
Abstract
Enicosanthellum pulchrum is a tropical plant from Malaysia and belongs to the Annonaceae family. This plant is rich in isoquinoline alkaloids. In the present study, liriodenine, an isoquinoline alkaloid, was examined as a potential anticancer agent, particularly in ovarian cancer. Liriodenine was isolated by preparative high-performance liquid chromatography. Cell viability was performed to determine the cytotoxicity, whilst the detection of morphological changes was carried out by acridine orange/propidium iodide assay. Initial and late apoptosis was examined by Annexin V-fluorescein isothiocyanate and DNA laddering assays, respectively. The involvement of pathways was detected via caspase-3, caspase-8, and caspase-9 analyses. Confirmation of pathways was further performed in mitochondria using a cytotoxicity 3 assay. Apoptosis was confirmed at the protein level, including Bax, Bcl-2, and survivin, while interruption of the cell cycle was used for final validation of apoptosis. The result showed that liriodenine inhibits proliferation of CAOV-3 cells at 37.3 μM after 24 hours of exposure. Changes in cell morphology were detected by the presence of cell membrane blebbing, chromatin condensation, and formation of apoptotic bodies. Early apoptosis was observed by Annexin V-fluorescein isothiocyanate bound to the cell membrane as early as 24 hours. Liriodenine activated the intrinsic pathway by induction of caspase-3 and caspase-9. Involvement of the intrinsic pathway in the mitochondria could be seen, with a significant increase in mitochondrial permeability and cytochrome c release, whereas the mitochondrial membrane potential was decreased. DNA fragmentation occurred at 72 hours upon exposure to liriodenine. The presence of DNA fragmentation indicates the CAOV-3 cells undergo late apoptosis or final stage of apoptosis. Confirmation of apoptosis at the protein level showed overexpression of Bax and suppression of Bcl-2 and survivin. Liriodenine inhibits progression of the CAOV-3 cell cycle in S phase. These findings indicate that liriodenine could be considered as a promising anticancer agent.
Introduction
Enicosanthellum pulchrum (King) Heusden belongs to the family Annonaceae, which is also known as a family of “mempisang” in Malaysia.Citation1 This species is often found in the middle of the highlands, and the distribution is mostly in the Cameron Highland, Malaysia, as reported by Chua et al.Citation2 The species is a medium-sized tree that can reach up to 3–5 m in height.Citation3 Phytochemical analysis of this plant has reported some known alkaloids in the bark and roots, including (−)-asimilobine, liriodenine, (−)-anonaine, (−)-norliridinine (−)-scoulerine,Citation4 and cleistopholine.Citation5 Biological activity has also been reported for the species and compounds, including anti-platelet activating factor, antibacterial, and antiulcer activity.Citation5–Citation7
Liriodenine (8H-[1,3]benzodioxolo[6,5,4-de]benzo[g] quinolin-8-one), is an isoquinoline alkaloid. This compound is widely distributed and acts as a chemotaxonomic marker in the Annonaceae family.Citation8 Biological studies in vivo indicate that liriodenine has antiarrhythmic activity,Citation9 and its potential as antimicrobial, antibacterial, antifungal,Citation10–Citation12 mutagenic, and antiplatelet agentsCitation13,Citation14 has been demonstrated in in vitro studies. Previous studies have also reported that liriodenine has prominent cytotoxic effects in several cancer cell lines, inducing G1 cell cycle arrest and repressing DNA synthesis in HepG2 and SK-Hep-1 cells.Citation15 A report by Chen et al showed liriodenine to have potent activity in colon cancer, and that this compound could inhibit the SW480 cell cycle through the nitric oxide-dependent and p53-dependent G1/S phase arrest pathway.Citation16 In addition, liriodenine suppressed proliferation of A549 human lung adenocarcinoma cells in a time-dependent fashion.Citation17 These early findings indicate the strong potential of liriodenine as a therapeutic agent for various types of cancers. The present study assessed liriodenine as an anticancer agent, particularly for human ovarian cancer which is the first conducted in-depth study for the mechanism of apoptosis in vitro.
Materials and methods
Plant materials
The plant E. pulchrum was from the Cameron Highlands Mountain Forest, Pahang, Malaysia. The specimen was identified by the late Kamaruddin Mat Salleh from the Faculty of Science and Technology, University Kebangsaan Malaysia. A voucher specimen (SM769) was lodged with the Botany Department Herbarium, University Kebangsaan Malaysia. The air-dried roots were ground to 40–60 mesh.
