
Abstract
Background
Diabetic patients have lower capacity to metabolize drugs in comparison to normal people. Therefore, the present study aimed to investigate the alterations in gene expression of drug and arachidonic acid metabolizing cytochrome p450s (cyp450s) in the livers of controlled (CDM) and uncontrolled (UDM) insulin-dependent diabetic mice.
Methods
Balb/c mice were treated with single dose of streptozocin (240 mg/kg) to induce diabetes and compared with control group, which was treated with citric buffer (pH =4.5). After 3 days, the blood glucose level was measured to confirm the induction of diabetes. Normalization of blood glucose level in diabetic mice was achieved after 0.1 mL/kg Mixtard® insulin therapy for more 5 days. Then, the mice livers were isolated to extract RNA and convert it to cDNA. The gene expression of 14 genes, which play a major role in drug and arachidonic acid metabolism, were measured using quantitative real-time polymerase chain reaction technique.
Results
It was found that the gene expression was downregulated (ANOVA test, P-value <0.05) in the livers of UDM mice. The most downregulated genes were cyp4a12, cyp1a2, and slc22a1 with more than 10-fold reduction. The livers of CDM mice showed significantly (P-value <0.05) higher levels of mRNA than UDM mice, but still lower than the non-diabetic mice.
Conclusion
This study concluded that hepatic gene expression of drug metabolizing and arachidonic acid- cyp450 enzymes is reduced in insulin-dependent diabetic mice, which can explain, at least in part, the variation in drug and fatty acid metabolism between normal and diabetic patients.
Introduction
Diabetes mellitus (DM) is an endocrine disorder, which elevates blood glucose level.Citation1 The prevalence of DM worldwide is high, including Middle Eastern countries. It is found that the prevalence of diabetes in Jordan and Syria is 17.1% and 15.6%, respectively.Citation2,Citation3
Diabetes may lead to micro- and macrovascular damage to several organs if it is uncontrolled.Citation4 It affects heart, kidney, eyes, nerves, blood vessels, immune, and liver, which is the main site of drug metabolism.Citation5–Citation7
Diabetic patients are at high risk of developing liver failure, steatosis, and cancer.Citation8,Citation9 In addition, DM was reported to alter the hepatic capacity toward drug metabolism. Irshaid et al, found that insulin-dependent diabetic patients have lower capacity to metabolize the antibiotic dapsone than non-insulin-dependent diabetic and healthy volunteers.Citation10 In addition, the amount of hepatic mRNA, protein, and enzymatic activity of human CYP/CYP450 (CYP450) 3A4 and UDP- glucuronosyltransferase (UGT) 2B7 were reduced in diabetic patients.Citation11,Citation12
In diabetic rat models using streptozotocin and alloxan, it has been observed that the metabolism of aminopyrine and CYP450 content were suppressed and insulin therapy could normalize the hepatic drug metabolism.Citation13
The role of arachidonic acid-CYP450 metabolizing enzymes, in the liver, is not fully understood.Citation14 However, it is found that accumulation of fats in the liver is associated with reduction of hepatic CYP4A expression, which indicated a protective role of CYP4A enzyme through metabolism of different fatty acids, such as arachidonic and lauric acid.Citation15 Although the gene expression and enzyme activity of arachidonic acid metabolizing hydroxylase and epoxygenase were altered in different organs among cardiovascular disease patients,Citation16 there is limited information regarding arachidonic acid-cyp450 pathway in the liver of diabetic patients. It is reported that CYP4A gene expression was upregulated in the liver of high-fat feed type II diabetic mice model.Citation17
The gene expression profile of Phase I and II drug- and arachidonic acid-metabolizing enzymes were not fully investigated in the livers of insulin-dependent diabetic patients. Alterations in the gene expression of these enzymes may influence the pharmacokinetics and hence the drug response and hepatotoxicity among diabetic patients.Citation18 Therefore, the present study aimed to analyze the gene expression of Phase I and II drug- and arachidonic acid-metabolizing enzymes in controlled and uncontrolled insulin-dependent diabetic mice models.
Methods
Chemicals
Streptozocin, citric acid, sodium citrate, isopropyl alcohol, and 75% ethanol were obtained from Sigma-Aldrich (St. Louis, MO, USA). Human insulin Mixtard®, which contained 30% of soluble insulin and 70% of isophane insulin, was purchased from Novo Nordisk (Bagsvaerd, Denmark). The Trizol solution and cDNA synthesis kit were bought from Thermo Fisher Scientific (Waltham, MA, USA). The polymerase chain reaction (PCR) primers were bought from Integrated DNA technologies (Coralville, IA, USA). The Taq polymerase buffer, deoxynucleotide triphosphates (dNTPs), nuclease-free water, DNA ladder, and MgCl2 were earned from (Promega, Madison, WI, USA). Agarose gel and gel electrophoresis buffer were bought from (Bio-basic, Markham, ON, Canada). Finally, the Redsafe dye was bought from (iNtRON Biotechnology, Seoul, South Korea).
