
Abstract
Background
The fat mass and obesity-related (FTO) gene has a strong relationship with obesity, extreme obesity and inflammatory state, and may also be associated with food intake regulation.
Objective
The aim of the present study was to evaluate the influence of the rs9939609 single-nucleotide polymorphism of the FTO gene on appetite, ghrelin, leptin, interleukin 6 (IL6), tumor necrosis factor α (TNFα) levels and food intake of morbidly obese women.
Materials and methods
The study comprised 70 women, aged between 20 and 48 years, from Rio de Janeiro, Brazil. The participants were selected according to the body mass index between 40 and 60 kg/m2. Anthropometric and biochemical data were measured during fasting. Hormones and inflammatory data were measured before and after the participants ate an isocaloric meal. Dietary records were calculated and analyzed using a nutritional assessment program. Visual analog scales were used for behaviors of the sensations of appetite and food preferences. The FTO rs9939609 variant was genotyped using real-time polymerase chain reaction.
Results
Participants with the AA genotype had lower values of ghrelin and IL6 and higher values of leptin than those with TT and TA in the postprandial period. Comparing the plasma concentrations of ghrelin, insulin, IL6 and TNFα intragenotypes, it was observed that those with TT had decreased leptin and increased IL6 at the postprandial period. Subjects with TA showed increased postprandial IL6, and those with AA had decreased postprandial ghrelin. There was no difference in TNFα intra- and intergenotypes. The postprandial sensations of hunger were lower in AA than those with TT. There were differences between genotypes regarding ingested grams of protein by weight, cholesterol, B3, B5, B6 and B12 vitamins, and selenium potassium and sodium minerals.
Conclusion
These findings suggest that genetics may exert an influence on physiologic factors and might alter eating behavior.
Introduction
Today, the prevalence of obesity continues to increase and is one of the most challenging public health problems. Recent data show that the percentage of obese people has doubled in more than 70 countries between 1980 and 2015.Citation1 Currently, more than 1.9 billion individuals are overweight, of which more than 600 million are obese.Citation2 Furthermore, the medical costs associated with obese individuals is ~30% greater than that of normal-weight people.Citation3
In Brazil, ~32 million adults had a body mass index (BMI) indicative of obesity in 2015.Citation4 Between 2013 and 2014, our country was ranked fifth in the world in terms of obesity.Citation5,Citation6 Recently, the Risk and Protective Factors Surveillance Survey for Chronic Diseases Telephone Survey indicated an increase in obesity in Brazil by 60% in the past 10 years, from 11.8% in 2006 to 18.9% in 2016, that is, almost one in five Brazilians are obese.Citation7 The prevalence for morbid obesity in the population is low (0.81%), but led to costs totaling $64.2 million in 2011, which amounts to almost a quarter of the costs attributable to obesity in the public health system.Citation8
Body weight gain in the population is due to changes in dietary patterns and lifestyle, with increased energy intake and decreased physical activity. These changes appear to be the major factors leading to the increase of the global obesity epidemic in recent decades. However, there are clear differences in susceptibility between individuals or communities.Citation9,Citation10
The contribution of genetics to the development of obesity has been extensively investigated in genome-wide association studies, where the most common forms of obesity are polygenic, with the discovery of at least 77 loci associated with this diseaseCitation11 and 32 loci with a strong association in the increase of BMI.Citation12 First described in 2007, it was demonstrated by bioinformatics tools that the fat mass and obesity-related (FTO) gene encodes a protein of the superfamily Fe (II) 2-oxoglutarate-dependent dioxygenases, with high similarity to the sequence of the DNA repair enzyme of Escherichia coli AlkB and its eukaryotic homologs AlkB 2 and 3 (ABH2 and 3). This gene also encodes a RNA adenosine demethylase, where the main substrate is N6-methyladenosine (m6A).Citation13 The enzyme m6A RNA demethylase influences nucleic acid demethylation, which is important for epigenetic regulation (modifications of chromatin and DNA that activate or inhibit certain genes).Citation14,Citation15 The FTO gene is located on the long arm of chromosome 16 on position 12.2 and consists of nine exons, occupying an area of >400 kb.Citation14