Root extraction
A total of 100 g of roots were extracted successively by the maceration technique using n-hexane, ethyl acetate (EtOAc), and methanol (MeOH) (Merck, Whitehouse Station, NJ, USA) solvents. The EtOAc extract (1.96 g) was taken and separated by vacuum liquid chromatography using silica gel type H (Sigma-Aldrich, St Louis, MO, USA) on a gradient of n-hexane-CHCl3 (100:0, 70:30, 50:50, 30:70, and 0:100) and CHCl3-MeOH (100:0, 50:50, and 0:100). In total, five fractions were obtained after 10 fractions were combined based on thin layer chromatography analysis.
Preparative HPLC separation
Preparative high-performance liquid chromatography (HPLC) was performed by injecting 2 mL of filtered MeOH fraction 4 onto a Prep Nova-Pak (10 mm ×20 mm ×30 mm) HR C-18 reversed-phase HPLC column (Waters, Milford, MA, USA). The fractions were eluted using a 322 Gilson pump (Gilson, Fairfield, NJ, USA) at a permanent flow rate of 12 mL/min. The solvent system consisted of 10% acetonitrile in water to 100% acetonitrile (v/v) and flow gradiently for 90 minutes. Liriodenine (8.0 mg, 0.4% of yield) was detected at 254 nm absorbance using a Gilson 156 ultraviolet-visible absorbance detector. The compound was collected at 26–28 minutes.
Cell viability assay
Human ovarian cancer (CAOV-3 and SKOV-3) and human hepatic (WRL-68) cell lines were purchased from the American Tissue Culture Collection (Manassas, VA, USA), and an immortalized human ovarian epithelial cell line (SV-40) was purchased from ABM Inc (Richmond, BC, Canada). Briefly, confluence cells were harvested and centrifuged at 1,800 rpm for 5 minutes. The concentration of cells was adjusted to 1×106 cells/mL. The cells were then treated with liriodenine at various concentrations in a 96-well plate and incubated for 24, 48, and 72 hours. A total of 20 μL of 3-(4,5-dimethylthiazol-2yl)-2,5-diphenyl tetrazolium bromide (MTT) reagent (Invitrogen, Carlsbad, USA) was added to each well and incubated for 3 hours. The plate was recorded at an absorbance of 570 nm to determine the IC50 value. Paclitaxel and cisplatin were used as the positive control in this assay.
Acridine orange/propidium iodide double staining
Apoptosis were counted by propidium iodide (PI) and acridine orange (AO) double staining according to the standard procedure and were examined under a fluorescent microscope (Leica, Wetzlar, Germany) attached to Q-Floro software. In brief, the assay was carried out in a 25 mL culture flask (Nunc®, Sigma-Aldrich). CAOV-3 cells were seeded at density of 1×106 cells/mL and treated with an IC50 concentration of liriodenine. The flasks were then incubated at 37°C for 24, 48, and 72 hours. The harvested cells were washed and stained with fluorescent dyes (AO/PI) comprising 10 μL of AO (10 μg/mL) and 10 μL of PI (10 μg/mL). The morphology of the cells was observed under an ultraviolet-fluorescent BX60 microscope (Olympus, Tokyo, Japan) within 30 minutes, before the intensity of fluorescent started to disappear.
Annexin-V-FITC assay
CAOV-3 cells (5×104 cells/mL) were seeded into a six-well plate. After exposure to liriodenine for 24, 48, and 72 hours, the cells were harvested and washed with phosphate-buffered saline. The assay was performed using an FITC (fluorescein isothiocyanate) Annexin V Apoptosis Detection Kit I (BD Pharmingen™, San Diego, CA, USA). The treated cells were centrifuged at 1,600 rpm for 5 minutes. Next, 100 μL of each sample were taken and placed into a tube containing 5 μL of FITC Annexin V and 10 μL of PI stain. The suspension was mixed, and 100 μL of 1× Assay buffer was added per tube. All samples were then analyzed using a flow cytometer (BD FACSCanto™ II, San Jose, CA, USA).
Caspase analysis
The caspase-3, caspase-8, and caspase-9 colorimetric assays were performed using a commercial kit (R&D Systems Inc, Minneapolis, MN, USA). The cells were seeded and treated with the IC50 concentration in 25 mL flasks for 24, 48, and 72 hours. Cells that had been stimulated for apoptosis were collected and centrifuged at 1,600 rpm for 10 minutes. The cell pellet was lysed by addition of lysis buffer. Prior to centrifugation, the lysate cells were incubated on ice for 10 minutes and then centrifuged at 10,000× g for 1 minute. The assay was carried out in a 96-well flat bottom microplate. Next, 50 μL of cell lysate and 50 μL of 2× reaction buffer 3, 8, or 9 were added in each well; 5 μL of caspase-3, caspase-8, or caspase-9 colorimetric substrate (LEHD-pNA) was then added to each reaction well and incubated at 37°C for 1 hour. The plate was read on a luminescence microplate reader (Infinite M200 PRO, Tecan, Männedorf, Switzerland) at a wavelength of 405 nm.