Experimental animals
Twenty eight Balb/c mice (Mus musculus), weighing 24–28 g and of similar age were purchased from the animal house of Applied Science Private University (Amman, Jordan). The mice were cared in accordance with the guide of Canadian Council on Animal Care,Citation19 and the study protocol was approved by the ethical committee at Al-Zaytoonah University of Jordan (2017–2016/22/40). The mice were housed at 23°C ± 1°C, and 12 hours light–12 hours dark cycle. All animals were separately caged and maintained on standard laboratory animal diet pellets ad libitum.
Experimental protocol
Following a period of stabilization (7 days), the mice were divided into 4 groups of, 7 mice each, as follows:
Control group, which was treated with the vehicle 0.1 M citric acid buffer (0.1 citric acid monohydrate and 0.1 M trisodium citrate and adjusted to pH =4.5) by intraperitoneal route.
Uncontrolled DM (UDM) insulin-dependent group, which was treated with single intraperitoneal 240 mg/ kg streptozocin, solubilized in cold and freshly prepared citric buffer.
Controlled DM (CDM) insulin-dependent group, which was treated with single intraperitoneal 240 mg/kg streptozocin. Then after 3 days, the diabetic mice were treated with 1 unit/kg/day of Mixtard® insulin for 5 continuous days. The dose of insulin used in this study was equivalent to the dose used in human and it was used previously on the experimental mice.Citation20
Insulin-treated hypoglycemic group, which received no streptozocin but exposed to 1 unit/kg/day of Mixtard® insulin for 5 continuous days.
Physical observation
Daily observation throughout the study was made for mortality and general well-being in the diabetic and non-diabetic groups. The mice body weight was monitored at the beginning of treatment and after 8 days of treatment. The consumption of water and food was monitored by weighting the average amount of food (in g) and water (in mL) consumed per mouse, respectively.
Analysis of blood glucose level
The blood glucose level was measured using Accu-Chek Performa® (Roche, Basel, Switzerland). The blood drops obtained from the tail of the experimental mice. The blood glucose analyses were done before streptozocin treatment, after 3 days of streptozocin treatment and after 5 days of insulin treatment. The mouse was considered as diabetic when the fasting blood glucose was >200 mg/dL.Citation21
RNA extraction and cDNA synthesis
Around 100 mg of the liver samples were isolated from the mice. Then, total RNA was extracted and purified using Trizol RNA extraction solution (Thermo Fisher Scientific), isopropyl alcohol, and 75% ethanol, according to the manufacturer’s instructions. After that, RNA was converted to cDNA using cDNA Synthesis Kit® (Thermo Fisher Scientific). Briefly, 3 mg of total RNA was added to a reaction mixture containing 100 pmol oligo deoxythymine, 2.5 mM dNTP, 0.1 M DTT, 1X reverse transcriptase buffer, and 100 unit of Moloney murine leukemia virus reverse transcriptase and incubated at 42°C for 1 hour. The RNA and cDNA concentrations were measured using the Nanodrop instrument Quawell DNA/ Protein Analyzer (Quawell, Sunnyvale, CA, USA).
Gene expression analysis
Quantitative real-time PCR was performed using Bio-Rad CFX96 machine (Bio-Rad, Berkeley, CA, USA). Briefly, 1 ng of cDNA was added to a reaction mixture containing SYBR™ Green PCR Master Mix (Thermo Fisher Scientific) and selective primers () for cDNA sequence of drug- and arachidonic acid-metabolizing enzymes. The PCR condition used was as follows: denaturation at 95°C for 3 minutes followed by 40 cycles of denaturation at 95°C for 10 seconds and annealing at 53°C for 30 seconds. The gene expression of the targeted genes was normalized to the expression of the house keeping gene gapdh, as described previously,Citation22 using the following formula: ΔCT = CT (metabolizing gene)−CT (gapdh gene), where Ct is the PCR threshold cycle at which the fluorescence level rises above a baseline. Then, the relative gene expression was calculated as 2−ΔΔCT. The ΔΔCT is the difference of ΔCT between the treated and control group.
Table 1 Primers names, sequence, applicant size, and annealing temperature used in real-time PCR
Conventional PCR
The PCR was done as prescribed previously.Citation23,Citation24 Briefly, synthesized cDNA (330 ng) underwent 35 cycles of PCR amplification in a 20 µL reaction volume containing 10 mM dNTPs, 25 mM MgCl2, 10 pmol from each of the forward and reverse primers of cyp4a12, nat2, slc22a1, and gapdh genes (), and 1.5 unit of Go-Taq DNA polymerase.