The FTO gene was recently described and is strongly associated with obesity,Citation14 extreme obesityCitation11 and inflammatory state,Citation16 and may also be associated with food intake regulation.Citation17FTO polymorphism has been studied in recent years and has been indicated as a predictor of obesity in several populations,Citation18–Citation20 where about 50% of the world population are the carriers of the “risk” allele.Citation21 These studies indicated the association of FTO gene variants with food intake, raising the hypothesis of influence on hunger and satiety regulationCitation17,Citation22 probably related to the postprandial plasma concentrations of leptin and ghrelin hormones.Citation23
The most studied FTO single-nucleotide polymorphism (SNP) is rs9939609, which comprises the A and T alleles, where the risk allele A seems to be directly related to increased accumulation of body fat, especially when presented as an AA homozygote.Citation18 In a pilot study published recently in Nigeria, rs9939609 was studied in a group of 103 obese and 98 controls, with mean age of 20 years. The results suggested that factors such as physical inactivity, high energy intake and sitting time had a mediating effect on the FTO gene variant for BMI, and that these variables may be playing an important role in the development and worsening of obesity.Citation24
Therefore, the aim of the present study was to evaluate the influence of rs9939609 on appetite, ghrelin, leptin, interleukin 6 (IL6), tumor necrosis factor α (TNFα) and food intake of morbidly obese women in Rio de Janeiro, Brazil.
Materials and methods
Study group: subjects
The study comprised 70 women, aged between 20 and 48 years, from Rio de Janeiro, Brazil. The participants were selected according to BMI (between 40 and 60 kg/m2) and the presence of obesity for at least 5 years. We excluded patients with renal failure, those with congestive heart failure, those diagnosed with dysgeusia, those who were in treatment for cancer, pregnant, those with a diagnosis of hypothyroidism or hyperthyroidism, those under the use of corticosteroids and medicines to lose weight or that influence the sensations of hunger and satiety (sibrutramin, orlistat, fluoxetine, topiramate and liraglutide), those in menopause, those who underwent surgery in the gastrointestinal tract, those who did not complete all of the steps described above, or who presented some intercurrence previously listed during the study. All participants provided written informed consent to participate in this study, and the protocol was performed according to the Declaration of Helsinki (1964). This study was approved by the Ethics and Research Committee of the University Hospital Clementino Fraga Filho, under the protocol CAAE no. 845.537 and registered in the Clinical Trials under the number NCT02598037.
Clinical measurements
The anthropometric data collected were body weight in kilograms, height, BMI, waist circumference (WC) and hip circumference (HC). The WC was measured at the midpoint between the lower margin of the last palpable rib and the top of the iliac crest. HC was measured around the widest portion of the buttocks.Citation25 Waist-to-hip ratios were calculated by dividing WC by the HC.Citation25
Serum samples were collected after 12 hours of an overnight fasting. Triglycerides (TGs), total cholesterol, high-density lipoproteins (HDLs), low-density lipoproteins (LDLs), very low-density lipoproteins (VLDLs), glucose and insulin fasting were measured. Ghrelin, leptin, IL6 and TNFα plasma were collected in tubes containing ethylenediaminetetraacetic acid and Pefabloc® (specific protein inhibitor), before and after the participants ate an isocaloric meal. The composition of the meals was elaborated with the same proportions of macronutrients (56% carbohydrates, 18% proteins and 26% lipids) and the same volume (350 mL) for each volunteer.
Dietary intake and physical activity
Three nonconsecutive 3-day dietary records were filled in, reporting two typical days (weekdays) and one atypical day (weekend or holiday).Citation26 Dietary records were calculated and analyzed using the nutritional assessment program AVANU-TRI® version 4.0.
The International Physical Activity Questionnaire – short version was completed, adapted to the Portuguese language of Brazil.Citation27 For the classification of the habitual practice of physical activity, the scores were considered: 1) sedentary, 2) insufficiently active, 3) active and 4) very active.
