
Abstract
Background
Paeonia lactiflora Pall. (PLP), a traditional Chinese herbal medicine, has been used for hepatic disease treatment over thousands of years. In our previous study, PLP was shown to demonstrate therapeutic effect on hepatitis with severe cholestasis. The aim of this study was to evaluate the antioxidative effect of PLP on alpha-naphthylisothiocyanate (ANIT)-induced cholestasis by activating NF-E2-related factor 2 (Nrf2) via phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway.
Materials and methods
Liquid chromatography-mass spectrometry (LC-MS) was performed to identify the main compounds present in PLP. The mechanism of action of PLP and its therapeutic effect on cholestasis, induced by ANIT, were further investigated. Serum indices such as total bilirubin (TBIL), direct bilirubin (DBIL), aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), γ-glutamyl transpeptidase (γ-GT), and total bile acid (TBA) were measured, and histopathology of liver was also performed to determine the efficacy of treatment with PLP. Moreover, in order to illustrate the underlying signaling pathway, liver glutathione (GSH) content and mRNA or protein levels of glutamate-cysteine ligase catalytic subunit (GCLc), glutamate-cysteine ligase modulatory subunit (GCLm), Akt, heme oxygenase-1 (HO-1), NAD(P)H/quinone oxidoreductase 1 (Nqo1), and Nrf2 were further analyzed. In addition, validation of PLP putative target network was also performed in silico.
Results
Four major compounds including paeoniflorin, albiflorin, oxypaeoniflorin, and benzoylpaeoniflorin were identified by LC-MS analysis in water extract of PLP. Moreover, PLP could remarkably downregulate serum levels of TBIL, DBIL, AST, ALT, ALP, γ-GT, and TBA, and alleviate the histological damage of liver tissue caused by ANIT. It enhanced antioxidative system by activating PI3K/Akt/Nrf2 pathway through increasing Akt, Nrf2, HO-1, Nqo1, GCLc, and GCLm expression. The putative targets network validation also confirmed the important role of PLP in activating Akt expression.
Conclusion
The potential mechanism of PLP in alleviating ANIT-induced cholestasis could to be related to the induction of GSH synthesis by activating Nrf2 through PI3K/Akt-dependent pathway. This indicates that PLP might be a potential therapeutic agent for cholestasis.
Introduction
Cholestasis, characterized by decreased bile flow and accumulation of bile acids, is one of the most common and devastating manifestations associated with a variety of hereditary and acquired liver diseases.Citation1,Citation2 This condition will ultimately lead to hepatic failure and cirrhosis without proper treatment.Citation3 Due to the serious outcome, it always brings heavy burden to patients and society all over the world.Citation4–Citation6 Cholestasis is observed in various dysfunctions, but current studies mainly concentrate on dysregulation of bile acid transporters, oxidative stress, and inflammation in hepatocytes. Among the three aspects, cholestasis associated with liver diseases is most commonly featured by the enhancement of reactive oxygen species (ROS) and reduction of antioxidant systems such as glutathione (GSH), which finally results in oxidative stress and directly induces the apoptosis of hepatocytes.
Recently, NF-E2-related factor 2 (Nrf2) has been proved to play a central role in defense against oxidative stress. Expression of many antioxidant proteins and detoxifying enzymes such as GSH, glutamate-cysteine ligase catalytic subunit (GCLc), and GCL modulatory subunit (GCLm) are primarily controlled by Nrf2.Citation7 It has been indicated that the translocation of Nrf2 can ultimately inhibit the oxidative stress via mediating the positive regulatory signal of GSH, GCLc, and GCLm.Citation8 Once the intracellular concentrations of GCLc and GCLm increase, GSH acting as the downstream signal, correspondingly reduces ROS and provides protection against oxidative stress. Given the importance of GSH in maintaining normal cell function, dysregulation in its synthesis generally worsens the disease process.Citation9,Citation10 Phosphatidylinositol 3-kinase (PI3K)/Akt pathway has been recently proved to be one of the potential upstream signaling regulators of Nrf2.Citation8 Accumulated evidence has shown that nuclear translocation of Nrf2 and GSH synthesis might be induced by phosphorylation of Akt.Citation11 Therefore, there seems to be a close relationship between the activation of GSH and PI3K/Akt/Nrf2 pathway in cholestasis.
Complementary and alternative medicine displays its unique role in the treatment of cholestasis along with the development of medical science. In the traditional Chinese medicine, the dried root of Paeonia lactiflora Pall. (PLP, Chishao in Chinese) has been used for the treatment of hepatic diseases for over thousands of years.Citation12 In our previous meta-analysis, PLP was shown to demonstrate excellent therapeutic effect on hepatitis associated with severe cholestasis.Citation6 However, the protective mechanism of PLP on cholestasis seems to be inadequate compared with clinical application. Current studies indicate that PLP might exhibit protective effect against liver diseases through antioxidation and free radicals scavenging mechanisms,Citation13 but the potential pathway still remains to be clarified. Therefore, in the current study, we aimed to investigate the role of PLP in alleviating alpha-naphthylisothiocyanate (ANIT)-induced cholestasis. To further explore its mechanism of antioxidative action in depth, we specifically investigated the PI3K/Akt and Nrf2 signaling pathways, which might provide a novel insight into improving the treatment for cholestatic liver injury ().
Figure 1 Work flow of present study.