Gel electrophoresis
The products of conventional reverse-transcriptase PCR were separated on 1.5% agarose gel after applying electrical current of 125 Ampere. The gel was stained using Redsafe dye. Comparison of the size was done between the PCR products and the standard size 100 bp DNA loading ladder. The DNA was seen by UV transilluminator (Bio Doc-ITTM, Cambridge, UK).
Statistical analysis
The results in this study were expressed by the average ± SD. ANOVA test was used as a statistical tool for comparison between the tested groups. Statistically significant changes compared with the control groups were indicated as *P<0.05.
Results
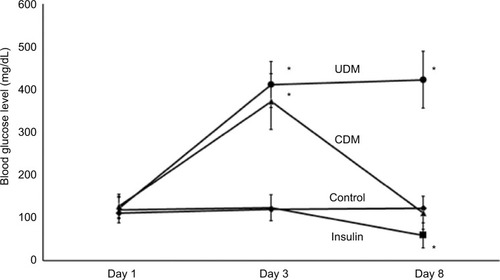
Streptozocin was used to induce diabetes in mice. After 3 days of 240 mg/kg streptozocin injection, the blood glucose levels of the streptozocin-treated animals were elevated significantly >400 mg/dL, while the blood glucose levels of the control group were within the normal levels <125 mg/ dL. shows that 5 days treatment of 1 unit/kg/day Mixtard® insulin normalized the blood glucose level (<125) in the CDM group. The last group was treated with only 1 unit/kg/day Mixtard® insulin, which exhibited hypoglycemic effect and reduced the blood glucose level below 59 mg/dL. This hypoglycemia was associated with the death of 40% of the treated mice and finally, this group contained 4 mice for gene expression analysis.
Figure 1 Blood glucose levels of the experimental animals.
Abbreviations: CDM, controlled diabetes mellitus; UDM, uncontrolled diabetes mellitus.
As shown in , the water consumption was increased >5-fold in the UDM group, which is a symptom of polydipsia among the diabetic mice. The polydipsia was less observed in the CDM group, where the water consumption was just 2-fold more than the control mice. The insulin-treated group showed higher water (3-fold) and food consumption (2-fold) than the control group ().
Table 2 Food and water consumption in the treated mice
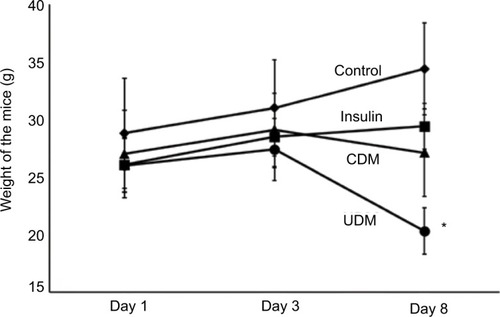
The weight of UDM group was reduced significantly (P-value <0.05) from 26±5 g at the first day of the experiment to 20.3±3 g at day 8 of this study (). The insulin treatment prevented the sharp decline in the body weight of the CDM, where the body weight was 26±3 g at day 8 of this experiment ().
Figure 2 The changes in body weight of the experimental mice.
Abbreviations: CDM, controlled diabetes mellitus; UDM, uncontrolled diabetes mellitus.
As diabetic animals showed polydipsia and reduction in body weight, these data indicated that the diabetic model induced in this study was insulin-dependent type I diabetes.Citation25
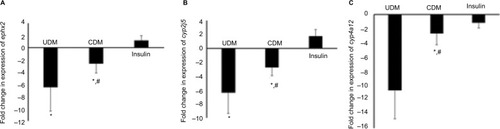
Regarding gene expression, the mRNA levels of arachidonic acid- and drug-metabolizing-cyp450s were significantly reduced in livers of diabetic mice (–). The most affected genes among arachidonic and drug-metabolizing cyp450 genes were cyp1a2 and cyp4a12 with down-regulation in gene expression by 16.9- and 10.7-fold, respectively. The gene expression of cyp2e1 and cyp2c29 was reduced by only 4.3- and 4.2-fold, respectively, which showed the lowest influence by DM compared with other cyp450s (). However, 5 days of insulin treatment attenuated significantly (P-value <0.05) the effect of DM on cyp450 gene expression (). The livers of hypoglycemic insulin-treated mice did not show significant difference compared with the control group, except that cyp3a11 was induced significantly (P-value <0.05) to >2-fold ().
Figure 3 Gene expression of Phase I drug-metabolizing (A–E) enzymes in the liver of the tested mice.