Assessment of binge eating and behaviors of sensations of appetite and food preferences
The binge eating scale was completed, adapted to the Portuguese language of Brazil.Citation28 For the classification of the periodic binge eating, the following scores were considered: ≤17 for without binge eating, 18–26 moderate binge eating and ≥27 severe binge eating.
Visual analog scales (VASs) were used for behaviors of the sensations of appetite and food preferences.Citation29 These sensations were assessed using a 10 cm VAS and were applied at intervals of 30 minutes during the study.
DNA extraction and genotyping
DNA was extracted from whole blood using a commercial DNA extraction kit (Invitrogen™ PureLink™ Genomic DNA). The FTO variant rs9939609 was genotyped using real-time polymerase chain reaction with a TaqMan® assay (Thermo Fisher Scientific, Waltham, MA, USA). Amplification was performed in the Step One Plus™ and the genotypes were identified by SDS 2.3 software. Negative controls (all components excluding DNA) were included in all plates.
Statistical analysis
The Kolmogorov–Smirnov test was used to verify the distribution of variables. Data with non-Gaussian distribution were presented as median and quartile interval. The Kruskal–Wallis tests were used to analyze the data between groups, Wilcoxon–Mann–Whitney for analyses of paired and unpaired data and chi-square to evaluate the association between variables. Friedman test was used to compare the distributions of the quantitative variables and to verify that there was a significant variation in TT, TA and AA over the postoperative period (30–180 minutes), for hunger, satiety, gastric fullness, desire to eat savory and appetizer in those who presented differences between genotypes. All results were analyzed with the statistical analysis program SPSS version 21.0, considering significant values of P<0.05.
Results
In this present study, we analyzed the FTO rs9939609 polymorphism in our sample. The calculation of the Hardy–Weinberg equilibrium was performed, and it showed that genotypes were in equilibrium (χ 2=0.045; P=0.832).Citation30,Citation31 Our sample was stratified into groups according to genotype distribution, in which the frequencies were TT (33%), TA (50%) and AA (17%). Moreover, the allele frequencies were T (58%) and A (42%). Low physical activity was observed in 57% of women with TT genotype, 69% with TA genotype and 75% with AA genotype.
Our results showed that women carrying the AA genotype have a higher level of preprandial insulin. There were no differences in other variables among genotypes ().
Table 1 Anthropometric and biochemical characteristics of the population
The effects of rs9939609 on hormone levels were analyzed in fasting and postprandial states. shows that there was a difference between genotypes for ghrelin, leptin and IL6, in which the AA had lower values of ghrelin and IL6 and higher values of leptin than TT and TA in the postprandial period.
Table 2 Hormone and cytokine data of the population
Comparing the plasma concentrations of ghrelin, insulin, IL6 and TNFα intragenotypes, it was observed that TT had decreased leptin and increased IL6 during the postprandial period. TA showed increased postprandial IL6 and AA decreased postprandial ghrelin. There was no difference in TNFα intra- and intergenotypes.
Considering the results found for the hormones previously referred to, it is important to complement with data related to the sensations of hunger and satiety, as well as food intake behavior.
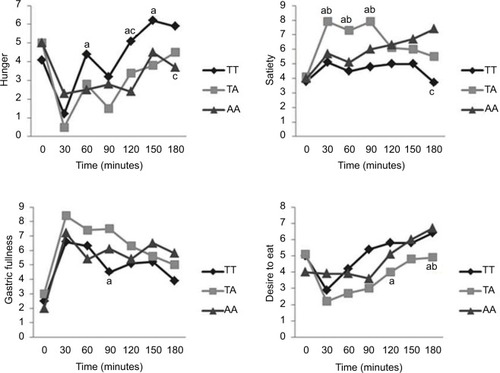
Our results showed that hunger was higher at 60, 120 and 150 minutes in TT than in TA. Hunger in AA was lower than TT at 120 and 180 minutes. Satiety was higher at 30, 60 and 90 minutes in TA than in TT and AA. At 180 minutes, AA shows higher satiety than TT. Gastric fullness was lower at 90 minutes in TT than in TA. Desire to eat was lower in TA at 120 minutes than in TT and at 180 minutes than in TT and AA ().
