPARγ monoclonal antibody (Cell Signaling Technology, Beverly, MA, USA) and rabbit antimouse RUNX2 polyclonal antibody (Bioworld Technology, Louis Park, MN, USA). Tissue sections were incubated in sodium citrate buffer (0.01 M, pH 6.0) for 10 minutes to retrieve antigen after deparaffinization and rehydration. The next step was quenching the endogenous peroxidase with 3% hydrogen peroxide prepared in 100% ethanol for 5 minutes. Nonspecific binding was blocked by addition of goat serum for 1 hour at room temperature. Then, sections were incubated with primary antibodies - either anti-PPARy or anti-RUNX2 - overnight at 4°C. After being washed in phosphate-buffered saline three times, sections were further incubated with biotinylated secondary antibody (antirabbit immunoglobulin G) for 30 minutes at room temperature. Sections were counterstained with hematoxylin. As negative control, nonimmune mouse immunoglobulin G was used as the primary antibody, and this gave uniformly negative results (data not shown). Pictures were obtained at 20× magnification using an Olympus BX51RF stereomicroscope. The quantification of the level of these two proteins was analyzed by Image-Pro Plus analysis software at 20× magnification.
Statistical analysis
Results were expressed as mean ± standard error of mean, and data were analyzed with the SPSS 13.0 software (IBM, Armonk, NY, USA) using one-way analysis of variance and least significant difference. Post hoc Tukey’s tests were performed with SPSS software directly. P-values < 0.05 were considered statistically significant.
Results
The effect of endogenous n-3 PUFAs on body weight and abdominal adipose tissue in OVX-induced osteoporosis model
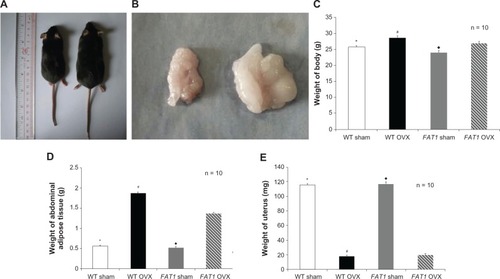
There was a difference in the weight of uteruses between the OVX and sham groups, which identified the sound establishment of the OVX-induced osteoporosis model in mice. At the beginning of this study, all mice were well matched for age and weight. The values for body weight and abdominal adipose tissue between FAT1 and WT OVX mice were different (P < 0.05) 3 months postoperatively, but there was no significant difference in body weight between FAT1 and WT sham mice (P > 0.05). OVX groups showed higher levels of body weight compared with sham groups ().
Figure 2 (A–E) Gross examination and tissue weight. Three months after ovariectomy, there was increased body weight (C) and abdominal adipose tissue (D) in the ovariectomized (OVX) group compared to the sham group. However, the FAT1 OVX group presented significant lower body weight (A and C) and abdominal adipose tissue (B and D) compared to the wild-type (WT) OVX group. The weight of uterus was significantly lower in the OVX group than the sham group (E).
P < 0.05 versus FAT1 OVX; #
P < 0.05 versus FAT1 OVX.
Effect of endogenous n-3 PUFAs on femur bone microstructure morphometries
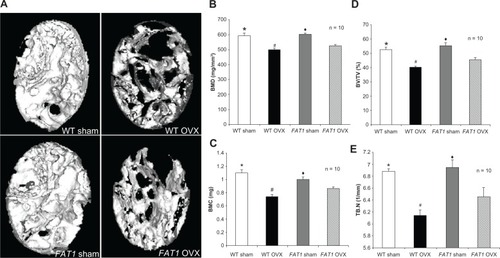
BMD, BMC, BV/TV, and Tb.N in the OVX groups were lower in the femoral region than the sham group. BMD, BV/TV, and BMC in the distal femur of FAT1 OVX mice were higher (P < 0.05) compared to WT OVX mice. However, the Tb.N in the WT OVX group mildly decreased (P > 0.05) compared with the FAT1 OVX group. There was no statistical significance between WT and FAT1 sham mice in femur bone microstructure morphometries ().
Figure 3 (A–E) Bone parameter assessment by micro-computed tomography scan. (A) Three-dimensional reconstructions of the distal femoral metaphysis of FAT1 and wild-type (WT) mice by micro-CT scan. The FAT1 ovariectomized (OVX) group exhibited more bone mass than the WT OVX group. (B–D) Quantitation of bone parameters of distal femur bone. The values of bone mineral density (BMD, B), bone mineral content (BMC, C), bone volume/total volume (BV/TV, D), and trabecular number (Tb.N) (E) in the OVX group were decreased compared with those in the sham group. The bone parameter (BMD, BMC, BV/TV) values of the FAT1 OVX group were significantly higher than the WT OVX group, but the Tb.N in the FAT1 OVX group only mildly increased than WT OVX group.