Abbreviations: CDM, controlled diabetes mellitus; PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
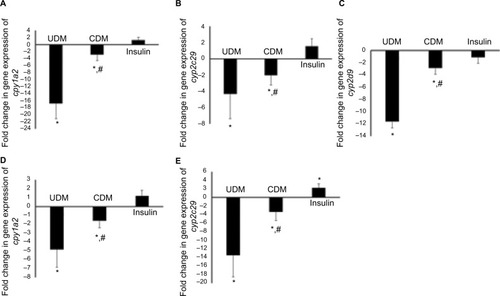
Figure 4 Gene expression of Phase II drug-metabolizing enzymes (A–E) in the liver of the tested mice.
Notes: Total RNA was isolated from the livers then converted to cDNA. The gene expression was analyzed using real-time PCR with SYPER green die. Further details are provided in the Methods section. “*” refers to significant difference (P-value<0.05) compared with the control group, while “#” means significant difference (P-value<0.05) compared with the UDM.
Abbreviations: CDM, controlled diabetes mellitus; PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
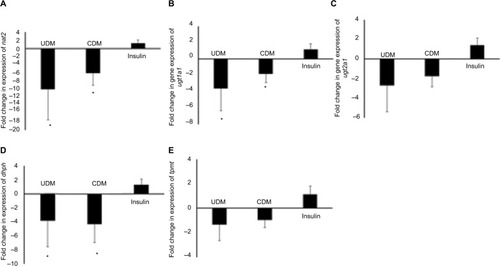
Figure 5 Gene expression of arachidonic acid-cyp450s (A–C) in the liver of the tested mice.
Abbreviations: CDM, controlled diabetes mellitus; PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
Phase II drug-metabolizing enzymes were also reduced in the livers of the UDM mice (). Although, Phase II drug-metabolizing enzymes were less influenced by DM than Phase I enzymes, the most influenced gene among Phase II metabolizing enzymes were nat2, where it was reduced by >10-fold in UDM (). The gene expression of ugt1a1, 2b1, and dhpd were reduced by 3.6-, 2.7-, and 3.8-fold, respectively (). In addition, insulin treatment failed to attenuate significantly (P-value >0.05) the effect of DM on nat2, ugt2b5, and dhpd genes (). The mRNA levels of tpmt gene did not differ significantly in all treated groups ().
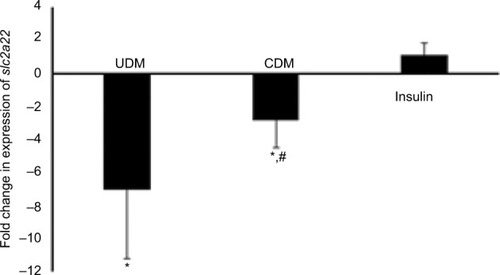
Furthermore, the gene expression of drug transporter gene slc22a1 was reduced significantly to 7-fold in UDM but attenuated significantly by insulin treatment ().
Figure 6 Gene expression of drug transporter gene slc22a1 in the liver of the tested mice.
Abbreviations: CDM, controlled diabetes mellitus; PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
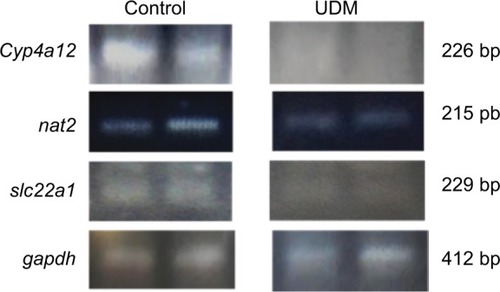
In addition, we confirmed the reduction in the gene expression of cyp4a12, nat2, and slc22a1 genes, which were highly influenced drug- and arachidonic acid-metabolizing enzyme genes, using reverse transcriptase PCR. As it is shown on , the intensity of the PCR bands of cyp4a12, nat2, and slc22a1 genes, obtained from the livers of control mice, was higher than of the PCR bands obtained from the UDM group.
Figure 7 Reverse transcriptase PCR for the major downregulated genes in the livers of diabetic mice.
Notes: The RNA was extracted from the livers of UDM and non-diabetic control group. Then, the RNA was converted to cDNA. The cyp4a12, nat2, and slc22a1 genes were amplified using conventional PCR. The gapdh gene was amplified as housekeeping gene to normalize the expression of the tested genes. Then, the PCR products were run on 1.5% agarose gel. The PCR bands were visualized after staining of the gel with Redsafe dye. Further details are in the Materials and Methods section.