Materials and methods
Chemicals and reagents
In this study, the dried root of PLP was used. PLP was purchased from Beijing Lvye Medicinal Materials Company (Beijing, People’s Republic of China). ANIT was purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Total bilirubin (TBIL), direct bilirubin (DBIL), aspartate transaminase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), γ-glutamyltranspeptidase (γ-GT), total bile acid (TBA), and GSH assay kits were obtained from Nanjing Jiancheng Bioengineering Institute (Nanjing, People’s Republic of China). All the other chemicals of analytical grade were purchased from commercial sources.
Identification of compounds present in PLP by ultra high performance liquid chromatography-quadruple-time of flight
The dried root of PLP was cut into small pieces, mixed, and soaked in water (1/10 w/v) for 0.5 hours; then the mixture was boiled and reflux extraction was performed two times (2 hours for the first time, 1.5 hours for the second time). The aqueous extract of PLP was decanted, filtered through six-layer gauze, and evaporated to dryness under reduced pressure. The weight ratio (w/w) of powder to raw herb was 25.8%. The powder was dissolved in purified water to obtain a final concentration of 0.02 g/mL, which was used as the injection sample.
Ultra high performance liquid chromatography- quadruple-time of flight (UHPLC-Q-TOF) was performed with the following conditions: column: 2.1×100 mm, 1.8 μm (Zorbax Eclipse Plus C18, Agilent, Santa Clara, CA, USA); injection volume: 2 μL; flow rate: 0.3 mL/min; solvents: A, H2O with 0.1% H3PO4 and B, 100% acetonitrile; gradient: 0–2 minutes, 95% A; 2–5 minutes, 85% A; 5–10 minutes, 75% A; 10–12 minutes, 70% A; 12–14 minutes, 62% A; 14–16 minutes, 20% A; 16–20 minutes, 95% A.
For operation in MS/MS mode, a mass spectrometer (MS) fitted with an orthogonal Z-spray ion interface was used for all the analyses. Ionization was achieved using electrospray technique. For analysis, the electrospray source parameters were fixed as follows: electrospray capillary voltage was 3.0 kV in negative ionization mode and 4.0 kV in positive ionization mode; the mass range was set from 80 m/z to 1,000 m/z; gas temperature was 200°C in negative ionization mode and 225°C in positive ionization mode; gas flow was 11 L/min; nebulizer was set to 35 pisg (negative) and 45 pisg (positive); sheath gas temperature was 350°C and sheath gas flow was 12 L/min; nozzle voltage was 500 V in both negative and positive modes.
Animals and treatments
All animal procedures were approved by the Animal Experiment Committee of 302 Military Hospital of People’s Liberation Army. Male Sprague Dawley rats weighing 200±20 g were obtained from the laboratory animal center of The Military Medical Science Academy of the PLA (Permission No SCXK-(A) 2012-0004). All the animals were acclimated for 1 week prior to the experiment and were kept under the same temperature (25°C±2°C) and lighting (12:12-hour light:dark cycle) conditions. Water and food were available for rats ad libitum. All the studies were performed in accordance with the guidelines of the Council on Animal Care of Academia Sinica. The animals were randomly divided into six groups with ten rats in each group. The aqueous extract of PLP was dissolved in normal saline (PLP group) and was given to rats intragastrically 80 g/kg, 40 g/kg and 20 g/kg doses respectively for 5 days. Rats in PLP groups were intragastrically treated with 60 mg/kg ANIT (dissolved in olive oil) (ANIT group) on the third day. PLP doses adopted in this study were based on preliminary experiments, and were proved to have no toxic effects.Citation12,Citation14 ANIT group was administrated with normal saline for five days and on the third day was treated with 60 mg/kg ANIT. On the third day, ANIT group was also treated with 60 mg/kg ANIT. The rats serving as negative control were administrated normal saline every day and intragastrical treatment with the vehicle (olive oil) alone on the third day. Ursodeoxycholic acid (UDCA), the positive control, was given to rats at a dosage of 60 mg/kg daily for 5 days and was given ANIT 60 mg/kg on the third day. Sample collection and liver functional assays.
Rats were sacrificed after the last administration. Blood samples were collected and centrifuged at 3,000× g for 10 minutes and serum was separated. The serum samples thus obtained were sterile, hemolysis-free, and stored at −70°C before determining the biochemical parameters. The serum levels of TBIL, DBIL, AST, ALT, ALP, γ-GT, and TBA were measured by Synergy Hybrid Reader (Biotek, Winooski, VT, USA) using assay kits, wherein assays were performed according to the manufacturer’s protocol.Citation15 Histological assessment of liver damage.
Liver tissues were excised and fixed in 10% phosphate-buffered saline (PBS)-buffered formalin. Three or four paraffin-embedded sections (4–5 μm thick) per specimen were prepared and stained with hematoxylin-eosin (HE). The stained sections were examined under Nikon microscope and analyzed by NIS-Elements (version F 4.0) software.