Figure 1 Behavior of sensations of appetite and satiety by genotype in the pre- and postprandial periods.
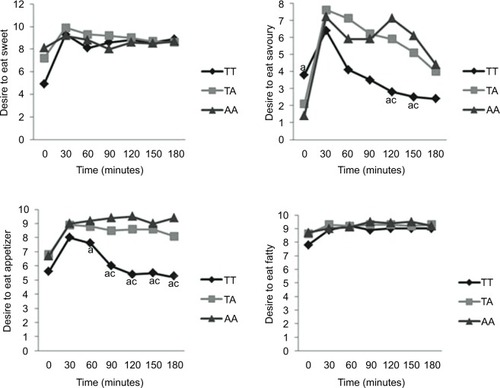
Desire to eat savory was lower at preprandial and postprandial 30, 120 and 150 minutes in TT than in TA and AA. In the postprandial period, desire to eat appetizer was lower at 60 minutes in TT than in TA and lower at 90, 120, 150 and 180 minutes than in TA and AA. There is no difference between genotypes for desire to eat sweet and fatty ().
Figure 2 Behavior of food preferences by genotype in the pre- and postprandial periods.
Additionally, it was observed that women carrying TT and TA genotypes were considered without binge eating and AA genotype with moderate binge eating according to the binge eating scores 14, 16 and 20, respectively.
Finally, 3-day dietary records were taken in order to analyze whether rs9939609 influences food intake of morbidly obese subjects. There were differences between genotypes regarding ingested grams of protein per kilogram of body weight (PTNg/kg BW); cholesterol; B3, B5, B6 and B12 vitamins and selenium potassium and sodium minerals. TT genotype consumed less PTNg/kg BW, B3 and B12 vitamins and potassium than TA and AA genotypes. Subjects carrying the AA genotype consumed larger quantities of cholesterol, B5 and B6 vitamins and selenium and sodium than those with TT and TA genotypes. There is no difference in the food intake of the other parameters ().
Table 3 Food intake data of the population expressed in median and quartile
Discussion
The purpose of this study was to investigate the hunger and satiety sensations, the release of hormones involved in this regulation, inflammatory cytokines and food intake in the different genotypes of FTO rs9939609 polymorphism. The main findings of the present study were a reduction of postprandial ghrelin in AA and higher postprandial ghrelin in TT and TA, compared to AA; increased postprandial leptin in AA, a decrease in this hormone in TT and higher postprandial leptin in AA than in TT; higher sensation of hunger in TT and greater sensation of satiety in AA during the postprandial period; desire to eat an appetizer during the postprandial period in individuals with risk allele (A) and lower protein intake per kilogram of weight in the TT genotype and higher consumption of cholesterol and sodium in AA.
The A allele proportion of the FTO rs9939609 reported in this study is in agreement with earlier studies.Citation18,Citation22 The research is contradictory in terms of differences in the anthropometric parameters. While several studies have shown an increase in body weight, waist and HCs, and especially in BMI in individuals with the risk allele,Citation10,Citation12,Citation15,Citation19,Citation24 other authors do not present differences between the genotypes,Citation22,Citation32 similarly to the present study, which can be explained by the population selection, that was homogeneous regarding these parameters.