Abbreviations: PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
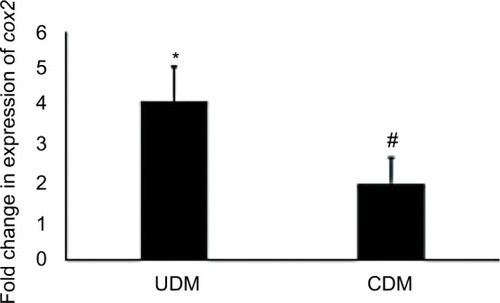
Finally, the current study analyzed the gene expression of the inflammation marker gene cyclo-oxygenase (cox2) and found its expression was increased >4-fold in UDM and attenuated to 1.8-fold after insulin therapy ().
Figure 8 Gene expression analysis of cox2 gene in the livers of diabetic mice.
Abbreviations: CDM, controlled diabetes mellitus; PCR, polymerase chain reaction; UDM, uncontrolled diabetes mellitus.
Discussion
Clinical studies showed that insulin-dependent diabetic patients have reduced capacity to metabolize drugs.Citation10,Citation12 The preset study investigated the alterations in gene expression of 14 genes involved in drug and arachidonic acid metabolism in the livers of insulin-dependent diabetic mice. We found that diabetic mice have significant decline in the mRNA levels of all tested drug- and arachidonic acid-metabolizing enzymes. Moreover, normalizing the blood glucose levels, using insulin, reversed significantly the decline in gene expression. It was also observed that hepatic cyp3a11 was induced in hypoglycemic mice. These alterations may explain, at least in part, the variation in drug metabolism and hence its response among insulin-dependent diabetic patients. The findings of this study may increase our understanding of the molecular changes in the hepatic metabolism of drugs and fatty acids among diabetic patients.
The present study used streptozocin as a chemical compound to induce diabetes in the mice. Streptozocin destroys the beta-pancreatic cells, which makes them unable to produce insulin and results into elevated blood glucose level.Citation26 The streptozocin-induced diabetes is the most common animal model used to study the morphological, biochemical, and molecular alterations induced by DM.Citation27 However, streptozocin itself can influence harmfully the liver cells.Citation28 The alterations in gene expression of drug and arachidonic and metabolizing enzymes, found in this study, might be due to the hyperglycemia rather than the direct effect of streptozocin on the liver; where the reduction in gene expression was attenuated by normalizing the blood glucose levels.
Diabetes is reported to cause hepatic impairment. The gene expression of drug- and arachidonic acid-metabolizing enzymes were altered in hepatic dysfunction and drug-induced hepatotoxicity. Some studies showed that the gene expression and the capacity of drug-metabolizing enzymes decreased in patients with hepatic dysfunctions.Citation29,Citation30 In addition, it is reported that the gene expression of drug-metabolizing enzymes was reduced in hepatic injury and diseases, such as cirrhosis and sepsis.Citation31,Citation32 The antituberculosis rifampin induced hepatotoxicity in mouse livers, which was coupled with down-regulation of arachidonic acid-cyp450s.Citation33 Furthermore, the inflammatory mediator interleukin, which is released during hepatic injury,Citation31 decreased the gene expression of Phase I and II drug-metabolizing enzymes and drug transporters.Citation34 It is reported that inflammatory mediators were elevated in the livers of diabetic patients,Citation35 and we showed in this study that gene expression of inflammatory biomarker cox2 was increased significantly (P-value <0.05) in the livers of UDM. These might indicate that the inflammatory mediators play a role in the down-regulation of drug- and arachidonic acid-metabolizing enzymes in the livers of diabetic mice.
The mouse cyp3a11, cyp2c29, and cyp2d9 are a homology of human CYP3A4, CYP2C9, and CYP2D6, respectively. These CYP450s are the major Phase I drug-metabolizing enzymes, which oxidize >80% of the drugs.Citation36 It is pointed out that CYP3A4 substrate antipyrine metabolism was lower in diabetic patients than normal volunteers.Citation37 In addition, midazolam and testosterone hydrolyzation were reduced in the human liver microsomes isolated from diabetic livers, which were associated with reduced CYP3A4 protein and mRNA levels.Citation11,Citation12 Furthermore, Kudo et al, found that intestinal cyp3a and hepatic cyp2c29 and cyp2d9 mRNA levels were reduced in diabetic mice.Citation38 The results of this study are in line with the previous studies where gene expression of drug-metabolizing cyp450s was significantly lower in UDM than control mice. This down-regulation of drug-metabolizing cyp450s may explain, at least in part, the reduction of oxidation capacity toward drugs among insulin-dependent diabetic patients. Further studies are needed to correlate between the gene expression levels of cyp450s and drug oxidation.