Multiple cytotoxicity assays
Multiple cytotoxicity assays were carried out using the Cellomics® Multiparameter Cytotoxicity 3 kit (Thermo Scientific, Pittsburgh, PA, USA). The assay was performed using a 96-well microplate. The cells were seeded in the plate at a concentration of 5×103 cells per well. The cells were then treated with liriodenine at concentrations of 20, 30, and 40 μM, respectively, and incubated overnight at 37°C and 5% CO2 saturation. Briefly, several solutions were added successively in each well containing 50 μL of live cell staining, 100 μL of fixation solution, 100 μL of 1× permeabilization buffer, and 100 μL of 1× blocking buffer for an incubation duration of 30, 20, 10, and 15 minutes, respectively. Two antibodies solutions (primary and secondary antibody) were used, whereby 50 μL of each solution were added to the wells. The plate was then read and the results were evaluated on an ArrayScan HCS reader.
Analysis of DNA fragmentation
The CAOV-3 cells were plated at a density of 5×104 cells/mL in a 25 mL flask and incubated for 24 hours. The cells were then treated with liriodenine for 24, 48, and 72 hours. A Suicide-Track™ DNA Ladder isolation kit (Calbiochem KgaA, Darmstadt, Germany) was used to analyze the DNA fragments (mononucleosome and oligonucleosomes) formed during apoptosis. In brief, the cells were trypsinized and centrifuged at 1,800 rpm for 5 minutes. The pellet was gently resuspended in 55 μL of solution 1, 20 μL of solution 2, and 25 μL of solution 3 (kit components). After incubation overnight at 50°C, the mixture was mixed with 500 μL of resuspension buffer. DNA laddering was detected by preparing 1.5% agarose gel in 1× TAE buffer with the staining reagent supplied in the kit. The gel was run consistently at 50 V until the dye front was 1–2 cm from the bottom of the gel. The DNA was visualized by ultraviolet light transilluminator and photographed.
Western blot analysis
The CAOV-3 cells were seeded in a 75 mL culture flask and treated with liriodenine at 24, 48, and 72 hours. The cells were then harvested by centrifugation at 13,000 rpm for 10 seconds and resuspended in 400 μL of Pro-Prep™ solution. Cell lysis was induced by incubation at −20°C for 20 minutes.
After incubation, the cell pellet was centrifuged at 13,000 rpm (4°C) for 5 minutes. Total proteins were taken and mixed with loading dye before separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis for 90 minutes. The gel was then transferred to a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA) and allowed to run for another 90 minutes. The membrane was blocked with 5 mL of 5% bovine serum albumin and incubated overnight. The appropriate primary antibodies for β-actin (1:1,000), Bax (1:1,000), Bcl-2 (1:1,000), and survivin (1:1,000) were used (Abcam, Inc, Burlingame, CA, USA) and conjugated with secondary antibody (anti-rabbit) for 1 hour at room temperature. The bound antibody was identified using peroxidase-conjugated and exposed for a few minutes to allow the appearance of bands. The polyvinylidene difluoride membrane was finally viewed and photographed using an ultraviolet light transilluminator.
Cell cycle analysis
The CAOV-3 cancer cells were seeded into a 25 mL culture flask at a concentration of 1×106 cells/mL and treated with liriodenine for 24, 48, and 72 hours. The cells were then centrifuged at 1,800 rpm for 5 minutes. The supernatant was removed and the pellet was washed with phosphate-buffered saline. The cells were fixed with fixation solution (700 μL of 90% cold ethanol [EtOH]) and kept at 4°C overnight to restore the integrity of the cells. EtOH was discarded by centrifuging at 1,800 rpm for 5 minutes. The cells were then rinsed, and 600 μL of phosphate-buffered saline was added. A total of 25 μL of RNase A (10 mg/mL) and 50 μL of PI (1 mg/mL) were added to the fixed cells, which were then incubated for 1 hour at 37°C. Next, the DNA content of the cells was analyzed by flow cytometer (BD FACS Canto™ II).