Measurement of GSH content in liver tissue
A portion of the liver tissue was homogenized with a buffer containing 0.15 M KCl to obtain a 1:10 (w/v) solution. The homogenates were then centrifuged at 12,000× g (4°C) for 20 minutes and the supernatants were collected. The supernatant was measured using the GSH assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, People’s Republic of China). All the steps were performed according to the manufacturer’s instructions for the determination of GSH content.Citation16
Western blot for detecting GLCc, GCLm, pAkt, and Nrf2 in liver tissue
Rat liver tissue (0.1 g) was homogenized and subsequently lysed in ice-cold lysis buffer containing 1 mM phenylmethylsulfonyl fluoride and a protease inhibitor mixture. The sample was subjected to centrifugation at 8,000× g for 10 minutes at 4°C to remove any debris. After centrifugation, the supernatant was aliquoted and stored at −80°C. These samples were used to perform Western blot assay to detect the presence of GLCc, GCLm, and pAkt. The nuclear and cytoplasmic extractions were performed using the nuclear and cytoplasmic extraction kits (Biosynthesis Biotechnology Company, Beijing, People’s Republic of China) following manufacturer’s instructions. Then, nuclear and cytoplasmic proteins were assayed by Western blot analysis for the presence of Nrf2. Fifty micrograms of protein was separated by 12% SDS–polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane. Immunodetection was performed using a rabbit anti-GCLc antibody (1:5,000), anti-GCLm antibody (1:5,000), anti-Akt antibody (1:2,000), anti-Nrf2 antibody (1:2,000), and mouse anti-GAPDH antibody (1:1,000) in a solution of 5% milk, Tris-buffered saline (TBS), and 0.05% Tween-20 to detect GCLc, GCLm, Akt, Nrf2, and GAPDH, respectively. After incubation with the appropriate peroxidase-conjugated secondary antibody, the membrane was washed in TBST (TBS with Tween 20) for 60 minutes, and the immunoreactive bands were visualized with chemiluminescence.
Quantitative RT-PCR analyses for GCLc, GCLm, Akt, Nrf2, Nqo1, and HO-1 in liver tissue
The effect of PLP on mRNA expressions of GCLc, GCLm, Akt, Nrf2, NAD(P)H/quinone oxidoreductase 1 (Nqo1), and heme oxygenase-1 (HO-1) in the liver tissue of ANIT-induced cholestasis rats were determined by quantitative reverse transcription polymerase chain reaction (RT-PCR). Total RNA was extracted from the liver tissues of each group using Trizol reagent following the manufacturer’s protocols. RNA concentration was determined by optical density measurement at 260 nm on a spectrophotometer. RNA (2 μg) was reverse-transcribed using a PrimeScript™ RT reagent kit, and 2 μL cDNA was used for the PCR reaction. List of primers used in our study are listed in . The subsequent PCR amplification was carried out by ABI Step One Plus (Applied Biosystems Inc, Carlsbad, CA, USA) PCR machine, running 40 cycles at 95°C for 5 seconds and 60°C for 60 seconds. Data were calculated for comparison through 2−ΔΔCT method.
Table 1 Primers sequences for RT-PCR
PLP putative target network validation
The ingredients of PLP were collected from TCM Database@ Taiwan (http://tcm.cmu.edu.tw/). Data about putative targets of PLP’s main ingredients were collected from Herbal Ingredients’ Targets Database (http://lifecenter.sgst.cn/hit/) and ChEMBL (https://www.ebi.ac.uk/chembl/), two well-known databases including information about the targets of herb ingredients. The targets were saved as Uniprot ID and uploaded into the David 6.7 (http://david.abcc.ncifcrf.gov/home.jsp) information tool to get the gene oncology and pathway information. The relationship among herb, ingredients, and targets were visualized by using Navigator software.
Statistical analysis
The data were expressed as the mean ± standard deviation (SD) and analyzed with the SPSS software program (version 20.0; SPSS Inc., Chicago, IL, USA). The differences between the group means were calculated by one-way analysis of variance. The differences were considered to be statistically significant when P<0.05 and highly significant when P<0.01. Principal component analysis (PCA) was also performed to analyze the comprehensive effect of seven liver function parameters.
Results
Identification of compounds present in PLP by UHPLC-Q-TOF
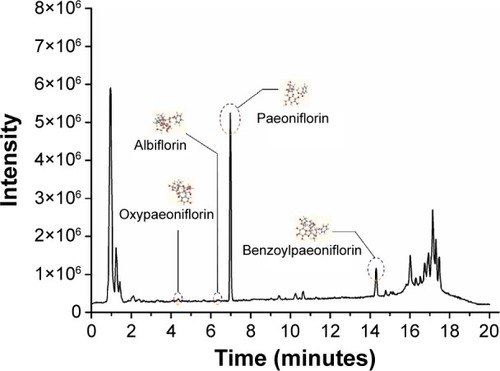
Both negative and positive electron spray (ES) modes were applied to analyze and identify the chemical components in the aqueous extract of PLP. The total current chromatogram at the positive ES mode is shown in . Four compounds including paeoniflorin, albiflorin, oxypaeoniflorin, and benzoylpaeoniflorin were detected and identified by comparing the MS fragments, characteristics of the compounds, with previous reference records and database (provided by Agilent) connected to Agilent MassHunter software ().Citation17–Citation19 The analyzed and identified compounds are listed in .
Table 2 Identification of compounds present in PLP by UHPLC-Q-TOF
Figure 2 Positive ES mode of LC–MS of PLP aqueous extract.
Abbreviations: ES, electron spray; PLP, Paeonia lactiflora Pall.; LC–MS, liquid chromatography–mass spectrometry; tR, retention time.