No differences were found between investigated rs9939609 SNP biochemical parameters (cholesterol total, LDL, HDL, VLDL, TG, glucose and Homeostatic Model Assessment of Insulin Resistance). However, it was observed that insulin levels were higher in AA than in TT and TA genotypes. Different findings in the literature for insulin have been cited. For example, De Luis et al found an increased release of insulin in the TT genotypeCitation33 and Shahid et al in the AA genotype, similar to our study.Citation34
Most of the women in this study were considered as inactive, which was more prevalent in women who have the risk allele, corroborating with other studies.Citation24,Citation35 Although the prevalence of inactive women is high in the AA genotype, studies show that there was no relation between FTO and the level of physical activity in adults,Citation23,Citation36 suggesting that the presence of the risk allele is not influencing the level of physical activity in these women. On the other hand, studies have shown that an obesity-increasing effect may be attenuated in ~30% of individuals who are physically active.Citation32,Citation36 For example, it has been shown that people who carry the A allele benefit from practicing physical activity.Citation36
Binge eating disorder is associated with current morbid obesity (BMI≥40 kg/m2), and it is observed in over 40% of such individuals.Citation37 Some studies evaluating the relationship between FTO gene polymorphism and the presence of eating disorders show the role of the rs9939609 SNP in satiety responsiveness,Citation38 which could be explained by high FTO expression in the hypothalamic regions of the brain associated with appetite regulation.Citation39 In addition, a more recent study of 250 lean subjects with eating disorders found that the presence of the risk allele was associated with binge eating behavior.Citation40 Interestingly, it was observed in the present study that patients with the AA genotype presented moderate binge eating according to the binge eating score.
In the literature, it has been established that ghrelin affects appetite and food intake. However, researches have shown that polymorphism may influence the release of this hormone in patients with obesity. The association between rs9939609 and ghrelin secretion is still unknown. Arrizabalaga et al observed that ghrelin at the baseline showed no difference between rs9939609 genotypes, similar to our findings.Citation41 Nevertheless, there was a decrease in ghrelin in the postprandial period, when compared to the release of this hormone pre- and postprandially in our morbidly obese population. The opposite ghrelin pattern was observed in a similar study.Citation42
This study evaluated pre- and postprandial leptin, showing a decrease of this hormone in the TT genotype 3 hours after an isocaloric meal. Some studies evaluating leptin in the baseline show that there is no difference between genotypes presenting the risk allele,Citation33,Citation43,Citation44 corroborating with our findings. Otherwise, other studies observed a positive association of rs9939609 with leptin.Citation34,Citation41,Citation45,Citation46
No differences were found in the literature on inflammatory cytokines between genotypes with or without the risk allele in the baseline.Citation33,Citation43,Citation45,Citation46 Our findings showed increased IL6 in TT and TA, and TT presented higher IL6 than AA. Among all adipokines related to inflammatory processes, IL6, TNFα, leptin and adiponectin have received special attention in the literature. Leptin synthesis is stimulated by TNFα and IL6, contributing to satiety and maintaining the inflammatory state.Citation16
Studies have shown that carriers of the rs9939609 A genotype have altered postprandial satiety levels as recorded by VAS completed periodically.Citation42,Citation47,Citation48 While some studies have shown lower postprandial fullnessCitation47 and higher postprandial appetite levels in AA homozygotes compared to TT homozygotes for rs9939609,Citation42 our results showed the opposite. The TT genotype women had elevated hunger and decreased satiety, agreeing with the study carried out by Den Hoed et al.Citation48 Surprisingly, fullness was lower in the TT genotype, and desire to eat an appetizer was higher in women with the risk allele throughout the postprandial period.
This is the first time an analysis of the food intake, among morbidly obese individuals, is performed in detail, showing the values of carbohydrates, fibers, proteins, saturated and unsaturated fats, dietary cholesterol, vitamins and minerals. Participants with the TT genotype showed lower consumption of protein per kilogram of body weight and increased consumption of cholesterol and sodium, suggesting higher intake of food from animal products, despite there being no differences in protein (%) and total fat intake (%).
Although the presence of polymorphism in rs9969309 seems to contribute to the alteration of appetite and food intake, several articles in the literature are controversial regarding the energy and macronutrients intake, and these divergences may be explained by inaccuracies of measuring dietary intake in the obese. While most of the research does not show differences in energy and macronutrient intake between genotypes,Citation23,Citation24,Citation33,Citation47,Citation49,Citation50 some authors note a high-energy intake in individuals carrying the risk allele.Citation22,Citation32,Citation51 However, it remains to be confirmed which components of food intake are predominantly FTO targets.