Arachidonic acid-metabolizing cyp450s convert arachidonic acid to cardioactive compounds. Cyp4a12 synthesize the platelet activator and vasoconstrictor 20-hydroxyeicosatetraenoic acid (20-HETE), while cyp2c29 and cyp2j5 metabolize arachidonic acid to the vasodilator epoxyeicosatrienoic acids (EETs).Citation39 All of the arachidonic acid-metabolizing cyp450 genes were reduced significantly in this study. However, 20-HETE-synthesizing genes were downregulated (>10-fold) more than EETs producing genes. It is showed that alterations of gene expression of arachidonic acid-metabolizing cyp450s, influenced on 20-HETE and EETs metabolite levels.Citation40 This may indicate that the ratio of 20-HETE to EETs synthesis was reduced in the livers of diabetic mice. The role of 20-HETE and EETs in the pathophysiology of the liver is still not understood. Although, it is suggested that down-regulation of cyp4a12 decreases the ability to metabolize fatty acid in the liver,Citation15 this reduction in fatty acid metabolism may deposit fatty acids in the liver. The fatty liver was reported in diabetic patients.Citation8 Therefore, the observed reduction in arachidonic acid-cyp450 gene expression, in this study, may contribute to the pathogenesis of the liver tissues among insulin-dependent diabetes.
UGTs also play a major role in endogenous and drug metabolism.Citation39 They metabolize 10% of all prescribed drugs, such as morphine.Citation41 In addition, they are reported to metabolize arachidonic acids.Citation39 It is found clinically that diabetic patients have lower capacity to form acyl glucuronide mycophenolic acid, which was associated with lower human UGT2B7 protein expression in the liver and kidney.Citation12 The present study found that mouse ugt2b1, which is a homology of human UGT2B7, was downregulated by 3.4-fold. In addition, we found for the first time that hepatic ugt1a1 gene, which encodes ugt1a1 enzyme and metabolizes the anticancer irinotecan,Citation42 was also downregulated among UDM. Therefore, it is speculated that the rate of activation of irinotecan is low among insulin-dependent diabetic patient on irinotecan therapy. Although, insulin therapy did not attenuate significantly the down-regulation of ugt genes, this might indicate that molecular regulation of ugt genes is different than cyp450s in the diabetic livers.
This study observed for the first time that gene expression of nat2 gene, which acetylates the drugs, was the most affected Phase II drug-metabolizing enzymes in the liver samples of diabetic mice. This finding is in line with the study of Irshaid et al, where the antimicrobial dapsone acetylation capacity was reduced among insulin-dependent patients.Citation10 Therefore, down-regulation of the hepatic nat2 gene may explain the reduced capacity of drug acetylation among diabetic human patients.
The organic cation transporter 1 (OCT1) mediates the transportation of some drugs, such as the antidiabetic metformin, to the hepatic tissue.Citation43 It is found in this study that OCT1 encoding gene slc22a1 was reduced in UDM but reversed, by insulin therapy, in CDM. It is reported that oct1 expression was reduced in streptozocin-induced diabetic rats and influenced the kinetics of tetraethylammonium.Citation44 Therefore, it is speculated that the pharmacokinetics and hence the response of oct1 substrates, such as metformin, is different between normal and diabetic patients. Further intensive studies are needed to find the correlation between hepatic slc22a1 expression and metformin response in diabetic patients.
It is shown that insulin treatment induced several gene expression in different organs, including the liver.Citation45 In this study, we showed that insulin treatment reversed significantly the decline in mRNA of arachidonic and drug-metabolizing enzymes. However, 5 days treatment of insulin therapy of diabetic mice did not show comparable mRNA levels of arachidonic and drug-metabolizing enzymes to the control group. This might be due to the short duration of insulin therapy (5 days) used in the experiment design of this study. However, the decline in the drug metabolism capacity was also observed among controlled diabetic human volunteers.Citation10 Moreover, it is found in this study that insulin induced significantly cyp3a12 mRNA levels in the livers of hypoglycemic mice. This finding confirmed what was found in Davidson et al study, that human CYP3A4 was induced in the primary human hepatocytes after insulin treatment.Citation46 This may indicate that hypoglycemic patients have higher capacity to metabolize CYP3A4 substrate drugs, which influence the pharmacokinetics and hence drug response. Further clinical studies are needed to find the metabolism rate of CYP3A4 substrates among hypoglycemic patients.
Conclusion
The present study investigated the alterations in gene expression of arachidonic and drug-metabolizing enzymes in the livers of insulin-controlled and non-controlled diabetic and hypoglycemic mice and found that hyperglycemia decreased sharply while insulin attenuated this down-regulation in arachidonic and drug-metabolizing enzymes in diabetic mice. In addition, insulin induced cyp3a11 in the hypoglycemic group. These molecular alterations may help us in understanding the variation in drug metabolism and drug response between controlled and non-controlled diabetic patients compared with healthy people.
Acknowledgments
This work was supported by Deanship of Scientific Research at Al-Zaytoonah University of Jordan with a grant fund number of 13-10-2016.