Statistical analysis
Each test was performed as three separate experiments, and the results are the reported as mean ± standard deviation. The data were evaluated by one-way analysis of variance followed by Dunnett’s test using Statistical Package for the Social Sciences version 17.0 software (Chicago, IL, USA) to determine statistical significance and GraphPad Prism version 4.0 (GraphPad Software Inc, La Jolla, CA, USA) to determine the IC50 values.
Results
Identification of liriodenine
Liriodenine () appeared in the form of yellow needles. The 1H and 13C nuclear magnetic resonance spectroscopic data of compound in this study was compared with liriodenine structure that has been identified in other species.Citation18 However, this current study reported nuclear magnetic resonance spectroscopic data in one- and two-dimensional nuclear magnetic resonance, which has never been reported before (heteronuclear single quantum coherence and heteronuclear multiple-bond correlation) techniques and electrospray ionization mass spectrometry ().
Table 1 1H (600 MHz) and 13C (150 MHz) nuclear magnetic resonance data for liriodenine
Figure 1 Structure of liriodenine.
Liriodenine inhibited CAOV-3 cells in vitro
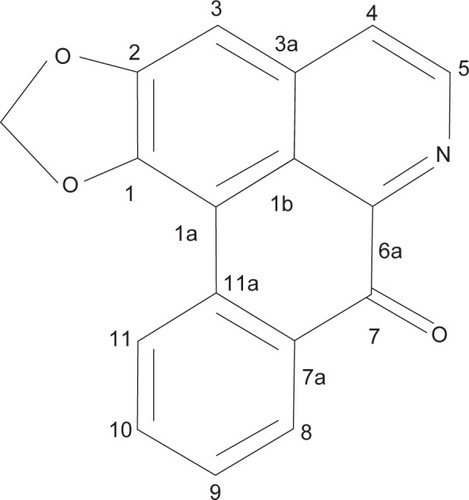
The cytotoxic effects of liriodenine were studied in CAOV-3, SKOV-3, and normal ovarian (WRL-68 and SV-40) cell lines. Two standard drugs, paclitaxel and cisplatin, were used as positive controls in this experiment (). The IC50 results show that liriodenine inhibited CAOV-3 cell growth at a concentration of 37.3±1.06 μM after 24 hours of exposure. However, the IC50 concentration decreased after the cells were exposed to liriodenine at 48 and 72 hours, with values of 26.3±0.07 μM and 23.1±1.62 μM, respectively (). The effect of liriodenine on CAOV-3 was significant when compared with cisplatin as a standard drug. Meanwhile, liriodenine exhibited less activity in SKOV-3 cells, with IC50 values of 68.0±1.56, 61.1±3.09, and 46.5±1.55 μM at 24, 48, and 72 hours, respectively. Additionally, both the normal ovarian cell lines displayed no evidence of cytotoxicity at a concentration higher than 100 μM.
Table 2 Inhibitory effects of study compounds at 24 hours
Figure 2 IC50 value of liriodenine at different time points in a CAOV-3 cell line.
Determination of changes in morphology using AO/PI double staining
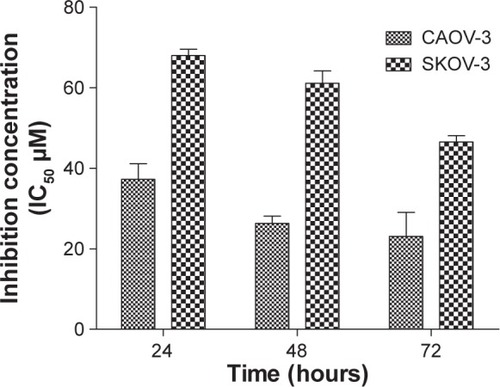
Morphological changes in CAOV-3 cells were observed under the fluorescence microscope after exposure to liriodenine. These changes could be seen at 24, 48, and 72 hours (). In comparison with untreated CAOV-3 cells, the treated cells showed an intact green nuclear structure with a round shape and no disruption. After 24 hours of treatment with liriodenine, the morphology of CAOV-3 cells showed cell membrane blebbing and fragmented DNA with bright green fluorescence. The changes could be clearly observed after 48 and 72 hours of treatment, with clear growth inhibition, increased cell membrane blebbing, presence of more apoptotic bodies, and also the appearance of a reddish-orange color due to PI being bound to denatured DNA cells, indicating dead cells. Furthermore, the numbers of CAOV-3 cells that underwent early and late apoptosis increased in a time-dependent manner.
Figure 3 Fluorescent micrographs and quantitative analysis of CAOV-3 cells double-stained with acridine orange and propidium iodide. Cells were treated with liriodenine in a time-dependent manner.