Effects of PLP on serum indices of liver function
The serum levels of ALT and AST were measured as the sensitive indices of liver damage. Rats administered with only ANIT displayed remarkable increase in the levels of ALT and AST (). The administration of PLP at both 80 g/kg and 40 g/kg significantly reduced the serum levels of ALT and AST, respectively, which is almost equal to the results obtained by administering UDCA. In addition, ALT, but not AST, levels were also altered by PLP at a dosage of 20 g/kg. As shown in , the serum levels of TBIL, DBIL, ALP, TBA, and γ-GT levels, which are crucial indices of cholestasis, were significantly higher in the ANIT-treated rats than in the control rats. Furthermore, the levels of ALP, TBIL, γ-GT, and TBA were remarkably decreased in rats treated with 80 g/kg, 40 g/kg, and 20 g/kg of PLP. Moreover, the serum level of DBIL decreased significantly in 80 g/kg and 40 g/kg PLP groups.
Figure 3 Effects of PLP on serum indices of liver function.
Abbreviations: PLP, Paeonia lactiflora Pall.; ANIT, alpha-naphthylisothiocyanate; UDCA, ursodeoxycholic acid; ALT, alanine aminotransferase; AST, aspartate aminotransferase; TBIL, total bilirubin; DBIL, direct bilirubin; TBA, total bile acid; ALP, alkaline phosphatase; γ-GT, γ-glutamyl transpeptidase; PCA, principal component analysis; SD, standard deviation.
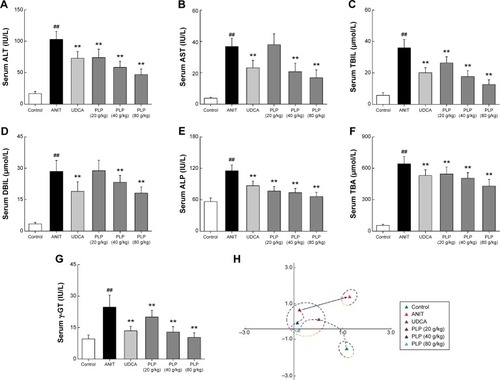
PCA was performed to analyze the comprehensive effect of seven liver function parameters. As shown in , there was a large Euclidean distance from control to ANIT group, which demonstrated the distinguished liver status and indicated the successful establishment of cholestatic hepatitis. Furthermore, the Euclidean distance from control group to other groups displayed as UDCA≈PLP 80 g/kg < PLP 40 g/kg < PLP 20 g/kg ≈ANIT. It indicated that PLP at a dosage of 80 g/kg presented a regulation effect of the comprehensive liver function similar to that shown by UDCA. In addition, PLP demonstrated weaker effect at 20 g/kg and 40 g/kg doses when compared with 80 g/kg dose.
Histological assessment of liver damage
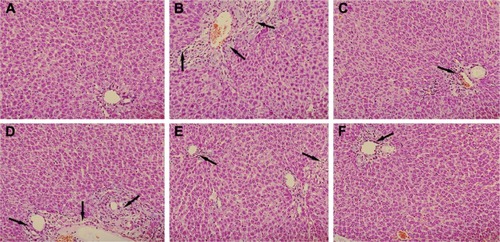
Histological evaluations provided the direct evidence for protective effect of PLP on ANIT-induced cholestasis. The liver tissue of control group exhibited normal structure with no abnormal morphological changes (). The specimens in the ANIT group showed acute infiltration with polymorphonuclear neutrophils, edema, sinusoid congestion, severe demolition or loss of the interlobular ducts, and hepatic necrosis (). Administration of UDCA and 80 g/kg PLP both exhibited a mild degree of bile duct epithelial damage and hepatocyte hydropic degeneration with less neutrophil infiltration (), which was almost similar to the control group. The specimens treated with 40 g/kg PLP displayed a moderately reduced severity of inflammatory cell infiltration and histological damage (). The liver damage in the specimens treated with 20 g/kg PLP was similar to that observed in ANIT-treated rats. PLP at 20 g/kg did not attenuate any portal tract edema, cholangitis, and bile duct epithelial damage ().
Figure 4 Effect of PLP on histological changes in the liver tissue of ANIT-induced rats.
Abbreviations: PLP, Paeonia lactiflora Pall.; ANIT, alpha-naphthylisothiocyanate; UDCA, ursodeoxycholic acid; HE, hematoxylin-eosin.
Regulation of PLP on antioxidation and PI3K/Akt/Nrf2 pathway
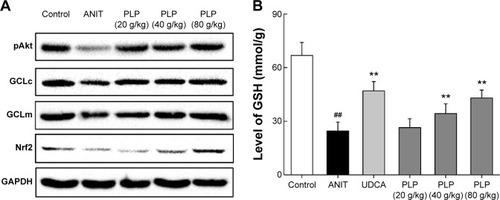
To study the role of PLP in cholestasis, the oxidative stress related to PI3K/Akt/Nrf2 pathway was explored. Results indicate that ANIT decreased the protein expressions of GCLm, GCLc, pAkt, and Nrf2, whereas PLP at 80 g/kg and 40 g/kg doses could promote the protein levels of GCLm, GCLc, p-Akt, and Nrf2 (). Furthermore, exposure to ANIT markedly decreased GSH content compared with control, suggesting the reduction of antioxidation in hepatocytes. PLP at 80 g/kg and 40 g/kg could significantly restore GSH content compared with ANIT-treated rats (). However, PLP at 20 g/kg presented no obvious change in ANIT-induced depletion of GSH.
Figure 5 GSH level and Western blot analysis of pAkt, GCLc, GCLm, and Nrf2.