In this study, we used a well-characterized cohort of participants. Furthermore, we analyzed the role of the genetic variant on hormone release and the feelings of hunger and satiety, as well as eating habits and physical activity in a population of Brazilian women with morbid obesity, who tend to be more difficult to recruit. However, limitations should also be observed: 1) this study was carried out in a group with a low level of schooling, which may influence the lack of understanding of certain answers; 2) many participants live in poorer communities and, therefore, may have increased difficulty accessing certain services (e.g., supermarkets, gyms and hospitals); and 3) only one FTO SNP was evaluated in this study, and it is not possible to clarify whether this gene has a direct relationship with feeding behavior.
Conclusion
The presence of the risk allele (A) occurred in almost half of the women with morbid obesity, and this risk allele increases the prevalence of low physical activity. AA homozygotes did not completely affect dietary intake, but dietary sodium and cholesterol intake was high in this group, deserving further investigation in relation to nutritional quality of the diet.
The levels of ghrelin decreased postprandially in women with AA genotypes, and these findings corroborated with the sensations of lower hunger observed in this group, as occurred in the TT genotype, where leptin levels increased in the postprandial period and the feeling of satiety was lower.
An interesting finding was in relation to the desire to eat an appetizer throughout the postprandial period – every 30 minutes – in women with the AA genotype and the presence of moderate binging. It is suggested that these women had the urge to eat all the time, even with the release of hormones involved in hunger and satiety. Additionally, the presence of the risk allele may interfere both in physiologic responses and in eating behavior in cases of morbid obesity.
These findings suggest that genetics may exert an influence on physiologic factors and might alter eating behavior. However, further clarification involving other genes, environmental factors and lifestyle is needed to better understand all the organic complexity involved in extreme obesity.
Acknowledgments
We thank the students of scientific initiation and, in particular, the volunteers. This work was supported by grants from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Disclosure
The authors report no conflicts of interest in this work.
References
- GBD 2015 Obesity Collaborators Afshin A Forouzanfar MH Reitsma MB Health effects of overweight and obesity in 195 Countries over 25 years N Engl J Med 2017 377 1 13 27 28604169
- WHO. World Health Organization Obesity and overweight 2016 Available from: http://www.who.int/mediacentre/factsheets/fs311/en/ Accessed June 26, 2017
- Withrow D Alter DA The economic burden of obesity worldwide: a systematic review of the direct costs of obesity Obes Rev 2011 12 2 131 141 20122135
- IMHA. Institute for Metrics and Health Assessment Available from: http://www.OverweightandObesityViz/ 2015 Accessed June 17, 2017
- Ng M Fleming T Robinson M Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: a systematic analysis for the Global Burden of Disease Study 2013 Lancet 2014 384 9945 766 781 24880830
- Cesare MD Bentham Jm Stevens GA Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants Lancet 2016 387 10026 1377 1396 27115820
- BRASIL VIGITEL – Fatores de risco e proteção para doenças crônicas não transmissíveis obtidos por inquérito telefônico 2016 Available from: http://portalarquivos.saude.gov.br/images/pdf/2017/junho/07/vigitel_2016_jun17.pdf Accessed June 16, 2017
- de Oliveira ML Santos LM da Silva EN Direct healthcare cost of obesity in Brazil: an application of the cost-of-illness method from the perspective of the public health system in 2011 PLoS One 2015 10 4 e0121160 25830909