Abbreviations: VI, viable cells; BL, blebbing of cell membrane; CC, chromatin condensation; EA, early apoptosis; LA, late apoptosis; AB, apoptotic bodies; VS, production of vesicles.
Annexin V analysis
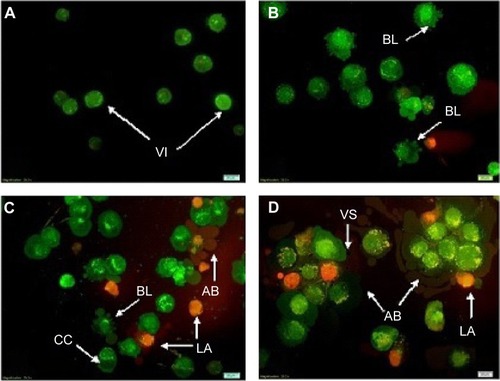
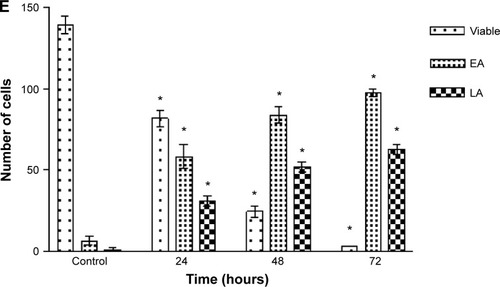
CAOV-3 cells undergoing early apoptosis after being treated with liriodenine was detected by Annexin V-FITC assay. The Annexin V-FITC assay showed an initial stimulation of apoptosis in a time-dependent manner (). The treated CAOV-3 cells exhibited a significant increase (P<0.05) of early apoptosis by 22.0%, 30.1%, and 36.6% at 24, 48, and 72 hours, respectively. Conversely, the percentages of viable cells were reduced from 54.7%, 41.3%, and 29.8%. The exposure of liriodenine in CAOV-3 cells at a longer period (24 to 72 hours) caused decrease in viable cells and increase in cell death.
Figure 4 Flow cytometry analysis of Annexin V-fluorescein isothiocyanate in CAOV-3 cells treated with liriodenine in a time-dependent manner.
Abbreviations: PI-A, propidium iodide; FITC, fluorescein isothiocyanate; Q, quadrant; Q1, secondary necrosis; Q2, late apoptosis; Q3, viable cells; Q4, early apoptosis.
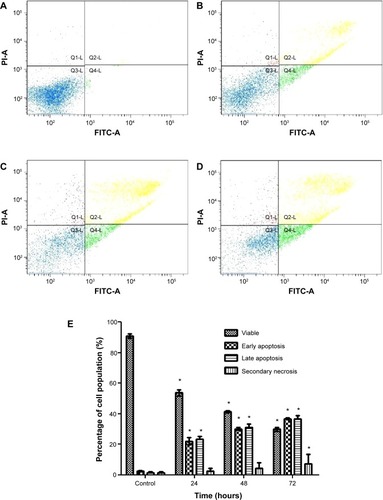
Liriodenine induced caspase activity
CAOV-3 cells treated with liriodenine were analyzed for induction of caspase-3, caspase-8, and caspase-9. The results showed that liriodenine stimulates both caspase-3 and caspase-9, as indicated by increasing values for luminescence units over time (). There was a significant difference (P<0.05) between treated and untreated cells for caspase-3 and caspase-9 during the three stipulated time periods, while the luminescence unit for caspase-8 decreased. Stimulation of caspase-3 and caspase-9 indicates that the apoptosis triggered by liriodenine involves the intrinsic pathway.
Figure 5 The colorimetric analysis of caspase-3, caspase-8, and caspase-9 in untreated and treated CAOV-3 cells with liriodenine at 24, 48, and 72 hours.
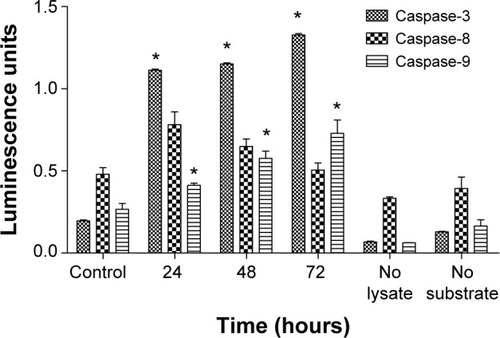
Liriodenine induced apoptosis on mitochondrial disruption
The Multiparameter Cytotoxicity 3 assay (Cellomics) was carried out to investigate four parameters that are important in the apoptosis process, ie, total nuclear intensity, mitochondrial membrane potential, cell permeability, and cytochrome c release (). The results indicated a reduction in total nuclei (Hoechst dye), which refers to the number of CAOV-3 cells present after being treated with liriodenine for 24 hours. The same situation occurred in the mitochondrial membrane potential, whereby there was a significant reduction in intensity (P<0.05) in cells treated with liriodenine 30 and 40 μM, respectively, at 24 hours. Meanwhile, cell permeability and cytochrome c were significantly increased (P<0.05) when compared with untreated CAOV-3 cells at the same concentrations.