Abbreviations: PLP, Paeonia lactiflora Pall.; ANIT, alpha-naphthylisothiocyanate; UDCA, ursodeoxycholic acid; Nrf2, NF-E2-related factor 2; GSH, glutathione; GCLc, glutamate-cysteine ligase catalytic subunit; GCLm, glutamate-cysteine ligase modulatory subunit; SD, standard deviation.
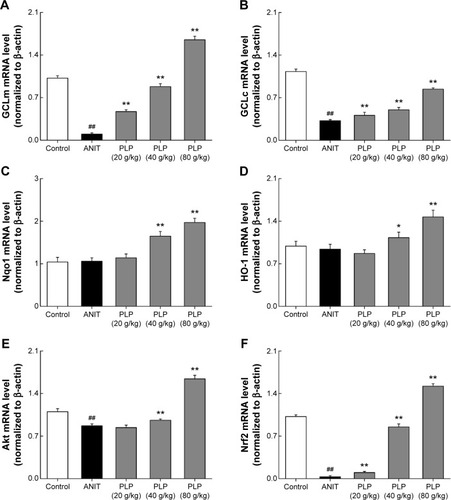
Expression of mRNAs involved in PI3K/Akt/Nrf2 signaling pathway
As shown in , we examined the effect of PLP on ANIT-reduced mRNA expressions of GCLc and GCLm. The results showed that PLP increased the mRNA expressions of GCLc and GCLm in a concentration-dependent manner. The downstream signaling of Nrf2, such as mRNA expressions of Nqo1 and HO-1, was not significantly altered by ANIT, but was upregulated by PLP at doses of 80 g/kg and 40 g/kg (). This indicated that PLP possibly affects Nqo1 and HO-1 upstream gene expression such as Nrf2. To elucidate the regulation of PI3K/Akt pathway and Nrf2, we further examined the effect of PLP on mRNA expressions of Akt and Nrf2 (). We observed that PLP could increase the mRNA expression of Nrf2 at all doses, and there was a significant increase in regulation of Akt in the group treated with PLP at doses of 80 g/kg and 40 g/kg.
Figure 6 mRNA expessions of GLCc, GLCm, Nqo1, HO-1, Akt, and Nrf2 (%).
Abbreviations: PLP, Paeonia lactiflora Pall.; ANIT, alpha-naphthylisothiocyanate; UDCA, ursodeoxycholic acid; Nrf2, NF-E2-related factor 2; GCLc, glutamate-cysteine ligase catalytic subunit; GCLm, glutamate-cysteine ligase modulatory subunit; HO-1, heme oxygenase-1; NQO1, NAD(P)H/quinone oxidoreductase 1; SD, standard deviation.
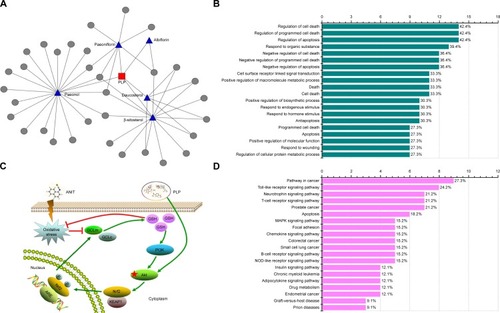
PLP putative targets network validation
Based on Herbal Ingredients’ Targets Database and ChEMBL, five main ingredients of PLP and 33 known targets of these similar ingredients as putative targets for PLP were identified (, ). As shown in , the putative targets were analyzed by David 6.7 information tool to obtain the gene annotation. demonstrates the top 20 gene functions of the putative targets. The top gene function was related to cell death or apoptosis, which occupied approximately 42.4% of the total genes. displays the top 20 pathways of the putative targets. The results indicated that cancer (27.3%), Toll-like receptor (24.2%), neurotrophin (21.2%), T-cell receptor (21.2%), and prostate cancer (21.2%) signaling pathways were relatively important of these 33 putative targets. Moreover, according to the annotation of KEGG, Akt was regulated in all the top five pathways, which indicated that Akt played an important role in PLP’s putative targets ().
Figure 7 PLP putative targets network validation and its possible mechanism.
Abbreviations: PLP, Paeonia lactiflora Pall.; ANIT, alpha-naphthylisothiocyanate; Nrf2, NF-E2-related factor 2; GSH, Glutathione; GCLc, glutamate-cysteine ligase catalytic subunit; GCLm, glutamate-cysteine ligase modulatory subunit.
Discussion
Cholestasis is characterized by intrahepatic accumulation of potentially toxic bile acids that are associated with several liver diseases as an effect of obstruction or destruction of bile ducts.Citation20 Cholestasis is one of the main features in chronically progressive hepatic disorders such as biliary atresia, primary biliary cirrhosis, and primary sclerosing cholangitis. The primary event of cholestasis has been implicated in the later development of hepatocellular injury, progressive hepatic fibrogenesis, cirrhosis, and death due to liver failure.Citation21 Accumulated evidence indicates that free radicals, oxidative stress, and lipid peroxidation are observed in cholestatic damage. Several studies further demonstrate the pro-oxidant potential of hydrophobic bile acids in hepatocytes.Citation22 It is therefore believed that oxidative stress is a decisive generating factor in the pathogenesis of cholestatic damage, and antioxidant therapy will be the recommended therapeutic strategy. ANIT is a hepatotoxicant commonly used in rodents to mimic human intrahepatic cholestasis, which acts by interrupting the bile flow and accumulating bile acids or other bile components in the liver. In ANIT-induced liver injury, the change of hepatic antioxidant defense system with enhancement of hepatic lipid peroxidation is suggested to be the primary contributor to the progress of liver damage.Citation23,Citation24 Therefore, ANIT-induced liver injury model was applied in this study to mimic the characteristics and mechanism of cholestatic hepatitis.