- Swinburn BA Sacks G Hall KD The global obesity pandemic: shaped by global drivers and local environments Lancet 2011 378 9793 804 814 21872749
- Xia Q Grant SFA The genetics of human obesity Ann N Y Acad Sci 2013 1281 1 178 190 23360386
- Fall T Ingelsson E Genome-wide association studies of obesity and metabolic syndrome Mol Cell Endocrinol 2014 382 1 740 757 22963884
- Speliotes EK Willer CJ Berndt SI Association analyses of 249,796 individuals reveal eighteen new loci associated with body mass index Nat Genet 2010 42 11 937 948 20935630
- Jia G Fu Y Zhao X N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO Nat Chem Biol 2011 7 12 885 887 22002720
- Gerken T Girard CA Tung Y-CL The obesity-associated FTO gene encodes a 2-oxoglutarate–dependent nucleic acid demethylase Science 2007 318 5855 1469 1472 17991826
- Ben-Haim MS Moshitch-Moshkovitz S Rechavi G FTO: linking m6A demethylation to adipogenesis Cell Res 2015 25 1 3 4 25475057
- Fruhbeck G Gomez-Ambrosi J Muruzabal FJ Burrell MA The adipocyte: a model for integration of endocrine and metabolic signaling in energy metabolism regulation Am J Physiol Endocrinol Metab 2001 280 6 827 847
- Fredriksson R Hagglund M Olszewski PK The obesity gene, FTO, is of ancient origin, up-regulated during food deprivation and expressed in neurons of feeding-related nuclei of the brain Endocrinology 2008 149 5 2062 2071 18218688
- Frayling TM Genome-wide association studies provide new insights into type 2 diabetes aetiology Nat Rev Genet 2007 8 9 657 662 17703236
- Scuteri A Sanna S Chen WM Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits PLoS Genet 2007 10 1 43 47
- Illangasekera YA Kumarasiri RP Fernando DJ Dalton CF Association of FTO and near MC4R variants with obesity measures in urban and rural dwelling Sri Lankans Obes Res Clin Pract 2016 10 Suppl 1 S117 S124 26948330
- Gulati P Yeo GS The biology of FTO: from nucleic acid demethylase to amino acid sensor Diabetologia 2013 56 10 2113 2121 23896822
- Speakman JR Rance KA Johnstone AM Polymorphisms of the FTO gene are associated with variation in energy intake, but not energy expenditure Obesity (Silver Spring) 2008 16 8 1961 1965 18551109
- Speakman JR The “Fat Mass and Obesity Related” (FTO) gene: mechanisms of impact on obesity and energy balance Curr Obes Rep 2015 4 1 73 91 26627093
- Oyeyemi BF Ologunde CA Olaoye AB Alamukii NA FTO gene associates and interacts with obesity risk, physical activity, energy intake, and time spent sitting: pilot study in a Nigerian population J Obes 2017 2017 3245270 28607773
- WHO. World Health Organization Waist Circumference and Waist-Hip Ratio 2011 Available from: http://apps.who.int/iris/bitstream/10665/44583/1/9789241501491_eng.pdf Accessed August 7, 2017
- Sampaio LR Silva MdCMd Roriz AKC Leite VR Inquérito Alimentar [Food Inquiry] Sampaio LR Avaliação Nutricional 1 EDUFBA 2012 Portuguese
- Matsudo S Araujo T Matsudo V Questionário Internacional de Atividade Física (IPAQ): Estudo de validade e reprodutibilidade no Brasil [International Physical Activity Questionnaire (IPAQ): Study of validity and reproducibility in Brazil] Revista Brasileira de Atividade Física e Saúde 2001 6 2 5 18 Portuguese
- Freitas S Lopes CS Coutinho W Appolinario JC Tradução e adaptação para o português da Escala de Compulsão Alimentar Periódica Revista Brasileira de Psiquiatria 2001 23 4 215 220 Portuguese
- Flint A Raben A Blundell JE Astrup A Reproducibility, power and validity of visual analogue scales in assessment of appetite sensations in single test meal studies Int J Obes Relat Metab Disord 2000 24 1 38 48 10702749
- Hardy GH Mendelian proportions in a mixed population Science 1908 28 706 49 50 17779291
- Weinberg W Über den Nachweis der Vererbung beim Menschen 64 Jahresh Wuertt Ver vaterl Natkd 1908
- Loos RJ Yeo GS The bigger picture of FTO – the first GWAS-identified obesity gene Nat Rev Endocrinol 2014 10 1 51 61 24247219