Figure 6 Representative images and quantitative analysis of CAOV-3 cells treated or not treated with liriodenine at 24 hours.
Liriodenine triggered DNA fragmentation in CAOV-3 cells
CAOV-3 cells treated with liriodenine at concentrations of 37 μM showed the presence of DNA laddering after 72 hours of treatment (). The presence of laddering indicated DNA fragmentation in CAOV-3 cells. The positive control used in this study comprised HL-60 cells treated with actinomycin D, which showed formation of a clear ladder and had close similarities to the marker.
Figure 7 Electrophoretic separation of fragmented DNA for CAOV-3 cells treated or not treated with liriodenine.
Abbreviation: bp, base pairs.

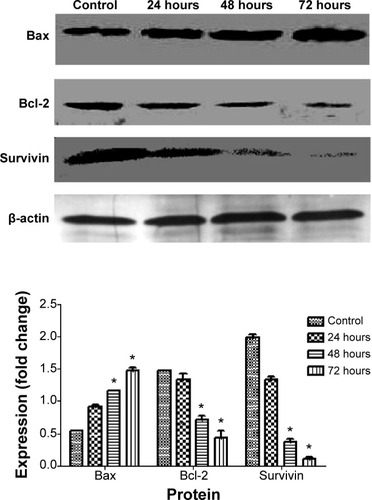
Western blot analysis
Involvement of the mitochondria in apoptosis was confirmed at the protein level by Western blotting. There were three proteins involved, ie, Bax, Bcl-2, and survivin. We observed upregulation of Bax protein in CAOV-3 cells treated with liriodenine when compared with untreated CAOV-3 cells. In contrast, Bcl-2 and survivin showed downregulation of protein expression after exposed to liriodenine at 24, 48, and 72 hours (). Moreover, there were significant differences (P<0.05) in all proteins at 72 hours between CAOV-3 cells treated or not treated with liriodenine.
Figure 8 Effect of liriodenine on level of apoptosis in regulatory proteins at 24, 48, and 72 hours, with β-actin as a loading control.
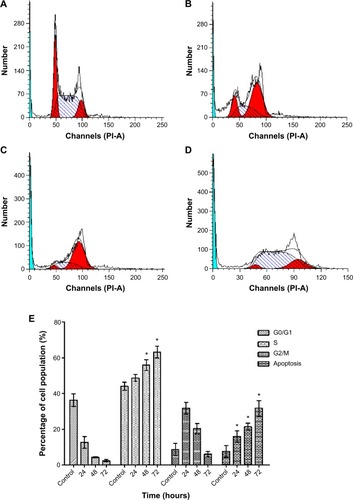
Liriodenine inhibited proliferation of CAOV-3 cells and arrested the cell cycle at S phase
Analysis of cell cycle arrest by liriodenine in CAOV-3 cells found that this alkaloid inhibits cell proliferation at S phase (). The results showed a significant (P<0.05) increase in the number of treated cells in S phase at 24–72 hours when compared with untreated cells. However, uneven changes occurred at the three time points for G0/G1 and G2/M phases, indicating that division of CAOV-3 cells halted during S phase.
Figure 9 Flow cytometric analysis of cell cycle distribution of CAOV-3 cells treated with liriodenine in a time-dependent manner.
Abbreviation: PI-A, propidium iodide.