PLP is one of the most well-known herbs in People’s Republic of China, Korea, and Japan for more than 1,200 years. PLP consistently exhibits wide pharmacological effects such as vasodilatation of thoracic aorta;Citation25 liver protection;Citation1 antiallergic,Citation26 anti-inflammation, and immunoregulation effects.Citation27 Current studies further indicate that PLP and its active components demonstrate multiple effects mainly through the function of antioxidation.Citation28,Citation29 Paeoniflorin, one of the active compounds in PLP, is proved to attenuate apoptosis in PC12 cells via suppressing ROS-mediated PKC/NF-κB pathway.Citation28 Kim also reported that the aqueous extracts of PLP was able to protect primary cultures of rat cortical cells exposed to oxidative stress induced by H2O2.Citation29 PLP and its components show the antioxidant effect not only in neurocytes, but also in other cells such as hepatocytes. The aqueous extract of PLP can protect against CCl4-induced liver injury with oxidative damage in rats, and the hepatoprotective effects might be correlated with its antioxidation and free radical scavenger effect.Citation12 This result has also been validated by our previous research. In our former study, we first proved that the prescriptions consisting of PLP are effective against cholestatic hepatitis in clinical application.Citation6 The efficacy of paeoniflorin on cholestasis is then validated in the ANIT-induced liver injury rats, where it acts as free radical scavenger.Citation1 These studies indicate that PLP is efficient in treating cholestatic hepatitis, which might be due to its antioxidant effect. Therefore, we further carried out this study in order to illustrate the mechanism of action of PLP on cholestatic hepatitis based on our previous result.
Our study showed that the pretreatment of PLP significantly and dose-dependently decreased ANIT-induced elevation of serum ALT, AST, ALP, TBIL, DBIL, TBA, and γ-GT levels. Furthermore, histological injuries were remarkably relieved after PLP pretreatment at the dose of 80 g/kg and 40 g/kg but with weaker effect at the dose of 20 g/kg. These results indicated that PLP was effective on ANIT-induced liver injury associated with cholestasis. GSH, involved in the detoxification of chemical substances conjugated by the catalytic action of GST and extracellular transport of conjugated compounds, is one of the major cellular molecules against ROS.Citation8 GSH synthesis occurs in the cytosol of all mammalian cells via two enzymatic steps: the formation of γ-glutamylcysteine from glutamate and cysteine catalyzed by glutamate cysteine ligase (GCL) and the formation of GSH from γ-glutamylcysteine and glycine catalyzed by GSH synthase. GCLc and GCLm, the constituents of GCL, are therefore believed to play crucial roles against oxidative stress. Previously, Mitsuyoshi indicated that UDCA, the only medication approved by the FDA for the treatment of cholestasis, increased the cellular content of GSH and induction of GCL to protect the cell from oxidative damage.Citation30 According to the important role of GSH in antioxidation, we examined the effect of PLP on the expression of GSH and its synthetic enzymes. From our experiment, it was observed that PLP significantly increased GSH levels and upregulated the mRNA/protein expressions of GCLc and GCLm in a dose-dependent manner. The result was also in accordance with our former study and Wang’s reports,Citation1,Citation31 which suggested that PLP might partly contribute to protecting ANIT-induced cholestasis from oxidative damage.
Nrf2, a key transcription factor, serves as a sensor for regulating the expression of GCL and other antioxidative stress genes in response to oxidative stimulations.Citation32 Under normal conditions, Nrf2 is repressed in the cytoplasm by the actin-binding protein Keap1. Upon oxidative stress, Nrf2 dissociates from Keap1 and translocates to the nucleus to bind to antioxidant response element, which is an enhancer element that initiates the transcription of a battery of genes encoding phase-II enzymes and antioxidant enzymes.Citation33 Eminent evidence has shown that Nrf2 activation protects against intrahepatic cholestasis by regulating expression of several detoxifying and antioxidant enzymes in both BDL and ANIT models.Citation34,Citation35 Further research proves that Nrf2, which serves as the endogenous therapeutic agent, is able to positively regulate the expression of GCL subunits and glutamine synthetase in cholestasis mice. In the present study, the expression of Nrf2 in ANIT-treated rats decreased significantly compared with control rats. This result was similar to Tanaka’s former report.Citation34 In order to further investigate the downstream regulation of Nrf2, we also detected the mRNA expressions of Nqo1 and HO-1, which are the prototypical Nrf2 target genes and crucial antioxidation elements. The results demonstrated that ANIT did not significantly alter the Nqo1 and HO-1 mRNA expressions. It suggested that the cholestasis induced by ANIT was not remarkably related to dysfunction of these two genes. However, PLP could significantly promote these target genes, suggesting the potent antioxidation effect of PLP through Nrf2. In addition, our results also showed that the expression of Nrf2 increased remarkably after pretreatment with PLP. All the results indicated that Nrf2 was the important target of GCLm and GCLc in cholestasis. It also proved the positive function of PLP in increasing Nrf2 expression in cholestasis treatment.