P < 0.05 versus FAT1 OVX; (B–E) *P < 0.05 versus WT OVX; (B–E) ♦
P < 0.05 versus FAT1 OVX.
Effect of endogenous n-3 PUFAs on bone marrow adiposity
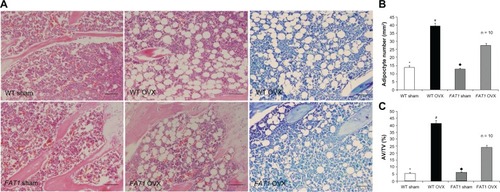
The adipocyte parameters in the WT and FAT1 groups are described in . There was increased AD# in the OVX group compared with the sham group (P < 0.05) for both WT and FAT1 mice. Adipocyte parameters such as AV/TV and AD# from histological analyses confirmed extensive fat accumulation in the WT OVX group versus the FAT1 OVX group (P < 0.05), but the adipocyte parameters between the FAT1 and WT sham groups remained stable ().
Figure 4 (A–C) Histological analysis of bone marrow adiposity. (A) Hematoxylin and eosin and toluidine blue staining for distal femur bone marrow. Images were taken at 200x magnification. There were fewer adipocytes in the FAT1 OVX group than the wild-type (WT) ovariectomized (OVX) group. (B) #, per mm2) and (C) adipocyte volume/tissue volume (AV/TV) were significantly reduced in the FAT1 OVX group. Three months after ovariectomy, the OVX group showed more bone marrow adiposity than the sham group (P < 0.05).
P < 0.05 versus FAT1 OVX. Scale bars, 100 μm
Effect of endogenous n-3 PUFAs on PPARγ and RUNX2 expression in bone marrow
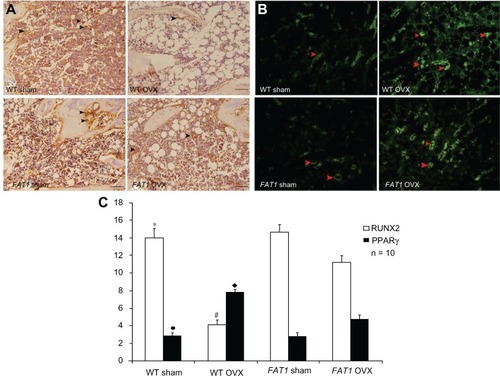
In agreement with immunofluorescence data, the intensity of positive staining for PPARγ was enhanced by OVX significantly (P < 0.05), and the level of PPARγ protein expression was lower in the FAT1 OVX group compared with the WT OVX group (P < 0.05). There was no significant difference between WT and FAT1 sham mice in the intensity of PPARγ-positive staining.
In immunohistochemistry data, positive immunoreaction of RUNX2 appearing in osteoblasts was stronger in the sham group than the OVX group (P < 0.05). In the WT OVX group, RUNX2-positive staining was elevated compared with FAT1 OVX mice (P < 0.05). There was no statistical significance between WT and FAT1 sham mice in the level of RUNX2 protein expression ().
Figure 5 (A–C) Immunohistochemistry and counting of positively stained cells in bone marrow three months after ovariectomy. The black arrows indicate the runt-related transcription factor 2 (RUNX2) positively stained cells, and the red arrows indicate peroxisome proliferator-activated receptor gamma (PPARγ) positively stained cells (200× magnification, scale bar = 100 μm) (A and C). The proportion of RUNX2-positive cells was significantly increased in the FAT1 ovariectomized (OVX) group compared to wild-type (WT mice). (B and C) The proportion of PPARγ-positive cells was significantly reduced in the FAT1 OVX group compared to WT mice.
P > 0.05 versus FAT1 sham. Scale bars, 100 μm
Discussion
In the present study, the effect of endogenous n-3 PUFAs on bone marrow adipogenesis was investigated in a FAT1 TG OVX model. Our data suggested that TG mice present significantly lower bone marrow adiposity (AV/TV, AD#) but increased bone parameters (BMD, BMC, BV/TV) in the distal femoral metaphysis.