Discussion
The main aim of this study was to evaluate liriodenine, which was isolated through the modern technique of preparative high-performance liquid chromatography, in the search for potential anticancer drugs particularly for human ovarian cancer. The effect of liriodenine on CAOV-3 cells in vitro was observed by investigation of the mechanisms of induction of apoptosis. The ability of liriodenine to inhibit 50% growth of CAOV-3 cells at concentrations as low as 37 μM after 24 hours of treatment showed that liriodenine and cisplatin are both comparable in terms of their activity. However, the findings of this study demonstrate that liriodenine appears to be better than cisplatin, an existing drug that is used extensively in the treatment of ovarian cancer. In light of the current findings, liriodenine can potentially overcome the resistance problems faced by some drugs.Citation19
Detection of apoptosis by morphological changes in CAOV-3 cells was observed using the AO/PI assay under fluorescence microscopy. Changes in the treated cells were similar to those described by previous researchers,Citation20,Citation21 which can be seen via formation of a dense and packed cytoplasm as well as chromatin condensation.Citation19 In addition, longer exposure of the cells to liriodenine can lead to aggressive changes in cell morphology, such as the presence of extensive cell membrane blebbing. After extensive cell membrane blebbing occurs, it will be followed by the formation of apoptotic bodies through the separation of cell fragments.Citation22 Annexin V-FITC is an important marker for confirmation of apoptosis in cells due to the presence of an intact cell membrane in the early phase of apoptosis.Citation22 This condition allows Annexin V-labeled FITC to be bound to the outer membrane of the cell, whereby the presence of translocated phosphatidylserine can later be detected by flow cytometry.Citation23,Citation24 This action happened as early as 24 hours, as shown in . Induction of apoptosis by liriodenine through the intrinsic pathway was also seen in activation of caspase-9. The intrinsic pathway is one of the crucial apoptotic pathways, as opposed to the extrinsic pathway.Citation22,Citation25 Once this pathway has been activated, several factors will cause the mitochondria to be involved in downstream apoptotic signaling.
Factors involved in downstream events of apoptosis can be explained as chaos in the inner mitochondrial membrane, leading to opening of the mitochondrial permeability transition pore and damage to the mitochondrial transmembrane potential, which finally allows cytochrome c to be released from the mitochondria into the cytoplasm.Citation26 Involvement of that mitochondria can also be supported through several proteins that trigger apoptosis, including Bax and Bcl-2. The Bcl-2 and Bax proteins play a vital role as antiapoptotic and proapoptotic factors, respectively. Both of these proteins have special significance in determination of apoptosis or otherwise.Citation27 In the present study, both proteins exhibited an opposing physiology action on cancer cells after exposure to liriodenine, suggesting that apoptosis was induced in CAOV-3 cells.
One of the common processes that is widely used in the determination of apoptosis is DNA fragmentation.Citation22 This phenomenon occurs when the cell enters the final phase of apoptosis. As shown in , the formation of a DNA ladder indicates DNA fragmentation occurring in treated-CAOV-3 cells after 72 hours of exposure. The existence of the DNA ladder provides evidence of DNA fragmentation as a result of endonuclease cleavage when caspase-3 breaks down.Citation28,Citation29
The effects of liriodenine on proliferation of CAOV-3 cells were also examined by cell cycle analysis and expression of survivin protein. This protein not only inhibits apoptosis but is also involved in regulation of cell division and enhancement of angiogenesis.Citation30 Survivin is expressed at high levels on many cancer cells, including ovarian cancer,Citation31 making this protein a good marker for use in this study. In the present study, liriodenine successfully decreased the expression of surviving, as shown in . A similar situation was observed in the cell cycle analysis, where inhibition of survivin caused the CAOV-3 cells to stop dividing due to arrest in S phase (). When the cell cycle is disturbed, DNA becomes damage and apoptosis is initiated.Citation32 Based on all the evidence and the results shown, liriodenine does positively induce apoptosis in CAOV-3 cells.
Conclusion
Liriodenine induced apoptosis in CAOV-3 cells via the mitochondrial signaling pathway by involvement of caspase-3 and caspase-9. Similarly, liriodenine was able to inhibit proliferation of CAOV-3 cells. This study demonstrates that liriodenine is a promising potential drug candidate for the treatment of ovarian cancer.
Acknowledgments
The authors thank the University of Malaya for providing the financial support (PG151-2012B) that enabled us to conduct this research. We would also wish to express our utmost gratitude and appreciation to the late Prof Datuk Dr A Hamid A Hadi for his help and support throughout the study.
Disclosure
The authors report no conflicts of interest in this work.