Akt is the primary mediator of PI3K-initiated signaling pathway. It commonly acts as an antiapoptotic signaling molecule in many different cell death paradigms when it is phosphorylated and activated at residue Ser473. In recent years, it is believed that the PI3K/Akt pathway creates a survival signal against oxidative stress-induced injuries by regulating Nrf2 expression in numerous cells.Citation11,Citation36 Even in HepG2 cells, the activity of GCL subunits is proved to be stimulated by the activation of PI3K/Akt signaling accompanied with nuclear translocation of Nrf2.Citation8 These results indicate that PI3K/Akt plays an important role in regulation of Nrf2 and oxidative stress. Therefore, we further investigated the relationship between Nrf2 and PI3K/Akt signaling pathway. As shown in our result, PLP could enhance the mRNA expression of Akt and protein expression of pAkt. Moreover, the result of PLP putative targets network validation also demonstrated that Akt was the crucial target of PLP. The results revealed the involvement of PI3K/Akt pathway in regulating GSH level and Nrf2 translocation (, ).
A phytochemical analysis was performed to determine the main active components of PLP. Four representative compounds including paeoniflorin, albiflorin, oxypaeoniflorin, and benzoylpaeoniflorin were identified by UPLC-MS. Paeoniflorin is the most bioactive component among the four compounds, and its liver protection effects have been demonstrated by many studies.Citation37–Citation39 Apart from paeoniflorin, oxypaeoniflorin also shows antioxidative and anti-inflammatory activities to a little extent.Citation40 Therefore, from the aspect of chemical composition, we might speculate that the protective effect of PLP on ANIT-induced cholestasis was due to the presence of active compounds such as paeoniflorin, and oxypaeoniflorin.
All the evidences mentioned earlier indicate that PLP is able to alleviate ANIT-induced cholestasis by downregulating oxidative stress via PI3K/Akt/Nrf2 signaling pathway.
Conclusion
In summary, various biochemical tests and histological assessments have revealed that PLP could provide protection against ANIT-induced cholestasis. The potential mechanism seems to be related to the induction of GSH synthesis by activating Nrf2 through PI3K/Akt-dependent pathway. The results indicate that PLP might be a potential therapeutic agent for cholestasis.
Acknowledgments
This work was financially supported by grants from National Natural Science Foundation of China (81173571 and 81303120), The “Twelfth Five Year Plan” Foundation of China People’s Liberation Army (CWS11C164), and the major projects of the National Science and Technology (2012ZX10005010-002-002).
Supplementary materials
Figure S1 Mass-to-charge (m/z) information of four compounds.
Table S1 PLP putative targets
References
- ChenCYTCM Database@Taiwan: the world’s largest traditional Chinese medicine database for drug screening in silicoPLoS One201161e1593921253603
- HaoYeLiYeKangHongHIT: linking herbal active ingredients to targetsNucleic Acids Res201139D1055D105921097881
- KrugerFARostomROveringtonJPMapping small molecule binding data to structural domainsBMC Bioinformatics201213S1123282026
Disclosure
The authors report no conflicts of interest in this work.
References
- ZhaoYZhouGWangJPaeoniflorin protects against ANIT-induced cholestasis by ameliorating oxidative stress in ratsFood Chem Toxicol20135824224823623840
- AnwerMSRole of protein kinase C isoforms in bile formation and cholestasisHepatology20146031090109724700589
- PaddaMSSanchezMAkhtarAJBoyerJLDrug-induced cholestasisHepatology20115341377138721480339
- YangHRamaniKXiaMDysregulation of glutathione synthesis during cholestasis in mice: molecular mechanisms and therapeutic implicationsHepatology20094961982199119399914
- WeerachayaphornJLuoYMennoneASorokaCJHarryKBoyerJLDeleterious effect of oltipraz on extrahepatic cholestasis in bile duct-ligated miceJ Hepatol201460116016623978715
- MaXWangJHeXLarge dosage of chishao in formulae for cholestatic hepatitis: a systematic review and meta-analysisEvid Based Complement Alternat Med2014201432815224987427
- XuWHellerbrandCKöhlerUAThe Nrf2 transcription factor protects from toxin-induced liver injury and fibrosisLab Invest200888101068107818679376
- ArisawaSIshidaKKameyamaNUrsodeoxycholic acid induces glutathione synthesis through activation of PI3K/Akt pathway in HepG2 cellsBiochem Pharmacol200977585886619073151
- LuSCRegulation of glutathione synthesisMol Aspects Med2009301–2425918601945
- KoKYangHNoureddinMChanges in S-adenosylmethionine and glutathione homeostasis during endotoxemia in miceLab Invest200888101121112918695670
- LiCPanZXuTZhangCWuQNiuYPuerarin induces the upregulation of glutathione levels and nuclear translocation of Nrf2 through PI3K/Akt/GSK-3β signaling events in PC12 cells exposed to leadNeurotoxicol Teratol2014461925195717
- ZhaoYMaXWangJLarge dose means significant effect – dose and effect relationship of Chi-Dan-Tui-Huang decoction on alpha-naphthylisothiocyanate-induced cholestatic hepatitis in ratsBMC Complement Altern Med20151510425889085