Disclosure
The authors report no conflicts of interest in this work
References
- Corsino L Dhatariya K Umpierrez G Management of Diabetes and Hyperglycemia in Hospitalized Patients Endotext De Groot LJ Chrousos G Dungan K MA South Dartmouth 2000
- Ajlouni K Khader YS Batieha A Ajlouni H El-Khateeb M An increase in prevalence of diabetes mellitus in Jordan over 10 years J Diabetes Complications 2008 22 5 317 324 18413210
- Albache N Al Ali R Rastam S Fouad FM Mzayek F Maziak W Epidemiology of Type 2 diabetes mellitus in Aleppo, Syria J Diabetes 2010 2 2 85 91 20923489
- Engelgau MM Geiss LS Saaddine JB The evolving diabetes burden in the United States Ann Intern Med 2004 140 11 945 950 15172919
- Obrosova IG Fathallah L Liu E Nourooz-Zadeh J Early oxidative stress in the diabetic kidney: effect of DL-alpha-lipoic acid Free Radic Biol Med 2003 34 2 186 195 12521600
- Yue KK Chung WS Leung AW Cheng CH Redox changes precede the occurrence of oxidative stress in eyes and aorta, but not in kidneys of diabetic rats Life Sci 2003 73 20 2557 2570 12967680
- American Diabetes Association Diagnosis and classification of diabetes mellitus Diabetes Care 2010 33 Suppl 1 S62 S69 20042775
- Bhatt HB Smith RJ Fatty liver disease in diabetes mellitus Hepatobiliary Surg Nutr 2015 4 2 101 108 26005676
- Mantovani A Targher G Type 2 diabetes mellitus and risk of hepatocellular carcinoma: spotlight on nonalcoholic fatty liver disease Ann Transl Med 2017 5 13 270 28758096
- Irshaid Y Al-Hadidi H Abuirjeie M Acetylator phenotypes of Jordanian diabetics Eur J Clin Pharmacol 1992 43 6 621 623 1493843
- Dostalek M Court MH Yan B Akhlaghi F Significantly reduced cytochrome P450 3A4 expression and activity in liver from humans with diabetes mellitus Br J Pharmacol 2011 163 5 937 947 21323901
- Dostalek M Court MH Hazarika S Akhlaghi F Diabetes mellitus reduces activity of human UDP-glucuronosyltransferase 2B7 in liver and kidney leading to decreased formation of mycophenolic acid acylglucuronide metabolite Drug Metab Dispos 2011 39 3 448 455 21123165
- Lee JH Yang SH Oh JM Lee MG Pharmacokinetics of drugs in rats with diabetes mellitus induced by alloxan or streptozocin: comparison with those in patients with type I diabetes mellitus J Pharm Pharmacol 2010 62 1 1 23 20722995
- Elshenawy OH Shoieb SM Mohamed A El-Kadi AO Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain: An Emerging Therapeutic Target Pharmaceutics 2017 9 1 9
- Leclercq IA Farrell GC Field J CYP2E1 and CYP4A as microsomal catalysts of lipid peroxides in murine nonalcoholic steatohepatitis J Clin Invest 2000 105 8 1067 1075 10772651
- Lukaszewicz KM Lombard JH Role of the CYP4A/20-HETE pathway in vascular dysfunction of the Dahl salt-sensitive rat Clin Sci 2013 124 12 695 700 23438293
- Pass GJ Becker W Kluge R Effect of hyperinsulinemia and type 2 diabetes-like hyperglycemia on expression of hepatic cytochrome p450 and glutathione s-transferase isoforms in a New Zealand obese-derived mouse backcross population J Pharmacol Exp Ther 2002 302 2 442 450 12130701
- Ahmed S Zhou Z Zhou J Chen SQ Pharmacogenomics of Drug Metabolizing Enzymes and Transporters: Relevance to Precision Medicine Genomics Proteomics Bioinformatics 2016 14 5 298 313 27729266
- Canadian Council on Animal Care Guide to the care and use of experimental animals Canadian Council on Animal Care Ottawa, Ont 1980
- Guo T Mao Y Li H Characterization of the gene expression profile of heterozygous liver-specific glucokinase knockout mice at a young age Biomed Pharmacother 2012 66 8 587 596 23085254
- Oliveira WH Nunes AK França ME Effects of metformin on inflammation and short-term memory in streptozotocin-induced diabetic mice Brain Res 2016 1644 149 160 27174003
- Livak KJ Schmittgen TD Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method Methods 2001 25 4 402 408 11846609
- Jarrar YB Shin JG Lee SJ Expression of arachidonic acid-metabolizing cytochrome P450s in human megakaryocytic Dami cells In Vitro Cell Dev Biol Anim 2013 49 7 492 500 23722412