Despite the fact that the intimate relationship between bone marrow adipogenesis and bone loss has been realized in osteoporosis pathogenesis,Citation30,Citation31 involving several studies on the potential effect of dietary or endogenous n-3 PUFAs on osteogenesis and osteoclastogenesis,Citation32–Citation34 the influence of endogenous n-3 PUFAs on adipogenesis in bone marrow is still poorly understood.
For better understanding of the effect of endogenous n-3 PUFAs, TG mice with FAT1 gene overexpression were developed, which was able to show a much lower n-6:n-3 PUFA ratio of ~1:1 compared to WT mice with ratio of 20–30:1.Citation23 Therefore, we used FAT1 transgenic mice in our study to investigate bone marrow adipogenesis under osteoporosis situations for the first time, taking advantage of its steady endogenous n-3 PUFA ratio.
Since adipogenesis and osteoblastogenesis from BMSCs are regulated by key transcription factors – PPARγ and RUNX2, respectively – expression of same could determine the establishment of adipocytes or osteoblasts. In this study, we observed a significantly lower expression of PPARγ but higher RUNX2 expression in bone marrow of TG mice when compared to WT mice, which confirmed the inhibitory effect of n-3 PUFAs on bone marrow adipogenesis theoretically.
With regard to the effect of fat on bone, peripheral adipose tissue and bone marrow should be taken into consideration. It has been suggested peripheral adipose tissue influences bone through stresses caused by mechanical loading.Citation35 Recent in vitro and in vivo studies have drawn a conclusion that lowering dietary the n-6:n-3 PUFA ratio is associated with lower intra-abdominal fat mass and lower area of adipocytes from retroperitoneal fat,Citation36 which suggests that n-3 PUFA additions can inhibit the formation of peripheral adipocytes hypertrophy and hyperplasia. However, the effect of endogenous n-3 PUFAs on bone marrow adiposity and their possible mechanism needs further investigation due to underlying endocrine functions in bone, since adipocytes in bone marrow are an important source of adipokines that could have a direct lipotoxic effect on BMSCs and osteoblasts, which leads to bone loss.Citation13 Our results revealed that endogenous n-3 PUFAs not only suppress the whole body weight of mice but also inhibit adiposity accumulation in bone marrow, preventing bone loss induced by ovariectomy in mice, indicating the potential effects of endogenous n-3 PUFAs on BMSCs, the unique precursors of bone marrow adipocytes. Future studies should look into the differentiation-modulation mechanism of BMSCs by endogenous n-3 PUFAs.
It has been suggested the current antiosteoporosis drugs, such as strontium ranelate and risedronate, which have been widely used in the prevention and treatment of osteoporosis in postmenopausal women,Citation37,Citation38 could inhibit bone loss partially by retarding bone marrow adipogenesis, with alteration of the RUNX2/PPARγ balance during BMSC differentiation. This study on FAT1 transgenic mice provides compelling evidence that bone marrow adipose is the critical therapeutic target of osteoporosis, and endogenous n-3 PUFAs are proposed as a novel treatment in preventing postmenopausal osteoporosis.
Some hypotheses about the mechanisms by which n-3 PUFAs prevent bone mass loss associated with menopause have been proposed – enhancement of osteoblastogenesis and reduction of osteoclast activity – but these mechanisms are not yet fully understood. Combined with their effect of alleviation of proinflammatory cytokines such as interleukin 6, interleukin 1, and tumor necrosis factor-α secreted by bone marrow adipocytesCitation39,Citation40 and the effect of the marrow microenvironment modulating osteoclastogenic activity and increasing bone resorption,Citation41–Citation45 endogenous n-3 PUFAs, which alleviated bone marrow adiposity in an osteoporosis animal model, may target bone marrow adipocytes by attenuating adipogenesis of BMSCs, subsequently increasing the osteoblastogenesis. However, the specific signaling pathway mechanism involved in this process should be further pursued.
In conclusion, in this study, we showed that the endogenous conversion of n-6 PUFA to n-3 PUFA and maintaining a lower ratio of n-6:n-3 PUFAs could attenuate adipogenesis within the bone marrow, which is associated with decreasing the level of PPARγ and increasing the level of RUNX2 expression.
Conclusion
Our findings provide us with a more comprehensive understanding of the mechanism of action of n-3 PUFAs on promoting osteoblastogenesis and perhaps will lead to the development of a therapeutic approach for osteoporosis aimed at reversing adipocyte differentiation from BMSCs.
Disclosure
The authors report no conflicts of interest in this work.