- LiRGuoWFuZDingGZouYWangZHepatoprotective action of Radix Paeoniae Rubra aqueous extract against CCl4-induced hepatic damageMolecules201116108684869422005833
- WeiS‘Dose-Effect-Response’ Relationships of Chi-Dan-Tui-Huang decoction on Acute Cholestatic Hepatitis [dissertation]ChengduChengdu University of Traditional Chinese Medicine2012
- WuQZhangDTaoNInduction of Nrf2 and metallothionein as a common mechanism of hepatoprotective medicinal herbsAm J Chin Med201442120722124467545
- LiJPengLDuHThe Protective Effect of Beraprost Sodium on Diabetic Cardiomyopathy through the Inhibition of the p38 MAPK Signaling Pathway in High-Fat-Induced SD RatsInt J Endocrinol2014201490143725435878
- JiangMZhouMHanYIdentification of NF-κB Inhibitors in Xuebijing injection for sepsis treatment based on bioactivity-integrated UPLC-Q/TOFJ Ethnopharmacol2013147242643323524166
- ZhangHJChengYYAn HPLC/MS method for identifying major constituents in the hypocholesterolemic extracts of Chinese medicine formula ‘Xue-Fu-Zhu-Yu decoction’Biomed Chromatogr200620882182616345012
- ZhangYQianDPanYComparisons of the pharmacokinetic profile of four bioactive components after oral administration of gan-sui-ban-xia decoction plus-minus gansui and gancao drug combination in normal ratsMolecules20152059295930826007184
- Gonzalez-SanchezEFirrincieliDHoussetCChignardNNuclear receptors in acute and chronic cholestasisDig Dis201533335736626045270
- OguzSKanterMErbogaMErenogluCProtective effects of thymoquinone against cholestatic oxidative stress and hepatic damage after biliary obstruction in ratsJ Mol Histol201243215115922270828
- Fuentes-BrotoLMartínez-BallarínEMiana-MenaJLipid and protein oxidation in hepatic homogenates and cell membranes exposed to bile acidsFree Radic Res200943111080108919669996
- CuiYJAleksunesLMTanakaYGoedkenMJKlaassenCDCompensatory induction of liver efflux transporters in response to ANIT-induced liver injury is impaired in FXR-null miceToxicol Sci20091101476019407337
- OhtaYKongoMSasakiEHaradaNChange in hepatic antioxidant defense system with liver injury development in rats with a single alpha-naphthylisothiocyanate intoxicationToxicology1999139326527510647926
- JinSNWenJFWangTTKangDGLeeHSChoKWVasodilatory effects of ethanol extract of Radix Paeoniae Rubra and its mechanism of action in the rat aortaJ Ethnopharmacol2012142118819322543176
- LeeBShinYWBaeEAAntiallergic effect of the root of Paeonia lactiflora and its constituents paeoniflorin and paeonolArch Pharm Res200831444545018449501
- HeDYDaiSMAnti-inflammatory and immunomodulatory effects of paeonia lactiflora pall: a traditional chinese herbal medicineFront Pharmacol201125210
- DongHLiRYuCXuTZhangXDongMPaeoniflorin inhibition of 6-hydroxydopamine-induced apoptosis in PC12 cells via suppressing reactive oxygen species-mediated PKCδ/NF-κB pathwayNeuroscience2015285708025446358
- KimSHLeeMKLeeKYSungSHKimJKimYCChemical constituents isolated from Paeonia lactiflora roots and their neuroprotective activity against oxidative stress in vitroJ Enzyme Inhib Med Chem20092451138114019772486
- MitsuyoshiHNakashimaTSumidaYUrsodeoxycholic acid protects hepatocytes against oxidative injury via induction of antioxidantsBiochem Biophys Res Commun1999263253754210491327
- WangTZhouZXSunLXResveratrol effectively attenuates α-naphthyl-isothiocyanate-induced acute cholestasis and liver injury through choleretic and anti-inflammatory mechanismsActa Pharmacol Sin201435121527153625418378
- OkadaKShodaJTaguchiKNrf2 counteracts cholestatic liver injury via stimulation of hepatic defense systemsBiochem Biophys Res Commun2009389343143619732748
- WangGXiuPLiFXinCLiKVitamin A supplementation alleviates extrahepatic cholestasis liver injury through Nrf2 activationOxid Med Cell Longev2014201427369225126202
- TanakaYAleksunesLMCuiYJKlaassenCDANIT-induced intrahepatic cholestasis alters hepatobiliary transporter expression via Nrf2-dependent and independent signalingToxicol Sci2009108224725719181614
- AleksunesLMSlittALMaherJMNuclear factor-E2-related factor 2 expression in liver is critical for induction of NAD(P)H: quinone oxidoreductase 1 during cholestasisCell Stress Chaperones200611435636317278884
- YehYHKuoCTChangGJRosuvastatin suppresses atrial tachycardia-induced cellular remodeling via Akt/Nrf2/heme oxygenase-1 pathwayJ Mol Cell Cardiol201582849225771143
- ZhaoYMaXWangJPaeoniflorin alleviates liver fibrosis by inhibiting HIF-1α through mTOR-dependent pathwayFitoterapia20149931832725454463
- ChenMCaoLLuoYPaeoniflorin protects against concanavalin A-induced hepatitis in miceInt Immunopharmacol2015241424925479726
- ChenXLiuCLuYPaeoniflorin regulates macrophage activation in dimethylnitrosamine-induced liver fibrosis in ratsBMC Complement Altern Med20121225423237422
- ZhangMHFengLZhuMMGuJFWuCJiaXBAntioxidative and anti-inflammatory activities of paeoniflorin and oxypaeoniflora on AGEs-induced mesangial cell damagePlanta Med20137914131932323881455