- De Luis DA Aller R Izaola O Primo D Romero E Association of the rs9939609 gene variant in FTO with insulin resistance, cardiovascular risk factor and serum adipokine levels in obese patients Nutr Hosp 2016 33 5 573 27759977
- Shahid A Rana S Saeed S Imran M Afzal N Mahmood S Common variant of FTO gene, rs9939609, and obesity in Pakistani females Biomed Res Int 2013 2013 324093 24102053
- Muc M Padez C Manco L Influence of physical activity on the association between the FTO variant rs9939609 and adiposity in young adults Am J Human Biol 2015 27 5 734 738 25809737
- Kilpelainen TO Qi L Brage S Physical activity attenuates the influence of FTO variants on obesity risk: a meta-analysis of 218,166 adults and 19,268 children PLoS Med 2011 8 11 e1001116 22069379
- Hudson JI Hiripi E Pope HGJr Kessler RC The prevalence and correlates of eating disorders in the National Comorbidity Survey Replication Biol Psychiatry 2007 61 3 348 358 16815322
- Wardle J Carnell S Haworth CMA Farooqi IS O’Rahilly S Plomin R Obesity associated genetic variation in FTO is associated with diminished satiety J Clin Endocrinol Metab 2008 93 9 3640 3643 18583465
- Jonassaint CR Szatkiewicz JP Bulik CM Specific common variants of the obesity-associated FTO gene are not associated with psychological and behavioral eating disorder phenotypes Am J Med Genet 2011 156 4 454 461
- Castellini G Franzago M Bagnoli S Fat mass and obesity-associated gene (FTO) is associated to eating disorders susceptibility and moderates the expression of psychopathological traits PLoS One 2017 12 3 e0173560 28282466
- Arrizabalaga M Larrarte E Margareto J Maldonado-Martin S Barrenechea L Labayen I Preliminary findings on the influence of FTO rs9939609 and MC4R rs17782313 polymorphisms on resting energy expenditure, leptin and thyrotropin levels in obese non-morbid pre-menopausal women J Physiol Biochem 2014 70 1 255 262 24307561
- Karra E O’Daly OG Choudhury AI A link between FTO, ghrelin, and impaired brain food-cue responsivity J Clin Invest 2013 123 8 3539 3551 23867619
- Olza J Ruperez AI Gil-Campos M Influence of FTO variants on obesity, inflammation and cardiovascular disease risk biomarkers in Spanish children: a case-control multicentre study BMC Med Genet 2013 14 123 24289790
- Labayen I Margareto J Maldonado-Martin S Independent and combined influence of the FTO rs9939609 and MC4Rrs17782313 polymorphisms on hypocaloric diet induced changes in body mass and composition and energy metabolism in non-morbid obese premenopausal women Nutr Hosp 2015 31 5 2025 2032 25929371
- Zimmermann E Skogstrand K Hougaard DM Influences of the common FTO rs9939609 variant on inflammatory markers throughout a broad range of body mass index PLoS One 2011 6 1 e15958 21246032
- de Luis DA Aller R Conde R Relation of the rs9939609 gene variant in FTO with cardiovascular risk factor and serum adipokine levels in morbid obese patients Nutr Hosp 2012 27 4 1184 1189 Spanish 23165561
- Douglas A Yaqoob P Givens DI Reynolds CK Minihane AM The impact of obesity-related SNP on appetite and energy intake Br J Nutr 2013 110 6 1151 1156 23433430
- den Hoed M Westerterp-Plantenga MS Bouwman FG Mariman EC Westerterp KR Postprandial responses in hunger and satiety are associated with the rs9939609 single nucleotide polymorphism in FTO Am J Clin Nutr 2009 90 5 1426 1432 19793853
- Gustavsson J Mehlig K Leander K FTO gene variation, macronutrient intake and coronary heart disease risk: a gene-diet interaction analysis Eur J Nutr 2016 55 1 247 255 25644180
- Qi Q Kilpeläinen TO Downer MK FTO genetic variants, dietary intake and body mass index: insights from 177 330 individuals Hum Mol Genet 2014 23 25 6961 6972 25104851
- Church C Moir L McMurray F Overexpression of Fto leads to increased food intake and results in obesity Nat Genet 2010 42 12 1086 1092 21076408