References
- [No authors listed] Consensus development conference: diagnosis, prophylaxis, and treatment of osteoporosis Am J Med 1993 94 646 650 8506892
- Meunier P Aaron J Edouard C Vignon G Osteoporosis and the replacement of cell populations of the marrow by adipose tissue. A quantitative study of 84 iliac bone biopsies Clin Orthop Relat Res 1971 80 147 154 5133320
- Griffith JF Yeung DK Antonio GE Vertebral marrow fat content and diffusion and perfusion indexes in women with varying bone density: MR evaluation Radiology 2006 241 831 838 17053202
- Verma S Rajaratnam JH Denton J Hoyland JA Byers RJ Adipocytic proportion of bone marrow is inversely related to bone formation in osteoporosis J Clin Pathol 2002 55 693 698 12195001
- Owen M Marrow stromal stem cells J Cell Sci Suppl 1988 10 63 76 3077943
- Rodríguez JP Montecinos L Ríos S Reyes P Martínez J Mesenchymal stem cells from osteoporotic patients produce a type I collagen-deficient extracellular matrix favoring adipogenic differentiation J Cell Biochem 2000 79 557 565 10996846
- Song L Tuan RS Transdifferentiation potential of human mesenchymal stem cells derived from bone marrow FASEB J 2004 18 980 982 15084518
- Muruganandan S Roman AA Sinal CJ Adipocyte differentiation of bone marrow-derived mesenchymal stem cells: cross talk with the osteoblastogenic program Cell Mol Life Sci 2009 66 236 253 18854943
- Beresford JN Bennett JH Devlin C Leboy PS Owen ME Evidence for an inverse relationship between the differentiation of adipocytic and osteogenic cells in rat marrow stromal cell cultures J Cell Sci 1992 102 341 351 1400636
- Clabaut A Delplace S Chauveau C Hardouin P Broux O Human osteoblasts derived from mesenchymal stem cells express adipogenic markers upon coculture with bone marrow adipocytes Differentiation 2010 80 40 45 20466479
- Maurin AC Chavassieux PM Frappart L Delmas PD Serre CM Meunier PJ Influence of mature adipocytes on osteoblast proliferation in human primary cocultures Bone 2000 26 485 489 10773588
- Maurin AC Chavassieux PM Vericel E Meunier PJ Role of polyunsaturated fatty acids in the inhibitory effect of human adipocytes on osteoblastic proliferation Bone 2002 31 260 266 12110443
- Elbaz A Wu X Rivas D Gimble JM Duque G Inhibition of fatty acid biosynthesis prevents adipocyte lipotoxicity on human osteoblasts in vitro J Cell Mol Med 2010 14 982 991 19382912
- Sakaguchi K Morita I Murota S Relationship between the ability to support differentiation of osteoclast-like cells and adipogenesis in murine stromal cells derived from bone marrow Prostaglandins Leukot Essent Fatty Acids 2000 62 319 327 10883064
- Zevenbergen H de Bree A Zeelenberg M Laitinen K van Duijn G Flöter E Foods with a high fat quality are essential for healthy diets Ann Nutr Metab 2009 54 15 24 19641346
- Watkins BA Li Y Lippman HE Seifert MF Omega-3 polyunsaturated fatty acids and skeletal health Exp Biol Med 2001 226 485 497
- Fernandes G Lawrence R Sun D Protective role of n-3 lipids and soy protein in osteoporosis Prostaglandins Leukot Essent Fatty Acids 2003 68 361 372 12798656
- Watkins BA Li Y Lippman HE Feng S Modulatory effect of omega-3 polyunsaturated fatty acids on osteoblast function and bone metabolism Prostaglandins Leukot Essent Fatty Acids 2003 68 387 398 12798659
- Sun D Krishnan A Zaman K Lawrence R Bhattacharya A Fernandes G Dietary n-3 fatty acids decrease osteoclastogenesis and loss of bone mass in ovariectomized mice J Bone Miner Res 2003 18 1206 1216 12854830
- Green KH Wong SC Weiler HA The effect of dietary n-3 long-chain polyunsaturated fatty acids on femur mineral density and biomarkers of bone metabolism in healthy, diabetic and dietary-restricted growing rats Prostaglandins Leukot Essent Fatty Acids 2004 71 121 130 15207529
- Watkins BA Reinwald S Li Y Seifert MF Protective actions of soy isoflavones and n-3 PUFAs on bone mass in ovariectomized rats J Nutr Biochem 2005 16 479 488 16043030