- Jarrar YB Balasmeh AA Jarrar W Sequence analysis of the N-acetyl-transferase 2 gene (NAT2) among Jordanian volunteers Libyan J Med 2018 13 1 1408381 29173142
- King AJ The use of animal models in diabetes research Br J Pharmacol 2012 166 3 877 894 22352879
- Cheng Y Kang H Shen J Beta-cell regeneration from vimentin+/ MafB+ cells after STZ-induced extreme beta-cell ablation Sci Rep 2015 5 11703 26129776
- Damasceno DC Netto AO Iessi IL Streptozotocin-induced diabetes models: pathophysiological mechanisms and fetal outcomes Biomed Res Int 2014 2014 819065 24977161
- Kume E Fujimura H Matsuki N Hepatic changes in the acute phase of streptozotocin (SZ)-induced diabetes in mice Exp Toxicol Pathol 2004 55 6 467 480 15384252
- Cobbina E Akhlaghi F Non-alcoholic fatty liver disease (NAFLD) -pathogenesis, classification, and effect on drug metabolizing enzymes and transporters Drug Metab Rev 2017 49 2 197 211 28303724
- Palatini P de Martin S Pharmacokinetic drug interactions in liver disease: An update World J Gastroenterol 2016 22 3 1260 1278 26811663
- Elbekai RH Korashy HM El-Kadi AO The effect of liver cirrhosis on the regulation and expression of drug metabolizing enzymes Curr Drug Metab 2004 5 2 157 167 15078193
- Higuchi S Wu R Zhou M Ravikumar TS Wang P Downregulation of hepatic cytochrome P-450 isoforms and PPAR-gamma: their role in hepatic injury and proinflammatory responses in a double-hit model of hemorrhage and sepsis J Surg Res 2007 137 1 46 52 17101152
- Kim JH Nam WS Kim SJ Mechanism Investigation of Rifam-picin-Induced Liver Injury Using Comparative Toxicoproteomics in Mice Int J Mol Sci 2017 18 7 E1417 28671602
- Klein M Thomas M Hofmann U A systematic comparison of the impact of inflammatory signaling on absorption, distribution, metabolism, and excretion gene expression and activity in primary human hepatocytes and HepaRG cells Drug Metab Dispos 2015 43 2 273 283 25480923
- Loria P Lonardo A Anania F Liver and diabetes. A vicious circle Hepatol Res 2013 43 1 51 64 23332087
- Almazroo OA Miah MK Venkataramanan R Drug Metabolism in the Liver Clin Liver Dis 2017 21 1 1 20 27842765
- Oltmanns D Dennin DE Pentz R Siegers CP Antipyrine clearance as a measure of drug metabolism in patients with diabetes mellitus Z Gastroenterol 1984 22 10 598 601 6506825
- Kudo T Toda T Ushiki T Differences in the pharmacokinetics of Cyp3a substrates in TSOD and streptozotocin-induced diabetic mice Xenobiotica 2010 40 4 282 290 20163193
- Jarrar YB Cha EY Seo KA Determination of major UDP-glucuronosyltransferase enzymes and their genotypes responsible for 20-HETE glucuronidation J Lipid Res 2014 55 11 2334 2342 25249502
- Alsaad AM Zordoky BN El-Sherbeni AA El-Kadi AO Chronic doxorubicin cardiotoxicity modulates cardiac cytochrome P450-mediated arachidonic acid metabolism in rats Drug Metab Dispos 2012 40 11 2126 2135 22867862
- de Gregori S de Gregori M Ranzani GN Morphine metabolism, transport and brain disposition Metab Brain Dis 2012 27 1 1 5 22193538
- Dean L Irinotecan Therapy and UGT1A1 Genotype Medical Genetics Summaries Pratt V McLeod H Dean L Malheiro A Rubinstein W MD Bethesda 2012
- Hakooz N Jarrar YB Zihlif M Effects of the genetic variants of organic cation transporters 1 and 3 on the pharmacokinetics of metfor-min in Jordanians Drug Metab Pers Ther 2017 32 3 157 162 28862982
- Grover B Buckley D Buckley AR Cacini W Reduced expression of organic cation transporters rOCT1 and rOCT2 in experimental diabetes J Pharmacol Exp Ther 2004 308 3 949 956 14718608
- Mastaitis JW Wurmbach E Cheng H Sealfon SC Mobbs CV Acute induction of gene expression in brain and liver by insulin-induced hypoglycemia Diabetes 2005 54 4 952 958 15793232
- Davidson MD Ballinger KR Khetani SR Long-term exposure to abnormal glucose levels alters drug metabolism pathways and insulin sensitivity in primary human hepatocytes Sci Rep 2016 6 28178 27312339