References
- BurkillIHA Dictionary of the Economic Products of the Malay Peninsula2nd edKuala Lumpur, MalaysiaMinistry of Agriculture and Cooperatives1966
- ChuaLCompleted data collection forms for endemic trees of Peninsular Malaysia1997
- The IUCN Red List of Threatened SpeciesDisepalum pulchrum2014 Available from: www.iucnredlist.orgAccessed December 15, 2014
- LavaultMGuinaudeauHBrunetonJSevenetTHadiHA(−)- Thaipetaline, a tetrahydroproto berberine from a Malayan AnnonaceaePhytochemistry19902938453847
- NordinNJalilJJantanIMuradSPlatelet-activating factor (PAF) receptor binding activity of the roots of Enicosanthellum pulchrumPharm Biol20125028429022103812
- NordinNSyam MohanNMZajmiAAntioxidant, anticancer and antimicrobial activities of methanolic extracts from Enicosanthellum pulchrum (King) HeusdenSains Malaysiana20144315151521
- NordinNSalamaSMGolbabapourSAnti-ulcerogenic effect of methanolic extracts from Enicosanthellum pulchrum (King) Heusden against ethanol-induced acute gastric lesion in animal modelsPLoS One20149e11192525379712
- PinheiroIMLBXavierCMde SouzaADLAcanthoic acid and other constituents from the stem of Annona amazonica (Annonaceae)J Braz Chem Soc20092010951102
- ChangGJWuMHWuYCSuMJElectrophysiological mechanisms for antiarrhythmic efficacy and positive inotropy of liriodenine, a natural aporphine alkaloid from Fissistigma glaucescensBr J Pharmacol1996118157115838842417
- RahmanMMLopaSSSadikGAntibacterial and cytotoxic compounds from the bark of Cananga odorataFitoterapia20057675876116242266
- NissankaAPKarunaratneVBandaraBAntimicrobial alkaloids from Zanthoxylum tetraspermum and caudatumPhytochemistry20015685786111324918
- KhanMKiharaMOmolosoAAntimicrobial activity of Michelia champacaFitoterapia20027374474812490248
- NozakaTWatanabeFTadakiS-IMutagenicity of isoquinoline alkaloids, especially of the aporphine typeMutat Res19902402672792184353
- ChenK-SKoF-NTengC-MWuY-CAntiplatelet and vasorelaxing actions of some aporphinoidsPlanta Med1996621331368657745
- HsiehT-JLiuT-ZChernC-LLiriodenine inhibits the proliferation of human hepatoma cell lines by blocking cell cycle progression and nitric oxide-mediated activation of p53 expressionFood Chem Toxicol2005431117112615833387
- ChenC-YChenS-YChenC-HLiriodenine induces G1/S cell cycle arrest in human colon cancer cells via nitric oxide-and p53-mediated pathwayProcess Biochem20124714601468
- ChangH-CChangF-RWuY-CLaiY-HAnti-cancer effect of liriodenine on human lung cancer cellsKaohsiung J Med Sci20042036537115473647
- Kithsiri WijeratneEHatanakaYKikuchiTTezukaYLeslie GunatilakaAA dioxoaporphine and other alkaloids of two annonaceous plants of Sri LankaPhytochemistry19964217031706
- BellatiFNapoletanoCRuscitoIComplete remission of ovarian cancer induced intractable malignant ascites with intraperitoneal bevacizumab. Immunological observations and a literature reviewInvest New Drugs20102888789419936983
- SavillJFadokVCorpse clearance defines the meaning of cell deathNature200040778478811048729
- KurosakaKTakahashiMWatanabeNKobayashiYSilent cleanup of very early apoptotic cells by macrophagesJ Immunol20031714672467914568942
- ElmoreSApoptosis: a review of programmed cell deathToxicol Pathol20073549551617562483
- Van EngelandMNielandLJRamaekersFCSchutteBReutelingspergerCPAnnexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposureCytometry199831199450519
- FadokVAVoelkerDRCampbellPACohenJJBrattonDLHensonPMExposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophagesJ Immunol1992148220722161545126
- IgneyFHKrammerPHDeath and anti-death: tumour resistance to apoptosisNat Rev Cancer2002227728812001989
- SaelensXFestjensNWalleLVVan GurpMvan LooGVandenabeelePToxic proteins released from mitochondria in cell deathOncogene2004232861287415077149
- CorySAdamsJMThe Bcl2 family: regulators of the cellular life-or-death switchNat Rev Cancer2002264765612209154
- LiuXZouHSlaughterCWangXDFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosisCell1997891751849108473
- MirzaAMcGuirkMHockenberryTNHuman survivin is negatively regulated by wild-type p53 and participates in p53-dependent apoptotic pathwayOncogene2002212613262211965534
- AltieriDCValidating survivin as a cancer therapeutic targetNat Rev Cancer20033465412509766
- ZhengY-LLoffredoCAAlbergAJLess efficient g2-m checkpoint is associated with an increased risk of lung cancer in African AmericansCancer Res2005659566957316230422
- PietenpolJStewartZCell cycle checkpoint signaling: cell cycle arrest versus apoptosisToxicology200218147548112505356