- Shen CL Yeh JK Rasty J Li Y Watkins BA Protective effect of dietary long-chain n-3 polyunsaturated fatty acids on bone loss in gonad-intact middle-aged male rats Br J Nutr 2006 95 462 468 16512931
- Rahman MM Bhattacharya A Banu J Kang JX Fernandes G Endogenous n-3 fatty acids protect ovariectomy induced bone loss by attenuating osteoclastogenesis J Cell Mol Med 2009 13 1833 1844 20141608
- Wood RJ Vitamin D and adipogenesis: new molecular insights Nutr Rev 2008 66 40 46 18254883
- Moerman EJ Teng K Lipschitz DA Lecka-Czernik B Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: the role of PPAR-gamma2 transcription factor and TGF-beta/BMP signaling pathways Aging Cell 2004 3 379 389 15569355
- Yoshida CA Furuichi T Fujita T Core-binding factor beta interacts with Runx2 and is required for skeletal development Nat Genet 2002 32 633 638 12434152
- Wei D Li J Shen M Cellular production of n-3 PUFAs and reduction of n-6-to-n-3 ratios in the pancreatic beta-cells and islets enhance insulin secretion and confer protection against cytokine-induced cell death Diabetes 2010 59 471 478 19933995
- Elbaz A Rivas D Duque G Effect of estrogens on bone marrow adipogenesis and Sirt1 in aging C57BL/6J mice Biogerontology 2009 10 747 755 19333775
- Syed FA Oursler MJ Hefferanm TE Peterson JM Riggs BL Khosla S Effects of estrogen therapy on bone marrow adipocytes in postmenopausal osteoporotic women Osteoporos Int 2008 19 1323 1330 18274695
- Meunier P Aaron J Edouard C Vignon A Osteoporosis and the replacement of cell populations of the marrow by adipose tissue Clin Orthop Rel Res 1971 80 147 154
- Justesen J Stenderup K Ebbesen EN Mosekilde L Steiniche T Kassem M Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis Biogerontology 2001 2 165 171 11708718
- Kesavalu L Vasudevan B Raghu B Omega-3 fatty acid effect on alveolar bone loss in rats J Dent Res 2006 85 648 652 16798867
- Bhattacharya A Rahman M Sun D Fernandes G Effect of fish oil on bone mineral density in aging C57BL/6 female mice J Nutr Biochem 2007 18 372 379 16963250
- Lau BY Ward WE Kang JX Ma DW Femur EPA and DHA are correlated with femur biomechanical strength in young fat-1 mice J Nutr Biochem 2009 20 453 461 18708283
- Taes YE Lapauw B Vanbillemont G Fat mass is negatively associated with cortical bone size in young healthy male siblings J Clin Endocrinol Metab 2009 94 2325 2331 19401374
- Massiera F Saint-Marc P Seydoux J Arachidonic acid and prostacyclin signaling promote adipose tissue development: a human health concern? J Lipid Res 2003 44 271 279 12576509
- Fournier C Perrier A Thomas M Reduction by strontium of the bone marrow adiposity in mice and repression of the adipogenic commitment of multipotent C3H10T1/2 cells Bone 2012 50 499 509 21839191
- Jin J Wang L Wang XK Risedronate inhibits bone marrow mesenchymal stem cell adipogenesis and switches RANKL/OPG ratio to impair osteoclast differentiation J Surg Res 2013 180 e21 e29 22487394
- Fried SK Bunkin DA Greenberg AS Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: depot difference and regulation by glucocorticoid J Clin Endocrinol Metab 1998 83 847 850 9506738
- Timper K Grisouard J Sauter NS Glucose-dependent insulinotropic polypeptide induces cytokine expression, lipolysis and insulin resistance in human adipocytes Am J Physiol Endocrinol Metab 2013 304 E1 E13 23092914
- Jilka RL Hangoc G Girasole G Increased osteoclast development after estrogen loss: mediation by interleukin-6 Science 1992 257 88 91 1621100
- Manolagas SC Role of cytokines in bone resorption Bone 1995 17 2 Suppl 63S 67S 8579900
- Lee YM Fujikado N Manaka H Yasuda H Iwakura Y IL-1 plays an important role in the bone metabolism under physiological conditions Int Immunol 2010 22 805 816 20679512
- Hashizume M Mihara M The roles of interleukin-6 in the pathogenesis of rheumatoid arthritis Arthritis 2011 2011 765624 22046525
- Maurin AC Chavassieux PM Frappart L Delmas PD Serre CM Meunier PJ Influence of mature adipocytes on osteoblast proliferation in human primary cocultures Bone 2000 26 485 489 10773588